
Developing Inside a Layer of Germs—A Potential Role for Multiciliated Surface Cells in Vertebrate Embryos


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
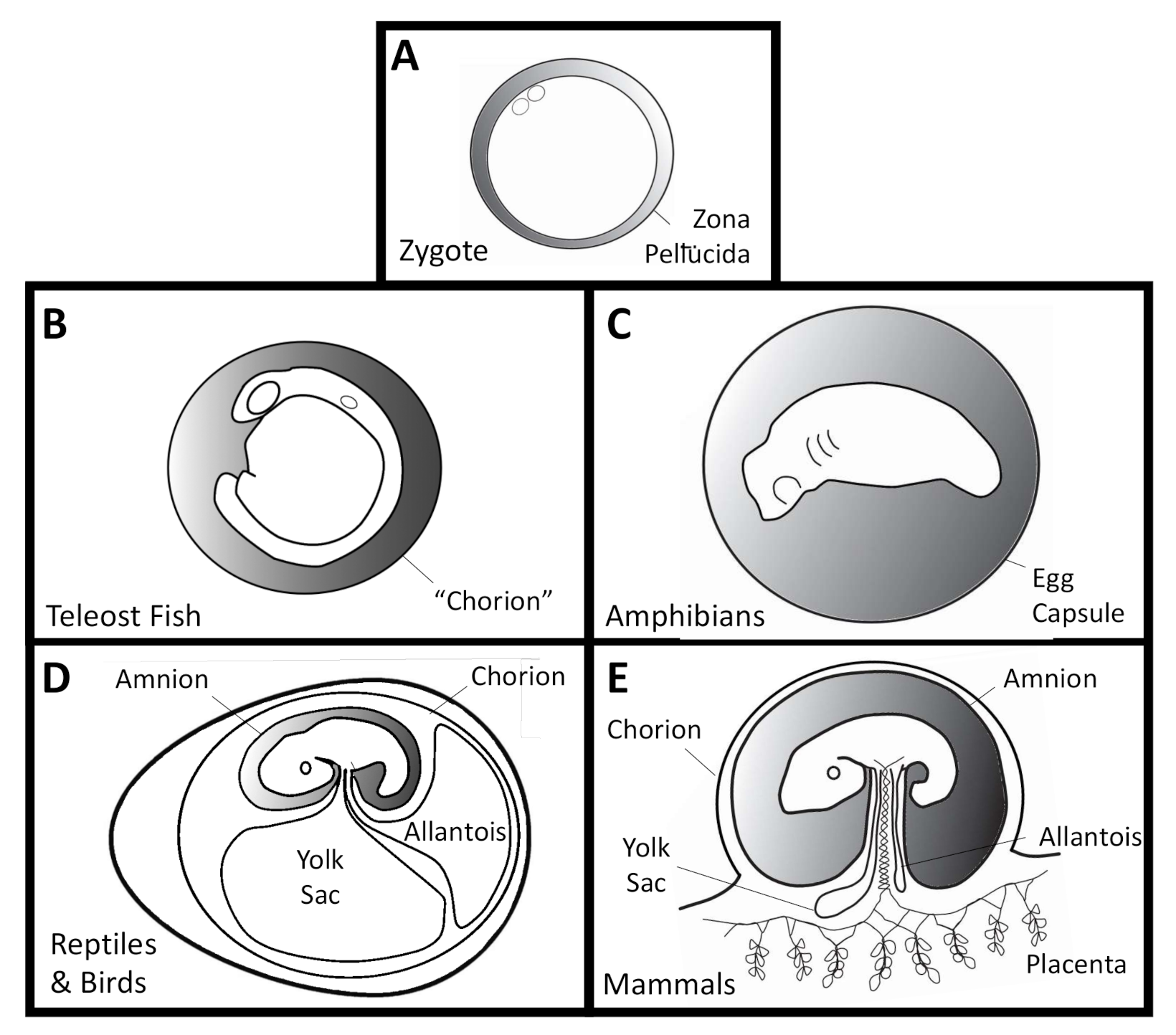
2. Terminology: In Ovo and In Utero
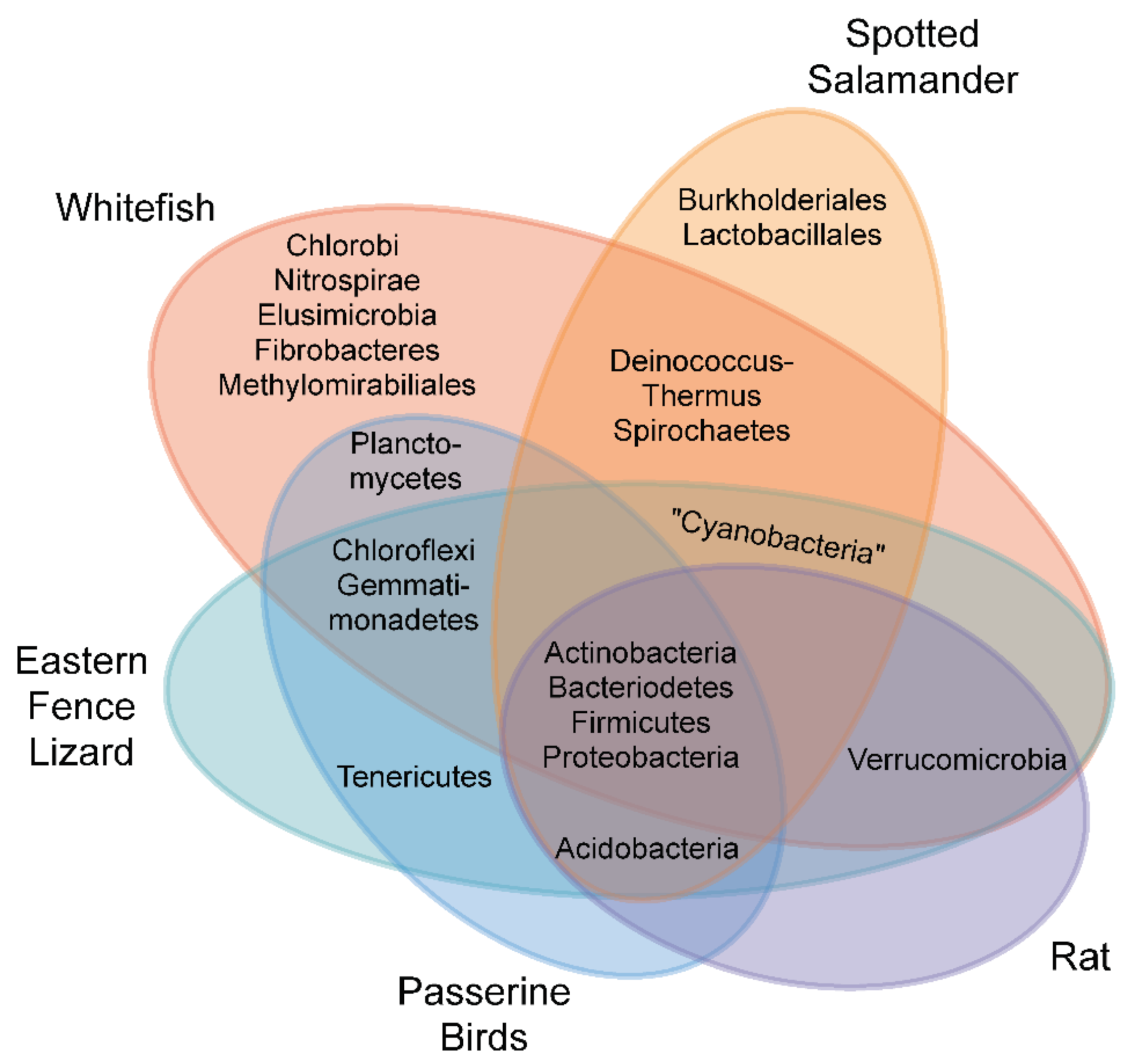
3. Methods for Identifying Microbial Consortia
4. Evidence for In Ovo and In Utero Microbiotas in Healthy Vertebrates
4.1. Fish
4.2. Amphibians
4.3. Reptiles and Birds
4.4. Mammals
5. Gnotobiototic Vertebrates
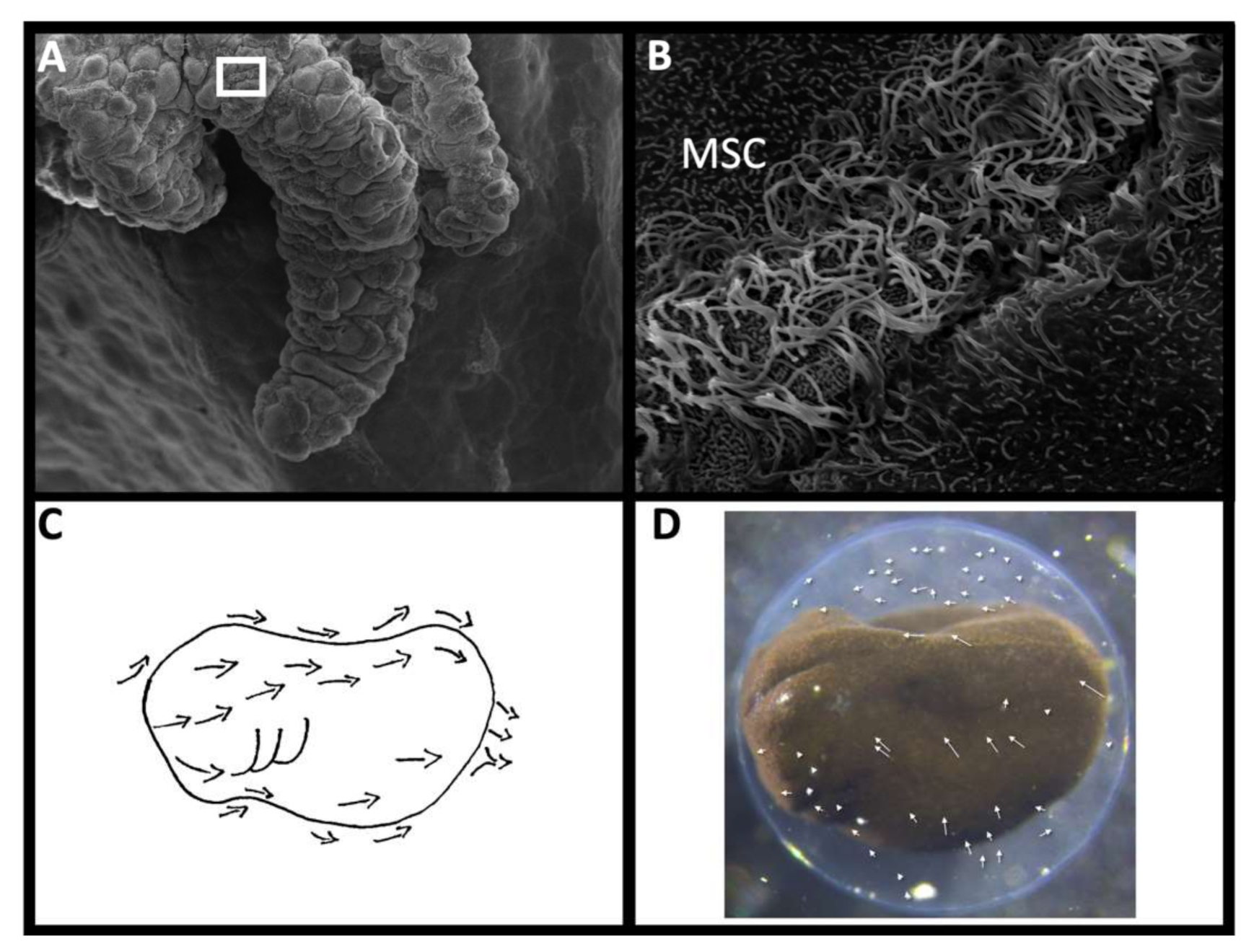
6. Multiciliated Surface Cells
6.1. Vertebrate Embryos with Multiciliated Surface Cells
6.2. Origin and Morphology of MSCs
6.3. Proposed Functions of MSCs
6.4. Boundary Layers in Microbial Recruitment
7. Conclusions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McFall-Ngai, M.; Bosch, T.C.G. Animal development in the microbial world: The power of experimental model systems. In Current Topics in Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2021; Volume 141, pp. 371–397. ISBN 978-0-12-814968-3. [Google Scholar]
- Bosch, T.C.G.; McFall-Ngai, M. Animal development in the microbial world: Re-thinking the conceptual framework. In Current Topics in Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2021; Volume 141, pp. 399–427. ISBN 978-0-12-814968-3. [Google Scholar]
- Popescu, L.; Cao, Z.-P. From Microscopy to Genomic Approach in Soil Biodiversity Assessment. Curr. Issues Mol. Biol. 2018, 27, 195–198. [Google Scholar] [CrossRef]
- Burnham, P.; Gomez-Lopez, N.; Heyang, M.; Cheng, A.P.; Lenz, J.S.; Dadhania, D.M.; Lee, J.R.; Suthanthiran, M.; Romero, R.; De Vlaminck, I. Separating the Signal from the Noise in Metagenomic Cell-Free DNA Sequencing. Microbiome 2020, 8, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, T.J.; Sanders, J.G.; Fierer, N. Not All Animals Need a Microbiome. FEMS Microbiol. Lett. 2019, 366. [Google Scholar] [CrossRef] [Green Version]
- Nyholm, S.V. In the Beginning: Egg–Microbe Interactions and Consequences for Animal Hosts. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190593. [Google Scholar] [CrossRef]
- Kellam, L.; Pastorelli, L.M.; Bastida, A.M.; Senkbeil, A.; Montgomery, S.; Fishel, S.; Campbell, A. Perivitelline Threads in Cleavage-Stage Human Embryos: Observations Using Time-Lapse Imaging. Reprod. Biomed. Online 2017, 35, 646–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, L.; Suter, M.J.-F.; Räsänen, K. Evolution of Egg Coats: Linking Molecular Biology and Ecology. Mol. Ecol. 2015, 24, 4052–4073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claw, K.G.; Swanson, W.J. Evolution of the Egg: New Findings and Challenges. Annu. Rev. Genom. Hum. Genet. 2012, 13, 109–125. [Google Scholar] [CrossRef]
- Alteri, A.; Viganò, P.; Maizar, A.A.; Jovine, L.; Giacomini, E.; Rubino, P. Revisiting Embryo Assisted Hatching Approaches: A Systematic Review of the Current Protocols. J. Assist. Reprod. Genet. 2018, 35, 367–391. [Google Scholar] [CrossRef]
- Altig, R.; McDiarmid, R.W. Morphological Diversity and Evolution of Egg and Clutch Structure in Amphibians. Herpetol. Monogr. 2007, 21, 1–32. [Google Scholar] [CrossRef]
- Laale, H.W. The Perivitelline Space and Egg Envelopes of Bony Fishes: A Review. Copeia 1980, 1980, 210–226. [Google Scholar] [CrossRef]
- Kirunda, D.F.K.; McKee, S.R. Relating Quality Characteristics of Aged Eggs and Fresh Eggs to Vitelline Membrane Strength as Determined by a Texture Analyzer. Poult. Sci. 2000, 79, 1189–1193. [Google Scholar] [CrossRef]
- Damaziak, K.; Kieliszek, M.; Bucław, M. Characterization of Structure and Protein of Vitelline Membranes of Precocial (Ring-Necked Pheasant, Gray Partridge) and Superaltricial (Cockatiel Parrot, Domestic Pigeon) Birds. PLoS ONE 2020, 15, e0228310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryder, J. Development of the Silver Gar (Belone Longirostris) with Observations on the Genesis of the Blood in Embryo Fishes and a Comparison of Fish Ova with Those of Other Vertebrates. Bull. U. S. Fish Comm. 1882, 1, 283–301. [Google Scholar]
- Salthe, S.N. The Egg Capsules in the Amphibia. J. Morphol. 1963, 113, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Szarski, H. The Origin of Vertebrate Foetal Membranes. Evolution 1968, 22, 211–214. [Google Scholar] [CrossRef] [PubMed]
- Escherich, T. The Intestinal Bacteria of the Neonate and Breast-Fed Infant. Infect. Dis. 1885, 11, 352–356. [Google Scholar] [CrossRef] [PubMed]
- Reasoner, D.J.; Geldreich, E.E. A New Medium for the Enumeration and Subculture of Bacteria from Potable Water. Appl. Environ. Microbiol. 1985, 49, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Browne, H.P.; Forster, S.C.; Anonye, B.O.; Kumar, N.; Neville, B.A.; Stares, M.D.; Goulding, D.; Lawley, T.D. Culturing of ‘Unculturable’ Human Microbiota Reveals Novel Taxa and Extensive Sporulation. Nature 2016, 533, 543–546. [Google Scholar] [CrossRef] [Green Version]
- Walker, A.W.; Duncan, S.H.; Louis, P.; Flint, H.J. Phylogeny, Culturing, and Metagenomics of the Human Gut Microbiota. Trends Microbiol. 2014, 22, 267–274. [Google Scholar] [CrossRef]
- Kerney, R.; Kim, E.; Hangarter, R.P.; Heiss, A.A.; Bishop, C.D.; Hall, B.K. Intracellular Invasion of Green Algae in a Salamander Host. Proc. Natl. Acad. Sci. USA 2011, 108, 6497–6502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seferovic, M.D.; Pace, R.M.; Carroll, M.; Belfort, B.; Major, A.M.; Chu, D.M.; Racusin, D.A.; Castro, E.C.C.; Muldrew, K.L.; Versalovic, J.; et al. Visualization of Microbes by 16S in Situ Hybridization in Term and Preterm Placentas without Intraamniotic Infection. Am. J. Obstet. Gynecol. 2019, 221, 146.e1–146.e23. [Google Scholar] [CrossRef] [PubMed]
