
Notes on Some Interesting Sporocarp-Inhabiting Fungi Isolated from Xylarialean Fungi in Japan

Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Sampling of Ascomata
2.2. Isolation of SCIF
2.3. DNA Extraction, Polymerase Chain Reaction (PCR) Amplification, and DNA Sequencing
2.4. Morphological Observation
2.5. Phylogenetic Analyses
3. Results
3.1. Field Sampling and Isolation
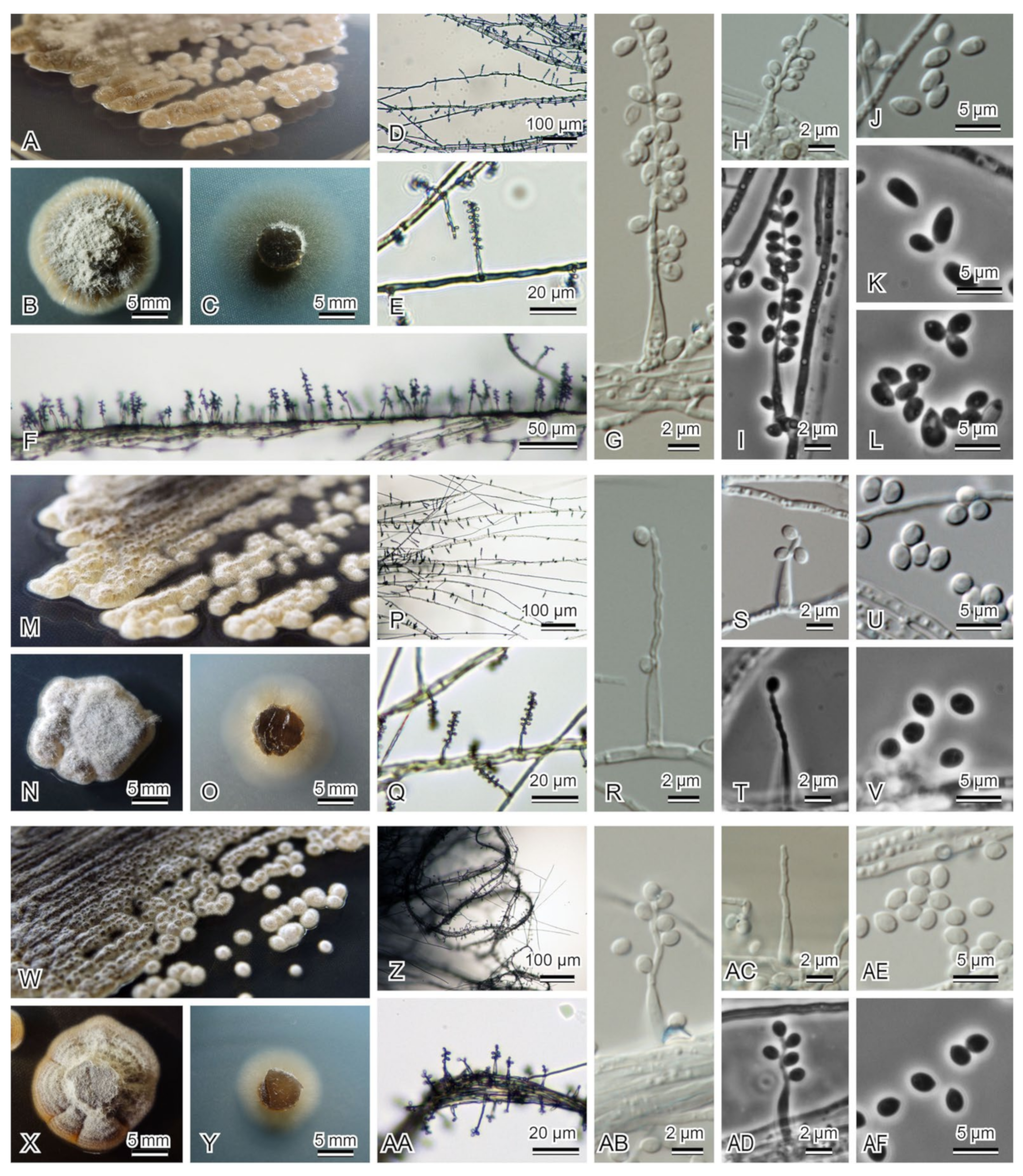
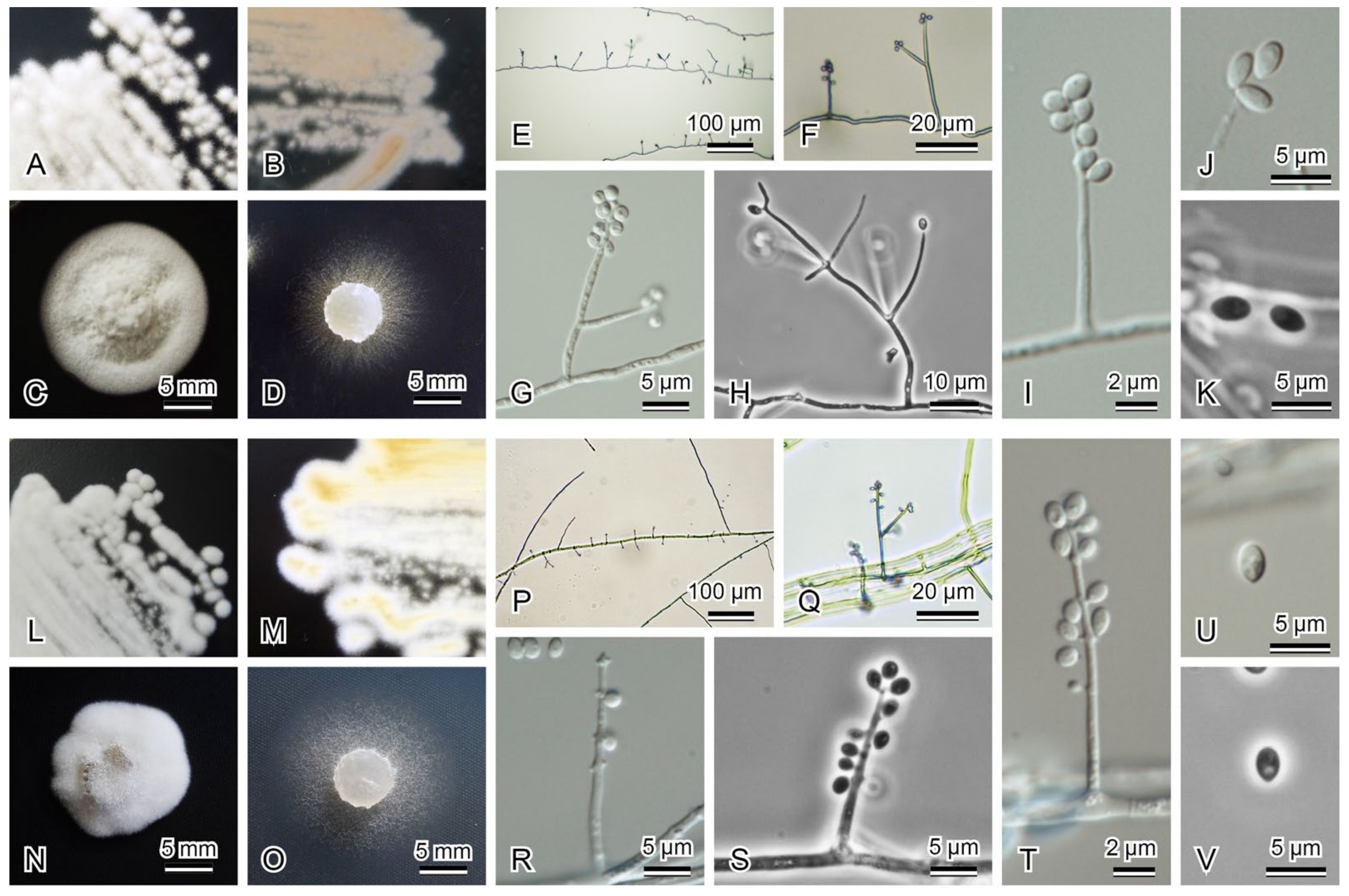
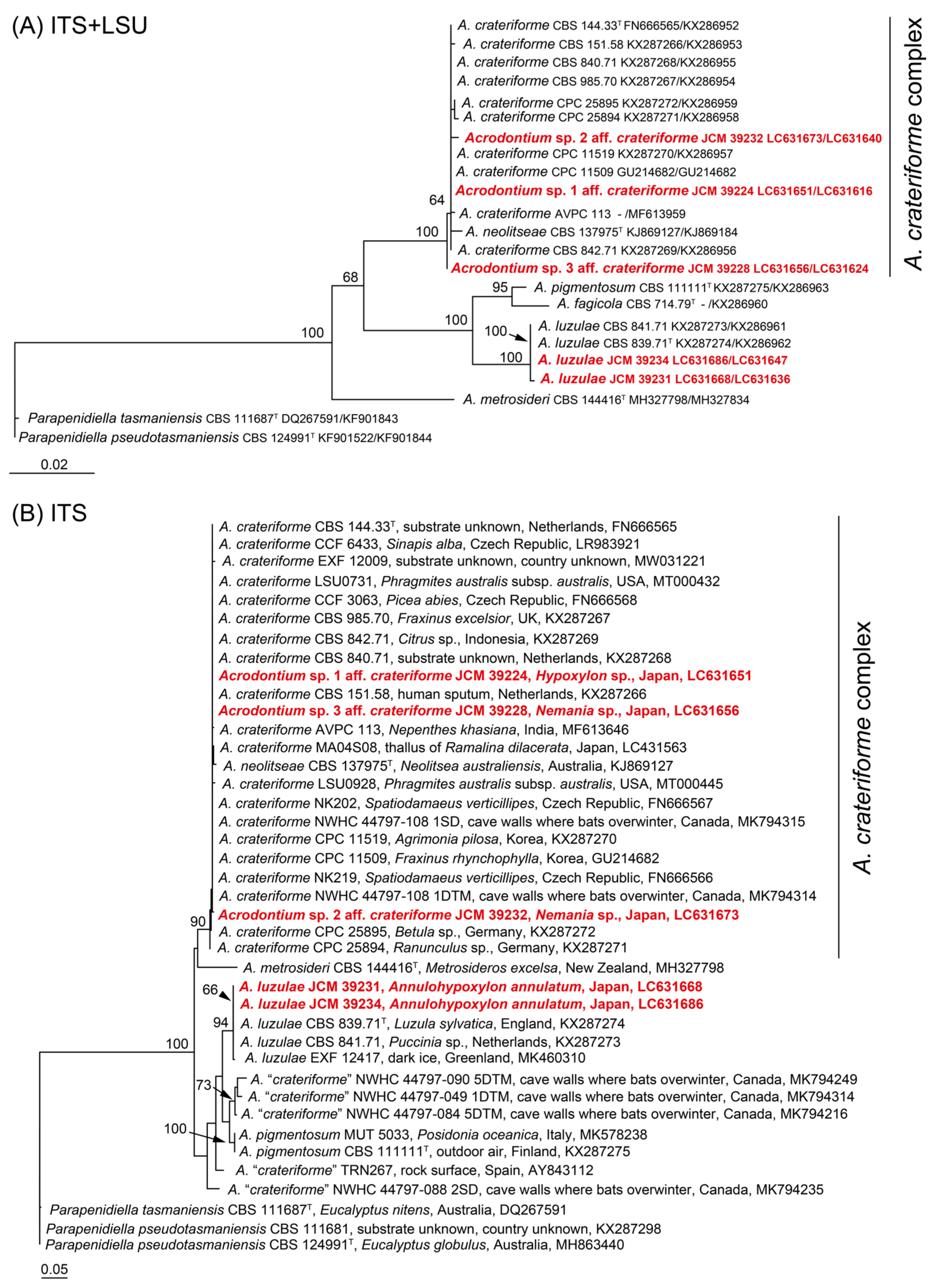
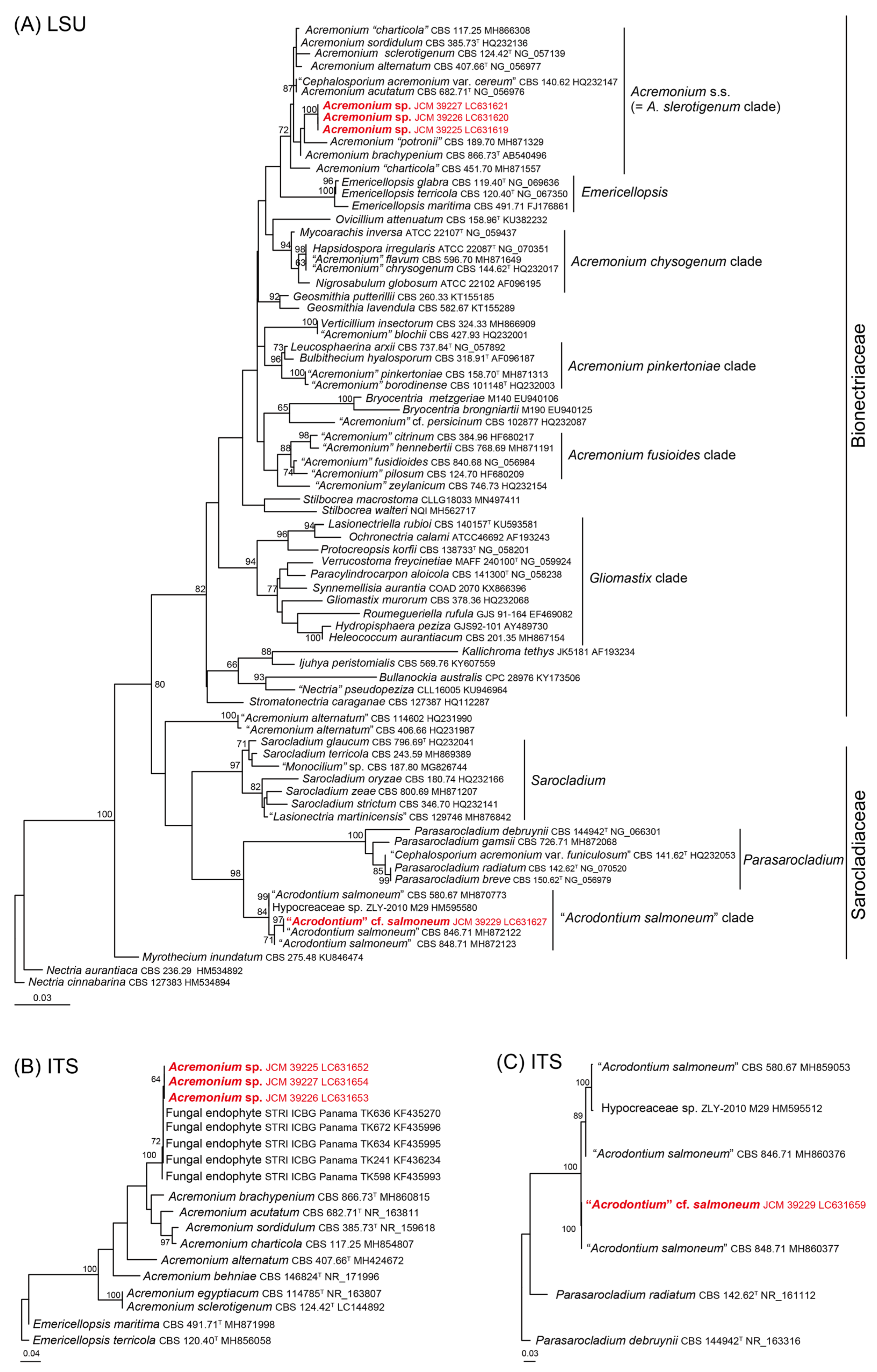
3.2. Morphological Observation and Molecular Phylogenetic Analyses of Noteworthy Fungal Species
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hawksworth, D.L.; Lücking, R. Fungal diversity revisited: 2.2 to 3.8 million species. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef]
- Grum-Grzhimaylo, A.A.; Georgieva, M.L.; Bondarenko, S.A.; Debets, A.J.M.; Bilanenko, E.N. On the diversity of fungi from soda soils. Fungal Divers. 2016, 76, 27–74. [Google Scholar] [CrossRef] [Green Version]
- Le Calvez, T.; Burgaud, G.; Mahé, S.; Barbier, G.; Vandenkoornhuyse, P. Fungal diversity in deep-sea hydrothermal ecosystems. Appl. Environ. Microbiol. 2020, 75, 6415–6421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sklenář, F.; Jurjević, Z.; Zalar, P.; Frisvad, J.C.; Visagie, C.M.; Kolařík, M.; Houbraken, J.; Chen, A.J.; Yilmaz, N.; Seifert, K.A.; et al. Phylogeny of xerophilic aspergilli (subgenus Aspergillus) and taxonomic revision of section Restricti. Stud. Mycol. 2017, 88, 161–236. [Google Scholar] [CrossRef] [Green Version]
- Zalar, P.; de Hoog, G.S.; Schroers, H.J.; Frank, J.M.; Gunde-Cimerman, N. Taxonomy and phylogeny of the xerophilic genus Wallemia (Wallemiomycetes and Wallemiales, cl. et ord. nov.). Antonie van Leeuwenhoek 2005, 87, 311–328. [Google Scholar] [CrossRef]
- Morgenstern, I.; Powlowski, J.; Ishmael, N.; Darmond, C.; Marqueteau, S.; Moisan, M.C.; Quenneville, G.; Tsang, A. A molecular phylogeny of thermophilic fungi. Fungal Biol. 2012, 116, 489–502. [Google Scholar] [CrossRef]
- Sun, J.Z.; Liu, X.Z.; McKenzie, E.H.C.; Jeewon, R.; Liu, J.K.; Zhang, X.L.; Zhao, Q.; Hyde, K.D. Fungicolous fungi: Terminology, diversity, distribution, evolution, and species checklist. Fungal Divers. 2019, 95, 337–430. [Google Scholar] [CrossRef]
- Gams, W.; Diederich, P.; Põldmaa, K. Fungicolous fungi. In Biodiversity of Fungi; Muller, G.M., Bills, G.F., Foster, M.S., Eds.; Academic Press: Amsterdam, The Netherlands, 2004; pp. 343–392. [Google Scholar] [CrossRef]
- Jeffries, P. Biology and ecology of mycoparasitism. Can. J. Bot. 1995, 73, 1284–1290. [Google Scholar] [CrossRef]
- Gilman, J.C.; Tiffany, L.H. Fungicolous Fungi from Iowa. Proc. Iowa Acad. Sci. 1952, 59, 99–110. [Google Scholar]
- Seeler, E.V., Jr. Several fungicolous fungi. Farlowia 1943, 1, 119–133. [Google Scholar] [CrossRef]
