
Two New Species of the Genus Longipedia Claus, 1863 (Copepoda: Harpacticoida: Longipediidae) from Korea, with an Update and a Key to Species †


Abstract
:1. Introduction
2. Material and Methods
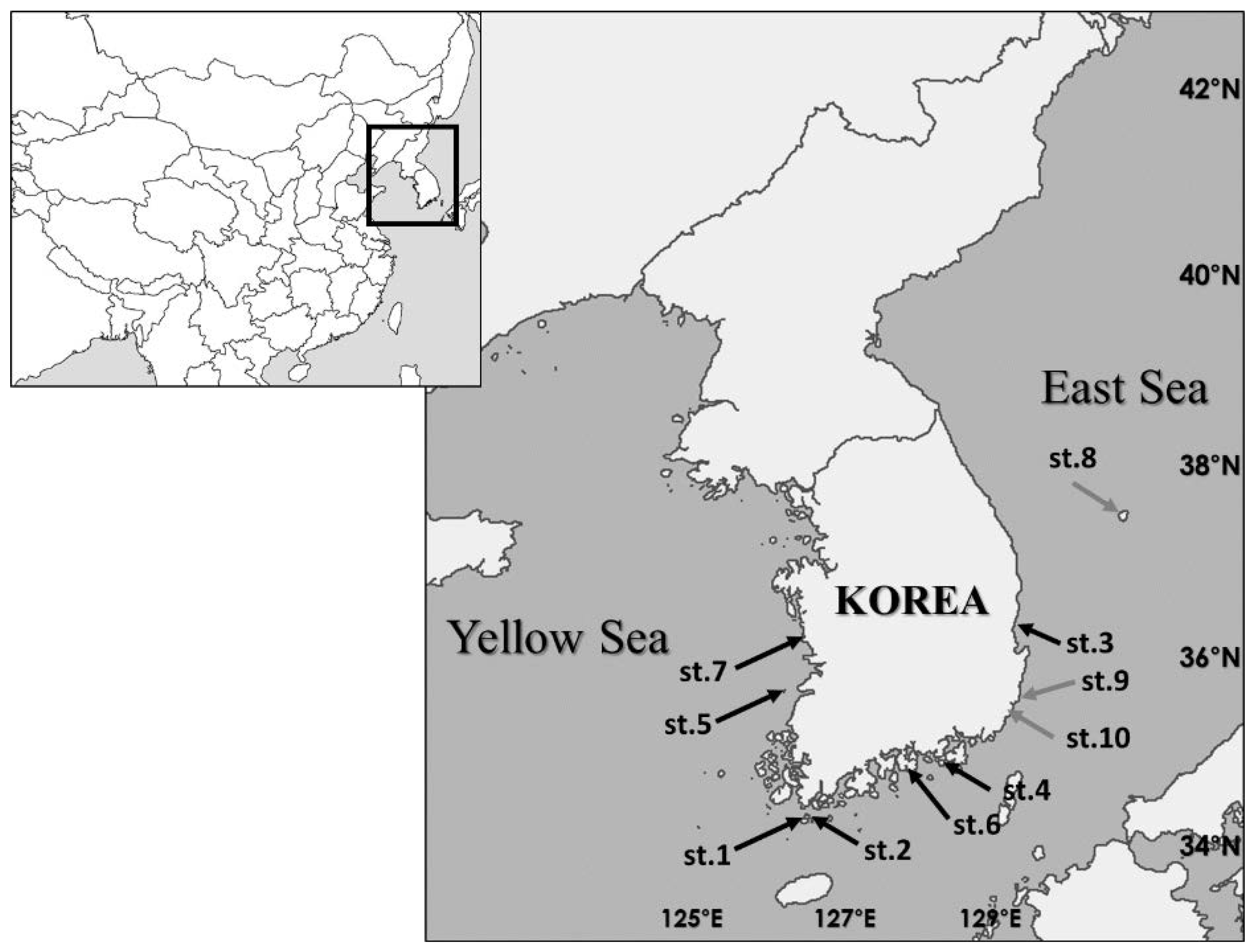
2.1. Sampling Locations
2.2. DNA Extraction and Sequencing
2.3. Morphological Characterization
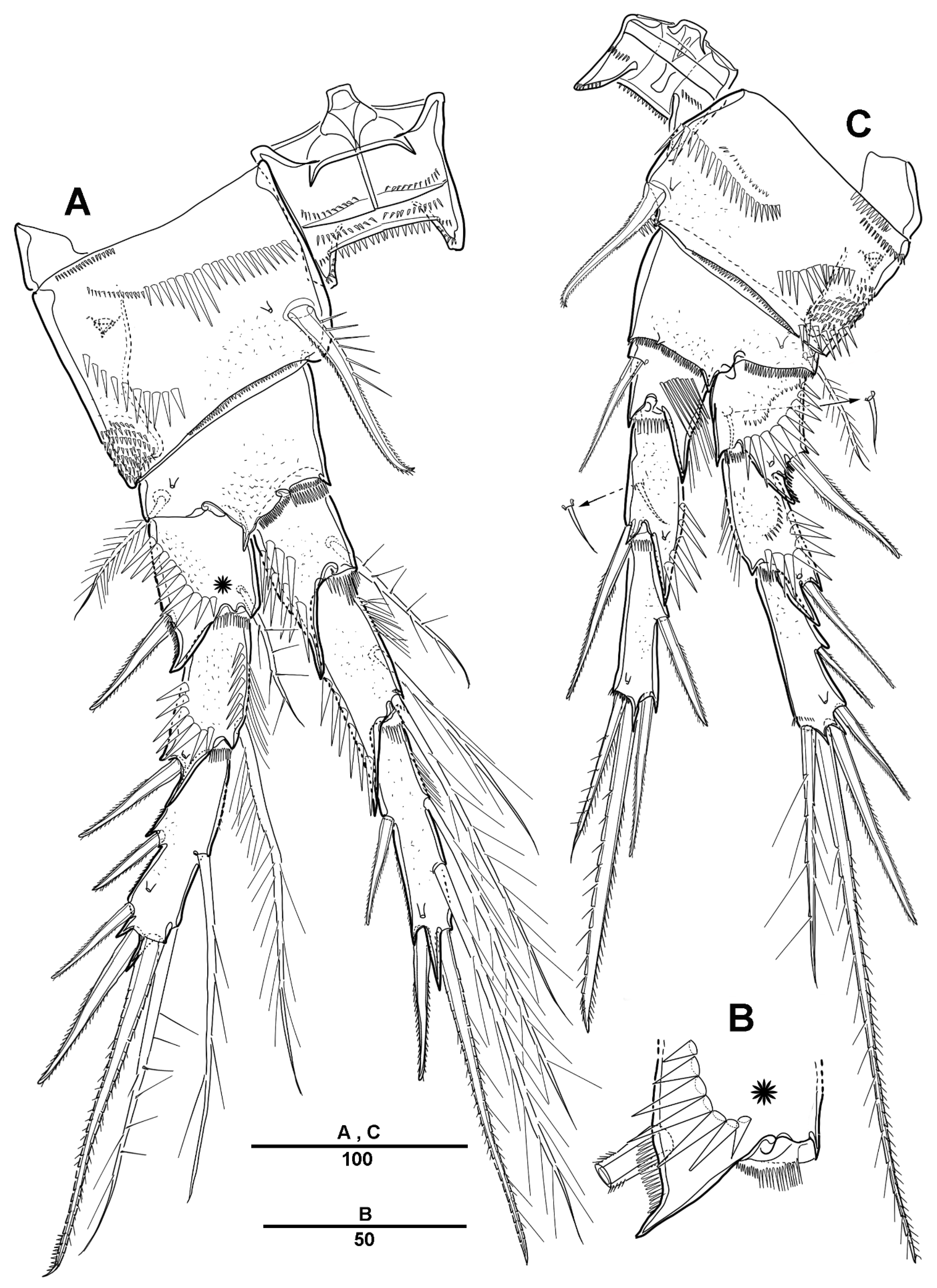
3. Results
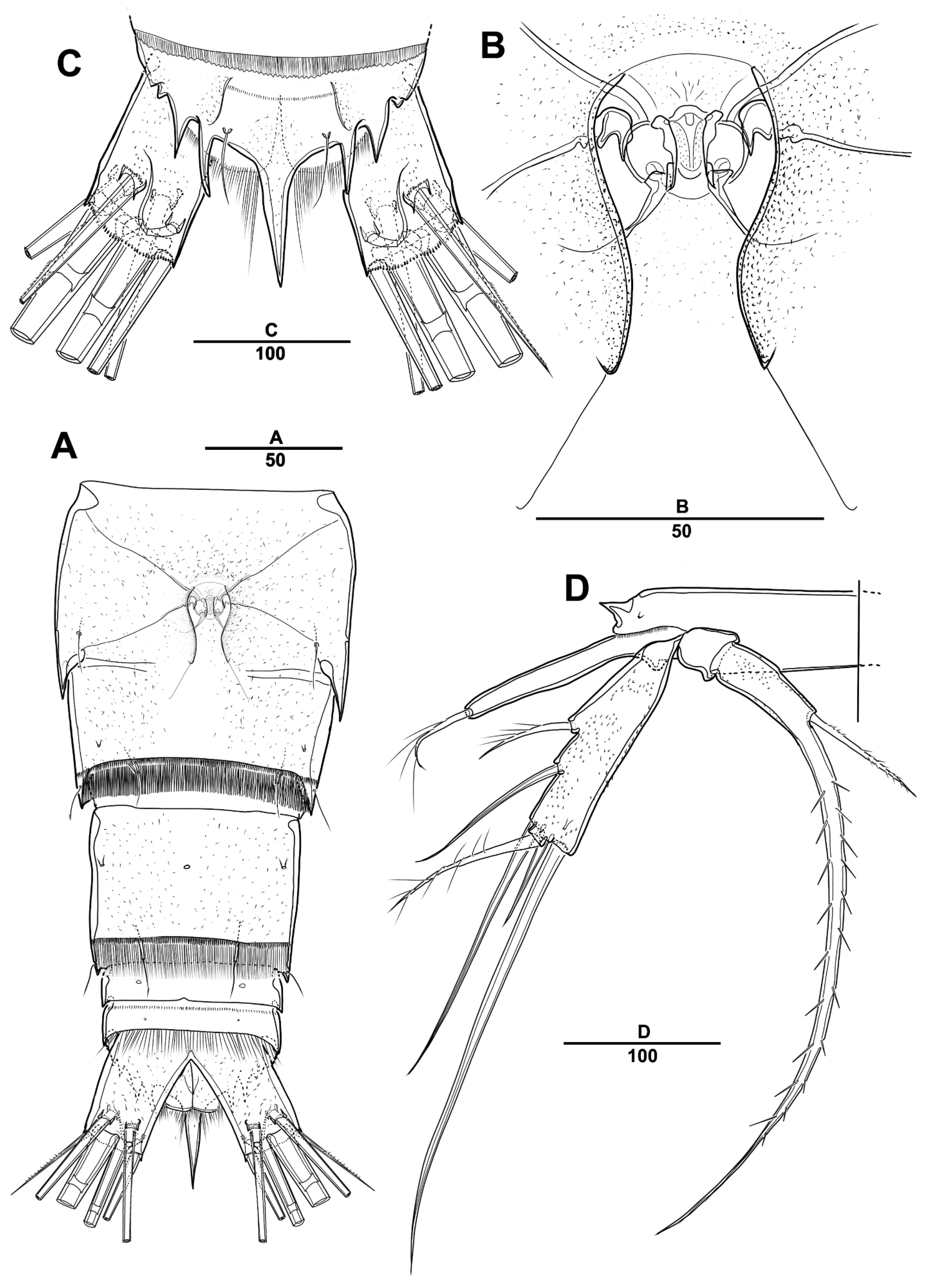
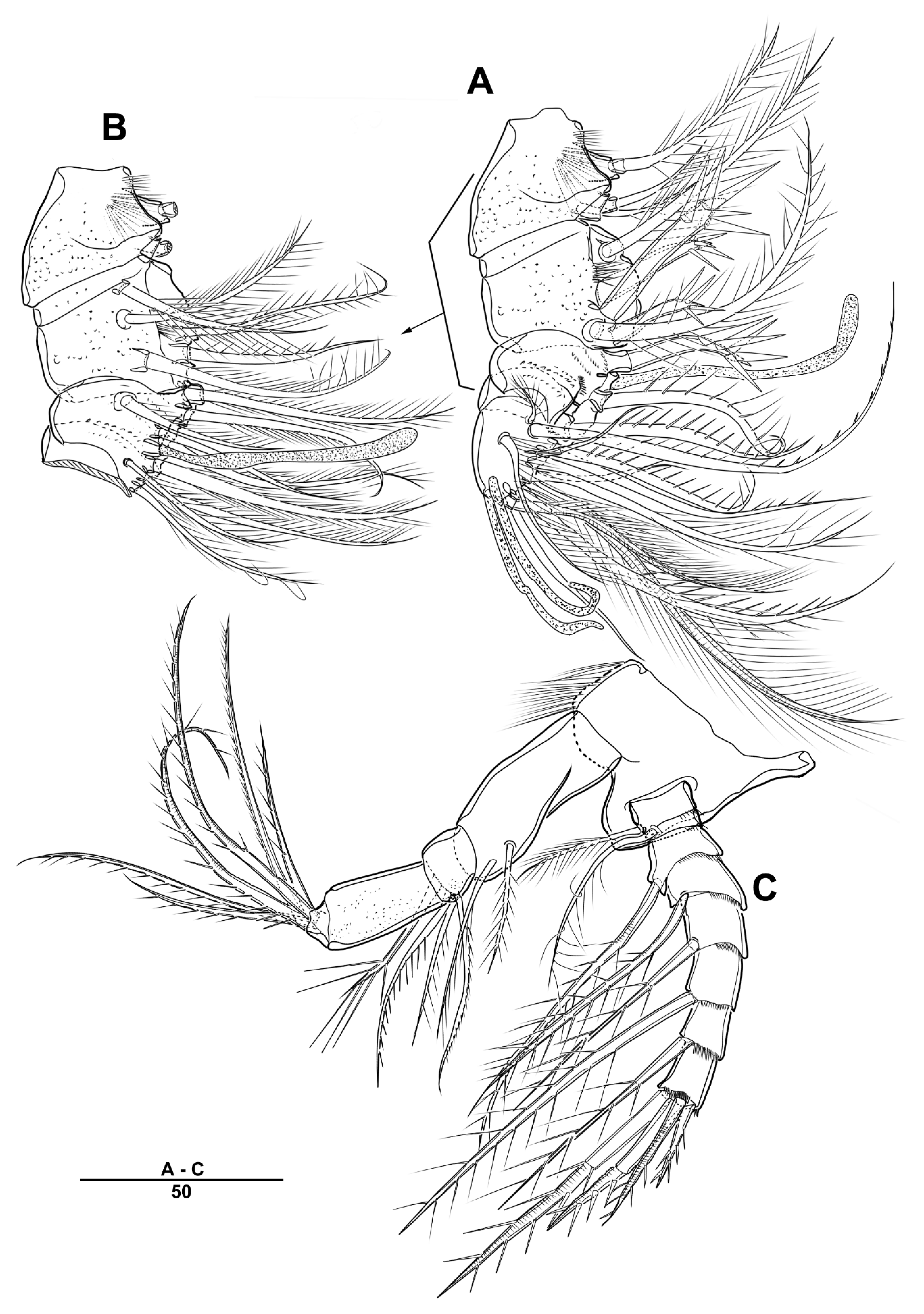
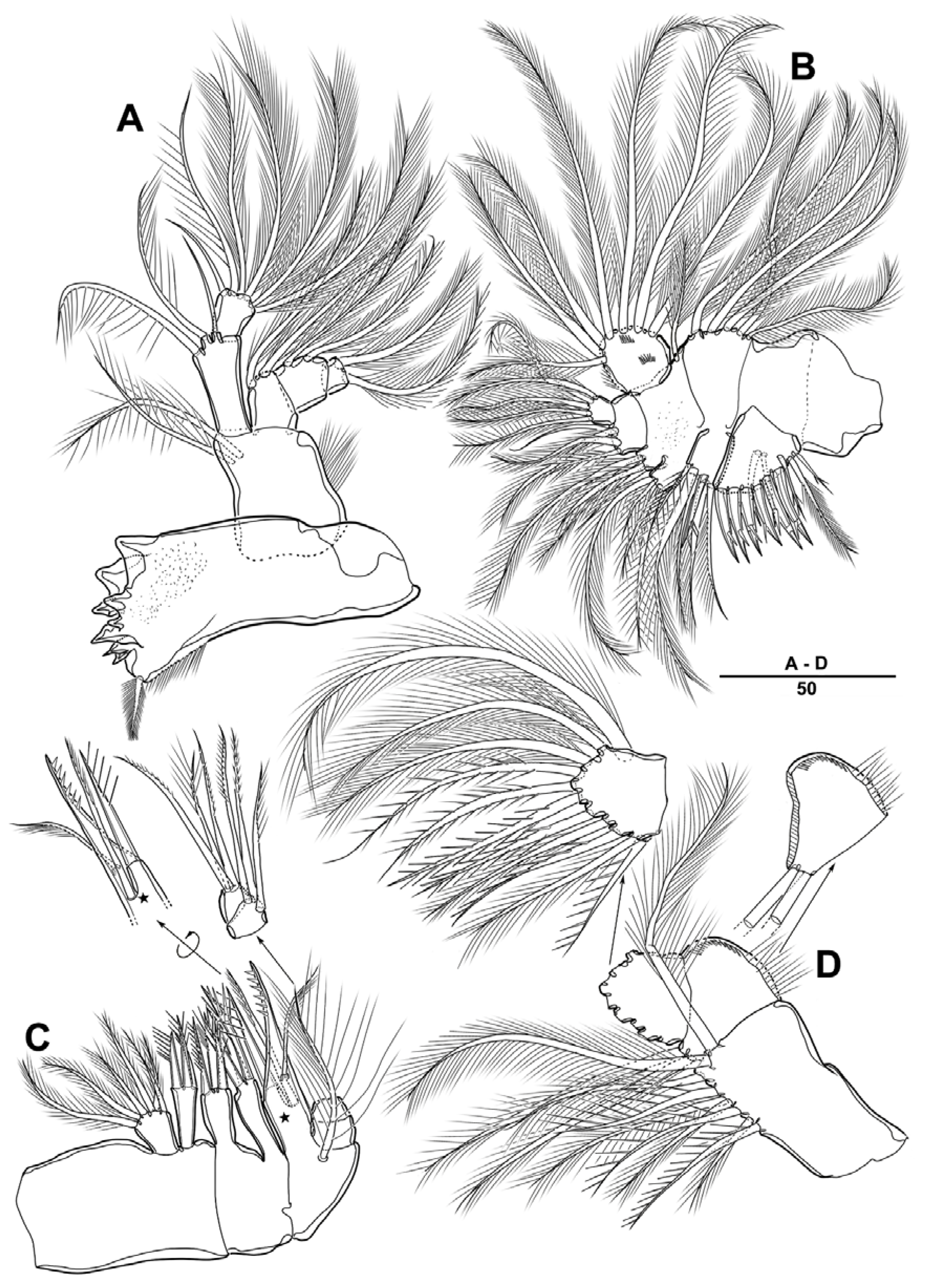
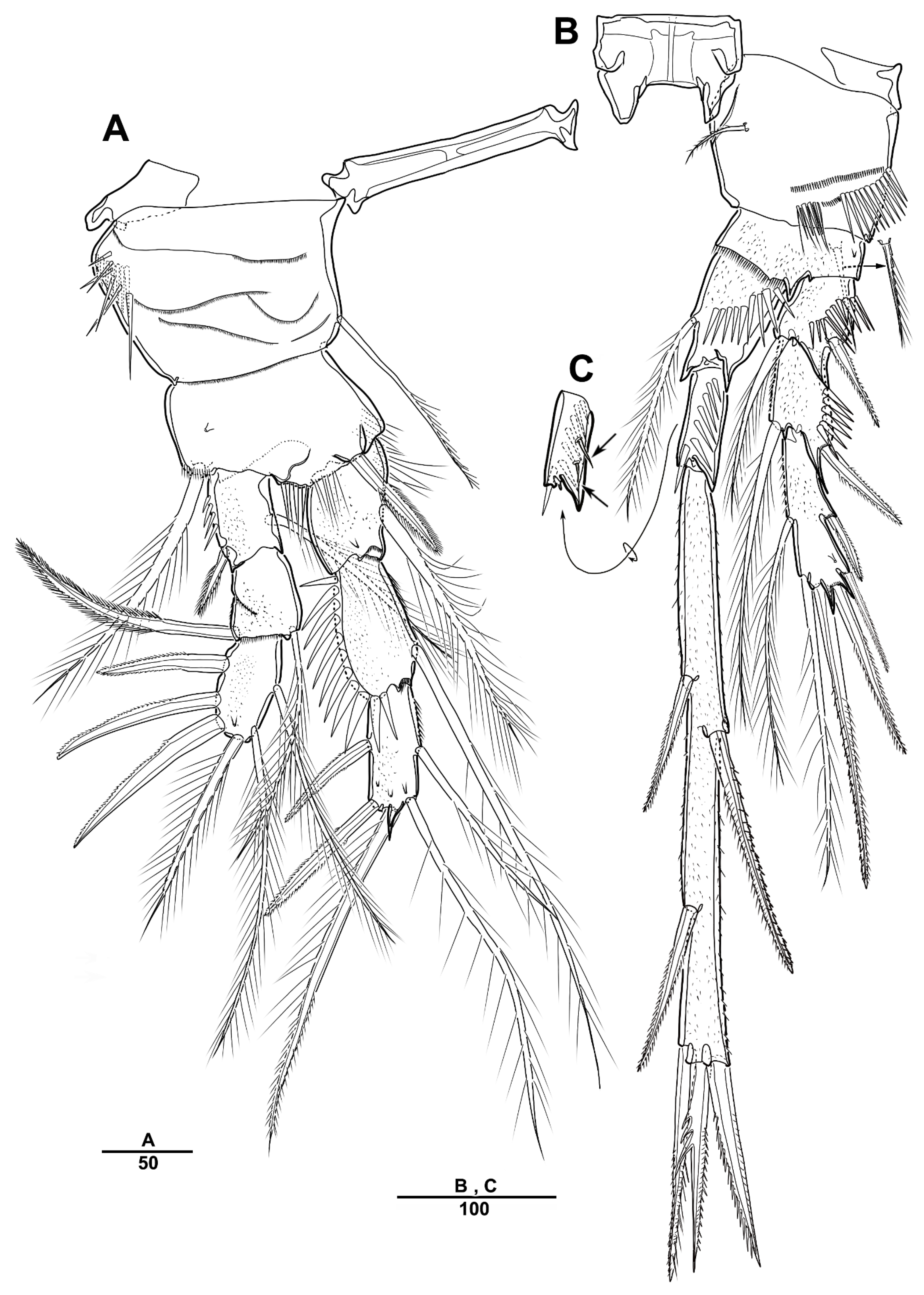
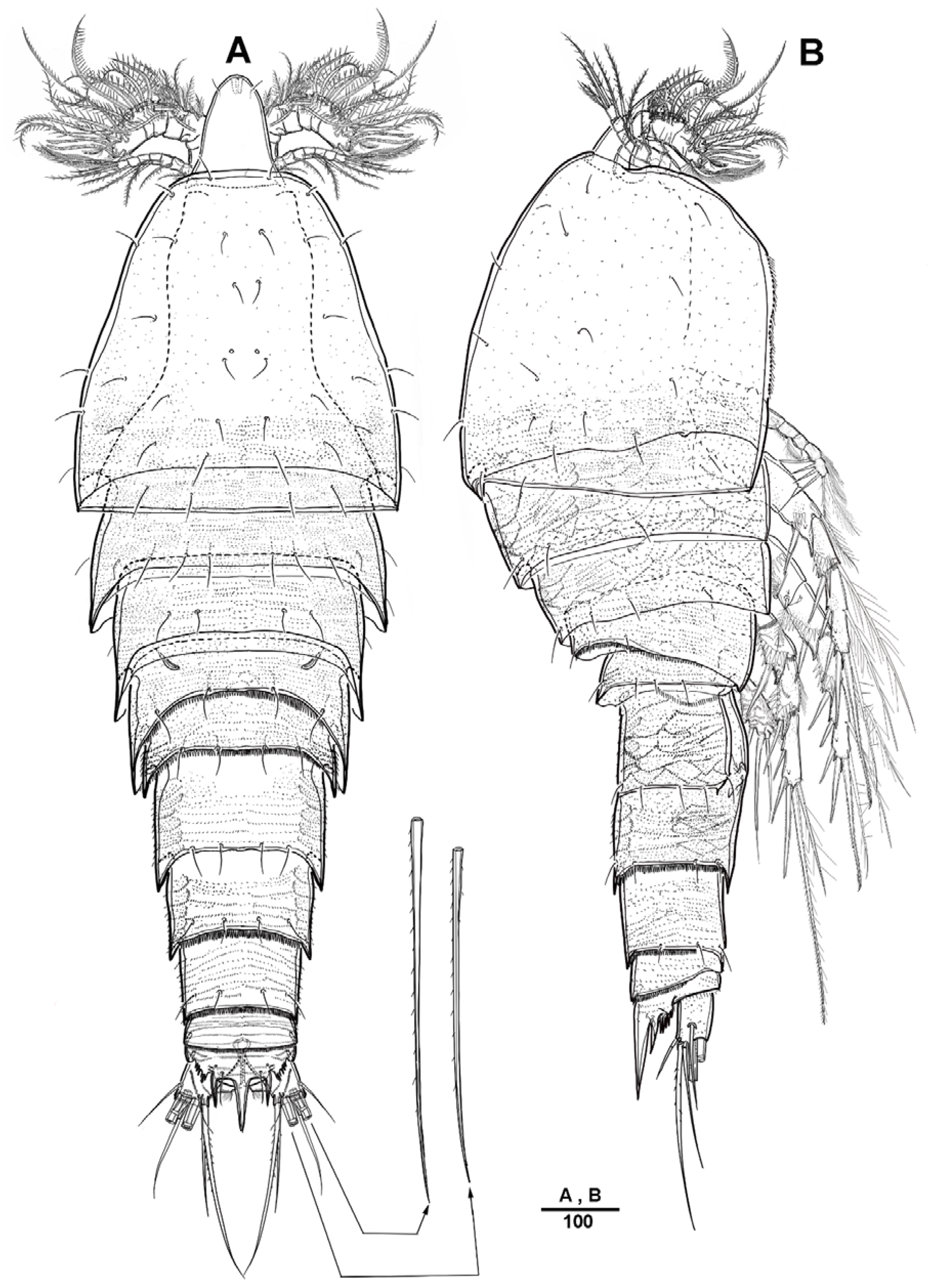
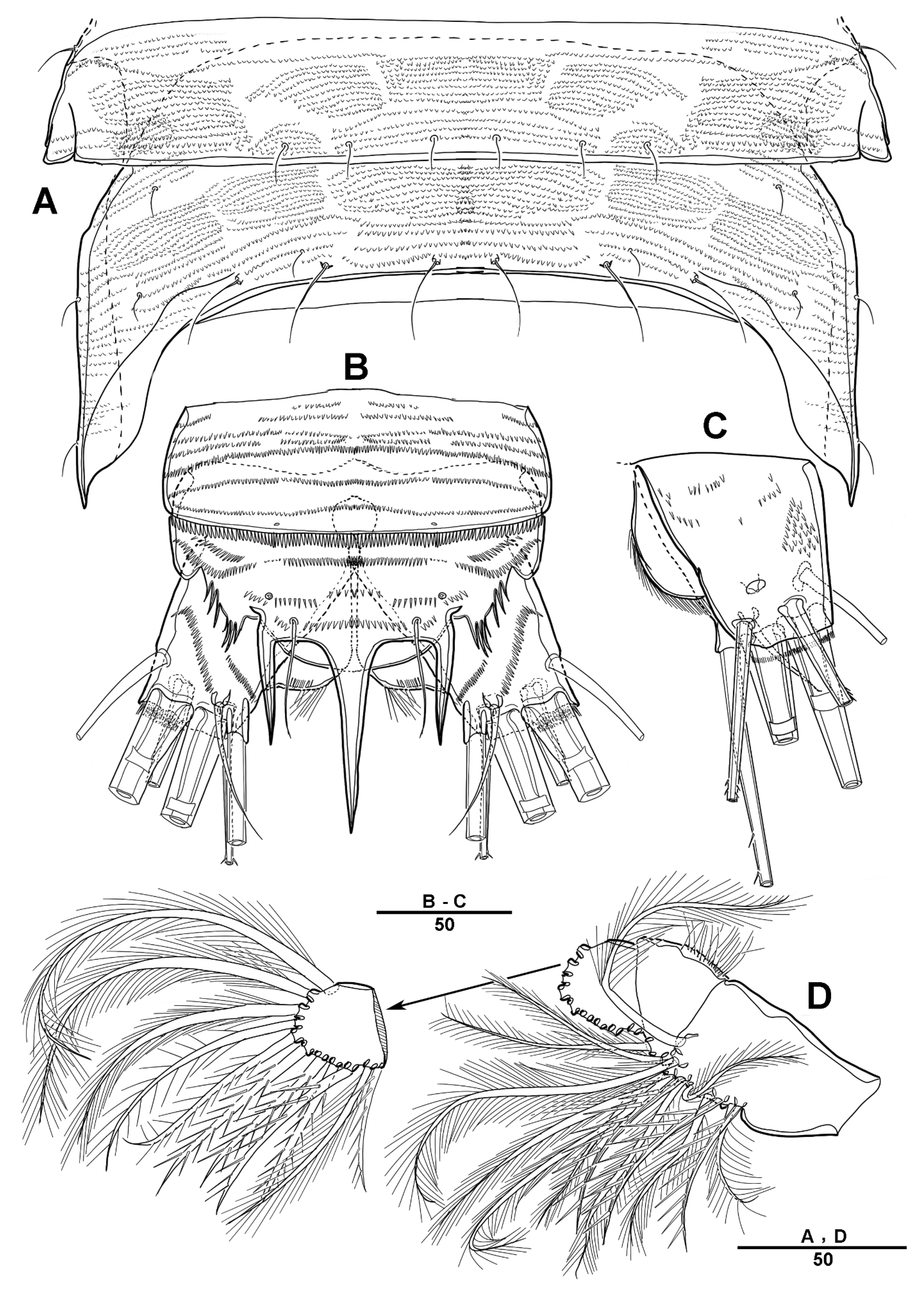
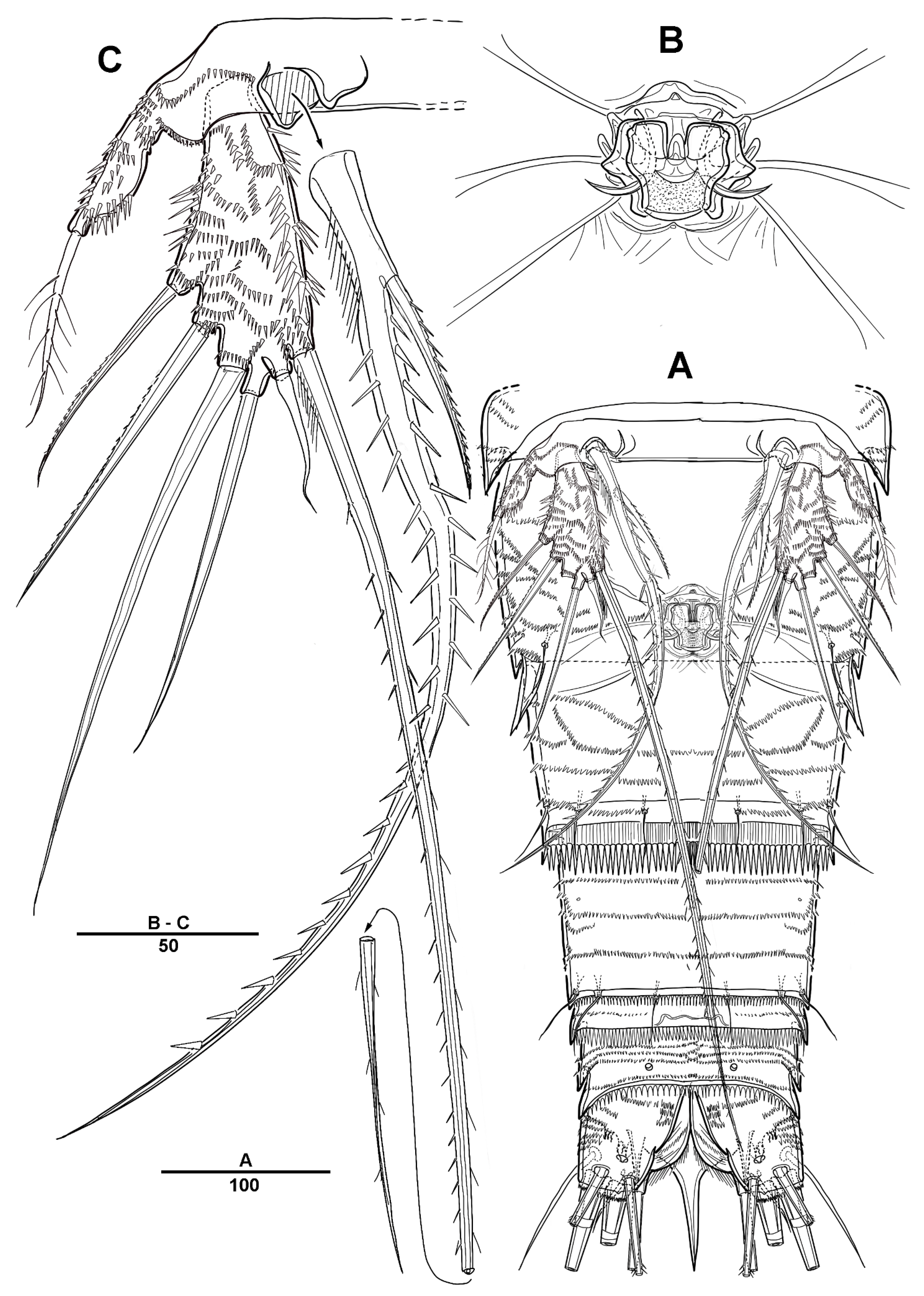
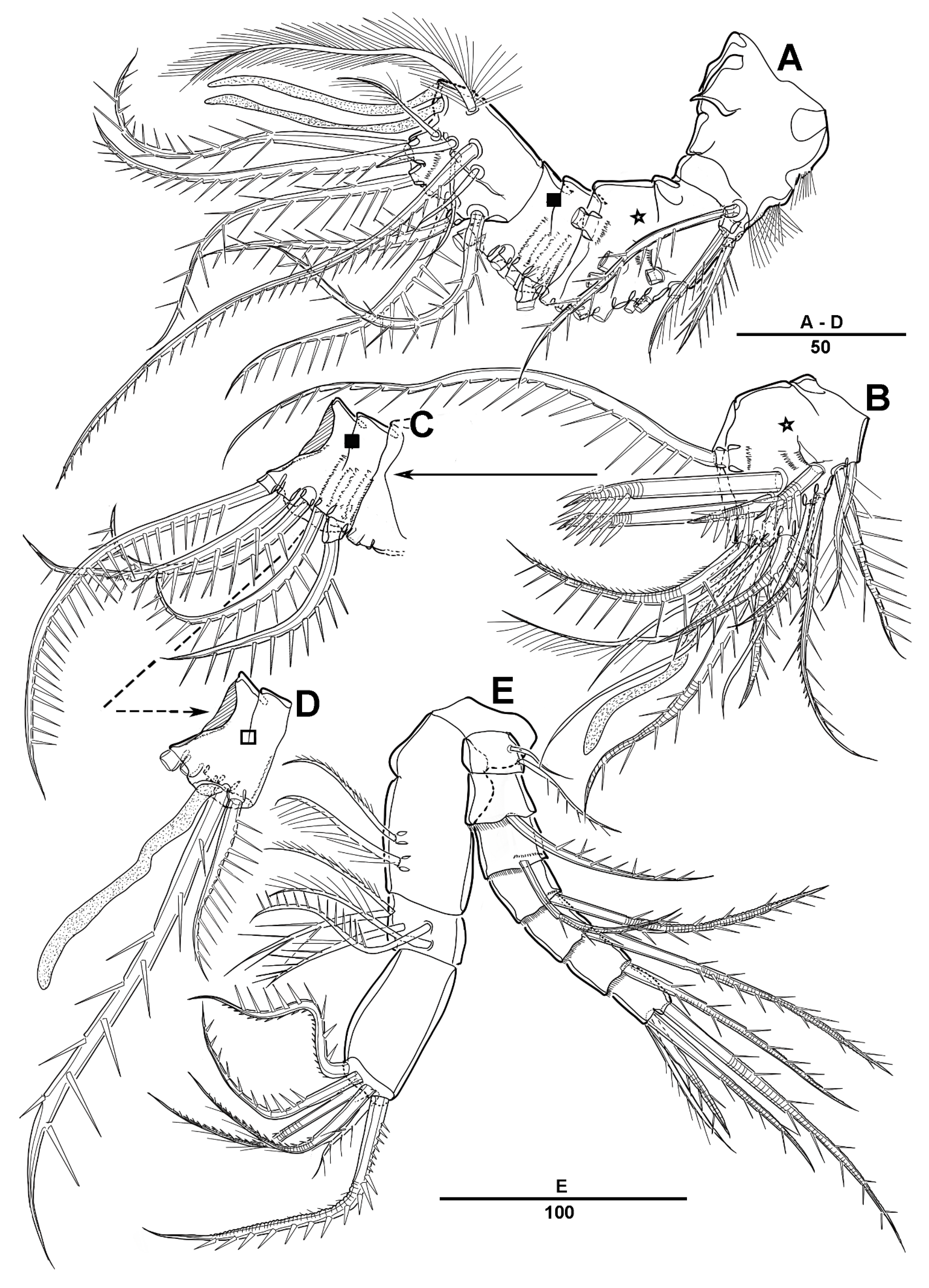
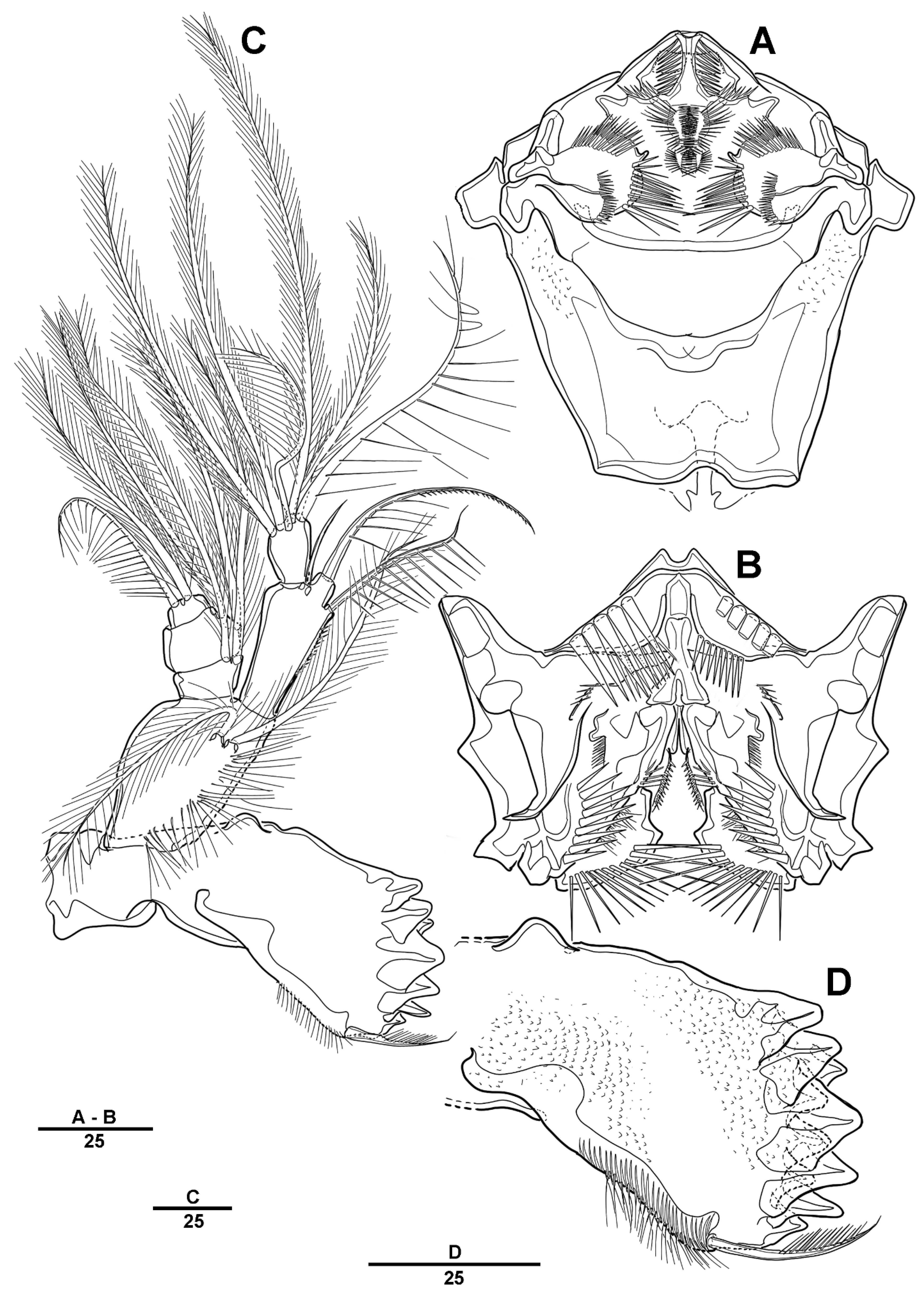
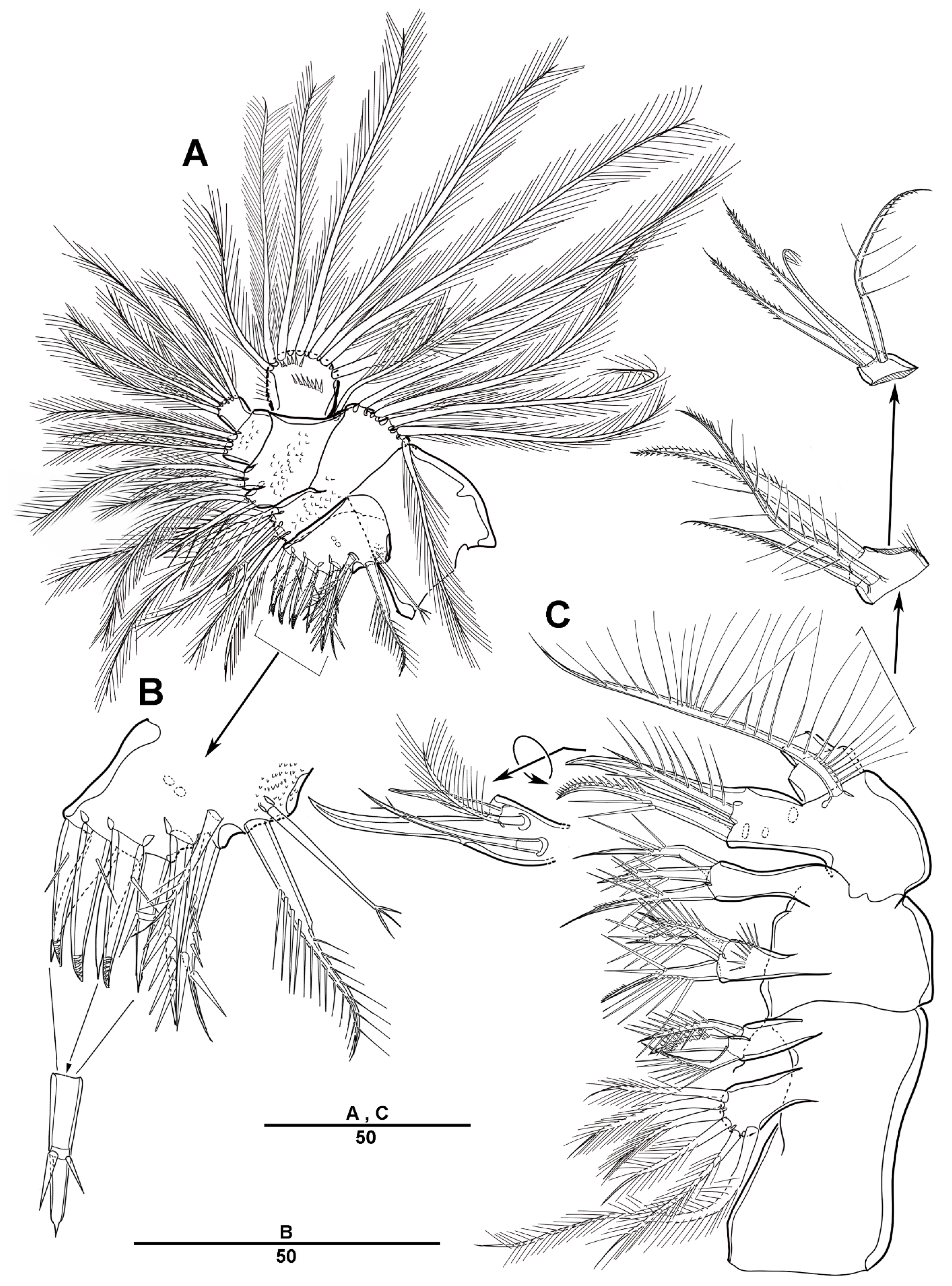
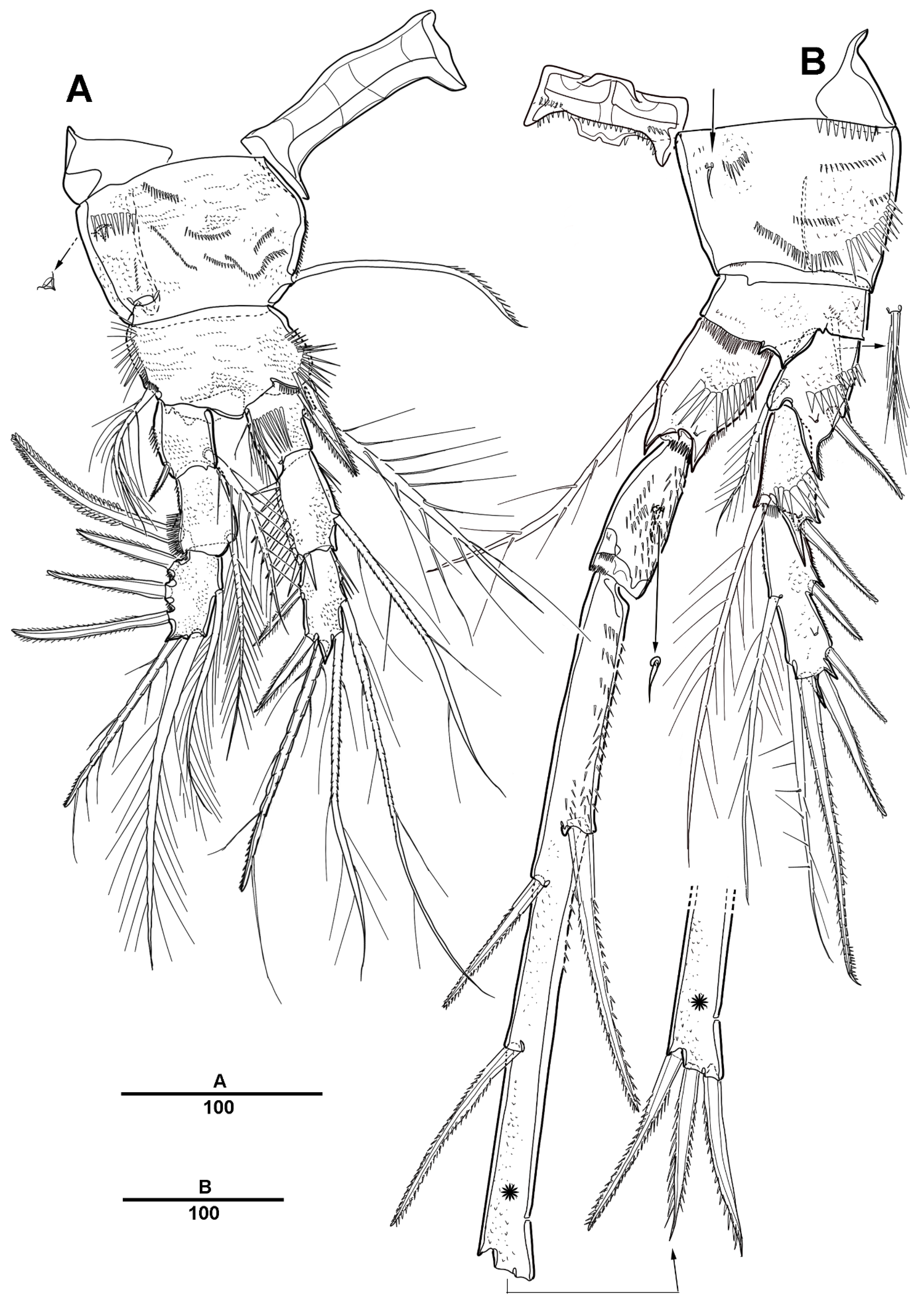
3.1. Longipedia koreana sp. nov.
| Exopod | Endopod | |
| P1 | 1.1.213 | 1.1.212 |
| P2 | 1.1.213 | 1.2.231 |
| P3 | 1.1.222 | 1.2.222 |
| P4 | 0.1.122 | 1.2.022 |
3.2. Longipedia ulleungensis sp. nov.
| Exopod | Endopod | |
| P1 | 1.1.123 | 1.1.212 |
| P2 | 1.1.213 | 1.1.231 |
| P3 | 1.1.222 | 1.2.212 |