- Borneman, J.; Hartin, R.J. PCR Primers That Amplify Fungal RRNA Genes from Environmental Samples. Appl. Environ. Microbiol. 2000, 66, 4356–4360. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.; Simpson, A.G.; Graham, L.E. Evolutionary Relationships of Apusomonads Inferred from Taxon-Rich Analyses of 6 Nuclear Encoded Genes. Mol. Biol. Evol. 2006, 23, 2455–2466. [Google Scholar] [CrossRef] [Green Version]
- Frank, J.A.; Reich, C.I.; Sharma, S.; Weisbaum, J.S.; Wilson, B.A.; Olsen, G.J. Critical Evaluation of Two Primers Commonly Used for Amplification of Bacterial 16S RRNA Genes. Appl. Environ. Microbiol. 2008, 74, 2461–2470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anslan, S.; Sachs, M.; Rancilhac, L.; Brinkmann, H.; Petersen, J.; Künzel, S.; Schwarz, A.; Arndt, H.; Kerney, R.; Vences, M. Diversity and Substrate-Specificity of Green Algae and Other Micro-Eukaryotes Colonizing Amphibian Clutches in Germany, Revealed by DNA Metabarcoding. Sci. Nat. 2021, 108, 29. [Google Scholar] [CrossRef] [PubMed]
- Collado, M.C.; Rautava, S.; Aakko, J.; Isolauri, E.; Salminen, S. Human Gut Colonization May Be Initiated in utero by Distinct Microbial Communities in the Placenta and Amniotic Fluid. Sci. Rep. 2016, 6, 23129. [Google Scholar] [CrossRef] [Green Version]
- Hazen, T.C.; Dubinsky, E.A.; DeSantis, T.Z.; Andersen, G.L.; Piceno, Y.M.; Singh, N.; Jansson, J.K.; Probst, A.; Borglin, S.E.; Fortney, J.L.; et al. Deep-Sea Oil Plume Enriches Indigenous Oil-Degrading Bacteria. Science 2010, 330, 204–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balvočiūtė, M.; Huson, D.H. SILVA, RDP, Greengenes, NCBI and OTT—How Do These Taxonomies Compare? BMC Genom. 2017, 18, 114. [Google Scholar] [CrossRef] [Green Version]
- Eisenhofer, R.; Minich, J.J.; Marotz, C.; Cooper, A.; Knight, R.; Weyrich, L.S. Contamination in Low Microbial Biomass Microbiome Studies: Issues and Recommendations. Trends Microbiol. 2019, 27, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, L.G.E.; Rogivue, A.; Fumagalli, L.; Wedekind, C. Declining Diversity of Egg-Associated Bacteria during Development of Naturally Spawned Whitefish Embryos (Coregonus Spp.). Aquat. Sci. 2015, 77, 481–497. [Google Scholar] [CrossRef]
- Bishop, C.; Jurga, E.; Graham, L. Patterns of Bacterial Diversity in Embryonic Capsules of the Spotted Salamander Ambystoma Maculatum: An Expanding View of a Symbiosis. FEMS Microbiol. Ecol. 2021, 97, fiab128. [Google Scholar] [CrossRef] [PubMed]
- Borghi, E.; Massa, V.; Severgnini, M.; Fazio, G.; Avagliano, L.; Menegola, E.; Bulfamante, G.P.; Morace, G.; Borgo, F. Antenatal Microbial Colonization of Mammalian Gut. Reprod. Sci. 2019, 26, 1045–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trevelline, B.K.; MacLeod, K.J.; Knutie, S.A.; Langkilde, T.; Kohl, K.D. In ovo Microbial Communities: A Potential Mechanism for the Initial Acquisition of Gut Microbiota among Oviparous Birds and Lizards. Biol. Lett. 2018, 14. [Google Scholar] [CrossRef] [PubMed]
- Bakke, I.; Attramadal, K.J.K.; Vestrum, R.I.; Vadstein, O. Controlling Factors for Community Assembly in Developing Cod Larvae (Gadus morhua). In Microbial Communities in Aquaculture Ecosystems; Derome, N., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 69–93. ISBN 978-3-030-16189-7. [Google Scholar]
- Cook, M.I.; Beissinger, S.R.; Toranzos, G.A.; Rodriguez, R.A.; Arendt, W.J. Trans–Shell Infection by Pathogenic Micro–Organisms Reduces the Shelf Life of Non–Incubated Bird’s Eggs: A Constraint on the Onset of Incubation? Proc. R. Soc. Lond. B Biol. Sci. 2003, 270, 2233–2240. [Google Scholar] [CrossRef] [Green Version]
- Theis, K.R.; Romero, R.; Greenberg, J.M.; Winters, A.D.; Garcia-Flores, V.; Motomura, K.; Ahmad, M.M.; Galaz, J.; Arenas-Hernandez, M.; Gomez-Lopez, N. No Consistent Evidence for Microbiota in Murine Placental and Fetal Tissues. mSphere 2020, 5. [Google Scholar] [CrossRef] [Green Version]
- Froese, R.; Pauly, D. FishBase. World Wide Web Electronic Publication; ScienceOpen, Inc.: Burlington, MA, USA, 2014. [Google Scholar]
- Balon, E. Epigenesis of an Epigeneticist: The Development of Some Alternative Concepts on the Early Ontogeny and Evolution of Fishes. Guelph Ichthyol. Rev. 1990, 1, 1–48. [Google Scholar]
- Kemp, A. Role of Epidermal Cilia in Development of the Australian Lungfish, Neoceratodus Forsteri (Osteichthyes: Dipnoi). J. Morphol. 1996, 228, 203–221. [Google Scholar] [CrossRef]
- De Schryver, P.; Vadstein, O. Ecological Theory as a Foundation to Control Pathogenic Invasion in Aquaculture. ISME J. 2014, 8, 2360–2368. [Google Scholar] [CrossRef] [Green Version]
- Gauthier, J.; Lavoie, C.; Charette, S.J.; Derome, N. Host-Microbiota Interactions and Their Importance in Promoting Growthand Resistance to Opportunistic Diseases in Salmonids. In Microbial Communities in Aquaculture Ecosystems: Improving Productivity and Sustainability; Derome, N., Ed.; Springer International Publishing: Cham, Switzerland, 2019; ISBN 978-3-030-16189-7. [Google Scholar]
