
Importance of Local Studies of Vascular Plant Communities in Conservation and Management: A Case Study in Susticacán, Zacatecas, Mexico

 ,
, 
Abstract
:
1. Introduction
2. Materials and Methods
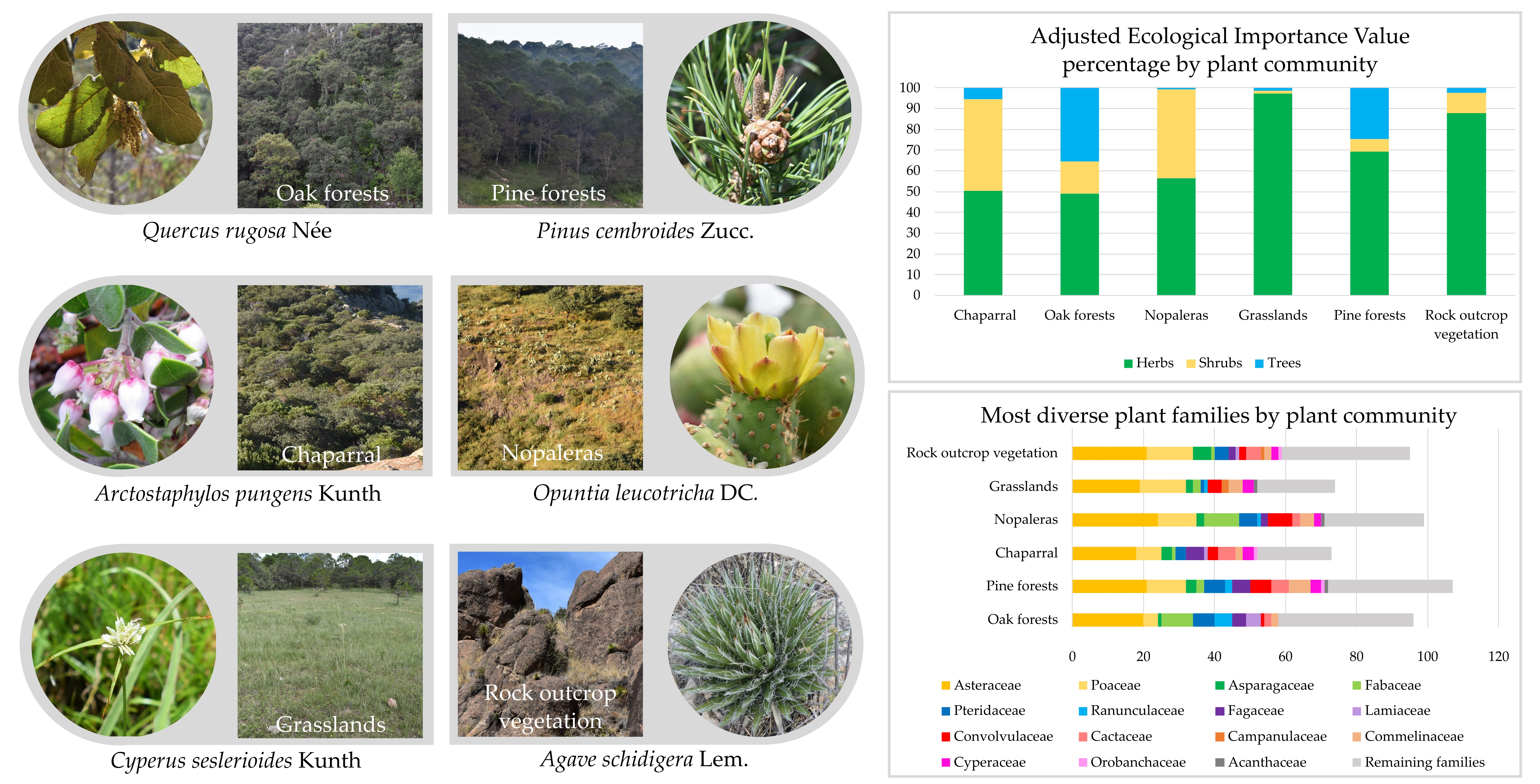
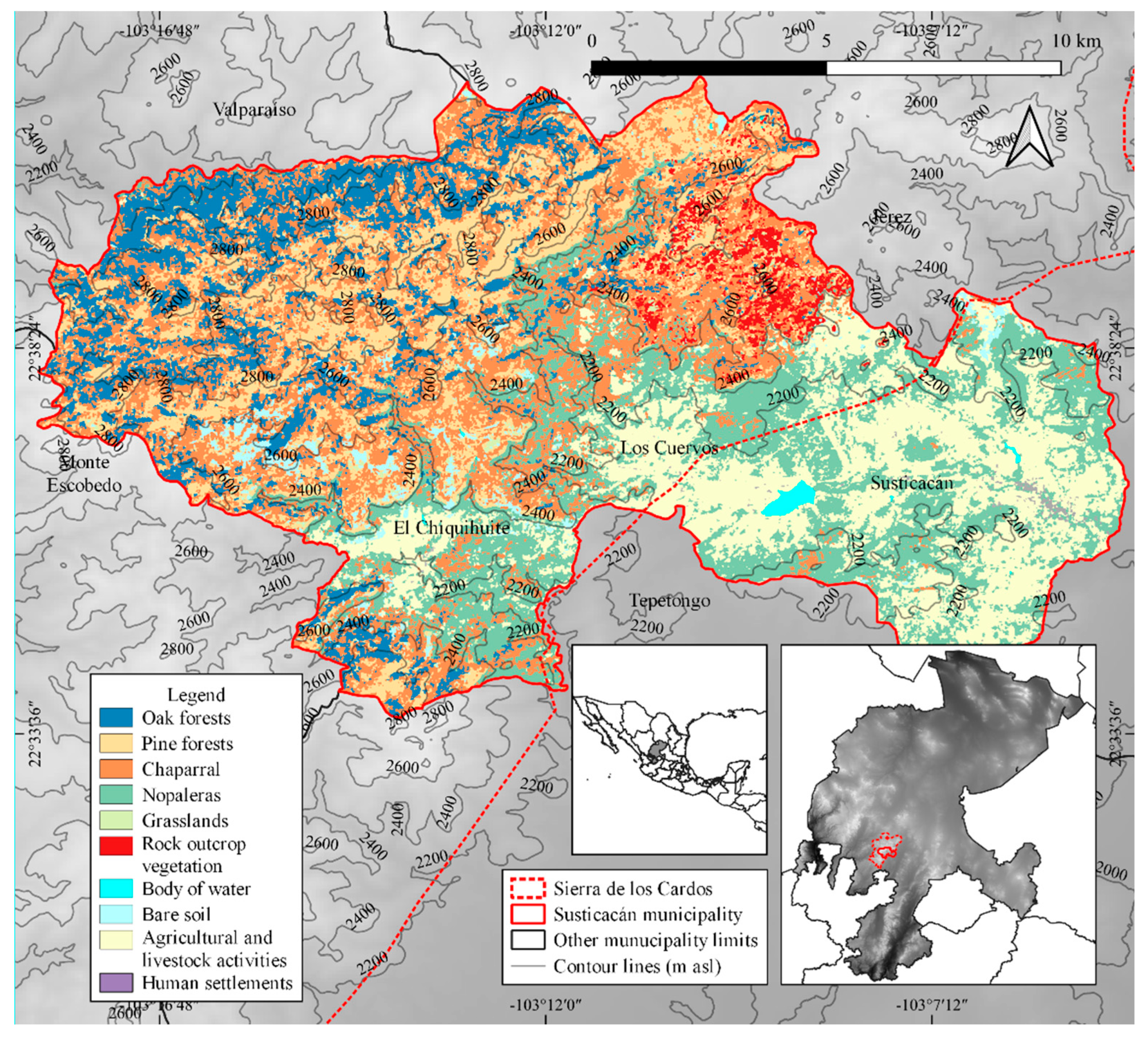
2.1. Study Site
2.2. Identification of Vascular Plant Communities
2.3. Characterization and Distribution of Vascular Plant Communities
3. Results
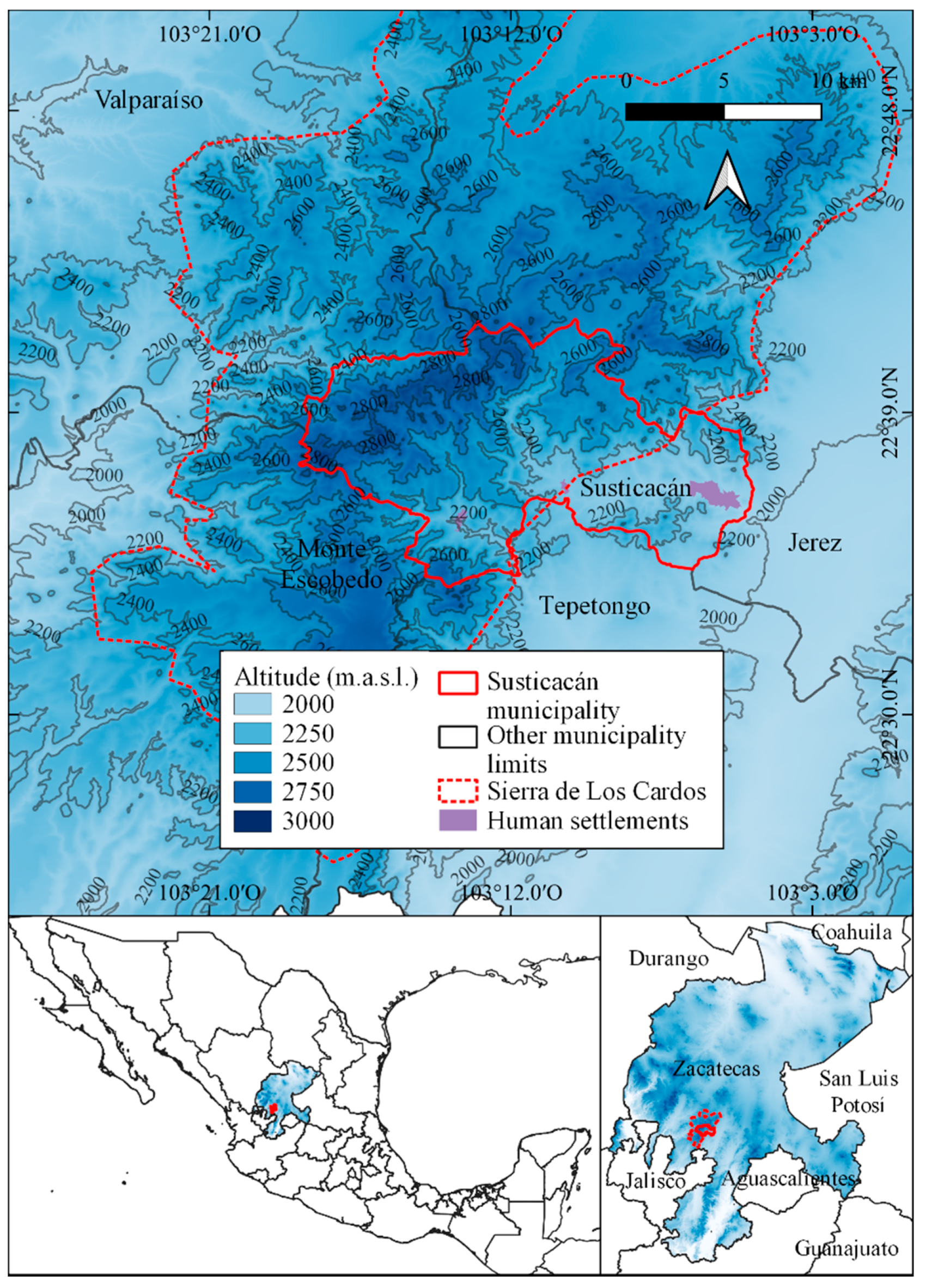
3.1. Identification of Vascular Plant Communities
3.2. Characterization and Distribution of Vascular Plant Communities
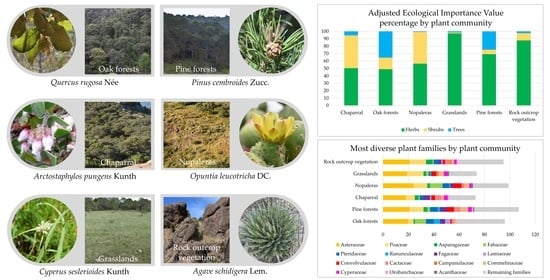
3.2.1. Oak Forests
3.2.2. Pine Forests
3.2.3. Chaparral
3.2.4. Nopaleras
3.2.5. Grasslands
3.2.6. Rock Outcrops Vegetation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chytrý, M.; Chiarucci, A.; Pärtel, M.; Pillar, V.D.; Bakker, J.P.; Mucina, L.; Peet, R.K.; White, P.S. Progress in vegetation science: Trends over the past three decades and new horizons. J. Veg. Sci. 2019, 30, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Pärtel, M.; Chiarucci, A.; Chytrý, M.; Pillar, V.D. Mapping plant community ecology. J. Veg. Sci. 2017, 28, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Damschen, E.I. Decoding plant communities across scales. Ecol. Evol. 2018, 2, 1844–1845. [Google Scholar] [CrossRef]
- Czaran, T.; Bartha, S. Spatiotemporal dynamic models of plant populations and communities. Trends Ecol. Evol. 1992, 7, 38–42. [Google Scholar] [CrossRef]
- Mori, A.S. Ecosystem management based on natural disturbances: Hierarchical context and non-equilibrium paradigm. J. Appl. Ecol. 2011, 48, 280–292. [Google Scholar] [CrossRef]
- Franklin, S.B.; Hunter, J.T.; De Cáceres, M.; Dengler, J.; Landucci, F.; Krestov, P. Introducing the IAVS Vegetation Classification Working Group. Phytocoenologia 2016, 46, 5–8. [Google Scholar] [CrossRef]
- Jensen, F.; Bergmeier, E.; Dengler, J.; Janišová, M.; Krestov, P.; Willner, W. Vegetation classification: A task for our time. Phytocoenologia 2016, 46, 1–4. [Google Scholar] [CrossRef]
- European Space Agency. Vegetation Classification Panel. Available online: https://www.esa.org/vegpanel/usnvc/ (accessed on 13 January 2021).
- Goodall, W.D. Numerical methods of classification. In Classification of Plant Communities; Whittaker, R.H., Ed.; Dr. W. Junk Publishers: Hague, The Netherlands, 1978; pp. 247–286. [Google Scholar]
- Whittaker, R.H. Communities and Ecosystems; Macmillan: New York, NY, USA, 1970; pp. 60–105. ISBN 978-002-4273-90-1. [Google Scholar]
- Rzedowski, J. Vegetación de México; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: Ciudad de México, Mexico, 2006; pp. 12–168. [Google Scholar]
- Sosa, V.; Loera, I. Influence of current climate, historical climate stability and topography on species richness and endemism in Mesoamerican geophyte plants. PeerJ 2017, 5, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Global Forest Watch. Mexico. Available online: https://www.globalforestwatch.org/dashboards/country/MEX/ (accessed on 4 December 2020).
