
Zobellia barbeyronii sp. nov., a New Member of the Family Flavobacteriaceae, Isolated from Seaweed, and Emended Description of the Species Z. amurskyensis, Z. laminariae, Z. russellii and Z. uliginosa

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolation and Cultivation
2.2. 16S rRNA Gene Sequencing and Phylogenetic Analysis
2.3. Multilocus Sequence Analysis (MLSA)
2.4. Genome Features of Strain 36-CHABK-3-33T and Phylogenomic Reconstruction
2.5. Physiology and Chemotaxonomy
3. Results and Discussion
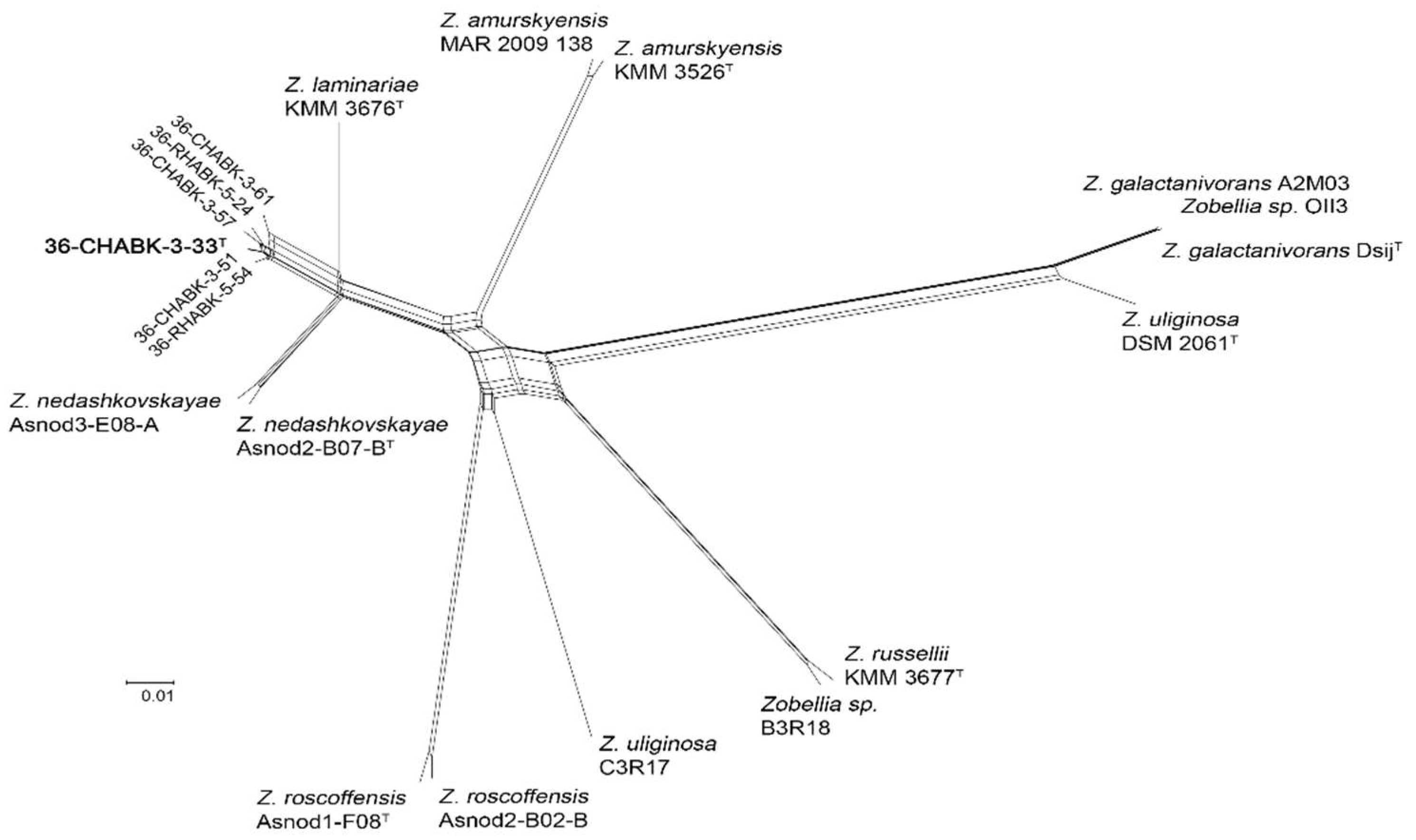
3.1. Phylogenetic Analysis
3.2. Multilocus Sequence Analysis of the Alga-Associated Isolates
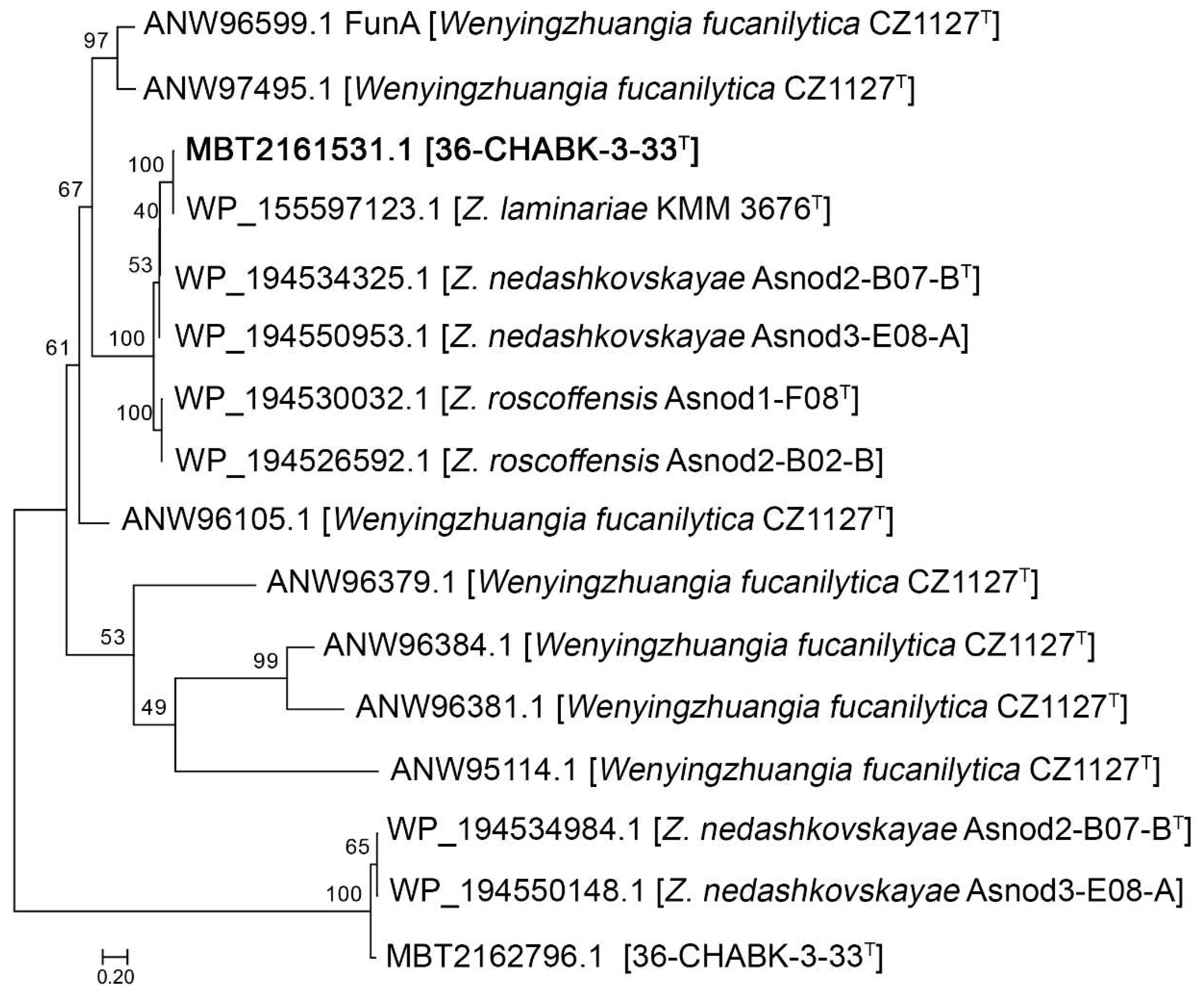
3.3. Genomic Characteristics and Phylogenomic Analysis
3.4. Morphological, Physiological and Biochemical Characteristics
3.5. Chemotaxonomic Characterization
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barbeyron, T.; l’Haridon, S.; Corre, E.; Kloareg, B.; Potin, P. Zobellia galactanovorans gen. nov., sp. nov., a marine species of Flavobacteriaceae isolated from a red alga, and classification of [Cytophaga] uliginosa (ZoBell and Upham 1944) Reichenbach 1989 as Zobellia uliginosa gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 2001, 51, 985–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LPSN—List of Prokaryotic Names with Standing in Nomenclature. Available online: https://www.bacterio.net/ (accessed on 1 September 2021).