- Derome, N.; Filteau, M. A Continuously Changing Selective Context on Microbial Communities Associated with Fish, from Egg to Fork. Evol. Appl. 2020, 13, 1298–1319. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn, M.S.; Boutin, S.; Hoseinifar, S.H.; Derome, N. Teleost Microbiomes: The State of the Art in Their Characterization, Manipulation and Importance in Aquaculture and Fisheries. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Rawls, J.F.; Samuel, B.S.; Gordon, J.I. Gnotobiotic Zebrafish Reveal Evolutionarily Conserved Responses to the Gut Microbiota. Proc. Natl. Acad. Sci. USA 2004, 101, 4596–4601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkins, L.G.E.; Fumagalli, L.; Wedekind, C. Effects of Host Genetics and Environment on Egg-Associated Microbiotas in Brown Trout (Salmo trutta). Mol. Ecol. 2016, 25, 4930–4945. [Google Scholar] [CrossRef] [Green Version]
- Wilkins, L.G.E.; Rogivue, A.; Schütz, F.; Fumagalli, L.; Wedekind, C. Increased Diversity of Egg-Associated Bacteria on Brown Trout (Salmo trutta) at Elevated Temperatures. Sci. Rep. 2015, 5, 17084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; de Bruijn, I.; Jack, A.L.; Drynan, K.; van den Berg, A.H.; Thoen, E.; Sandoval-Sierra, V.; Skaar, I.; van West, P.; Di guez-Uribeondo, J.; et al. Deciphering Microbial Landscapes of Fish Eggs to Mitigate Emerging Diseases. ISME J. 2014, 8, 2002–2014. [Google Scholar] [CrossRef]
- Hansen, G.H.; Olafsen, J.A. Bacterial Colonization of Cod (Gadus morhua L.) and Halibut (Hippoglossus hippoglossus) Eggs in Marine Aquaculture. Appl. Environ. Microbiol. 1989, 55, 1435–1446. [Google Scholar] [CrossRef] [Green Version]
- Stephens, W.Z.; Burns, A.R.; Stagaman, K.; Wong, S.; Rawls, J.F.; Guillemin, K.; Bohannan, B.J.M. The Composition of the Zebrafish Intestinal Microbial Community Varies across Development. ISME J. 2016, 10, 644–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eddy, F.B.; Ward, M.R.; Talbot, C.; Primmett, D. Ionic Movements across the Chorion in Newly Shed Salmon Eggs (Salmo salar L.). J. Comp. Physiol. B 1990, 159, 771–776. [Google Scholar] [CrossRef]
- Groot, E.P.; Alderdice, D.F. Fine Structure of the External Egg Membrane of Five Species of Pacific Salmon and Steelhead Trout. Can. J. Zool. 1985, 63, 552–566. [Google Scholar] [CrossRef]
- Yanagimachi, R.; Harumi, T.; Matsubara, H.; Yan, W.; Yuan, S.; Hirohashi, N.; Iida, T.; Yamaha, E.; Arai, K.; Matsubara, T.; et al. Chemical and Physical Guidance of Fish Spermatozoa into the Egg through the Micropyle. Biol. Reprod. 2017, 96, 780–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Stockhammer, O.W.; de Boer, L.; Vischer, N.O.E.; Spaink, H.P.; Grijpma, D.W.; Zaat, S.A.J. The Zebrafish Embryo as a Model to Quantify Early Inflammatory Cell Responses to Biomaterials: The Zebrafish Embryo Model. J. Biomed. Mater. Res. A 2017, 105, 2522–2532. [Google Scholar] [CrossRef]
- Benard, E.L.; van der Sar, A.M.; Ellett, F.; Lieschke, G.J.; Spaink, H.P.; Meijer, A.H. Infection of Zebrafish Embryos with Intracellular Bacterial Pathogens. J. Vis. Exp. 2012, 3781. [Google Scholar] [CrossRef] [PubMed]
- van der Sar, A.M.; Spaink, H.P.; Zakrzewska, A.; Bitter, W.; Meijer, A.H. Specificity of the Zebrafish Host Transcriptome Response to Acute and Chronic Mycobacterial Infection and the Role of Innate and Adaptive Immune Components. Mol. Immunol. 2009, 46, 2317–2332. [Google Scholar] [CrossRef] [PubMed]
- De la Paz, J.F.; Anguita-Salinas, C.; Díaz-Celis, C.; Chávez, F.P.; Allende, M.L. The Zebrafish Perivitelline Fluid Provides Maternally-Inherited Defensive Immunity. Biomolecules 2020, 10, 1274. [Google Scholar] [CrossRef] [PubMed]
- Roth, O.; Beemelmanns, A.; Barribeau, S.M.; Sadd, B.M. Recent Advances in Vertebrate and Invertebrate Transgenerational Immunity in the Light of Ecology and Evolution. Heredity 2018, 121, 225–238. [Google Scholar] [CrossRef] [Green Version]
- Gensollen, T.; Iyer, S.S.; Kasper, D.L.; Blumberg, R.S. How Colonization by Microbiota in Early Life Shapes the Immune System. Science 2016, 352, 539–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archetti, M.; Ubeda, F.; Fudenberg, D.; Green, J.; Pierce, N.E.; Yu, D.W. Let the Right One in: A Microeconomic Approach to Partner Choice in Mutualisms. Am. Nat. 2011, 177, 75–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wedekind, C.; Gessner, M.O.; Vazquez, F.; Maerki, M.; Steiner, D. Elevated Resource Availability Sufficient to Turn Opportunistic into Virulent Fish Pathogens. Ecology 2010, 91, 1251–1256. [Google Scholar] [CrossRef] [Green Version]
- Clark, E.S.; Wilkins, L.G.E.; Wedekind, C. MHC Class I Expression Dependent on Bacterial Infection and Parental Factors in Whitefish Embryos (Salmonidae). Mol. Ecol. 2013, 22, 5256–5269. [Google Scholar] [CrossRef] [Green Version]
- Clark, E.S.; Pompini, M.; Marques da Cunha, L.; Wedekind, C. Maternal and Paternal Contributions to Pathogen Resistance Dependent on Development Stage in a Whitefish (Salmonidae). Funct. Ecol. 2014, 28, 714–723. [Google Scholar] [CrossRef]
- Beemelmanns, A.; Poirier, M.; Bayer, T.; Kuenzel, S.; Roth, O. Microbial Embryonal Colonization during Pipefish Male Pregnancy. Sci. Rep. 2019, 9, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wells, K. The Ecology and Behavior of Amphibians; University of Chicago Press: Chicago, IL, USA, 2007. [Google Scholar]
- Beattie, R.C. A Physico-Chemical Investigation of the Jelly Capsules Surrounding Eggs of the Common Frog (Rana temporaria temporaria). J. Zool. 2009, 190, 1–25. [Google Scholar] [CrossRef]
- Gomez-Mestre, I.; Touchon, J.C.; Warkentin, K.M. Amphibian Embryo and Parental Defenses and a Larval Predator Reduce Egg Mortality from Water Mold. Ecology 2006, 87, 2570–2581. [Google Scholar] [CrossRef]