| P4 | 1.0.122 | 1.2.022 |
3.3. DNA Sequences of Two New Species
4. Discussion
- 1
- P4 enp-1 with setiform inner element ……………… helgolandica species-group… 2
- -
- P4 enp-1 with spiniform inner element ………………………………………………………… 5
- 2
- P2 coxa with well-developed inner seta; P5 exopod rectangular …………………………………………………………………………… Longipedia helgolandica Klie, 1949
- -
- P2 coxa with small or slender inner seta; P5 exopod triangular, apex wider than base ……………………………………………………………………………………3
- 3
- P5 baseoendopod not articulated ………………………………L. americana Wells, 1980
- -
- P5 baseoendopod articulated, with 2-segment endopodal lobe ……………………4
- 4
- P1 basis with long setules on distal edge; P2 coxa with very reduced inner seta ……………………………………………………………… L. corteziensis Gómez, 2001
- -
- P1 basis distal edge smooth; P2 coxa with slender and relatively longer inner seta ……………………………………………… L. santacruzensis Mielke, 1979
- 5
- P2 enp-1 with no projection on distal corner of anterior surface …………………………………………………………………… L. minor Scott T. & Scott A., 1893
- -
- P2 enp-1 with unguiform projection on corner of anterior surface ……………………………………………………………………………………………………………………… 6