- Gams, W. Cephalosporium-like hyphomycetes: Some tropical species. Trans. Br. Mycol. Soc. 1975, 64, 389–404. [Google Scholar] [CrossRef]
- Gams, W.; Zare, R. A revision of Verticillium sect. Prostrata. III. Generic classification. Nova Hedwigia 2001, 72, 329–337. [Google Scholar] [CrossRef]
- Zare, R.; Gams, W.; Culham, A. A revision of Verticillium sect. Prostrata, I. Phylogenetic studies using ITS sequences. Nova Hedwigia 2000, 71, 465–480. [Google Scholar] [CrossRef]
- Zare, R.; Gams, W. A revision of Verticillium section Prostrata. IV. The genera Lecanicillium and Simplicillium gen. nov. Nova Hedwigia 2001, 71, 1–50. [Google Scholar] [CrossRef]
- Tubaki, K. Studies on the Japanese hyphomycetes (II) fungicolous group. Nagaoa 1955, 5, 11–40. [Google Scholar]
- Porter, C.L. Concerning the characteristics of certain fungi as exhibited by their growth in the presence of other fungi. Am. J. Bot. 1924, 11, 168–188. [Google Scholar] [CrossRef]
- Waksman, S.A. Associative and antagonistic effects of microorganisms. I. Historical review of antagonistic relationships. Soil Sci. 1936, 43, 51–68. [Google Scholar] [CrossRef]
- Hoch, H.C.; Provvidenti, R. Mycoparasitic relationship: Cytology of the Sphaerotheca fuligena-Tilletiospis sp. interaction. Phytopathology 1979, 69, 359–362. [Google Scholar] [CrossRef]
- Santos, A.F.D.; Dhingra, O.D. Pathogenicity of Trichoderma spp. on the sclerotia of Sclerotinia sclerotiorum. Can. J. Bot. 1982, 60, 472–475. [Google Scholar] [CrossRef]
- Marletto, F. The micoflora of rhizosphere in truffle culture sites I. Truffle and rhizosphere yeast in truffle culture sites. Allionia 1969, 15, 155–171. [Google Scholar]
- Buzzini, P.; Gasparetti, C.; Turchetti, B.; Cramarossa, M.R.; Vaughan-Martini, A.; Martini, A.; Pagnoni, U.M.; Forti, L. Production of volatile organic compounds (VOCs) by yeasts isolated from the ascocarps of black (Tuber melanosporum Vitt.) and white (Tuber magnatum Pico) truffles. Arch. Microbiol. 2005, 184, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Pacioni, G.; Leonardi, M. Truffle-inhabiting fungi. Soil Biol. 2016, 47, 283–299. [Google Scholar] [CrossRef]
- Perlińska-Lenart, U.; Piłsyk, S.; Gryz, E.; Turło, J.; Hilszczańska, D.; Kruszewska, J.S. Identification of bacteria and fungi inhabiting fruiting bodies of Burgundy truffle (Tuber aestivum Vittad.). Arch. Microbiol. 2020, 202, 2727–2738. [Google Scholar] [CrossRef] [PubMed]
- Danon, M.; Chen, Y.; Hadar, Y. Ascomycete communities associated with suppression of Sclerotium rolfsii in compost. Fungal Ecol. 2010, 3, 20–30. [Google Scholar] [CrossRef]
- Jones, E.E.; Stewart, A. Selection of mycoparasites of sclerotia of Sclerotinia sclerotiorum isolated from New Zealand soils. N. Z. J. Crop Hortic. Sci. 2000, 28, 105–114. [Google Scholar] [CrossRef]