- Hamdoun, A.; Epel, D. Embryo Stability and Vulnerability in an Always Changing World. Proc. Natl. Acad. Sci. USA 2007, 104, 1745–1750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebollar, E.A.; Martínez-Ugalde, E.; Orta, A.H. The Amphibian Skin Microbiome and Its Protective Role Against Chytridiomycosis. Herpetologica 2020, 76, 167. [Google Scholar] [CrossRef]
- Loudon, A.H.; Venkataraman, A.; Van Treuren, W.; Woodhams, D.C.; Parfrey, L.W.; McKenzie, V.J.; Knight, R.; Schmidt, T.M.; Harris, R.N. Vertebrate Hosts as Islands: Dynamics of Selection, Immigration, Loss, Persistence, and Potential Function of Bacteria on Salamander Skin. Front. Microbiol. 2016, 7, 333. [Google Scholar] [CrossRef]
- Hughey, M.C.; Delia, J.; Belden, L.K. Diversity and Stability of Egg-Bacterial Assemblages: The Role of Paternal Care in the Glassfrog Hyalinobatrachium colymbiphyllum. Biotropica 2017, 49, 792–802. [Google Scholar] [CrossRef]
- Banning, J.L.; Weddle, A.L.; Wahl III, G.W.; Simon, M.A.; Lauer, A.; Walters, R.L.; Harris, R.N. Antifungal Skin Bacteria, Embryonic Survival, and Communal Nesting in Four-Toed Salamanders, Hemidactylium scutatum. Oecologia 2008, 156, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Walke, J.B.; Becker, M.H.; Loftus, S.C.; House, L.L.; Cormier, G.; Jensen, R.V.; Belden, L.K. Amphibian Skin May Select for Rare Environmental Microbes. ISME J. 2014, 8, 2207–2217. [Google Scholar] [CrossRef] [PubMed]
- Prest, T.L.; Kimball, A.K.; Kueneman, J.G.; McKenzie, V.J. Host-Associated Bacterial Community Succession during Amphibian Development. Mol. Ecol. 2018, 27, 1992–2006. [Google Scholar] [CrossRef]
- Orr, H. Note on the Development of Amphibians, Chiefly Concerning the Central Nervous System; with Additional Observations on the Hypophysis, Mouth, and the Appendages and Skeleton of the Head. J. Cell Sci. 1888, 115, 295–324. [Google Scholar]
- Gilbert, P. Observations on the Eggs of Ambystoma maculatum with Especial Reference to the Green Algae Found Within the Egg Envelopes. Ecology 1942, 23, 215–227. [Google Scholar] [CrossRef] [Green Version]
- Hutchison, V.H.; Hammen, C.S. Oxygen Utilization in the Symbiosis of Embryos of the Salamander, Ambystoma maculatum and the Alga, Oophila amblystomatis. Biol. Bull. 1958, 115, 483–489. [Google Scholar] [CrossRef]
- Jurga, E.; Graham, L.; Bishop, C.D. Oophila Is Monophyletic within a Three-Taxon Eukaryotic Microbiome in Egg Masses of the Salamander Ambystoma maculatum. Symbiosis 2020, 81, 187–199. [Google Scholar] [CrossRef]
- Ibelings, B.W.; De Bruin, A.; Kagami, M.; Rijkeboer, M.; Brehm, M.; Donk, E.V. Host Parasite Interactions between Freshwater Phytoplankton and Chytrid Fungi (Chyridiomycota). J. Phycol. 2004, 40, 437–453. [Google Scholar] [CrossRef]
- Blaustein, A.R.; Belden, L.K. Amphibian Defenses against Ultraviolet-B Radiation. Evol. Dev. 2003, 5, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Cook, M.I.; Beissinger, S.R.; Toranzos, G.A.; Arendt, W.J. Incubation Reduces Microbial Growth on Eggshells and the Opportunity for Trans-Shell Infection. Ecol. Lett. 2005, 8, 532–537. [Google Scholar] [CrossRef]
- Godard, R.D.; Morgan Wilson, C.; Frick, J.W.; Siegel, P.B.; Bowers, B.B. The Effects of Exposure and Microbes on Hatchability of Eggs in Open-Cup and Cavity Nests. J. Avian Biol. 2007, 38, 709–716. [Google Scholar] [CrossRef]
- Van Veelen, H.P.J.; Salles, J.F.; Tieleman, B.I. Microbiome Assembly of Avian Eggshells and Their Potential as Transgenerational Carriers of Maternal Microbiota. ISME J. 2018, 12, 1375–1388. [Google Scholar] [CrossRef]
- Grond, K.; Lanctot, R.B.; Jumpponen, A.; Sandercock, B.K. Recruitment and Establishment of the Gut Microbiome in Arctic Shorebirds. FEMS Microbiol. Ecol. 2017, 93, fix142. [Google Scholar] [CrossRef]
- Wielen, P.W.J.J.; Keuzenkamp, D.A.; Lipman, L.J.A.; Knapen, F.; Biesterveld, S. Spatial and Temporal Variation of the Intestinal Bacterial Community in Commercially Raised Broiler Chickens During Growth. Microb. Ecol. 2002, 44, 286–293. [Google Scholar] [CrossRef]
- Soler, J.J.; Martín-Vivaldi, M.; Peralta-Sánchez, J.M.; Arco, L.; Juárez-García-Pelayo, N. Hoopoes Color Their Eggs with Antimicrobial Uropygial Secretions. Naturwissenschaften 2014, 101, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Soler, J.J.; Martín-Vivaldi, M.; Ruiz-Rodríguez, M.; Valdivia, E.; Martín-Platero, A.M.; Martínez-Bueno, M.; Peralta-Sánchez, J.M.; Méndez, M. Symbiotic Association between Hoopoes and Antibiotic-Producing Bacteria That Live in Their Uropygial Gland. Funct. Ecol. 2008, 22, 864–871. [Google Scholar] [CrossRef] [Green Version]
- Martín-Vivaldi, M.; Soler, J.J.; Peralta-Sánchez, J.M.; Arco, L.; Martín-Platero, A.M.; Martínez-Bueno, M.; Ruiz-Rodríguez, M.; Valdivia, E. Special Structures of Hoopoe Eggshells Enhance the Adhesion of Symbiont-Carrying Uropygial Secretion That Increase Hatching Success. J. Anim. Ecol. 2014, 83, 1289–1301. [Google Scholar] [CrossRef] [PubMed]
- Martín-Platero, A.M.; Valdivia, E.; Ruíz-Rodríguez, M.; Soler, J.J.; Martín-Vivaldi, M.; Maqueda, M.; Martínez-Bueno, M. Characterization of Antimicrobial Substances Produced by Enterococcus faecalis MRR 10-3, Isolated from the Uropygial Gland of the Hoopoe (Upupa epops). Appl. Environ. Microbiol. 2006, 72, 4245–4249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Law-Brown, J.; Meyers, P.R. Enterococcus phoeniculicola Sp. Nov., a Novel Member of the Enterococci Isolated from the Uropygial Gland of the Red-Billed Woodhoopoe, Phoeniculus purpureus. Int. J. Syst. Evol. Microbiol. 2003, 53, 683–685. [Google Scholar] [CrossRef] [PubMed]
