
An MPA Design Approach to Benefit Fisheries: Maximising Larval Export and Minimising Redundancy


Abstract
:1. Introduction
2. Materials and Methods
2.1. Complementarity Algorithm Design
2.2. Assumptions
2.3. Measures of Performance
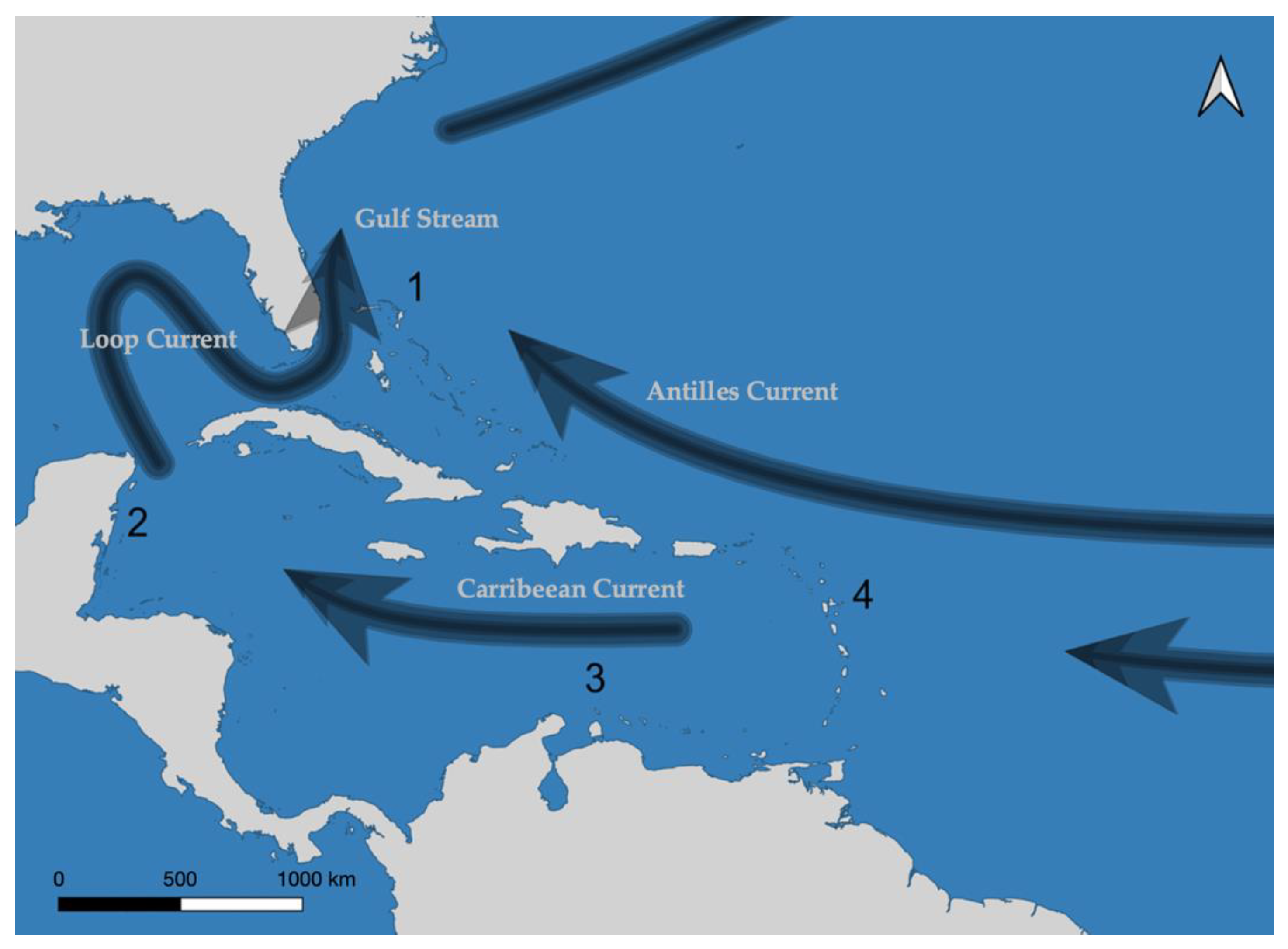
2.4. Application to Caribbean Coral Reef Dataset
3. Results
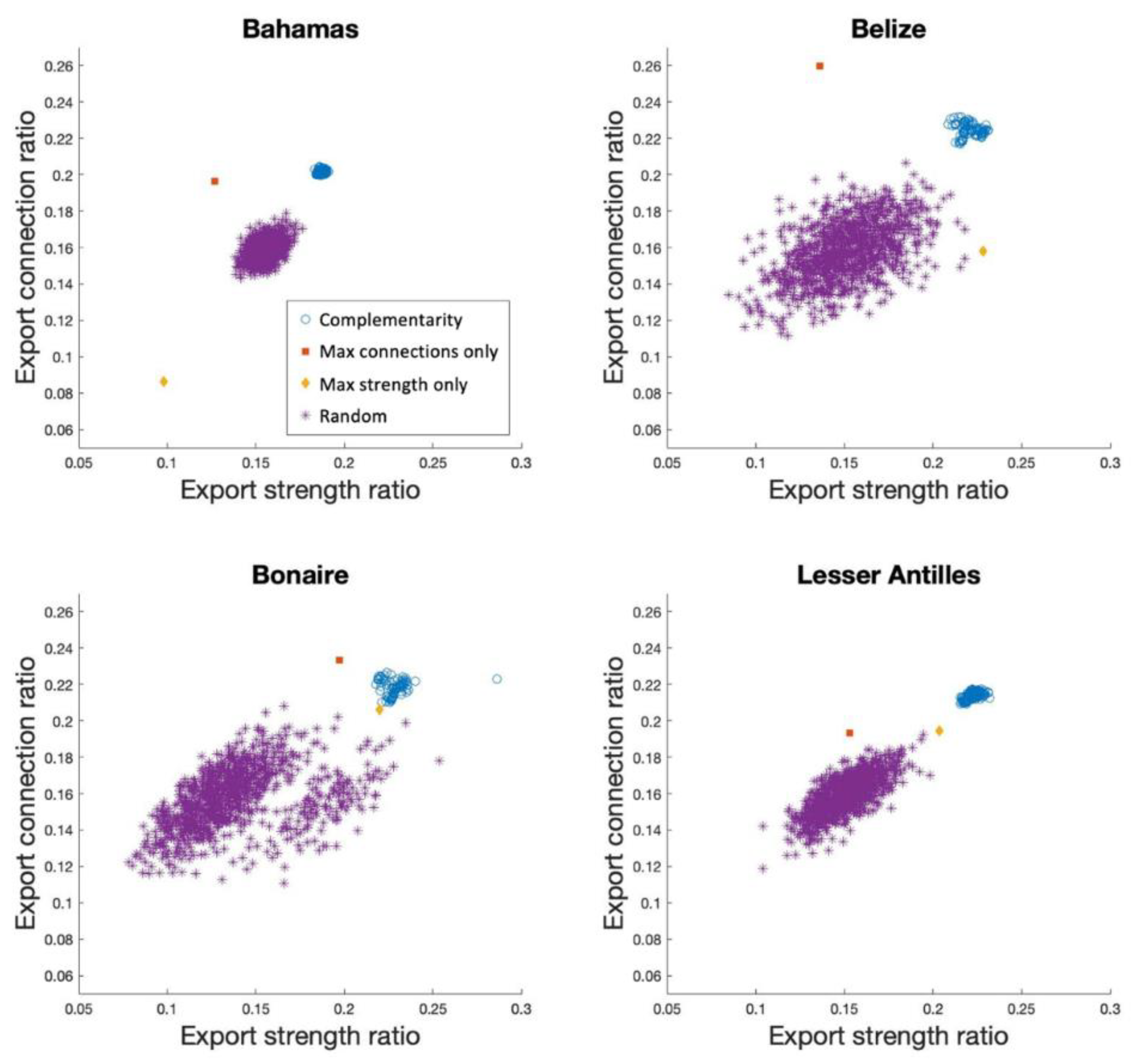
3.1. Connectivity Network Analysis
3.2. Non-Modelling Measures of Performance
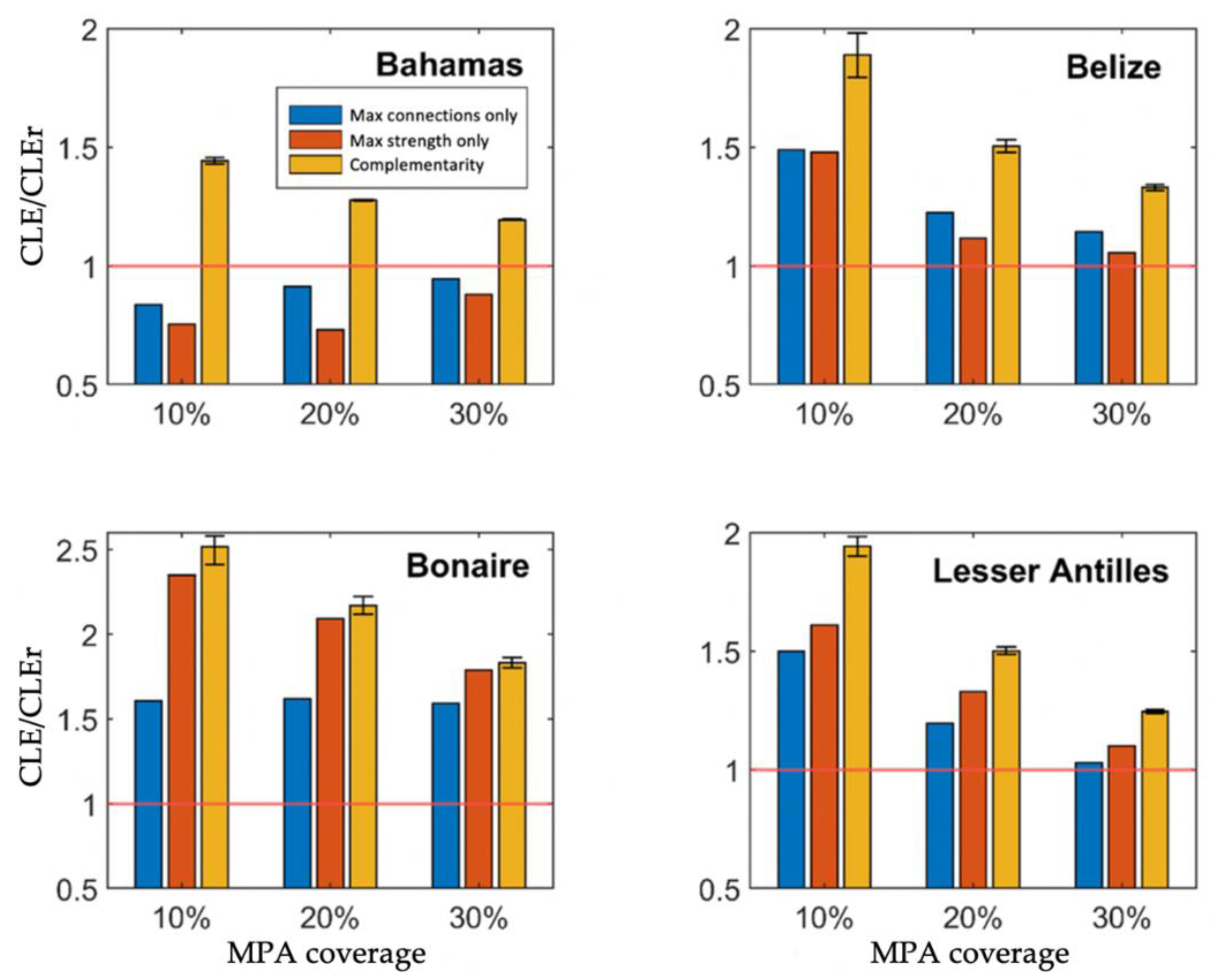
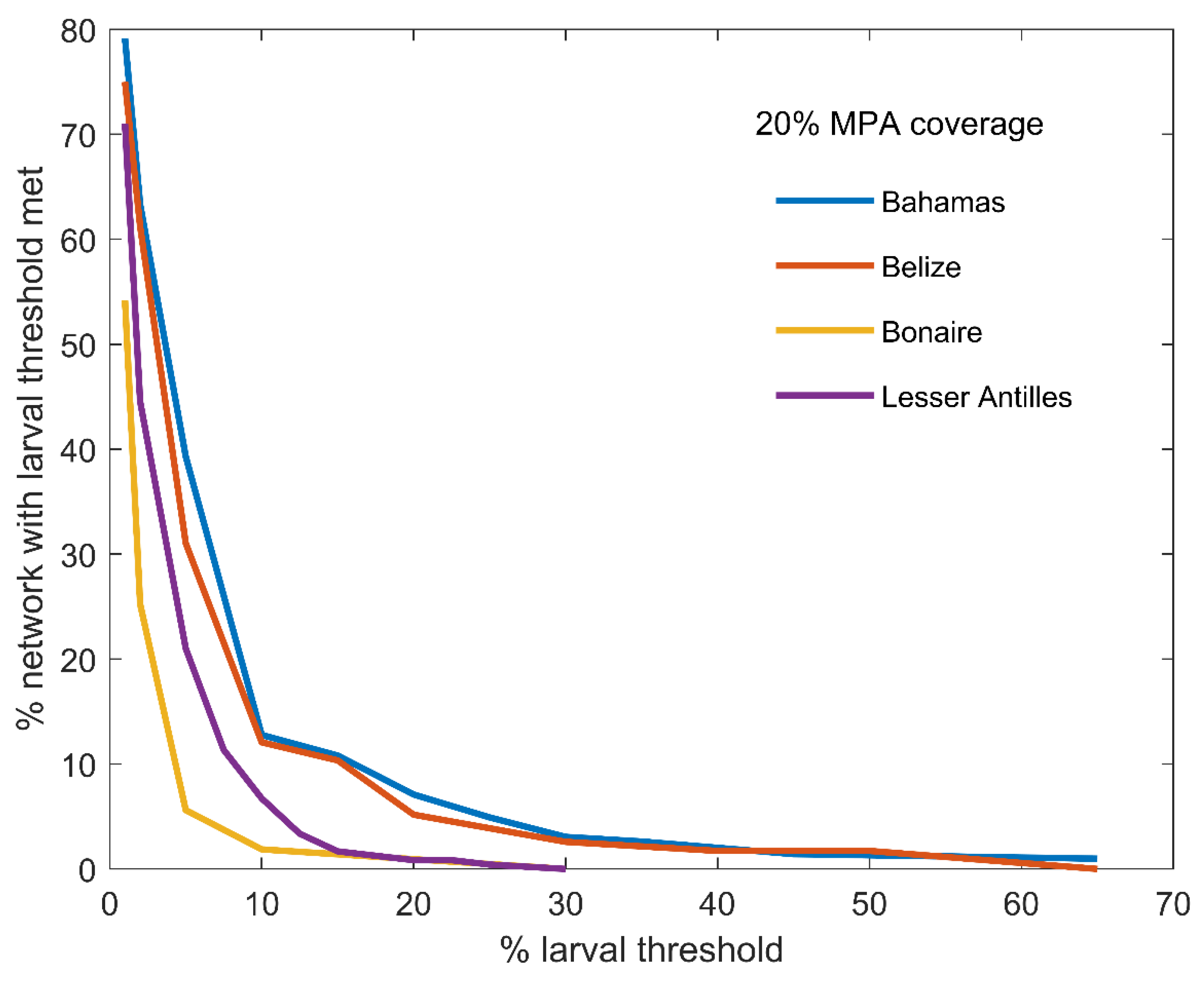
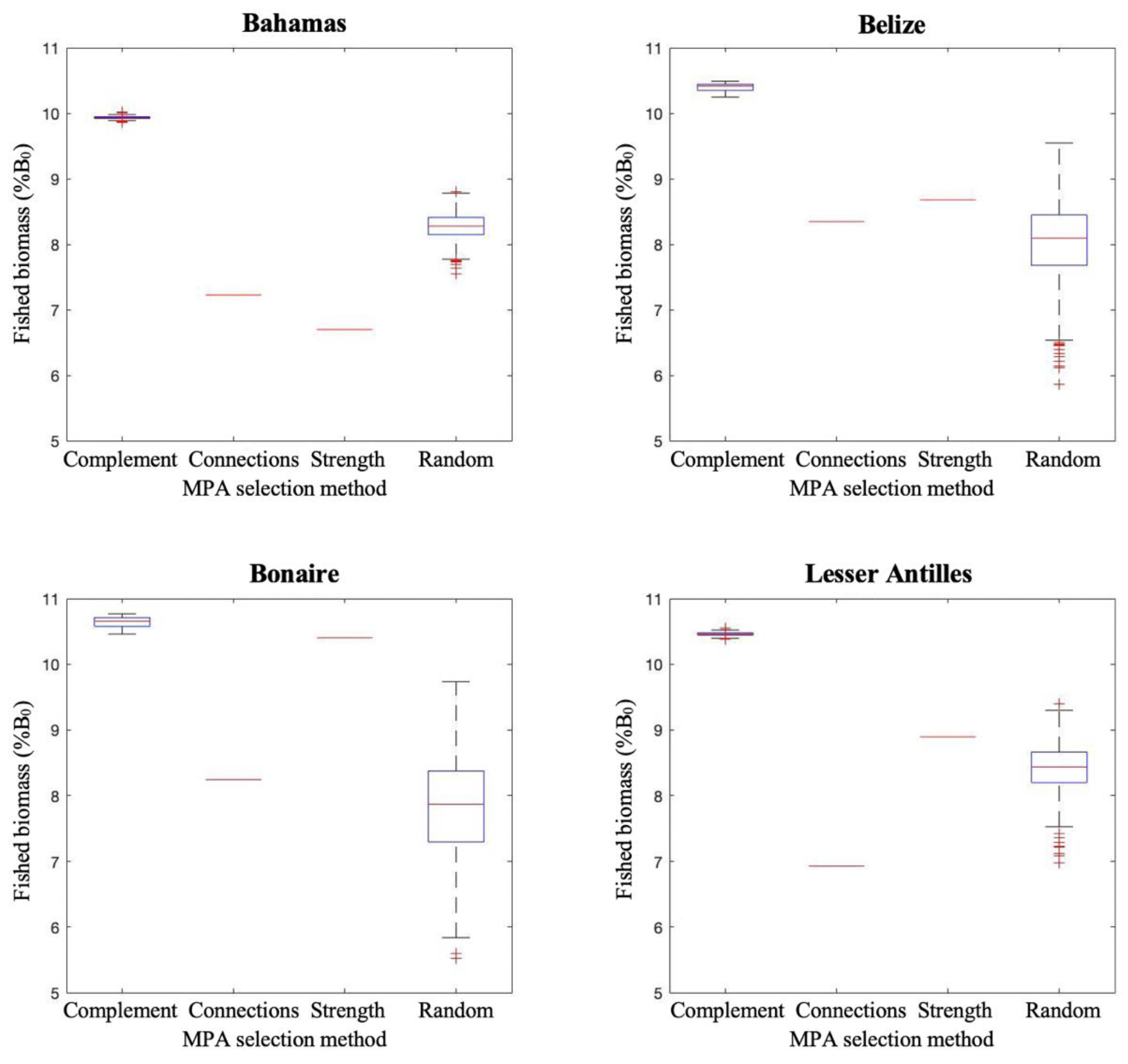
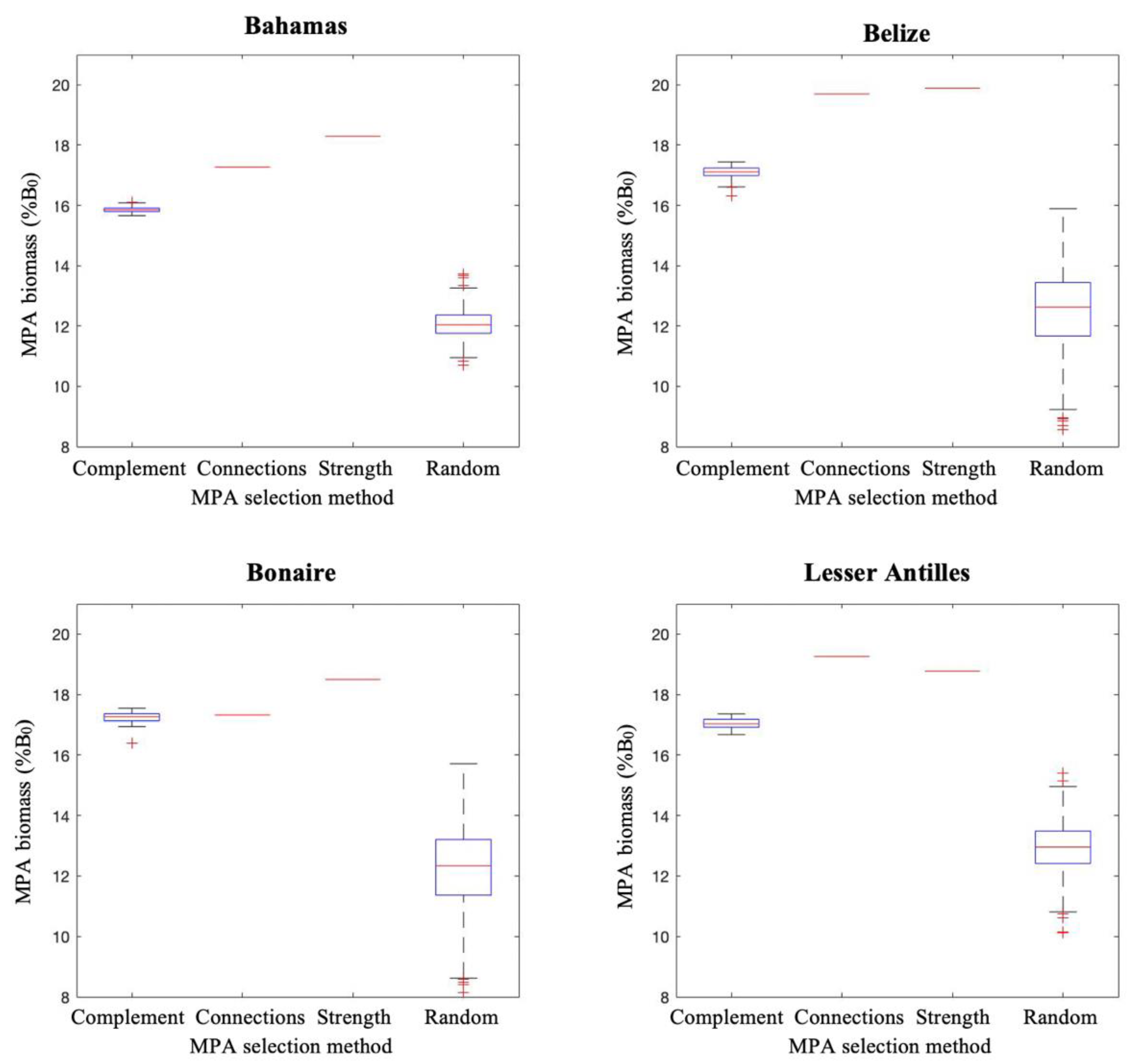
3.3. Modelling Measures of Performance
Population Modelling
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carr, M.H.; Neigel, J.E.; Estes, J.A.; Andelman, S.; Warner, R.R.; Largier, J.L. Comparing marine and terrestrial ecosystems: Implications for the design of coastal marine reserves. Ecol. Appl. 2003, 13 (Suppl. 1), 90–107. [Google Scholar] [CrossRef] [Green Version]
- Brown, C.J.; Harborne, A.R.; Paris, C.B.; Mumby, P.J. Uniting paradigms of connectivity in marine ecology. Ecology 2016, 97, 2447–2457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Righton, D.; Mills, C. Application of GIS to investigate the use of space in coral reef fish: A comparison of territorial behaviour in two red sea butterflyfishes. Int. J. Geogr. Inf. Sci. 2006, 20, 215–232. [Google Scholar] [CrossRef]