- Tu, J.C. Gliocladium virens, a destructive mycoparasite of Sclerotinia sclerotium. Am. Phytopathol. Soc. 1980, 70, 670–674. [Google Scholar] [CrossRef]
- Whipps, J.M.; Budge, S.P. Screening for sclerotial mycoparasites of Sclerotinia sclerotiorum. Mycol. Res. 1990, 94, 607–612. [Google Scholar] [CrossRef]
- Obase, K.; Douhan, G.W.; Matsuda, Y.; Smith, M.E. Culturable fungal assemblages growing within Cenococcum sclerotia in forest soils. FEMS Microbiol. Ecol. 2014, 90, 708–717. [Google Scholar] [CrossRef] [Green Version]
- Petrini, L.; Petrini, O. Xylariaceous fungi as endophytes. Sydowia 1985, 38, 216–234. [Google Scholar]
- Ellis, M.B.; Ellis, J.P. Microfungi on Miscellaneous Substrates; (Enlarged edition); The Richmond Publishing: Slough, UK, 1998. [Google Scholar]
- Læssoe, T.; Peterson, J.H. Fungi of Temperate Europe; Princeton University Press: Princeton, NJ, USA, 2019. [Google Scholar]
- Becker, K.; Lambert, C.; Wieschhus, J.; Stadler, M. Phylogenetic assignment of the fungicolous Hypoxylon invadens (Ascomycota, Xylariales) and investigation of its secondary metabolites. Microorganisms 2020, 8, 1397. [Google Scholar] [CrossRef]
- Nordén, B.; Læssøe, T.; Jordal, J.B. Chlorostroma vestlandicum sp. nov., a host-specific mycoparasite on Hypoxylon vogesiacum from western Norway. Karstenia 2014, 54, 9–13. [Google Scholar] [CrossRef] [Green Version]
- Vasilyeva, L.N.; Rogers, J.D.; Miller, A.N. Pyrenomycetes of the Great Smoky Mountains National Park. V. Annulohypoxylon and Hypoxylon (Xylariaceae). Fungal Divers. 2007, 27, 231–245. [Google Scholar]
- Izumitsu, K.; Hatoh, K.; Sumita, T.; Kitade, Y.; Morita, A.; Tanaka, C.; Gafur, A.; Ohta, A.; Kawai, M.; Yamanaka, T.; et al. Rapid and simple preparation of mushroom DNA directly from colonies and fruiting bodies for PCR. Mycoscience 2012, 53, 396–401. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., Thomas, J.W., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from Several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- De Hoog, G.S.; Gerrits van den Ende, A.H.G. Molecular diagnostics of clinical strains of filamentous Basidiomycetes. Mycoses 1998, 41, 183–189. [Google Scholar] [CrossRef]
- Gams, W.; van der Aa, H.A.; van der Plaats-Niterink, A.J.; Samson, R.A.; Stalpers, J.A. CBS Course of Mycology; Centraalbureau voor Shimmelcultures: Utrecht, The Netherlands, 1987. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Bensch, K.; Braun, U.; Groenewald, J.Z.; Crous, P.W. The genus Cladosporium. Stud. Mycol. 2012, 72, 1–401. [Google Scholar] [CrossRef] [Green Version]