- Grabherr, G.; Gottfried, M.; Pauli, H. Climate effects on mountain plants. Nature 1994, 369, 448. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.M.; Diez, J.M.; Levine, J.M. Novel competitors shape species’ responses to climate change. Nature 2015, 525, 515–518. [Google Scholar] [CrossRef]
- Steinmann, V.W.; Arredondo-Amezcua, L.; Hernández-Cárdenas, R.A.; Ramírez-Amezcua, Y. Diversity and origin of the central mexican alpine flora. Diversity 2021, 13, 31. [Google Scholar] [CrossRef]
- Descroix, L.; González Barrios, J.L.; Estrada Ávalos, J. La Sierra Madre Occidental, una Fuente de Agua Amenazada; Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias: Gómez Palacio, Mexico, 2004; pp. 25–85. [Google Scholar]
- González-Elizondo, M.S.; González-Elizondo, M.; Ruacho González, L.; Lopez Enriquez, L.L.; Retana Rentería, F.I.; Tena Flores, J.A. Ecosystems and Diversity of the Sierra Madre Occidental. In Merging Science and Management in a Rapidly Changing World: Biodiversity and Management of the Madrean Archipelago, 3rd ed.; Gottfried, G.J., Ffolliott, P.F., Gebow, B.S., Eskew, L.G., Collins, L.C., Eds.; U.S. Department of Agriculture: Fort Collins, CO, USA, 2013; pp. 204–211. [Google Scholar]
- Instituto Nacional de Estadística y Geografía. Prontuario de información Geográfica Municipal de los Estados Unidos Mexicanos, Zacatecas, Jerez, 32020. Available online: http://www3.inegi.org.mx/contenidos/app/mexicocifras/datos_geograficos/32/32020.pdf (accessed on 21 May 2021).
- Instituto Nacional de Estadística y Geografía. Prontuario de Información Geográfica Municipal de los Estados Unidos Mexicanos, Zacatecas, Monte Escobedo, 32031. Available online: http://www3.inegi.org.mx/contenidos/app/mexicocifras/datos_geograficos/32/32031.pdf (accessed on 21 May 2021).
- Instituto Nacional de Estadística y Geografía. Prontuario de Información Geográfica Municipal de los Estados Unidos Mexicanos, Zacatecas, Susticacán, 32043. Available online: http://www3.inegi.org.mx/contenidos/app/mexicocifras/datos_geograficos/32/32043.pdf (accessed on 21 May 2021).
- Instituto Nacional de Estadística y Geografía. Prontuario de Información Geográfica Municipal de los Estados Unidos Mexicanos, Zacatecas, Tepetongo, 32046. Available online: http://www3.inegi.org.mx/contenidos/app/mexicocifras/datos_geograficos/32/32046.pdf (accessed on 21 May 2021).
- Instituto Nacional de Estadística y Geografía. Prontuario de Información Geográfica Municipal de los Estados Unidos Mexicanos, Zacatecas, Valparaíso, 32049. Available online: http://www3.inegi.org.mx/contenidos/app/mexicocifras/datos_geograficos/32/32049.pdf (accessed on 21 May 2021).
- Fernández-Borja, H.; Giménez-de Azcárate, J. Ruta Histórico Cultural del Pueblo Huichol. Available online: https://cartocritica.org.mx/2014/ruta-a-wirikuta/ (accessed on 17 May 2021).
- Giménez-de Azcárate, J.; Escamilla, M. Las comunidades edafoxerófilas (enebrales y zacatonales) en las montañas del centro de México. Phytocoenologia 1999, 29, 449–468. [Google Scholar]
- Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. Índice de Impacto Humano en la Biodiversidad Terrestre. Available online: http://geoportal.conabio.gob.mx/metadatos/doc/html/indihum14gw.html (accessed on 4 April 2021).
- Parmesan, C. Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef] [Green Version]
- Wilson, R.J.; Gutierrez, D.; Gutierrez, J.; Monserrat, V.J. An elevational shift in butterfly species richness and composition accompanying recent climate change. Glob. Change Biol. 2007, 13, 1873–1887. [Google Scholar] [CrossRef]
- Lenoir, J.; Gégout, J.C.; Marquet, P.A.; De Ruffray, P.; Brisse, H. A significant upward shift in plant species optimum elevation during the 20th century. Science 2008, 320, 1768–1771. [Google Scholar] [CrossRef] [PubMed]
- Parolo, G.; Rossi, G. Upward migration of vascular plants following a climate warming trend in the Alps. Basic Appl. Ecol. 2008, 9, 100–107. [Google Scholar] [CrossRef]
- Jump, A.S.; Huang, T.J.; Chou, C.H. Rapid altitudinal migration of mountain plants in Taiwan and its implications for high altitude biodiversity. Ecography 2012, 35, 204–210. [Google Scholar] [CrossRef]
- Instituto Nacional de Estadística y Geografía. Conjunto de Datos Vectoriales Fisiográficos, Continuo Nacional Serie I, Sistema Topoformas. Available online: https://datos.gob.mx/busca/dataset/conjunto-de-datos-vectoriales-fisiograficos-continuo-nacional-escala-1-1-000-000-serie-i/resource/d82b7280-5680-4968-95ec-6755590ae0a2. (accessed on 21 May 2021).
- Instituto Nacional de Estadística y Geografía. Condensado Estatal Zacatecas. Available online: https://portalanterior.ine.mx/archivos1/Cartografia/2014/CED/32_ZAC/CED32_CARTA_110614.pdf (accessed on 21 May 2021).
- García, E. Modificaciones al Sistema de Clasificación Climatológica de Köppen; Instituto de Geografía: Ciudad de México, Mexico, 1981; pp. 3–34. [Google Scholar]
- United States Geological Survey. Earth Explorer. Available online: https://earthexplorer.usgs.gov/ (accessed on 10 November 2020).