- Nedashkovskaya, O.I.; Suzuki, M.; Vancanneyt, M.; Cleenwerck, I.; Lysenko, A.M.; Mikhailov, V.V.; Swings, J. Zobellia amurskyensis sp. nov., Zobellia laminariae sp. nov. and Zobellia russellii sp. nov., novel marine bacteria of the family Flavobacteriaceae. Int. J. Syst. Evol. Microbiol. 2004, 54, 1643–1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbeyron, T.; Thiébaud, M.; Le Duff, N.; Martin, M.; Corre, E.; Tanguy, G.; Vandenbol, M.; Thomas, F. Zobellia roscoffensis sp. nov. and Zobellia nedashkovskayae sp. nov., two flavobacteria from the epiphytic microbiota of the brown alga Ascophyllum nodosum, and emended description of the genus Zobellia. Int. J. Syst. Evol. Microbiol. 2021, 71, 004913. [Google Scholar] [CrossRef]
- ZoBell, C.E. A list of marine bacteria including descriptions of sixty new species. Bull. Scripps. Inst. Oceanog. Univ. Calif. 1944, 5, 239–292. [Google Scholar]
- Reichenbach, H.; Genus, I. Cytophaga Winogradsky 1929, 577AL emend. In Bergey’s Manual of Systematic Bacteriology; Staley, J.T., Bryant, M.P., Eds.; Williams & Wilkins: Baltimore, MD, USA, 1989; Volume 3, pp. 2015–2050. [Google Scholar]
- Bowman, J.P. Description of Cellulophaga algicola sp. nov., isolated from the surfaces of Antarctic algae, and reclassification of Cytophaga uliginosa (ZoBell and Upham 1944) Reichenbach 1989 as Cellulophaga uliginosa comb. nov. Int. J. Syst. Evol. Microbiol. 2000, 50, 1861–1868. [Google Scholar] [CrossRef] [Green Version]
- Nedashkovskaya, O.I.; Suzuki, M.; Genus, L.X.I. Zobellia. In Bergey’s Manual of Systematic Bacteriology; Krieg, N.R., Ludwig, W., Eds.; Springer: New York, NY, USA, 2011; Volume 4, pp. 292–295. [Google Scholar]
- Barbeyron, T.; Thomas, F.; Barbe, V.; Teeling, H.; Schenowitz, C.; Dossat, C.; Goesmann, A.; Leblanc, C.; Glöckner, F.O.; Czjzek, M.; et al. Habitat and taxon as driving forces of carbohydrate catabolism in marine heterotrophic bacteria: Example of the model algae-associated bacterium Zobellia galactanivorans DsijT. Environ. Microbiol. 2016, 18, 4610–4627. [Google Scholar] [CrossRef] [Green Version]
- Chernysheva, N.; Bystritskaya, E.; Stenkova, A.; Golovkin, I.; Nedashkovskaya, O.; Isaeva, M. Comparative genomics and CAZyme genome repertoires of marine Zobellia amurskyensis KMM 3526T and Zobellia laminariae KMM 3676T. Mar. Drugs 2019, 17, 661. [Google Scholar] [CrossRef] [Green Version]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley & Sons: New York, NY, USA, 1991; pp. 115–147. [Google Scholar]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Huson, D.H.; Kloepper, T.; Bryant, D. SplitsTree 4.0-Computation of phylogenetic trees and networks. Bioinformatics 2008, 14, 68–73. [Google Scholar] [CrossRef] [PubMed]
- FastQC. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 26 April 2018).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [Green Version]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef] [PubMed]
- Asnicar, F.; Thomas, A.M.; Beghini, F.; Mengoni, C.; Manara, S.; Manghi, P.; Zhu, Q.; Bolzan, M.; Cumbo, F.; May, U.; et al. Precise phylogenetic analysis of microbial isolates and genomes from meta-genomes using PhyloPhlAn 3.0. Nat. Commun. 2020, 11, 2500. [Google Scholar] [CrossRef]
- Rodriguez-R, L.M.; Konstantinidis, K.T. The enveomics collection: A toolbox for specialized analyses of microbial genomes and metagenomes. PeerJ Prepr. 2016, 4, e1900v1. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Göker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef]