- Moncalvo, J.M.; Nilsson, R.H.; Koster, B.; Dunham, S.M.; Bernauer, T.; Matheny, P.B.; Porter, T.M.; Margaritescu, S.; Weiß, M.; Garnica, S.; et al. The cantharelloid clade: Dealing with incongruent gene trees and phylogenetic reconstruction methods. Mycologia 2006, 98, 937–948. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. Corrigendum to: IQ-TREE 2: New models and efficient methods for phylogenetic inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Videira, S.I.R.; Groenewald, J.Z.; Braun, U.; Shin, H.D.; Crous, P.W. All that glitters is not Ramularia. Stud. Mycol. 2016, 83, 49–163. [Google Scholar] [CrossRef] [Green Version]
- De Hoog, G.S. The genera Beauveria, Isaria, Tritirachium, and Acrodontium gen. nov. Stud. Mycol. 1972, 1, 1–41. [Google Scholar]
- Perini, L.; Gostinčar, C.; Anesio, A.M.; Williamson, C.; Tranter, M.; Gunde-Cimerman, N. Darkening of the Greenland ice sheet: Fungal abundance and diversity are associated with algal bloom. Front. Microbiol. 2019, 10, 557. [Google Scholar] [CrossRef] [Green Version]
- Koukol, O. What do we know about Acrodontium crateriforme? Mykologické Listy 2010, 111, 12–18. [Google Scholar]
- Glenn, A.E.; Bacon, C.W.; Price, R.; Hanlin, R.T. Molecular phylogeny of Acremonium and its taxonomic implications. Mycologia 1996, 88, 369–383. [Google Scholar] [CrossRef] [Green Version]
- Lucheta, A.R.; de Souza Cannavan, F.; Roesch, L.F.; Tsai, S.M.; Kuramae, E.E. Fungal community assembly in the Amazonian dark earth. Microb. Ecol. 2016, 71, 962–973. [Google Scholar] [CrossRef] [Green Version]
- Manandhar, J.B.; Thapliyal, P.N.; Cavanaugh, K.J.; Sinclair, J.B. Interaction between pathogenic and saprobic fungi isolated from soybean roots and seeds. Mycopathologia 1987, 98, 69–75. [Google Scholar] [CrossRef]
- He, P.; He, X.; Zhang, C. Interactions between Psilocybe fasciata and its companion fungus Acremonium strictum. Ecol. Res. 2006, 21, 387–395. [Google Scholar] [CrossRef]
- Pacioni, G.; Leonardi, M.; Aimola, P.; Ragnelli, A.M.; Rubini, A.; Paolocci, F. Isolation and characterization of some mycelia inhabiting Tuber ascomata. Mycol. Res. 2007, 111, 1450–1460. [Google Scholar] [CrossRef]
- Summerbell, R.C.; Gueidan, C.; Schroers, H.J.; de Hoog, G.S.; Starink, M.; Rosete, Y.A.; Guarro, J.; Scott, J.A. Acremonium phylogenetic overview and revision of Gliomastix, Sarocladium, and Trichothecium. Stud. Mycol. 2011, 68, 139–162. [Google Scholar] [CrossRef] [PubMed]
- Kozlova, E.V.; Mazina, S.E. Biodiversity of Fungi in the photic and aphotic zones of Montenegro caves. Aerobiologia 2020, 36, 589–604. [Google Scholar] [CrossRef]
- Steiman, R.; Benoit-Guyod, J.L.; Guiraud, P.; Seigle-Murandi, F. Evaluation of acrodontiolamide, a chlorinated compound produced by Acrodontium salmoneum de Hoog for cytotoxicity and antimicrobial activity. Pharmazie 1995, 50, 693–695. [Google Scholar] [CrossRef]
- Stejskal, V.; Hubert, J.; Kubátová, A. Associated-food-hazards: Storage fungi and mites in poppy, mustard, lettuce and wheat. Plant Prot. Sci. 2017, 38, 673–680. [Google Scholar] [CrossRef]