- Hurtado-Reveles, L.; Burgos-Hernández, M.; Vázquez-Sánchez, M.; López-Acosta, J.C. Contribución al conocimiento florístico de la Sierra de los Cardos, Zacatecas, México. Bot Sci. in press. 1–45. [CrossRef]
- Van der Maarel, E. Transformation of cover-abundance values in phytosociology and its effects on community similarity. Vegetatio 1979, 39, 97–114. [Google Scholar]
- Kruskal, J.B. Multidimensional scaling by optimizing goodness of fit to a nonmetric hypothesis. Psychometrika 1964, 29, 1–27. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Similarity-based testing for community pattern: The two-way layout with no replication. Mar. Biol. 1994, 118, 167–176. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. A further biodiversity index applicable to species lists: Variation in taxonomic distinctness. Mar. Ecol-Prog Ser. 2001, 216, 265–278. [Google Scholar] [CrossRef]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Miranda, F.; Hernández-Xolocotzi, E. Los tipos de vegetación de México y su clasificación. Bot. Sci. 1963, 28, 29–179. [Google Scholar] [CrossRef]
- Guzmán, G.; Vela-Gálvez, L. Contribución al conocimiento de la vegetación del suroeste del estado de Zacatecas (República Mexicana). Bot. Sci. 1960, 25, 46–61. [Google Scholar] [CrossRef] [Green Version]
- Enríquez-Enríquez, D.E.; Koch, S.D.; González-Elizondo, M.S. Flora y vegetación de la Sierra de Órganos, municipio de Sombrerete, Zacatecas, México. Acta Bot. Mex. 2003, 64, 45–89. [Google Scholar] [CrossRef] [Green Version]
- González-Elizondo, M.S.; González-Elizondo, M.; Tena-Flores, J.A.; Ruacho-González, L.; López-Enríquez, I.L. Vegetación de la Sierra Madre Occidental, México: Una síntesis. Acta Bot. Mex. 2012, 100, 351–403. [Google Scholar] [CrossRef] [Green Version]
- Comisión Técnico Consultiva para la Determinación Regional de los Coeficientes de Agostadero. Coeficientes de Agostadero de la República Mexicana, estados Aguascalientes y Zacatecas; Secretaría de Agricultura y Recursos Hidráulicos: Ciudad de México, Mexico, 1980; pp. 22–30. [Google Scholar]
- Herrera-Arrieta, Y.; Pámanes-García, D.S. Guía de los pastos de Zacatecas; Instituto Politécnico Nacional: Zacatecas, Mexico, 2010; pp. 19–149. [Google Scholar]
- Villaseñor, J.L. Checklist of the native vascular plants of Mexico. Rev. Mex Biodivers. 2016, 87, 559–902. [Google Scholar] [CrossRef] [Green Version]
- Villaseñor, J.L.; Espinosa-García, J.F. The alien flowering plants of Mexico. Divers. Distrib. 2004, 10, 113–123. [Google Scholar] [CrossRef]
- Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. Malezas de México. Available online: http://www.conabio.gob.mx/malezasdemexico/2inicio/paginas/lista-plantas-abr2006.htm (accessed on 8 June 2021).
- Burgos-Hernández, M.; Castillo-Campos, G.; Vergara Tenorio, M.D.C. Potentially useful flora from the tropical rainforest in central Veracruz, Mexico: Considerations for their conservation. Acta Bot Mex. 2014, 109, 55–77. [Google Scholar] [CrossRef] [Green Version]
- Clark Labs. TerrSet. Available online: https://clarklabs.org/ (accessed on 13 January 2021).
- QGIS Association. QGIS Geographic Information System. Available online: http://www.qgis.org (accessed on 13 January 2021).
- Yang, J.; Jin, Z.; Yang, J.Y.; Zhang, D.; Frangi, A.F. Essence of kernel Fisher discriminant: KPCA plus LDA. Comm. Com. Inf. Sc. 2004, 37, 2097–2100. [Google Scholar] [CrossRef]
- Rzedowski, J. Diversidad y orígenes de la flora fanerogámica de México. Acta Bot. Mex. 1991, 14, 3–21. [Google Scholar] [CrossRef] [Green Version]
- Koleff, P.; Soberón, J.; Smith, A. Madrean Pine-Oax Woodlands. In Hotspots Revisited; Mittermeier, R.A., Robles Gil, P., Hoffman, M., Pilgrim, J., Brooks, T., Goettsch Mittermeier, C., Lamoreux, J., da Fonseca, G.A.B., Eds.; CEMEX: Ciudad de México, Mexico, 2004; pp. 103–112. [Google Scholar]
- Sabas-Rosales, J.L. Encinos (Quercus: Fagaceae) de Zacatecas: Taxonomía, Diversidad y Distribución. Ph.D. Thesis, Universidad Autónoma de Aguascalientes, Aguascalientes, Mexico, 2016. [Google Scholar]
- Granados-Sánchez, D.; Sánchez-González, A.; Granados Victorino, R.L.; Borja de la Rosa, A. Ecología de la vegetación del desierto chihuahuense. Rev. Chapingo Ser. Cie. 2011, 17, 111–130. [Google Scholar] [CrossRef]
- Barrera-Zubiaga, E.J.; Granados-Sánchez, D.; Granados-Victorino, R.L.; Luna-Cavazos, M. Characterization of four pinyon pine forests in the state of Zacatecas, Mexico. Rev. Chapingo Ser. Cie. 2018, 24, 275–296. [Google Scholar] [CrossRef]
- González-Elizondo, M.S.; González-Elizondo, M.; Márquez-Linares, M.A. Vegetación y Ecorregiones de Durango; Instituto Politécnico Nacional: Durango, Mexico, 2007; pp. 71–109. [Google Scholar]
- Acosta-Hernández, A.C. Papel Sucesional de Dodonea viscosa en la Cuenca alta del Río San Pedro Mezquital. Master’s Thesis, Instituto Politécnico Nacional, Durango, Mexico, 2016. [Google Scholar]
- Rivas, G.R. Disturbios y Variables Ambientales Asociados a la Presencia de Dodonaea viscosa en los Municipios de Durango y Mezquital, Durango. Master’s Thesis, Instituto Politécnico Nacional, Durango, Mexico, 2019. [Google Scholar]
- Márquez-Linares, M.A.; Escobar-Flores, J.G.; Sandoval, S.; Pérez-Verdín, G. Multitemporal distribution analysis of Dodonaea viscosa (L.) Jacq. by remote sensing in Durango, Mexico. AGROProductividad 2020, 13, 61–67. [Google Scholar] [CrossRef]
- Reyes-Agüero, J.A.; Vallejo, M. Matorral crasicaule. In San Luis Potosí, Estudio de Estado, 2nd ed.; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad La Biodiversidad: Mexico, Mexico, 2019; pp. 47–54. [Google Scholar]
- Possley, J.; Maschinski, J. Competitive effects of the invasive grass Rhynchelytrum repens (Willd.) C.E. Hubb. on pine rockland vegetation. Nat. Area J. 2006, 26, 391–395. [Google Scholar] [CrossRef]
- Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. Rynchelyctrum repens (Willd.) C. E. Hubb. Available online: http://www.conabio.gob.mx/malezasdemexico/poaceae/rhynchelytrum (accessed on 7 June 2021).