- Zhang, H.; Yohe, T.; Huang, L.; Entwistle, S.; Wu, P.; Yang, Z.; Busk, P.K.; Xu, Y.; Yin, Y. dbCAN2: A meta server for automated carbohydrate-active enzyme annotation. Nucleic Acids Res. 2018, 46, W95–W101. [Google Scholar] [CrossRef] [Green Version]
- Smibert, R.M.; Krieg, N.R. Phenotypic characterization. In Methods for General and Molecular Bacteriology; Gerhardt, P., Murray, R.G.E., Wood, W.A., Krieg, N.R., Eds.; American Society for Microbiology: Washington, DC, USA, 1994; pp. 607–654. [Google Scholar]
- Lemos, M.L.; Toranzo, A.E.; Barja, J.L. Modified medium for the oxidation-fermentation test in the identification of marine bacteria. Appl. Environ. Microbiol. 1985, 49, 1541–1543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fautz, E.; Reichenbach, H. A simple test for flexirubin-type pigments. FEMS Microbiol. Lett. 1980, 8, 87–90. [Google Scholar] [CrossRef]
- Sasser, M. Identification of Bacteria by Gas Chromatography of Cellular Fatty Acids; MIDI Technical Note 101; MIDI Inc.: Newark, DE, USA, 1990. [Google Scholar]
- Minnikin, D.E.; Patel, P.V.; Alshamaony, L.; Goodfellow, M. Polar lipid composition in the classification of Nocardia and related bacteria. Int. J. Syst. Evol. Microbiol. 1977, 27, 104–117. [Google Scholar] [CrossRef] [Green Version]
- Konstantinidis, K.T.; Tiedje, J.M. Genomic insights that advance the species definition for prokaryotes. PNAS 2005, 102, 2567–2572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. PNAS 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [Green Version]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [Green Version]
- Chernysheva, N.; Bystritskaya, E.; Likhatskaya, G.; Nedashkovskaya, O.; Isaeva, M. Genome-wide analysis of PL7 alginate lyases in the genus Zobellia. Molecules 2021, 26, 2387. [Google Scholar] [CrossRef]
- Shen, J.; Chang, Y.; Zhang, Y.; Mei, X.; Xue, C. Discovery and characterization of an endo-1,3-fucanase from marine bacterium Wenyingzhuangia fucanilytica: A novel glycoside hydrolase family. Front. Microbiol. 2020, 11, 1674. [Google Scholar] [CrossRef]
- Bernardet, J.F.; Nakagawa, Y.; Holmes, B. Proposed minimal standards for describing new taxa of the family Flavobacteriaceae and emended description of the family. Int. J. Syst. Evol. Microbiol. 2002, 52, 1049–1070. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feature | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
|---|---|---|---|---|---|---|---|---|
| Assembly level | Contig | Contig | Contig | Contig | Contig | Contig | Complete Genome | Contig |
| Genome size (Mb) | 4.98 | 4.94 | 5.15 | 5.14 | 4.98 | 5.30 | 5.52 | 4.92 |
| Number of contigs | 37 | 2 | 27 | 120 | 3 | 27 | 1 | 49 |
| G+C Content (%) | 36.7 | 36.8 | 36.8 | 38.0 | 37.6 | 42.6 | 42.8 | 39.0 |
| N50 (bp) | 927.759 | 4.937.885 | 904.812 | 94.524 | 4.613.932 | 914.779 | 644.008 | |
| L50 (bp) | 3 | 1 | 3 | 17 | 1 | 3 | 3 | |
| Coverage | 24 | 224 | 75 | 16 | 253 | 254 | 27 | |
| Total genes | 4114 | 4074 | 4285 | 4241 | 4041 | 4381 | 4486 | 4025 |
| Protein coding genes | 4053 | 4013 | 4177 | 4157 | 3981 | 4298 | 4416 | 3967 |
| rRNAs(5S/16S/23S) | 1/1/1 | 2/1/1 | 1/1/1 | 1/1/1 | 2/1/1 | 1/1/1 | 2/2/2 | 1/1/2 |
| tRNA | 39 | 39 | 39 | 37 | 38 | 37 | 40 | 36 |
| checkM completeness (%) | 100.00 | 100.00 | 99.68 | 100.00 | 100.00 | 100.00 | 100.00 | 99.68 |