- Yuan, Z.L.; Rao, L.B.; Chen, Y.C.; Zhang, C.L.; Wu, Y.G. From pattern to process: Species and functional diversity in fungal endophytes of Abies beshanzuensis. Fungal Biol. 2011, 115, 197–213. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, K.; Kaifuchi, S.; Ōmura, S.; Masuma, R. Five new Simplicillium species (Cordycipitaceae) from soils in Tokyo, Japan. Mycoscience 2013, 54, 42–53. [Google Scholar] [CrossRef]
- Serradilla, M.J.; Villalobos, M.D.; Hernández, A.; Martín, A.; Lozano, M.; Córdoba, M.D. Study of microbiological quality of controlled atmosphere packaged ‘Ambrunés’ sweet cherries and subsequent shelf-life. Int. J. Food Microbiol. 2013, 166, 85–92. [Google Scholar] [CrossRef]
- Jaber, S.; Mercier, A.; Knio, K.; Brun, S.; Kambris, Z. Isolation of fungi from dead arthropods and identification of a new mosquito natural pathogen. Parasites Vectors 2016, 9, 491. [Google Scholar] [CrossRef] [Green Version]
- Maurice, S.; Arnault, G.; Nordén, J.; Botnen, S.S.; Miettinen, O.; Kauserud, H. Fungal sporocarps house diverse and host-specific communities of fungicolous fungi. ISME J. 2021, 15, 1445–1457. [Google Scholar] [CrossRef]
- Dennis, R.W.G. British Ascomycetes; J. Cramer: Vaduz, Liechtenstein, 1981. [Google Scholar]
- Hattori, T. Diversity of wood-inhabiting polypores in temperate forests with different vegetation types in Japan. Fungal Divers. 2005, 18, 73–88. [Google Scholar]
- Justo, A.; Miettinen, O.; Floudas, D.; Ortiz-Santana, B.; Sjökvist, E.; Lindner, D.; Nakasone, K.; Niemelä, T.; Larsson, K.H.; Ryvardeng, L.; et al. A revised family-level classification of the Polyporales (Basidiomycota). Fungal Biol. 2017, 121, 798–824. [Google Scholar] [CrossRef]
- Greslebin, A.G.; Rajchenberg, M. Diversity of Corticiaceae sens. lat. in Patagonia, Southern Argentina. N. Z. J. Bot. 2003, 41, 437–446. [Google Scholar] [CrossRef]
- Maekawa, N. Taxonomic study of Japanese Corticiaceae (Aphyllophorales) I. Rep. Tottori Mycol. Inst. 1993, 31, 1–149. [Google Scholar] [CrossRef]
- Hao, Y.; Chen, S.Y.; Blanchette, R.A.; Liu, X.Z. Sistotrema brinkmannii, a psychrotolerant fungus from Antarctic soil. Mycosystema 2010, 29, 864–868. [Google Scholar]
- Gräfenhan, T.; Schroers, H.J.; Nirenberg, H.I.; Seifert, K.A. An overview of the taxonomy, phylogeny, and typification of nectriaceous fungi in Cosmospora, Acremonium, Fusarium, Stilbella, and Volutella. Stud. Mycol. 2011, 68, 79–113. [Google Scholar] [CrossRef] [Green Version]
- Samson, R.A.; Visagie, C.M.; Houbraken, J.; Hong, S.B.; Hubka, V.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Susca, A.; Tanney, J.B.; et al. Phylogeny, identification and nomenclature of the genus Aspergillus. Stud. Mycol. 2014, 78, 141–173. [Google Scholar] [CrossRef] [Green Version]
- Collado, J.; Platas, G.; Gonzalez, I.; Pelaez, F. Geographical and seasonal influences on the distribution of fungal endophytes in Quercus ilex. New Phytol. 1999, 144, 525–532. [Google Scholar] [CrossRef]