- Sarmiento-Ramírez, J.M.; van der Voort, M.; Raaijmakers, J.M.; Diéguez-Uribeondo, J. Unravelling the Microbiome of Eggs of the Endangered Sea Turtle Eretmochelys imbricata Identifies Bacteria with Activity against the Emerging Pathogen Fusarium falciforme. PLoS ONE 2014, 9, e95206. [Google Scholar] [CrossRef]
- Bézy, V.S.; Valverde, R.A.; Plante, C.J. Olive Ridley Sea Turtle Hatching Success as a Function of the Microbial Abundance in Nest Sand at Ostional, Costa Rica. PLoS ONE 2015, 10, e0118579. [Google Scholar] [CrossRef]
- Shine, R. Life-History Evolution in Reptiles. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 23–46. [Google Scholar] [CrossRef] [Green Version]
- Kohl, K.D.; Brun, A.; Magallanes, M.; Brinkerhoff, J.; Laspiur, A.; Acosta, J.C.; Caviedes-Vidal, E.; Bordenstein, S.R. Gut Microbial Ecology of Lizards: Insights into Diversity in the Wild, Effects of Captivity, Variation across Gut Regions and Transmission. Mol. Ecol. 2017, 26, 1175–1189. [Google Scholar] [CrossRef]
- Yockey, L.J.; Lucas, C.; Iwasaki, A. Contributions of Maternal and Fetal Antiviral Immunity in Congenital Disease. Science 2020, 368, 608–612. [Google Scholar] [CrossRef]
- Feiner, N.; de Souza-Lima, S.; Jorge, F.; Naem, S.; Aubret, F.; Uller, T.; Nadler, S.A. Vertical Transmission of a Nematode from Female Lizards to the Brains of Their Offspring. Am. Nat. 2020, 195, 918–926. [Google Scholar] [CrossRef] [PubMed]
- Hubert, J. The embryology of the squamata. In Biology of Reptilia; Wiley: New York, NY, USA, 1985; pp. 1–34. [Google Scholar]
- Gantois, I.; Ducatelle, R.; Pasmans, F.; Haesebrouck, F.; Gast, R.; Humphrey, T.J.; Van Immerseel, F. Mechanisms of Egg Contamination by Salmonella enteritidis. FEMS Microbiol. Rev. 2009, 33, 718–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Reu, K.; Grijspeerdt, K.; Messens, W.; Heyndrickx, M.; Uyttendaele, M.; Debevere, J.; Herman, L. Eggshell Factors Influencing Eggshell Penetration and Whole Egg Contamination by Different Bacteria, Including Salmonella enteritidis. Int. J. Food Microbiol. 2006, 112, 253–260. [Google Scholar] [CrossRef]
- Cox, N.A.; Richardson, L.J.; Maurer, J.J.; Berrang, M.E.; Fedorka-Cray, P.J.; Buhr, R.J.; Byrd, J.A.; Lee, M.D.; Hofacre, C.L.; O’Kane, P.M.; et al. Evidence for Horizontal and Vertical Transmission in Campylobacter Passage from Hen to Her Progeny. J. Food Prot. 2012, 75, 1896–1902. [Google Scholar] [CrossRef]
- Salter, S.J.; Cox, M.J.; Turek, E.M.; Calus, S.T.; Cookson, W.O.; Moffatt, M.F.; Turner, P.; Parkhill, J.; Loman, N.J.; Walker, A.W. Reagent and Laboratory Contamination Can Critically Impact Sequence-Based Microbiome Analyses. BMC Biol. 2014, 12, 87. [Google Scholar] [CrossRef] [Green Version]
- Tyndale-Biscoe, H.; Renfree, M. Breeding biology of marsupials by family. In Reproductive Physiology of Marsupials; Cambridge University Press: Cambridge, UK, 1987; pp. 14–94. [Google Scholar]
- Yadav, M.; Stanley, N.F.; Waring, H. The Microbial Flora of the Gut of the Pouch-Young and the Pouch of a Marsupial, Setonix brachyurus. J. Gen. Microbiol. 1972, 70, 437–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deakin, J.E.; Cooper, D.W. Characterization of and Immunity to the Aerobic Bacteria Found in the Pouch of the Brushtail Possum Trichosurus vulpecula. Comp. Immunol. Microbiol. Infect. Dis. 2004, 27, 33–46. [Google Scholar] [CrossRef]
- Osawa, R.; Blanshard, W.H.; O’Callaghan, P.G. Microflora of the Pouch of the Koala (Phascolarctos cinereus). J. Wildl. Dis. 1992, 28, 276–280. [Google Scholar] [CrossRef]
- Weiss, S.; Taggart, D.; Smith, I.; Helgen, K.M.; Eisenhofer, R. Host Reproductive Cycle Influences the Pouch Microbiota of Wild Southern Hairy-Nosed Wombats (Lasiorhinus latifrons). Anim. Microbiome 2021, 3, 13. [Google Scholar]
- Chhour, K.-L.; Hinds, L.A.; Jacques, N.A.; Deane, E.M. An Observational Study of the Microbiome of the Maternal Pouch and Saliva of the Tammar Wallaby, Macropus Eugenii, and of the Gastrointestinal Tract of the Pouch Young. Microbiology 2010, 156, 798–808. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Fox, S.; Pemberton, D.; Hogg, C.; Papenfuss, A.T.; Belov, K. The Tasmanian Devil Microbiome—Implications for Conservation and Management. Microbiome 2015, 3, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capellini, I.; Nunn, C.L.; Barton, R.A. Microparasites and Placental Invasiveness in Eutherian Mammals. PLoS ONE 2015, 10, e0132563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malmuthuge, N.; Griebel, P.J. Fetal Environment and Fetal Intestine Are Sterile during the Third Trimester of Pregnancy. Vet. Immunol. Immunopathol. 2018, 204, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, E.; Fernández, L.; Marín, M.L.; Martín, R.; Odriozola, J.M.; Nueno-Palop, C.; Narbad, A.; Olivares, M.; Xaus, J.; Rodríguez, J.M. Isolation of Commensal Bacteria from Umbilical Cord Blood of Healthy Neonates Born by Cesarean Section. Curr. Microbiol. 2005, 51, 270–274. [Google Scholar] [CrossRef]
- Jiménez, E.; Marín, M.L.; Martín, R.; Odriozola, J.M.; Olivares, M.; Xaus, J.; Fernández, L.; Rodríguez, J.M. Is Meconium from Healthy Newborns Actually Sterile? Res. Microbiol. 2008, 159, 187–193. [Google Scholar] [CrossRef]
- Kustner, O. Beitrag Zur Lehre von Der Puerperalen Infection Der Neugeborenen. Arch Gynakol 1877, 11, 256–263. [Google Scholar] [CrossRef]