- Trevizo, H.E. Impacto Ecológico del Zacate Rosado (Melinis repens (Willd.)) con Diferentes Años de Invasión en Pastizales de Chihuahua, México. Master’s Thesis, Universidad Autónoma de Chihuahua, Chihuahua, Mexico, 2018. [Google Scholar]
- Rapoport, E.H. Remarks on the biogeography of land invasions. Rev. Chil. Hist. Nat. 2000, 73, 367–380. [Google Scholar] [CrossRef]
- Instituto Nacional de Estadística y Geografía. Densidad de Población. Available online: http://cuentame.inegi.org.mx/poblacion/densidad.aspx?tema=P (accessed on 28 September 2021).
- Instituto Nacional de Estadística y Geografía. División Municipal. Available online: http://cuentame.inegi.org.mx/monografias/informacion/zac/territorio/div_municipal.aspx?tema=me&e=32 (accessed on 28 September 2021).
- Ramírez-Prieto, J.; Koch, S.D.; Balleza-Cadengo, J.D.J.; Adame-González, M.; Romero-Nápoles, J. Flora de la cima de la Mesa Alta, Jerez, Zacatecas, México. Bot. Sci. 2016, 94, 357–375. [Google Scholar] [CrossRef] [Green Version]
- Kerns, B.K.; Tortorelli, C.; Day, M.A.; Nietupski, T.; Barros, A.M.; Kim, J.B.; Krawchuk, M.A. Invasive grasses: A new perfect storm for forested ecosystems? Forest Ecol. Manag. 2020, 463, 1–15. [Google Scholar] [CrossRef]
- McDonald, J.A.; Martinez, J.; Nesom, G.L. Alpine flora of Cerro Mohinora, Chihuahua, Mexico. J. Bot. Res. Inst. Texas 2011, 5, 701–705. [Google Scholar]
- Rzedowski, J. Vegetación de México; Editorial Limusa: Ciudad de México, Mexico, 1978; pp. 247–327. [Google Scholar]
- Granados-Sánchez, D.; Sánchez-González, A. Clasificación fisonómica de la vegetación de la Sierra de Catorce, San Luis Potosí, a lo largo de un gradiente altitudinal. Terra Latinoamericana 2003, 21, 321–332. [Google Scholar]
- Márquez-Linares, M.A.; Jurado Ybarra, E.; González-Elizondo, M.S. Algunos aspectos de la biología de la manzanita (Arctostaphylos pungens HBK) y su papel en el desplazamiento de bosques templados por chaparrales. Ciencia UANL 2006, 9, 57–64. [Google Scholar]
- Casas, S.R.; González-Elizondo, S.; Tena-Flores, J.A. Estructura y tendencias sucesionales en vegetación de clima templado semi-seco en Durango, México. Madroño 1995, 42, 501–515. [Google Scholar]
- Challenger, A. Utilización y Conservación de los Ecosistemas Terrestres de México: Pasado Presente y Futuro; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: Ciudad de México, Mexico, 1998; pp. 727–795. [Google Scholar]
- Meave, J.A.; Rincón, A.; Romero-Romero, M.A. Oak forests of the hyper-humid region of La Chinantla, Northern Oaxaca Range, Mexico. In Ecology and Conservation of Neotropical Montane Oak Forests; Kappelle, M., Ed.; Springer: Berlin, Germany, 2006; pp. 113–125. [Google Scholar]
- Oldfield, S.; Eastwood, A. The Red List of Oaks; Fauna & Flora International: Cambridge, UK, 2007; pp. 5–7. [Google Scholar]
- Canada Centre for Remote Sensing. Cobertura del Suelo de México, 2011, a 250 Metros. Available online: http://www.conabio.gob.mx/informacion/gis/ (accessed on 7 June 2021).