| checkM contamination (%) | 1.07 | 1.07 | 1.07 | 0.97 | 0.65 | 0.81 | 1.07 | 0.00 |
| Accession number | JACATN000000000.1 | JADDXR000000000.1 | RCNS00000000.1 | RCNR00000000.1 | JADDXT000000000.1 | FTOB00000000.1 | FP476056.1 | JACSOI000000000.1 |
| Characteristic | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| Source of isolation * | Green and red algae | Brown alga | Brown alga | Red alga |
| Temperature range for growth (°C) | 4–32 | 4–40 * | 4–30 | 4–42 |
| Salinity range for growth (% NaCl) | 0.5–8 | 3–6 * | 1.5–6 | 0.5–8 |
| Degradation of | ||||
| Casein | − | − | − | + |
| Gelatin | + | − | + | + |
| Starch | − | − * | − | + |
| DNA | + | + | − | − |
| Tween 20 | + | + | − | + |
| Tween 40 | − | + * | + | − |
| Tween 80 | + | v | − | − |
| Acid formation from | ||||
| L-Rhamnose | + | + * | + | − |
| Raffinose | − | − * | + | − |
| D-Xylose | + | + | − | − |
| N-Acetyl-glucosamine | − | + * | − | − |
| Cystine arylamidaseactivity | + | + * | − | + |
| α-Chymotrypsin | − | + * | − | − |
| DNA G+C content (mol%) | 36.7 | 37.6–37.7 * | 36.8 | 42.8 |
| Fatty Acid | 1 | 2 * | 3 | 4 |
|---|---|---|---|---|
| Branched | ||||
| iso-C15:0 | 14.0 | 20.9 | 17.4 | 17.1 |
| anteiso-C15:0 | 1.7 | 1.3 | 2.0 | 1.9 |
| iso-C15:1 G | 13.5 | 10.7 | 14.4 | 14.9 |
| iso-C17:1 ω8c | 3.8 | – | 3.0 | 7.5 |
| iso-C17:1 ω9c | – | 7.9 | – | – |
| Saturated | ||||
| C15:0 | 11.1 | 8.6 | 5.8 | 5.5 |
| C16:0 | – | 1.1 | 1.1 | |
| Unsaturated | ||||
| C15:1 ω6c | 3.9 | 1.7 | 1.0 | 1.8 |
| C17:1 ω6c | 1.9 | 1.0 | – | tr |
| C18:1 ω5c | – | 1.5 | – | – |
| Hydroxy | ||||
| iso-C15:0 3-OH | 5.2 | 3.6 | 5.4 | 6.4 |
| iso-C17:0 3-OH | 21.1 | 21.2 | 29.1 | 27.1 |
| C15:0 3-OH | 1.1 | – | – | tr |
| C16:0 3-OH | 3.5 | 1.1 | 5.6 | 2.6 |
| Summed feature 3 | 17.1 | 13.8 | 13.0 | 9.6 |
| Summed feature 4 | 1.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nedashkovskaya, O.; Otstavnykh, N.; Zhukova, N.; Guzev, K.; Chausova, V.; Tekutyeva, L.; Mikhailov, V.; Isaeva, M. Zobellia barbeyronii sp. nov., a New Member of the Family Flavobacteriaceae, Isolated from Seaweed, and Emended Description of the Species Z. amurskyensis, Z. laminariae, Z. russellii and Z. uliginosa. Diversity 2021, 13, 520. https://doi.org/10.3390/d13110520
Nedashkovskaya O, Otstavnykh N, Zhukova N, Guzev K, Chausova V, Tekutyeva L, Mikhailov V, Isaeva M. Zobellia barbeyronii sp. nov., a New Member of the Family Flavobacteriaceae, Isolated from Seaweed, and Emended Description of the Species Z. amurskyensis, Z. laminariae, Z. russellii and Z. uliginosa. Diversity. 2021; 13(11):520. https://doi.org/10.3390/d13110520
Chicago/Turabian StyleNedashkovskaya, Olga, Nadezhda Otstavnykh, Natalia Zhukova, Konstantin Guzev, Viktoria Chausova, Liudmila Tekutyeva, Valery Mikhailov, and Marina Isaeva. 2021. "Zobellia barbeyronii sp. nov., a New Member of the Family Flavobacteriaceae, Isolated from Seaweed, and Emended Description of the Species Z. amurskyensis, Z. laminariae, Z. russellii and Z. uliginosa" Diversity 13, no. 11: 520. https://doi.org/10.3390/d13110520
APA StyleNedashkovskaya, O., Otstavnykh, N., Zhukova, N., Guzev, K., Chausova, V., Tekutyeva, L., Mikhailov, V., & Isaeva, M. (2021). Zobellia barbeyronii sp. nov., a New Member of the Family Flavobacteriaceae, Isolated from Seaweed, and Emended Description of the Species Z. amurskyensis, Z. laminariae, Z. russellii and Z. uliginosa. Diversity, 13(11), 520. https://doi.org/10.3390/d13110520