- Perez-Muñoz, M.E.; Arrieta, M.-C.; Ramer-Tait, A.E.; Walter, J. A Critical Assessment of the “Sterile Womb” and “in utero Colonization” Hypotheses: Implications for Research on the Pioneer Infant Microbiome. Microbiome 2017, 5, 1–19. [Google Scholar] [CrossRef]
- Funkhouser, L.J.; Bordenstein, S.R. Mom Knows Best: The Universality of Maternal Microbial Transmission. PLoS Biol. 2013, 11, e1001631. [Google Scholar] [CrossRef]
- Aagaard, K.; Ma, J.; Antony, K.M.; Ganu, R.; Petrosino, J.; Versalovic, J. The Placenta Harbors a Unique Microbiome. Sci. Transl. Med. 2014, 6, 237ra65. [Google Scholar] [CrossRef] [Green Version]
- Kliman, H.J. Comment on “The Placenta Harbors a Unique Microbiome”. Sci. Transl. Med. 2014, 6, 254le4. [Google Scholar] [CrossRef]
- Lauder, A.P.; Roche, A.M.; Sherrill-Mix, S.; Bailey, A.; Laughlin, A.L.; Bittinger, K.; Leite, R.; Elovitz, M.A.; Parry, S.; Bushman, F.D. Comparison of Placenta Samples with Contamination Controls Does Not Provide Evidence for a Distinct Placenta Microbiota. Microbiome 2016, 4, 29. [Google Scholar] [CrossRef] [Green Version]
- Antony, K.M.; Ma, J.; Mitchell, K.B.; Racusin, D.A.; Versalovic, J.; Aagaard, K. The Preterm Placental Microbiome Varies in Association with Excess Maternal Gestational Weight Gain. Am. J. Obstet. Gynecol. 2015, 212, 653.e1–653.e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.; Xiao, X.; Zhang, Q.; Mao, L.; Yu, M.; Xu, J. The Placental Microbiome Varies in Association with Low Birth Weight in Full-Term Neonates. Nutrients 2015, 7, 6924–6937. [Google Scholar] [CrossRef] [PubMed]
- Amarasekara, R.; Jayasekara, R.W.; Senanayake, H.; Dissanayake, V.H.W. Microbiome of the Placenta in Pre-Eclampsia Supports the Role of Bacteria in the Multifactorial Cause of Pre-Eclampsia: Pre-Eclampsia and Infectious Agents. J. Obstet. Gynaecol. Res. 2015, 41, 662–669. [Google Scholar] [CrossRef] [PubMed]
- Bassols, J.; Serino, M.; Carreras-Badosa, G.; Burcelin, R.; Blasco-Baque, V.; Lopez-Bermejo, A.; Fernandez-Real, J.-M. Gestational Diabetes Is Associated with Changes in Placental Microbiota and Microbiome. Pediatr. Res. 2016, 80, 777–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burrage, S. Bacteria in the Supposedly Sterile Meconium. J. Bacteriol. 1927, 13, 47–48. [Google Scholar]
- Hall, I.C. Bacterial Flora of First Specimens of Meconium Passed by Fifty New-Born Infants. Arch. Pediatr. Adolesc. Med. 1934, 47, 1279–1285. [Google Scholar] [CrossRef]
- Hansen, R.; Scott, K.P.; Khan, S.; Martin, J.C.; Berry, S.H.; Stevenson, M.; Okpapi, A.; Munro, M.J.; Hold, G.L. First-Pass Meconium Samples from Healthy Term Vaginally-Delivered Neonates: An Analysis of the Microbiota. PLoS ONE 2015, 10, e0133320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, J.W.; Brown, J.H. The Bacterial Content of the Uterus at Cesarean Section. Am. J. Obstet. Gynecol. 1927, 13, 133–143. [Google Scholar] [CrossRef]
- Stinson, L.F.; Keelan, J.A.; Payne, M.S. Identification and Removal of Contaminating Microbial DNA from PCR Reagents: Impact on Low-biomass Microbiome Analyses. Lett. Appl. Microbiol. 2019, 68, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Stinson, L.F.; Keelan, J.A.; Payne, M.S. Comparison of Meconium DNA Extraction Methods for Use in Microbiome Studies. Front. Microbiol. 2018, 9, 270. [Google Scholar] [CrossRef] [Green Version]
- Collado, M.C.; Segata, N. Initial Exploration of in utero Microbial Colonization. Nat. Med. 2020, 26, 469–470. [Google Scholar] [CrossRef]
- Rackaityte, E.; Halkias, J.; Fukui, E.M.; Mendoza, V.F.; Hayzelden, C.; Crawford, E.D.; Fujimura, K.E.; Burt, T.D.; Lynch, S.V. Viable Bacterial Colonization Is Highly Limited in the Human Intestine in utero. Nat. Med. 2020, 26, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-E.; Jardine, L.; Gottgens, B.; Teichmann, S.A.; Haniffa, M. Prenatal Development of Human Immunity. Science 2020, 368, 600–603. [Google Scholar] [CrossRef] [PubMed]
- Ganal-Vonarburg, S.C.; Hornef, M.W.; Macpherson, A.J. Microbial–Host Molecular Exchange and Its Functional Consequences in Early Mammalian Life. Science 2020, 368, 604–607. [Google Scholar] [CrossRef]
- Uchimura, Y.; Fuhrer, T.; Li, H.; Lawson, M.A.; Zimmermann, M.; Yilmaz, B.; Zindel, J.; Ronchi, F.; Sorribas, M.; Hapfelmeier, S.; et al. Antibodies Set Boundaries Limiting Microbial Metabolite Penetration and the Resultant Mammalian Host Response. Immunity 2018, 49, 545–559.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, I.; Miyamoto, J.; Ohue-Kitano, R.; Watanabe, K.; Yamada, T.; Onuki, M.; Aoki, R.; Isobe, Y.; Kashihara, D.; Inoue, D.; et al. Maternal Gut Microbiota in Pregnancy Influences Offspring Metabolic Phenotype in Mice. Science 2020, 367, eaaw8429. [Google Scholar] [CrossRef] [PubMed]
- Pasteur, L. Observations Relatives à La Note Précédente de M. Comptes Rendus Seances Acad. Sci. 1885, 100, 68. [Google Scholar]
- Teah, B.A. Bibliography of Germfree Research 1885–1963; Supplement 1977; Defense Technical Information Center: Fort Belvoir, VA, USA, 1977. [Google Scholar]
- Gordon, H.A. The Germ-Free Animal. Its Use in the Study of “Physiologic” Effects of the Normal Microbial Flora on the Animal Host. Am. J. Dig. Dis. 1960, 5, 841–867. [Google Scholar] [CrossRef] [PubMed]
- Gordon, H.A.; Pesti, L. The Gnotobiotic Animal as a Tool in the Study of Host Microbial Relationships. Bacteriol. Rev. 1971, 35, 390–429. [Google Scholar] [CrossRef] [PubMed]
- Timmons, E.H.; Olmsted, G.M.; Kaplan, H.M. The Germfree Leopard Frog (Rana pipiens): Preliminary Report. Lab. Anim. Sci. 1977, 27, 518–521. [Google Scholar] [PubMed]