- 6
- P2 enp-2 without armature element on the posterior surface …………………………………………………… L. thailandensis Chullasorn & Kangtia, 2008
- -
- P2 enp-2 with one tiny seta on the posterior surface …………………………………… 7
- -
- P2 enp-2 with two tiny setae on the posterior surface …………………………………… 9
- 7
- P4 exp-2 without inner seta; P2 coxa with much reduced inner seta; anal operculum with a long median projection (much longer than lateral projections), and a single spine and group of outer spines on each side……………………………………………………………….. L. ulleungensis sp. nov
- -
- P4 exp-2 with inner seta; P2 coxa with well-developed inner seta; anal operculum with a short median projection (no longer than lateral projections), and two spines on each side … L. brevispinosa Gurney, 1927
- -
- P4 exp-2 with inner seta; P2 coxa with well-developed inner seta; anal operculum with a long median projection (much longer than lateral projections), and two spines on each side ………………………………………… 8
- 8
- P5 exopod about three times as long as wide; P2 enp-3 with a terminaldentate spine (median apical spine with a large tooth about halfway along) ……………………………………………………………………………… L. spinulosa Itô, 1981
- -
- P5 exopod about twice as long as wide; P2 enp-3 with serrated apicalspines ………………………………………………………………………… L. weberi Scott A., 1909
- 9
- P2 coxa with small or reduced inner seta, or without inner seta ………………… 10
- -
- P2 coxa with well-developed inner seta ………………………………………………………11
- 10
- P2 coxa without inner seta; P4 exp-1 with a minute inner seta; P5 exopod about twice as long as wide; P5 endopod with well-developed inner seta ……………………………………………… L. andamanica Wells, 1980
- -
- P2 coxa with very small inner seta, originated from the posterior surface; P4 exp-1 with a minute inner seta; P5 exopod at most 1.3 times as long as wide; P5 endopod with small and bare inner seta…………………………………………………………………………………………… L. scotti Sars, 1903
- -
- P2 coxa with small inner seta on anterior surface; P4 exp-1 without armature element; P5 exopod at least 3.5 times as long as wide; P5endopod with well-developed inner seta………………………… L. koreana sp. nov.
- 11
- P1 coxa distal spinule much longer than the proximal spinule; P2 enp-3 median terminal spine with a large tooth about halfway along its length; P5 exopod about three times as long as wide……………………………………………………………………………………… L. nichollsi Wells, 1980