- Claisse, J.T.; Clark, T.B.; Schumacher, B.D.; McTee, S.A.; Bushnell, M.E.; Callan, C.K.; Parrish, J.D. Conventional tagging and acoustic telemetry of a small surgeonfish, Zebrasoma flavescens, in a structurally complex coral reef environment. Environ. Biol. Fishes 2011, 91, 185–201. [Google Scholar] [CrossRef]
- Green, A.L.; Maypa, A.P.; Almany, G.R.; Rhodes, K.L.; Weeks, R.; Abesamis, R.A.; White, A.T. Larval dispersal and movement patterns of coral reef fishes, and implications for marine reserve network design. Biol. Rev. 2015, 90, 1215–1247. [Google Scholar] [CrossRef]
- Nash, K.L.; Welsh, J.Q.; Graham, N.A.; Bellwood, D.R. Home-range allometry in coral reef fishes: Comparison to other vertebrates, methodological issues and management implications. Oecologia 2015, 177, 73–83. [Google Scholar] [CrossRef]
- Hanski, I.; Ovaskainen, O. Metapopulation theory for fragmented landscapes. Theor. Popul. Biol. 2003, 64, 119–127. [Google Scholar] [CrossRef]
- Sale, P.F.; Cowen, R.K.; Danilowicz, B.S.; Jones, G.P.; Kritzer, J.P.; Lindeman, K.C.; Steneck, R.S. Critical science gaps impede use of no-take fishery reserves. Trends Ecol. Evol. 2005, 20, 74–80. [Google Scholar] [CrossRef]
- Hock, K.; Wolff, N.H.; Ortiz, J.C.; Condie, S.A.; Anthony, K.R.N.; Blackwell, P.G.; Mumby, P.J. Connectivity and systemic resilience of the great barrier reef. PLoS Biol. 2017, 15, e2003355. [Google Scholar] [CrossRef]