- Hedrich, H.J.; Hardy, P. The Laboratory Mouse; Elsevier: London, UK, 2012. [Google Scholar]
- Melancon, E.; De La Torre Canny, S.G.; Sichel, S.; Kelly, M.; Wiles, T.J.; Rawls, J.F.; Eisen, J.S.; Guillemin, K. Best Practices for Germ-Free Derivation and Gnotobiotic Zebrafish Husbandry. Methods Cell Biol. 2017, 138, 61–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinn, I.K.; Shearer, W.T. Severe Combined Immunodeficiency Disorders. Immunol. Allergy Clin. N. Am. 2015, 35, 671–694. [Google Scholar] [CrossRef] [PubMed]
- Nokhbatolfoghahai, M.; Downie, J.R. Ciliated Cells on the Surface of Embryos of Scyliorhinus canicula. J. Fish Biol. 2003, 63, 523–527. [Google Scholar] [CrossRef]
- Nokhbatolfoghahai, M.; Downie, J.R.; Clelland, A.K.; Rennison, K. The Surface Ciliation of Anuran Amphibian Embryos and Early Larvae: Patterns, Timing Differences and Functions. J. Nat. Hist. 2005, 39, 887–929. [Google Scholar] [CrossRef]
- Nokhbatolfoghahai, M.; Downie, J.R.; Ogilvy, V. Surface Ciliation of Anuran Amphibian Larvae: Persistence to Late Stages in Some Species but Not Others. J. Morphol. 2006, 267, 1248–1256. [Google Scholar] [CrossRef]
- Kessel, R.G.; Beams, H.W.; Shih, C.Y. The Origin, Distribution and Disappearance of Surface Cilia during Embryonic Development of Rana pipiens as Revealed by Scanning Electron Microscopy. Am. J. Anat. 1974, 141, 341–359. [Google Scholar] [CrossRef] [PubMed]
- Morgan, T.H. The Development of the Frog’s Egg; Macmillan and Co.: New York, NY, USA, 1897. [Google Scholar]
- Dünker, N.; Wake, M.H.; Olson, W.M. Embryonic and Larval Development in the Caecilian Ichthyophis Kohtaoensis (Amphibia, Gymnophiona): A Staging Table. J. Morphol. 2000, 243, 3–34. [Google Scholar] [CrossRef]
- Fox, H. Early Development of Caecilian Skin with Special Reference to the Epidermis. J. Herpetol. 1986, 20, 154–167. [Google Scholar] [CrossRef]
- Harrison, R.G. Harrison Stages and Description of the Normal Development of the Spotted Salamander, Amblystoma punctatum. In Organization and Development of the Embryo; Yale University Press: New Haven, CT, USA, 1969; pp. 44–66. [Google Scholar]
- Landström, U. On the Differentiation of Prospective Ectoderm to a Ciliated Cell Pattern in Embryos of Ambystoma mexicanum. J. Embryol. Exp. Morphol. 1977, 41, 23–32. [Google Scholar]
- Tattersall, G.J.; Spiegelaar, N. Embryonic Motility and Hatching Success of Ambystoma maculatum Are Influenced by a Symbiotic Alga. Can. J. Zool. 2008, 86, 1289–1298. [Google Scholar] [CrossRef] [Green Version]
- Lannoo, M.J. Neuromast Topography in Anuran Amphibians. J. Morphol. 1987, 191, 115–129. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, J.I.; Doran, S.A.; Shartau, R.B.; Pon, J.R.; Ali, D.W.; Tam, R.; Kuang, S. Integrative Biology of an Embryonic Respiratory Behaviour in Pond Snails: The ‘embryo Stir-Bar Hypothesis’. J. Exp. Biol. 2008, 211, 1729–1736. [Google Scholar] [CrossRef] [Green Version]
- Wiley, A. Amphioxous and the Ancestry of Vertebrates; Macmillan: New York, NY, USA, 1894. [Google Scholar]
- Webb, J.F.; Northcutt, R.G. Ciliated Epidermal Cells in Non-Teleost Actinopterygian Fish. Acta Zool. 1991, 72, 107–111. [Google Scholar] [CrossRef]
- Bartsch, P.; Gemballa, S.; Piotrowski, T. The Embryonic and Larval Development of Polypterus senegalus Cuvier, 1829: Its Staging with Reference to External and Skeletal Features, Behaviour and Locomotory Habits. Acta Zool. 1997, 78, 309–328. [Google Scholar] [CrossRef]
- Song, J. The Lateral Line System in the Florida Gar, Lepisosteus platyrhincus, de Kay. Ph.D. Thesis, University of Michigan, Ann Arbor, MI, USA, 1989. [Google Scholar]
- Song, J.; Northcutt, R.G. Morphology, Distribution and Innervation of the Lateral-Line Receptors of the Florida Gar, Lepiosteus platyrhinchus. Brain. Behav. Evol. 1991, 37, 10–37. [Google Scholar] [CrossRef]
- Bemis, W.E.; Grande, L. Early Development of the Actinopterygian Head. I. External Development and Staging of the Paddlefish Polyodon spathula. J. Morphol. 1992, 213, 47–83. [Google Scholar] [CrossRef] [PubMed]
- Brooks, E.R.; Wallingford, J.B. Multiciliated Cells. Curr. Biol. 2014, 24, R973–R982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klos Dehring, D.A.; Vladar, E.K.; Werner, M.E.; Mitchell, J.W.; Hwang, P.; Mitchell, B.J. Deuterosome-Mediated Centriole Biogenesis. Dev. Cell 2013, 27, 103–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whiting, H.P.; Bone, Q. Ciliary Cells in the Epidermis of the Larval Australian Dipnoan, Neoceratodus. Zool. J. Linn. Soc. 1980, 68, 125–137. [Google Scholar] [CrossRef]
- Lauder, G.V.; Liem, K.F. The Evolution and Interrelationships of the Actinopterygian Fishes. Bull. Mus. Comp. Zool. Harv. Coll. 1983, 150, 95–197. [Google Scholar]
- Balfour, F.M. Chapter VII—Amphibia. In A Treatise on Comparative Embryology; MacMillan: London, UK, 1881; pp. 99–119. [Google Scholar]
- Walentek, P.; Quigley, I.K. What We Can Learn from a Tadpole about Ciliopathies and Airway Diseases: Using Systems Biology in Xenopus to Study Cilia and Mucociliary Epithelia. Genesis 2017, 55, e23001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuyen, A.; Rulquin, C.; Daian, F.; Thomé, V.; Clément, R.; Kodjabachian, L.; Pasini, A. The Scf/Kit Pathway Implements Self-Organized Epithelial Patterning. Dev. Cell 2021, 56, 795–810.e7. [Google Scholar] [CrossRef] [PubMed]