- -
- P1 coxa distal spinule much longer than the proximal spinule; P2 enp-3 median terminal spine with a large tooth about halfway along, or P2 enp-3 with three terminal spinulose claws; P5 exopod about twice as long as wide ……………………………………………… L. kikuchii Itô, 1980
- -
- P1 coxa with row of similar sized setules on outer side; P2 enp-3 with three terminal spinulose spines; P5 exopod about twice as long as wide ………………………………………………………………… L. coronata Claus, 1863
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wells, J.B.J. A revision of the genus Longipedia Claus (Crustacea: Copepoda: Harpacticoida). Zool. J. Linn. Soc. 1980, 70, 103–189. [Google Scholar] [CrossRef]
- Boxshall, G.A.; Halsey, S.H. An Introduction to Copepod Diversity; Ray Society: London, UK, 2004; pp. 1–966. [Google Scholar]
- Hulme, P.E. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Ganapati, P.N.; Shanthakumari, K. The systematics and distribution of planktonic copepods in the Lawson’s Bay, Waltair. J. Mar. Biol. Assoc. India 1961, 3, 6–18. [Google Scholar]
- Itô, T. A new subspecies of Longipedia andamanica Wells from the Pacific coast of Japan, with reference to the morphology of L. coronata Claus (Copepoda: Harpacticoida). Publ. Seto Mar. Biol. Lab. 1985, 30, 307–324. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.G. Taxonomic Study on the Harpacticoids (Crustacea: Copepoda: Harpacticoida) from Marine and Brackish Waters in Korea. Doctoral Dissertation; Chosun University: Gwangju, Korea, 2017. [Google Scholar]
- Itô, T. Two species of the genus Longipedia Claus from Japan, with reference to the taxonomic status of L. weberi previously reported from Amakusa, southern Japan (Copepoda: Harpacticoida). J. Nat. Hist. 1980, 14, 17–32. [Google Scholar] [CrossRef]
- Kim, S.H. Invertebrate fauna of Korea. In Arthropoda: Maxillopoda: Copepoda: Harpacticoida. Marine Harpacticoida II; National Institute of Biological Resources: Incheon, Korea, 2013; Volume 21, Number 27; pp. 1–113. [Google Scholar]
- Burgess, R. An improved protocol for separating meiofauna from sediments using colloidal silica sols. Mar. Ecol. Prog. Ser. 2001, 214, 161–165. [Google Scholar] [CrossRef]