- Hock, K.; Doropoulos, C.; Gorton, R.; Condie, S.A.; Mumby, P.J. Split spawning increases robustness of coral larval supply and inter-reef connectivity. Nat. Commun. 2019, 10, 3463. [Google Scholar] [CrossRef] [Green Version]
- Riginos, C.; Hock, K.; Matias, A.M.; Mumby, P.J.; van Oppen, M.J.H.; Lukoschek, V. Asymmetric dispersal is a critical element of concordance between biophysical dispersal models and spatial genetic structure in Great Barrier Reef corals. Divers. Distrib. 2019, 25, 1684–1696. [Google Scholar] [CrossRef] [Green Version]
- White, C.; Selkoe, K.A.; Watson, J.; Siegel, D.A.; Zacherl, D.C.; Toonen, R.J. Ocean currents help explain population genetic structure. Proc. R. Soc. B Biol. Sci. 2010, 277, 1685–1694. [Google Scholar] [CrossRef] [Green Version]
- Cowen, R.K.; Paris, C.B.; Srinivasan, A. Scaling of connectivity in marine populations. Science 2006, 311, 522–527. [Google Scholar] [CrossRef] [Green Version]
- Watson, J.R.; Kendall, B.E.; Siegel, D.A.; Mitarai, S. Changing seascapes, stochastic connectivity, and marine metapopulation dynamics. Am. Nat. 2012, 180, 99–112. [Google Scholar] [CrossRef] [Green Version]
- Green, A.L.; Fernandes, L.; Almany, G.; Abesamis, R.; McLeod, E.; Aliño, P.M.; Pressey, R.L. Designing marine reserves for fisheries management, biodiversity conservation, and climate change adaptation. Coast. Manag. 2014, 42, 143–159. [Google Scholar] [CrossRef]
- Balbar, A.C.; Metaxas, A. The current application of ecological connectivity in the design of marine protected areas. Glob. Ecol. Conserv. 2019, 17, e00569. [Google Scholar] [CrossRef]