- Trejo-Vázquez, I. El clima de la selva baja caducifolia en México. Investigaciones Geográficas 1999, 39, 40–52. [Google Scholar] [CrossRef]
- Secretaría de Medio Ambiente y Recursos Naturales. Inventario Estatal Forestal y de Suelos—Zacatecas 2014; Secretaría de Medio Ambiente y Recursos Naturales: Ciudad de México, Mexico, 2015; pp. 65–165.
- Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. Selvas Secas. Available online: https://www.biodiversidad.gob.mx/ecosistemas/selvaSeca (accessed on 14 June 2021).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oak Forests | Pine Forests | Chaparral | Nopaleras | Grasslands | Rock Outcrop Vegetation | |
|---|---|---|---|---|---|---|
| Oak forests | 93 | 0.0001 | 0.0001 | 0.0003 | 0.0001 | 0.0001 |
| Pine forests | 0.8224 | 107 | 0.0001 | 0.0001 | 0.0001 | 0.0001 |
| Chaparral | 0.8841 | 0.9938 | 73 | 0.0001 | 0.0001 | 0.0001 |
| Nopaleras | 0.8716 | 0.9244 | 0.9653 | 99 | 0.0001 | 0.0002 |
| Grasslands | 0.962 | 0.9262 | 0.9689 | 0.9733 | 74 | 0.0001 |
| Rock outcrop vegetation | 0.9191 | 0.8702 | 0.9462 | 0.8626 | 0.808 | 95 |
| Plant Communities | Indicator Species | IndVal | Contribution to Total Dissimilarity (%) |
|---|---|---|---|
| Oak forests | Quercus rugosa Née | 69.93 | 14.41 |
| Pine forests | Pinus cembroides Zucc. | 99.01 | 11.63 |
| Chaparral | Arctostaphylos pungens Kunth | 99.18 | 8.93 |
| Nopaleras | Opuntia leucotricha DC | 78.85 | 3.62 |
| Grasslands | Cyperus seslerioides Kunth | 85.20 | 2.66 |
| Rock outcrop vegetation | Agave schidigera Lem. | 92.00 | 0.66 |
| Total: 41.90 | |||
| Plant Communities | Family | Species | H | MH | C/A | F | AEIV |
|---|---|---|---|---|---|---|---|
| Oak forests | Fagaceae | Quercus potosina Trel. | A | 7 | 7 | 20 | 8.24 |
| Oak forests | Fagaceae | Quercus eduardi Trel. | A | 8 | 7 | 20 | 7.18 |
| Oak forests | Pinaceae | Pinus cembroides Zucc. | A | 3 | 2 | 30 | 1.87 |
| Pine forests | Asparagaceae | Dasylirion acrotrichum (Schiede) Zucc. | Ar | 0.9 | 2 | 10 | 0.43 |
| Pine forests | Asteraceae | Schkuhria pinnata (Lam,) Kuntze ex Tgell. | H | 0.6 | 2 | 30 | 1.27 |
| Pine forests | Asteraceae | Tagetes micrantha Cav. | H | 0.2 | 2 | 30 | 1.35 |
| Pine forests | Cactaceae | Opuntia leucotricha DC. | Ar | 0.6 | 1 | 20 | 0.85 |
| Pine forests | Cupressaceae | Juniperus deppeana Steud. | A | 9 | 2 | 20 | 0.9 |
| Pine forests | Cupressaceae | Juniperus flaccida Schltdl. | A | 8 | 4 | 80 | 9.82 |
| Pine forests | Fagaceae | Quercus potosina Trel. | A | 7 | 2 | 10 | 0.86 |
| Pine forests | Fagaceae | Quercus grisea Liebm. | A | 5 | 3 | 20 | 1.29 |
| Pine forests | Pinaceae | Pinus cembroides Zucc. | A | 11 | 8 | 100 | 60.5 |
| Chaparral | Ericaceae | Arctostaphylos pungens Kunth | Ar | 4 | 8 | 100 | 82.7 |
| Chaparral | Ericaceae | Comarostaphylis polifolia (Kunth) Zucc. Ex Klotzsch | Ar | 1.5 | 4 | 30 | 4.99 |
| Chaparral | Fagaceae | Quercus potosina Trel. | A | 4.5 | 7 | 10 | 4.9 |
| Nopaleras | Amaranthaceae | Gomphrena serrata L. | H | 0.1 | 2 | 20 | 0.98 |
| Nopaleras | Asparagaceae | Yucca decipiens Trel. | A | 6 | 2 | 10 | 0.49 |
| Nopaleras | Asteraceae | Brickellia secundiflora var. nepetifolia (Kunth) B. L. Rob. | Ar | 1.5 | 2 | 40 | 2.03 |
| Nopaleras | Asteraceae | Heterosperma pinnatum Cav. | H | 0.05 | 2 | 10 | 0.77 |
| Nopaleras | Cactaceae | Mammillaria heyderi Muehlenpf. | H | 0.2 | 3 | 60 | 3.01 |
| Nopaleras | Cactaceae | Opuntia leucotricha DC | Ar | 4 | 6 | 80 | 22.5 |
| Nopaleras | Convolvulaceae | Dichondra argéntea Humb, and Bonpl. Ex Willd. | H | 0.05 | 2 | 10 | 0.44 |
| Nopaleras | Fabaceae | Dalea bicolor Humb. and Bonpl. Ex Willd. | Ar | 2 | 4 | 50 | 4.47 |
| Nopaleras | Fabaceae | Mimosa monancistra Benth. | Ar | 1.0 | 3 | 60 | 8.57 |