- Lim, B.J.; Bang, H.W.; Moon, H.; Back, J. Integrative description of Diosaccus koreanus sp. nov. (Hexanauplia, Harpacticoida, Miraciidae) and integrative information on further Korean species. ZooKeys 2020, 927, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huys, R.; Gee, J.M.; Moore, C.G.; Hamond, R. Marine and Brackish Water Harpacticoid copepods. In Synopses of the British Fauna (New Series); Field Studies Council: Shrewsbury, UK, 1996; Part 1; pp. 4–19. [Google Scholar]
- Boeck, A. Oversigt over de ved Norges Kyster jagttagne Copepoder henhörende til Calanidernes, Cyclopidernes og Harpactidernes Familier. Forhandlinger i Videnskabsselskabet i Kristiania 1865, 1864, 226–282. [Google Scholar]
- Lang, K. Monographie der Harpacticiden; Håkan Ohlssons Boktryckeri: Lund, Sweden, 1948; pp. 1–1682. [Google Scholar]
- Siegfried, S. Phylogeny of Harpacticoida (Copepoda): Revision of “Maxillipedasphalea” and Exanechentera; Cuvillier Verlag: Göttingen, Germany, 2003; pp. 1–259. [Google Scholar]
- Walter, T.C.; Boxshall, G.; World of Copepods database. Longipedia Claus. 1863. Available online: http://www.marinespecies.org/copepoda/aphia.php?p=taxdetails&id=115403 (accessed on 18 October 2021).