- Ospina-Alvarez, A.; de Juan, S.; Alós, J.; Basterretxea, G.; Alonso-Fernández, A.; Follana-Berná, G.; Catalán, I.A. MPA network design based on graph theory and emergent properties of larval dispersal. Mar. Ecol. Prog. Ser. 2020, 650, 309–326. [Google Scholar] [CrossRef]
- Jones, G.P.; Almany, G.R.; Russ, G.R.; Sale, P.F.; Steneck, R.S.; Van Oppen, M.J.H.; Willis, B.L. Larval retention and connectivity among populations of corals and reef fishes: History, advances and challenges. Coral Reefs 2009, 28, 307–325. [Google Scholar] [CrossRef]
- Krueck, N.C.; Ahmadia, G.N.; Possingham, H.P.; Riginos, C.; Treml, E.A.; Mumby, P.J. Marine reserve targets to sustain and rebuild unregulated fisheries. PLoS Biol. 2017, 15, e2000537. [Google Scholar] [CrossRef] [Green Version]
- Harrison, H.B.; Williamson, D.H.; Evans, R.D.; Almany, G.R.; Thorrold, S.R.; Russ, G.R.; Jones, G.P. Larval export from marine reserves and the recruitment benefit for fish and fisheries. Curr. Biol. 2012, 22, 1023–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halpern, B.S.; Warner, R.R. Matching marine reserve design to reserve objectives. Proc. R. Soc. London Ser. B Biol. Sci. 2003, 270, 1871–1878. [Google Scholar] [CrossRef] [Green Version]