- Marcet, B.; Chevalier, B.; Luxardi, G.; Coraux, C.; Zaragosi, L.-E.; Cibois, M.; Robbe-Sermesant, K.; Jolly, T.; Cardinaud, B.; Moreilhon, C.; et al. Control of Vertebrate Multiciliogenesis by MiR-449 through Direct Repression of the Delta/Notch Pathway. Nat. Cell Biol. 2011, 13, 693–699. [Google Scholar] [CrossRef]
- Stubbs, J.L.; Vladar, E.K.; Axelrod, J.D.; Kintner, C. Multicilin Promotes Centriole Assembly and Ciliogenesis during Multiciliate Cell Differentiation. Nat. Cell Biol. 2012, 14, 140–147. [Google Scholar] [CrossRef] [Green Version]
- Dubaissi, E.; Rousseau, K.; Lea, R.; Soto, X.; Nardeosingh, S.; Schweickert, A.; Amaya, E.; Thornton, D.J.; Papalopulu, N. A Secretory Cell Type Develops alongside Multiciliated Cells, Ionocytes and Goblet Cells, and Provides a Protective, Anti-Infective Function in the Frog Embryonic Mucociliary Epidermis. Dev. Camb. Engl. 2014, 141, 1514–1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elgeti, J.; Gompper, G. Emergence of Metachronal Waves in Cilia Arrays. Proc. Natl. Acad. Sci. USA 2013, 110, 4470–4475. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, B.; Stubbs, J.L.; Huisman, F.; Taborek, P.; Yu, C.; Kintner, C. The PCP Pathway Instructs the Planar Orientation of Ciliated Cells in the Xenopus Larval Skin. Curr. Biol. 2009, 19, 924–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assheton, M.A. Notes on the Ciliation of the Ectoderm of the Amphibian Embryo. Q. J. Microsc. Sci. 1896, 38, 465–484. [Google Scholar]
- Burggren, W.W. Gas, Exchange, Metabolism, and “Ventilation”; in Gelatinous Frog Egg Masses. Physiol. Zool. 1985, 58, 503–514. [Google Scholar] [CrossRef]
- Feder, M.E.; Burggren, W.W. Cutaneous Gas Exchange in Vertebrates: Design, Patterns, Control and Implications. Biol. Rev. 1985, 60, 1–45. [Google Scholar] [CrossRef] [PubMed]
- Seymour, R.S.; Bradford, D.F. Gas Exchange through the Jelly Capsule of the Terrestrial Eggs of the Frog, Pseudophryne bibroni. J. Comp. Physiol. B 1987, 157, 477–481. [Google Scholar] [CrossRef]
- Twitty, V.C. Experimental Studies on the Ciliary Action of Amphibian Embryos. J. Exp. Zool. 1928, 50, 319–344. [Google Scholar] [CrossRef]
- Cartwright, J.H.E.; Piro, O.; Tuval, I. Fluid Dynamics in Developmental Biology: Moving Fluids That Shape Ontogeny. HFSP J. 2009, 3, 77–93. [Google Scholar] [CrossRef] [Green Version]
- Ciuhandu, C.S.; Wright, P.A.; Goldberg, J.I.; Stevens, E.D. Parameters Influencing the Dissolved Oxygen in the Boundary Layer of Rainbow Trout (Oncorhynchus mykiss) Embryos and Larvae. J. Exp. Biol. 2007, 210, 1435–1445. [Google Scholar] [CrossRef] [Green Version]
- Liem, K. Larvae of Air-Breathing Fishes as Countercurrent Flow Devices in Hypoxic Environments. Science 1981, 211, 1177–1179. [Google Scholar] [CrossRef] [PubMed]
- Lanzing, W.J.R. A Temporary Respiratory Organ in the Tail of Tilapia mossambica Fry. Copeia 1976, 1976, 800–802. [Google Scholar] [CrossRef]
- Secchi, E.; Vitale, A.; Miño, G.L.; Kantsler, V.; Eberl, L.; Rusconi, R.; Stocker, R. The Effect of Flow on Swimming Bacteria Controls the Initial Colonization of Curved Surfaces. Nat. Commun. 2020, 11, 2851. [Google Scholar] [CrossRef] [PubMed]
- Thurber, A.R.; Jones, W.J.; Schnabel, K. Dancing for Food in the Deep Sea: Bacterial Farming by a New Species of Yeti Crab. PLoS ONE 2011, 6, e26243. [Google Scholar] [CrossRef]
- Nawroth, J.C.; van der Does, A.M.; Ryan (Firth), A.; Kanso, E. Multiscale Mechanics of Mucociliary Clearance in the Lung. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190160. [Google Scholar] [CrossRef]
- Erb-Downward, J.R.; Thompson, D.L.; Han, M.K.; Freeman, C.M.; McCloskey, L.; Schmidt, L.A.; Young, V.B.; Toews, G.B.; Curtis, J.L.; Sundaram, B.; et al. Analysis of the Lung Microbiome in the “Healthy” Smoker and in COPD. PLoS ONE 2011, 6, e16384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawroth, J.C.; Guo, H.; Koch, E.; Heath-Heckman, E.A.C.; Hermanson, J.C.; Ruby, E.G.; Dabiri, J.O.; Kanso, E.; McFall-Ngai, M. Motile Cilia Create Fluid-Mechanical Microhabitats for the Active Recruitment of the Host Microbiome. Proc. Natl. Acad. Sci. USA 2017, 114, 9510–9516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murillo-Rincon, A.P.; Klimovich, A.; Pemöller, E.; Taubenheim, J.; Mortzfeld, B.; Augustin, R.; Bosch, T.C.G. Spontaneous Body Contractions Are Modulated by the Microbiome of Hydra. Sci. Rep. 2017, 7, 15937. [Google Scholar] [CrossRef]
- Gilbert, S.F.; Bosch, T.C.G.; Ledón-Rettig, C. Eco-Evo-Devo: Developmental Symbiosis and Developmental Plasticity as Evolutionary Agents. Nat. Rev. Genet. 2015, 16, 611–622. [Google Scholar] [CrossRef]
- Pennisi, E. Mysteries of Development. How Do Microbes Shape Animal Development? Science 2013, 340, 1159–1160. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kerney, R. Developing Inside a Layer of Germs—A Potential Role for Multiciliated Surface Cells in Vertebrate Embryos. Diversity 2021, 13, 527. https://doi.org/10.3390/d13110527
Kerney R. Developing Inside a Layer of Germs—A Potential Role for Multiciliated Surface Cells in Vertebrate Embryos. Diversity. 2021; 13(11):527. https://doi.org/10.3390/d13110527
Chicago/Turabian StyleKerney, Ryan. 2021. "Developing Inside a Layer of Germs—A Potential Role for Multiciliated Surface Cells in Vertebrate Embryos" Diversity 13, no. 11: 527. https://doi.org/10.3390/d13110527
APA StyleKerney, R. (2021). Developing Inside a Layer of Germs—A Potential Role for Multiciliated Surface Cells in Vertebrate Embryos. Diversity, 13(11), 527. https://doi.org/10.3390/d13110527