- Mikhailov, K.V.; Ivanenko, V.N. Low support values and lack of reproducibility of molecular phylogenetic analysis of Copepoda orders. Arthropoda Sel. 2021, 30, 39–42. [Google Scholar] [CrossRef]
- Itô, T. Descriptions and records of marine harpacticoid copepods from Hokkaido, VIII. J. Fac. Sci. Hokkaido Univ. 1981, 22, 422–450. [Google Scholar]
- Gómez, S. Longipedia corteziensis sp. nov. (Copepoda, Harpacticoida, Longipediidae) from a coastal lagoon in northwestern Mexico, with the definition of the helgolandica species-group of the genus Longipedia Claus, 1863. Hydrobiologia 2001, 453/454, 483–496. [Google Scholar]
- Chullasorn, S.; Kangtia, P. Longipedia thailandensis sp. nov. (Copepoda, Harpacticoida) from a brackish water treatment pond, Samut Sakhon, Thailand. Crustaceana 2008, 81, 207–225. [Google Scholar]
- Schizas, N.V.; Dahms, H.U.; Kangtia, P.; Corgosinho, P.H.; Galindo Estronza, A.M. A new species of Longipedia Claus, 1863 (Copepoda: Harpacticoida: Longipediidae) from Caribbean mesophotic reefs with remarks on the phylogenetic affinities of Polyarthra. Mar. Biol. Res. 2015, 11, 789–803. [Google Scholar] [CrossRef]
- Onbé, T. The developmental stages of Longipedia americana (Copepoda: Harpacticoida) reared in the laboratory. J. Crustac. Biol. 1984, 4, 615–631. [Google Scholar] [CrossRef]
- Yeom, J.; Lee, W. A new species of the genus Sarsamphiascus Huys, 2009 (Copepoda: Harpacticoida: Miraciidae) from a sublittoral zone of Hawaii. PeerJ 2020, 8, e8506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wells, J.B. An annotated checklist and keys to the species of Copepoda Harpacticoida (Crustacea). Zootaxa 2007, 1568, 1–872. [Google Scholar] [CrossRef] [Green Version]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Date | Locality | Methods (Depth) | Specimen Nos. |
|---|---|---|---|---|
| L. koreana sp. nov. | 29 August 2018 | St. 1: N 34°13′34.68″ E 126°31′35.37″ | Grab (20 m) | MABIK CR00248546 |
| MABIK CR00248533 | ||||
| MABIK CR00248534 | ||||
| MABIK CR00248535 | ||||
| MABIK CR00248536 | ||||
| St. 2: N 34°12′15.82″ E 126°32′50.87″ | Grab (8 m) | MABIK CR00248532 | ||
| MABIK CR00248539 | ||||
| MABIK CR00248540 | ||||
| 21 November 2018 | St. 3: N 36°22′11.69″ E 129°24′28.73″ | Scuba diving (15 m) | MABIK CR00248531 | |
| MABIK CR00248545 | ||||
| 27 June 2019 | St. 4: N 34°47′7″ E 128°27′32.42″ | Grab (15 m) | MABIK CR00248537 | |
| 26 September 2019 | St. 5: N 35°34′16.74″ E 126°16′49.94″ | Grab (9 m) | MABIK CR00248538 | |
| 21 October 2020 | St. 6: N 34°42′58.0″ E 127°59′29.0″ | Scuba diving (9 m) | MABIK CR00248541 | |
| 13 May 2021 | St. 7: N 36°13′18.0″ E 126°31’32.0” | Beach (intertidal) | MABIK CR00248542 | |
| L. ulleungensis sp. nov. | 25 July 2017 | St. 8: N 37°29′13.14″ E 130°55′19.62″ | Scuba diving (15 m) | MABIK CR00248543 |
| 26 April 2018 | St. 9: N 35°28′47.52″ E 129°25′38.88″ | Grab (10 m) | MABIK CR00248544 | |
| MABIK CR00248530 | ||||
| 27 April 2018 | St. 10: N 35°20′53.9″ E 129°20′19.5″ | Grab (10 m) | MABIK CR00248528 | |
| MABIK CR00248529 |
| Gene | Primer | Primer Sequence | PCR Condition |
|---|---|---|---|
| COI | LCO149 HCO2198 | GGTCAACAAATCATAAAGATATTGG TAAACTTCAGGGTGACCAAAAAATCA | 94 °C, 300 s; 40 cycles × (94 °C, 60 s; 48 °C, 120 s; 72 °C, 180 s; 72 °C, 600 s) |
| 18S rRNA | 18S F1 18S R9 18S F2 18S F3 18S F4 18S R6 18S R7 18S R8 | TACCTGGTTGATCCTGCCAG GATCCTTCCGCAGGTTCACCTAC CCTGAGAAACGGCTRCCACAT GYGRTCAGATACCRCCSTAGTT GGTCTGTGATGCCCTYAGATGT TYTCTCRKGCTBCCTCTCC GYYARAACTAGGGCGGTATCTG ACATCTRAGGGCATCACAGACC | 94 °C, 300 s; 40 cycle × (94 °C, 30 s; 50 °C, 30 s; 72 °C, 60 s); 72 °C, 420 s Internal primers, 18S F2–F4 and 18S R6–R8, were used for primer walking to sequence over 1700 bps |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bang, H.W.; Moon, H.; Back, J. Two New Species of the Genus Longipedia Claus, 1863 (Copepoda: Harpacticoida: Longipediidae) from Korea, with an Update and a Key to Species. Diversity 2021, 13, 590. https://doi.org/10.3390/d13110590
Bang HW, Moon H, Back J. Two New Species of the Genus Longipedia Claus, 1863 (Copepoda: Harpacticoida: Longipediidae) from Korea, with an Update and a Key to Species. Diversity. 2021; 13(11):590. https://doi.org/10.3390/d13110590
Chicago/Turabian StyleBang, Hyun Woo, Heejin Moon, and Jinwook Back. 2021. "Two New Species of the Genus Longipedia Claus, 1863 (Copepoda: Harpacticoida: Longipediidae) from Korea, with an Update and a Key to Species" Diversity 13, no. 11: 590. https://doi.org/10.3390/d13110590
APA StyleBang, H. W., Moon, H., & Back, J. (2021). Two New Species of the Genus Longipedia Claus, 1863 (Copepoda: Harpacticoida: Longipediidae) from Korea, with an Update and a Key to Species. Diversity, 13(11), 590. https://doi.org/10.3390/d13110590