- Krueck, N.C.; Ahmadia, G.N.; Green, A.; Jones, G.P.; Possingham, H.P.; Riginos, C.; Mumby, P.J. Incorporating larval dispersal into MPA design for both conservation and fisheries. Ecol. Appl. 2017, 27, 925–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaines, S.D.; White, C.; Carr, M.H.; Palumbi, S.R. Designing marine reserve networks for both conservation and fisheries management. Proc. Natl. Acad. Sci. USA 2010, 107, 18286–18293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, C.M.; Branch, G.; Bustamante, R.H.; Castilla, J.C.; Dugan, J.; Halpern, B.S.; Warner, R.R. Application of ecological criteria in selecting marine reserves and developing reserve networks. Ecol. Appl. 2003, 13 (Suppl. 1), 215–228. [Google Scholar] [CrossRef] [Green Version]
- White, J.W.; Schroeger, J.; Drake, P.T.; Edwards, C.A. The value of larval connectivity information in the static optimization of marine reserve design. Conserv. Lett. 2014, 7, 533–544. [Google Scholar] [CrossRef]
- Beger, M.; Linke, S.; Watts, M.; Game, E.; Treml, E.; Ball, I.; Possingham, H.P. Incorporating asymmetric connectivity into spatial decision making for conservation. Conserv. Lett. 2010, 3, 359–368. [Google Scholar] [CrossRef]
- Mumby, P.J. Bleaching and hurricane disturbances to populations of coral recruits in Belize. Mar. Ecol. Prog. Ser. 1999, 190, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Magris, R.A.; Treml, E.A.; Pressey, R.L.; Weeks, R. Integrating multiple species connectivity and habitat quality into conservation planning for coral reefs. Ecography 2016, 39, 649–664. [Google Scholar] [CrossRef]