| Nopaleras | Fabaceae | Vachellia schaffneri (S. Watson) Seigler and Ebinger | Ar | 3.5 | 5 | 40 | 6.06 |
| Grasslands | Asteraceae | Schkuhria pinnata (Lam.) Kuntze | H | 0.15 | 3 | 40 | 3.72 |
| Grasslands | Poaceae | Aristida schiedeana Trin. and Rupr. | H | 0.4 | 3 | 10 | 0.78 |
| Grasslands | Poaceae | Bouteloua repens (Kunth) Scribn. and Merr. | H | 0.16 | 3 | 60 | 3.94 |
| Grasslands | Poaceae | Eragrostis intermedia Hitchc. | H | 0.2 | 3 | 10 | 0.78 |
| Grasslands | Poaceae | Muhlenbergia phleoides (Kunth) Columbus | H | 0.35 | 6 | 60 | 13.5 |
| Grasslands | Poaceae | Muhlenbergia rigida (Kunth) Kunth | H | 0.8 | 7 | 60 | 11.87 |
| Rock outcrop vegetation | Asparagaceae | Agave schidigera Lem. | H | 0.5 | 6 | 100 | 11.65 |
| Rock outcrop vegetation | Asteraceae | Schkuhria pinnata (Lam.) Kuntze | H | 0.2 | 1 | 20 | 1.08 |
| Rock outcrop vegetation | Asteraceae | Stevia salicifolia Cav. | H | 0.5 | 2 | 30 | 1.64 |
| Rock outcrop vegetation | Asteraceae | Tagetes lunulata Ortega | H | 0.35 | 2 | 30 | 1.85 |
| Rock outcrop vegetation | Asteraceae | Tagetes micrantha Cav. | H | 0.15 | 5 | 70 | 5.22 |
| Rock outcrop vegetation | Asteraceae | Tridax palmeri A. Gray | H | 0.9 | 3 | 10 | 0.77 |
| Rock outcrop vegetation | Cactaceae | Echinocereus acifer (Otto ex Salm-Dyck) J.N.Haage | H | 0.22 | 3 | 50 | 4.45 |
| Rock outcrop vegetation | Cactaceae | Mammillaria moelleriana Boed. | H | 0.12 | 3 | 40 | 2.46 |
| Rock outcrop vegetation | Cactaceae | Stenocactus ochoterenianus Tiegel | H | 0.2 | 5 | 90 | 8.74 |
| Rock outcrop vegetation | Cistaceae | Crocanthemum glomeratum (Lag.) Janch. | H | 0.22 | 3 | 40 | 2.87 |
| Rock outcrop vegetation | Cyperaceae | Cyperus seslerioides Kunth | H | 0.12 | 6 | 40 | 5.72 |
| Rock outcrop vegetation | Ericaceae | Arctostaphylos pungens Kunth | Ar | 2 | 1 | 10 | 0.62 |
| Rock outcrop vegetation | Fagaceae | Quercus potosina Trel. | Ar | 1.5 | 1 | 10 | 0.62 |
| Rock outcrop vegetation | Pinaceae | Pinus cembroides Zucc. | A | 2 | 2 | 60 | 3.69 |
| Rock outcrop vegetation | Poaceae | Muhlenbergia emersleyi Vasey | H | 0.45 | 9 | 10 | 26.9 |
| Plant Communities | Families | Genera | Species | Endemic Species | Exclusive Species | Exotic Species |
|---|---|---|---|---|---|---|
| Oak forests | 38 | 74 | 96 | 33 | 58 | 1 |
| Pine forest | 42 | 76 | 107 | 33 | 20 | 1 |
| Chaparral | 29 | 52 | 73 | 29 | 17 | 0 |
| Nopaleras | 34 | 81 | 99 | 24 | 40 | 2 |
| Grasslands | 30 | 54 | 74 | 22 | 16 | 2 |
| Rock outcrops vegetation | 42 | 74 | 95 | 31 | 25 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hurtado-Reveles, L.; Burgos-Hernández, M.; López-Acosta, J.C.; Vázquez-Sánchez, M. Importance of Local Studies of Vascular Plant Communities in Conservation and Management: A Case Study in Susticacán, Zacatecas, Mexico. Diversity 2021, 13, 492. https://doi.org/10.3390/d13100492
Hurtado-Reveles L, Burgos-Hernández M, López-Acosta JC, Vázquez-Sánchez M. Importance of Local Studies of Vascular Plant Communities in Conservation and Management: A Case Study in Susticacán, Zacatecas, Mexico. Diversity. 2021; 13(10):492. https://doi.org/10.3390/d13100492
Chicago/Turabian StyleHurtado-Reveles, Leopoldo, Mireya Burgos-Hernández, Juan Carlos López-Acosta, and Monserrat Vázquez-Sánchez. 2021. "Importance of Local Studies of Vascular Plant Communities in Conservation and Management: A Case Study in Susticacán, Zacatecas, Mexico" Diversity 13, no. 10: 492. https://doi.org/10.3390/d13100492
APA StyleHurtado-Reveles, L., Burgos-Hernández, M., López-Acosta, J. C., & Vázquez-Sánchez, M. (2021). Importance of Local Studies of Vascular Plant Communities in Conservation and Management: A Case Study in Susticacán, Zacatecas, Mexico. Diversity, 13(10), 492. https://doi.org/10.3390/d13100492