- De Gusmáo, N.B.; Kaouadji, M.; Seigle-Murandi, F.; Steiman, R.; Thomasson, F. Acrodontiolamide, a chlorinated fungal metabolite from Acrodontium salmoneum. Spectrosc. Lett. 1993, 26, 1373–1380. [Google Scholar] [CrossRef]
- Liang, X.; Nong, X.H.; Huang, Z.H.; Qi, S.H. Antifungal and antiviral cyclic peptides from the deep-sea-derived fungus Simplicillium obclavatum EIODSF 020. J. Agric. Food Chem. 2017, 65, 5114–5121. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample No. | Species | Substrate | Sampling Site |
|---|---|---|---|
| AM002 | Hypoxylon sp. | Cryptomeria japonica | JAPAN, Ibaraki, Sakuragawa, near Ibaraki prefecture road route 41, 36°14′26.195″ N 140°03′57.412″ E |
| AM003 | Hypoxylon sp. | Alnus sp. | JAPAN, Ibaraki, Sakuragawa, near Ibaraki prefecture road route 41, dried river, 36°14′54.103″ N 140°05′58.300″ E |
| AM007 | Nemania sp. | Cryptomeria japonica | JAPAN, Ibaraki, Sakuragawa, around Ibaraki prefecture road route 41, near Onogawa-river, 36°14′13.736″ N 140°06′05.799″ E |
| WS34 | Annulohypoxylon annulatum | Zelkova serrata | JAPAN, Ibaraki, Tsukuba, near Takasaki Nature Park, 36°00′10.4″ N 140°06′49.5″ E |
| WS35 | Nemania sp. | Zelkova serrata | JAPAN, Ibaraki, Tsukuba, near Takasaki Nature Park, 36°00′10.4″ N 140°06′49.5″ E |
| WS36 | Annulohypoxylon annulatum | Zelkova serrata | JAPAN, Ibaraki, Tsukuba, near Takasaki Nature Park, 36°00′10.4″ N 140°06′49.5″ E |
| Sample No. | Isolate No. (JCM No.) | OTU Name | Closest Match from GenBank Using Megablast | GenBank No. | |
|---|---|---|---|---|---|
| ITS | LSU | ||||
| AM002 | 1_1_B_As_1000 | Moesziomyces antarcticus | Moesziomyces antarcticus (MH873351) 822/822 (100.00%) in LSU | LC631649 | LC631614 |
| 1_1_A_St_1000 | Herpotrichiellaceae sp. | “Capronia” pilosella (DQ826737) 535/542 (98.70%) in ITS | LC631650 | LC631615 | |
| 1_2_B_As_1000 (=JCM 39224) | Acrodontium sp. 1 aff. crateriforme | Acrodontium crateriforme (NR_152320) 457/457 (100%) in ITS | LC631651 | LC631616 | |
| 1_2_A_St_1000 | Tilletiopsis washingtonensis | Tilletiopsis washingtonensis (MH868275) 867/869 (99.77%) in LSU | - | LC631617 | |
| 1_8_A_St_1000 | Robbauera albescens | Robbauera albescens (MH873380) 809/810 (99.77%) in LSU | - | LC631618 | |
| AM003 | 1_1_A_As_10000 (=JCM 39225) | Acremonium sp. | Acremonium charticola (MH859034) 554/587 (94.38%) including 9 gaps in ITS | LC631652 | LC631619 |
| 1_1_B_As_1000 (=JCM 39226) | Acremonium sp. | Acremonium charticola (MH859034) 545/578(94.29%) including 9 gaps in ITS | LC631653 | LC631620 | |
| 1_2_A_As_1000 (=JCM 39227) | Acremonium sp. | Acremonium charticola (MH859034) 557/590 (94.40%) including 9 gaps in ITS | LC631654 | LC631621 | |
| AM007 | 2_1_A_As_1000 | Dactylospora sp. | Dactylospora parasitica (KY661666) 766/788(97.20%) including 1 gap in LSU | - | LC631622 |