- Daigle, R.M.; Metaxas, A.; Balbar, A.C.; McGowan, J.; Treml, E.A.; Kuempel, C.D.; Beger, M. Operationalizing ecological connectivity in spatial conservation planning with marxan connect. Methods Ecol. Evol. 2020, 11, 570–579. [Google Scholar] [CrossRef] [Green Version]
- Ball, I.R.; Possingham, H.P.; Watts, M. Marxan and relatives: Software for spatial conservation prioritisation. Spat. Conserv. Prioritisation Quant. Methods Comput. Tools 2009, 14, 185–196. [Google Scholar]
- Beverton, R.; Holt, S. On the Dynamics of Exploited Fish Populations; Springer: Dordrecht, The Netherlands, 1993. [Google Scholar]
- Holstein, D.M.; Paris, C.B.; Mumby, P.J. Consistency and inconsistency in multispecies population network dynamics of coral reef ecosystems. Mar. Ecol. Prog. Ser. 2014, 499, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Hilborn, R.; Walters, C.J. Quantitative Fisheries Stock Assessment: Choice, Dynamics, and Uncertainty; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1992. [Google Scholar]
- Paris, C.B.; Helgers, J.; Van Sebille, E.; Srinivasan, A. Connectivity modeling system: A probabilistic modeling tool for the multi-scale tracking of biotic and abiotic variability in the ocean. Environ. Model. Softw. 2013, 42, 47–54. [Google Scholar] [CrossRef]
- Walters, C.J.; Hilborn, R.; Parrish, R. An equilibrium model for predicting the efficacy of marine protected areas in coastal environments. Can. J. Fish. Aquat. Sci. 2007, 64, 1009–1018. [Google Scholar] [CrossRef]
- Krueck, N.; Abdurrahim, A.; Adhuri, D.; Mumby, P.; Ross, H. Quantitative decision support tools facilitate social-ecological alignment in community-based marine protected area design. Ecol. Soc. 2019, 24, 6. [Google Scholar] [CrossRef] [Green Version]
- Myers, R.A.; Bowen, K.G.; Barrowman, N.J. Maximum reproductive rate of fish at low population sizes. Can. J. Fish. Aquat. Sci. 1999, 56, 2404–2419. [Google Scholar] [CrossRef]
- Bode, M.; Leis, J.M.; Mason, L.B.; Williamson, D.H.; Harrison, H.B.; Choukroun, S.; Jones, G.P. Successful validation of a larval dispersal model using genetic parentage data. PLoS Biol. 2019, 17, e3000380. [Google Scholar] [CrossRef] [Green Version]
- Taebi, S.; Lowe, R.J.; Pattiaratchi, C.B.; Ivey, G.N.; Symonds, G. A numerical study of the dynamics of the wave-driven circulation within a fringing reef system. Ocean Dyn. 2012, 62, 585–602. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metrics | Definition |
|---|---|
| Average connections | average number of connections at any site of the network, calculated as the sum of all connections divided by the number of sites in the network. |
| Average strength | average larval flow from any site in the network calculated as the sum of all connection weights divided by the number of sites in the network. |