| 2_3_A_As_1000 | Cladosporium sp. aff. cladosporioides | Cladosporium cladosporioides complex sensu Bensch et al. [42] | LC631655 | LC631623 | |
| 2_4_A_As_1000 (=JCM 39228) | Acrodontium sp. 3 aff. crateriforme | Acrodontium crateriforme (NR_152320) 441/442 (99.77%) in ITS | LC631656 | LC631624 | |
| 2_5_A_As_1000 | Skeletocutis odora | Skeletocutis odora (KY948893) 805/805 (100%) in LSU | LC631657 | LC631625 | |
| WS34 | 1_2_A_As_10000 | Burgella sp. | Burgella flavoparmeliae (KC336075) 708/710 (99.72%) in LSU | LC631658 | LC631626 |
| 2_1_A_As_1000 (=JCM 39229) | Acrodontium cf. salmoneum | Acrodontium salmoneum (MH860376) 563/564 (99.82%) including 1 gaps in ITS | LC631659 | LC631627 | |
| 2_1_A_As_10000 | Penicillium sp. 1 | Penicillium citreosulfuratum (NR_153252) 576/578 (99.65%) in ITS | LC631660 | LC631628 | |
| 2_1_B_As_1000 | Sistotrema sp. aff. brinkmanni | Sistotrema brinkmannii-oblongisporum group sensu Moncalvo et al. [43] | LC631661 | LC631629 | |
| 2_1_B_As_10000 | Acaromyces ingoldii | Acaromyces ingoldii (NG_058540) 445/446 (99%) in LSU | LC631662 | LC631630 | |
| 2_2_A_As_10000 (=JCM 39230) | Simplicillium sp. 1 aff. sympodiophorum | Simplicillium sympodiophorum (NR_111027) 554/555 (99.82%) in ITS | LC631663 | LC631631 | |
| 2_2_A_As_1000 | Neoantrodiella gypsea | Neoantrodiella gypsea (KT203312) 773/774 (99.87% in LSU | - | LC631632 | |
| 2_2_B_As_1000 | Penicillium sp. 2 | Penicillium mallochii (NR_111674) 533/534 (99.81%) including 1 gap in ITS | LC631664 | - | |
| 2_2_B_As_10000 | Phlebia livida | Phlebia livida subsp. tuberculate (MW732462) 781/781 (100%) in LSU | LC631665 | LC631633 | |
| 2_3_A_As_10000 | Lenzites betulinus | Lenzites betulinus (MT644927) 785/785 (100%) in LSU | LC631666 | LC631634 | |
| 2_4_A_As_10000 | Microporus sp. aff. xanthopus | Microporus xanthopus (KX880659) 813/813 (100%) in LSU | LC631667 | LC631635 | |
| 2_5_A_As_10000 (=JCM 39231) | Acrodontium luzulae | Acrodontium luzulae (NR_154720) 526/526 (100%) in ITS | LC631668 | LC631636 | |
| 3_1_A_As_10000 | Robbauera albescens | Robbauera albescens (NR_138401) 618/618 (100%) in ITS | LC631669 | - | |
| WS35 | 1_1_A_As_10000 | Robbauera albescens | Robbauera albescens (MH873380) 488/488 (100%) in LSU | LC631670 | LC631637 |
| 1_2_A_As_10000 | Neoantrodiella gypsea | Neoantrodiella gypsea (KT203312) 750/750 (100%) in LSU | LC631671 | LC631638 | |
| 1_3_A_As_10000 | Incrustoporia chrysella | Incrustoporia chrysella (KP135286) 760/762 (99.74%) in LSU | LC631672 | LC631639 | |
| 1_5_B_As_10000 (=JCM 39232) | Acrodontium sp. 2 aff. crateriforme | Acrodontium neolitseae (NR_168148) 532/536 (99.25%) in ITS | LC631673 | LC631640 | |
| 2_1_B_As_1000 | Penicillium sp. 3 | Penicillium steckii (NR_111488) 373/373 (100%) in ITS | LC631674 | - | |