| Biconnected components | a measure of network decomposition into a tree of biconnected components. Biconnected components are connected nodes where if any one node is removed, the remaining nodes will still be connected. Articulation points are the nodes which join biconnected components together to form the global network. Therefore, fewer biconnected components suggests a network which is more cohesive and less disjointed. |
| Region | Average Connections | Average Strength | Biconnected Components |
|---|---|---|---|
| Bahamas | 102 | 0.2 | 3 |
| Belize | 43 | 0.12 | 1 |
| Bonaire | 36 | 0.04 | 6 |
| Lesser Antilles | 46 | 0.08 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tong, C.; Hock, K.; Krueck, N.C.; Tyazhelnikov, V.; Mumby, P.J. An MPA Design Approach to Benefit Fisheries: Maximising Larval Export and Minimising Redundancy. Diversity 2021, 13, 586. https://doi.org/10.3390/d13110586
Tong C, Hock K, Krueck NC, Tyazhelnikov V, Mumby PJ. An MPA Design Approach to Benefit Fisheries: Maximising Larval Export and Minimising Redundancy. Diversity. 2021; 13(11):586. https://doi.org/10.3390/d13110586
Chicago/Turabian StyleTong, Colm, Karlo Hock, Nils C. Krueck, Vladimir Tyazhelnikov, and Peter J. Mumby. 2021. "An MPA Design Approach to Benefit Fisheries: Maximising Larval Export and Minimising Redundancy" Diversity 13, no. 11: 586. https://doi.org/10.3390/d13110586
APA StyleTong, C., Hock, K., Krueck, N. C., Tyazhelnikov, V., & Mumby, P. J. (2021). An MPA Design Approach to Benefit Fisheries: Maximising Larval Export and Minimising Redundancy. Diversity, 13(11), 586. https://doi.org/10.3390/d13110586