| 2_1_A_As_10000 | Trametes sp. aff. versicolor | Trametes versicolor (KC176306) 607/611 (99.35%) in ITS | LC631675 | - | |
| 2_2_B_As_10000 | Neoantrodiella gypsea | Neoantrodiella gypsea (KT203312) 670/671 (99.85%) in LSU | LC631676 | LC631641 | |
| 2_3_A_As_10000 | Neoantrodiella gypsea | Neoantrodiella gypsea (KT203291) 519/521 (99.62%) including 1 gap in ITS | LC631677 | - | |
| 2_4_B_As_1000 | Talaromyces sp. | Talaromyces aurantiacus (NR_103681) 464/474 (97.89%) including 6 gaps in ITS | LC631678 | LC631642 | |
| 2_6_B_As_1000 | Beauveria bassiana | Beauveria bassiana (NR_111594) 501/504 (99.40%) including 1 gap in ITS | LC631679 | LC631643 | |
| 3_6_B_As_10000 (=JCM 39233) | Simplicillium sp. 2 aff. sympodiophorum | Simplicillium sympodiophorum (NR_111027) 559/561 (99.64%) including 1 gap in ITS | LC631680 | - | |
| 2_4_B_As_1000 | Talaromyces sp. | Talaromyces aurantiacus (NR_103681) 464/474 (97.89%) including 6 gaps in ITS | LC631678 | LC631642 | |
| 3_8_B_As_10000 | Moesziomyces antarcticus | Moesziomyces antarcticus (NR_155406) 686/689 (99.56%) including 2 gaps in ITS | LC631681 | - | |
| WS36 | 1_1_B_As_10000 | Aspergillus sp. aff. versicolor | Aspergillus versicolor (KU729039) 506/508 (99.60%) including 2 gaps in ITS | LC631682 | - |
| 1_2_B_As_10000 | Cerrena zonata | Cerrena zonata (MW785060) 711/712 (99.86%) including 1 gap in LSU | LC631683 | LC631644 | |
| 1_4_A_As_10000 | Sistotrema sp. | Sistotrema coroniferum (KF218968) 628/646 (97.21%) including 4 gaps in LSU | LC631684 | LC631645 | |
| 2_3_b_As_10000 | Neoantrodiella gypsea | Neoantrodiella gypsea (KT203312) 750/751 (99.87%) including 1 gap in LSU | LC631685 | LC631646 | |
| 3_1_A_As_10000 (=JCM 39234) | Acrodontium luzulae | Acrodontium luzulae (NR_154720) 480/480 (100%) in ITS | LC631686 | LC631647 | |
| 3_3_A_As_1000 | Meira sp. | Meira miltonrushii (NG_060234) 470/472 (99.58%) including 1 gap in LSU | - | LC631648 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sa’diyah, W.; Hashimoto, A.; Okada, G.; Ohkuma, M. Notes on Some Interesting Sporocarp-Inhabiting Fungi Isolated from Xylarialean Fungi in Japan. Diversity 2021, 13, 574. https://doi.org/10.3390/d13110574
Sa’diyah W, Hashimoto A, Okada G, Ohkuma M. Notes on Some Interesting Sporocarp-Inhabiting Fungi Isolated from Xylarialean Fungi in Japan. Diversity. 2021; 13(11):574. https://doi.org/10.3390/d13110574
Chicago/Turabian StyleSa’diyah, Wasiatus, Akira Hashimoto, Gen Okada, and Moriya Ohkuma. 2021. "Notes on Some Interesting Sporocarp-Inhabiting Fungi Isolated from Xylarialean Fungi in Japan" Diversity 13, no. 11: 574. https://doi.org/10.3390/d13110574
APA StyleSa’diyah, W., Hashimoto, A., Okada, G., & Ohkuma, M. (2021). Notes on Some Interesting Sporocarp-Inhabiting Fungi Isolated from Xylarialean Fungi in Japan. Diversity, 13(11), 574. https://doi.org/10.3390/d13110574