
Validity of Pampus liuorum Liu & Li, 2013, Revealed by the DNA Barcoding of Pampus Fishes (Perciformes, Stromateidae)

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Species Identification
2.2. DNA Extraction, Amplification and Sequencing
2.3. Phylogenetic Inference and Barcoding Gaps
2.4. Species Delimitation
3. Results
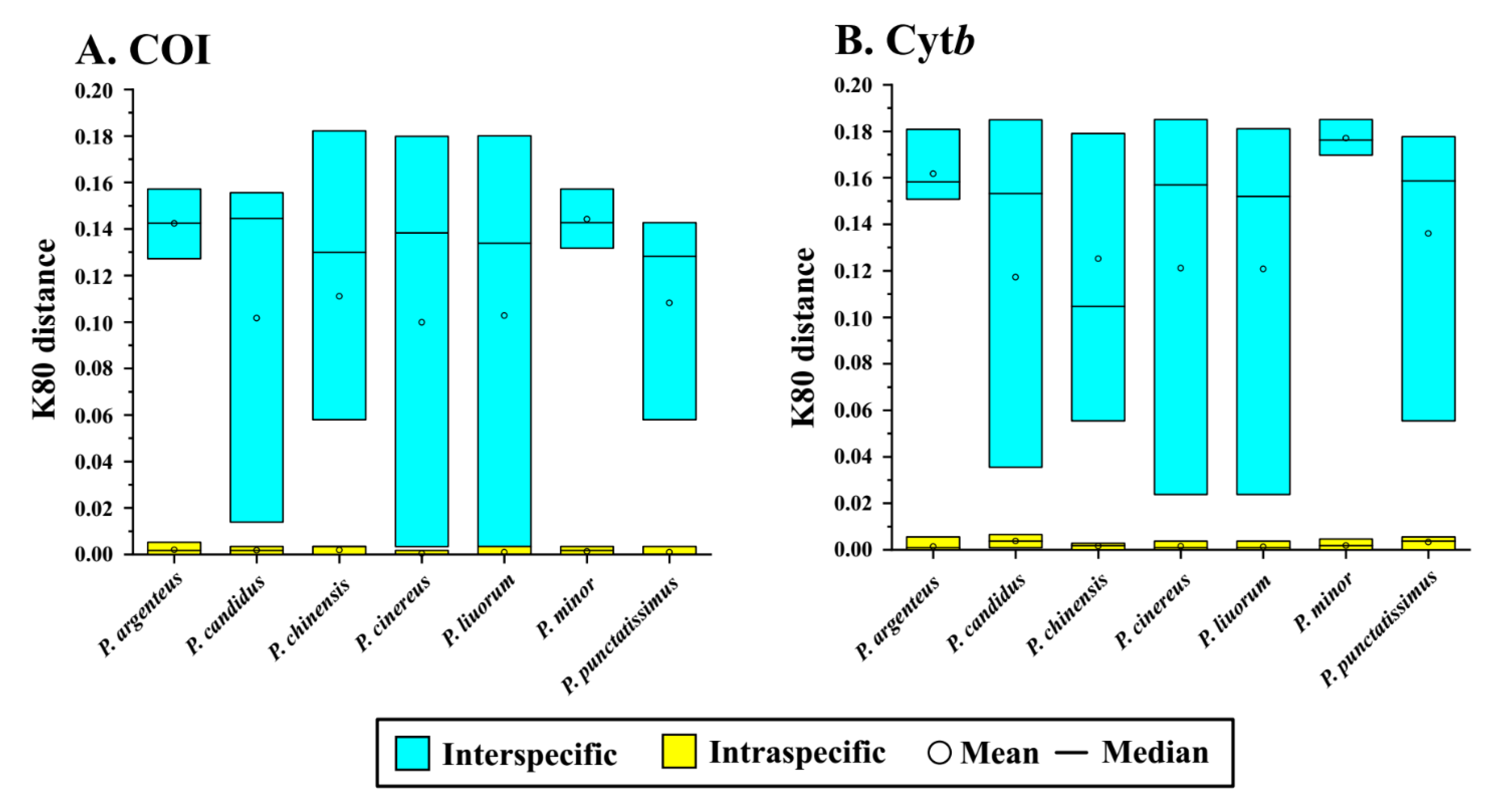
3.1. Sequence Variation Indices and Barcoding Gaps of COI and Cytb
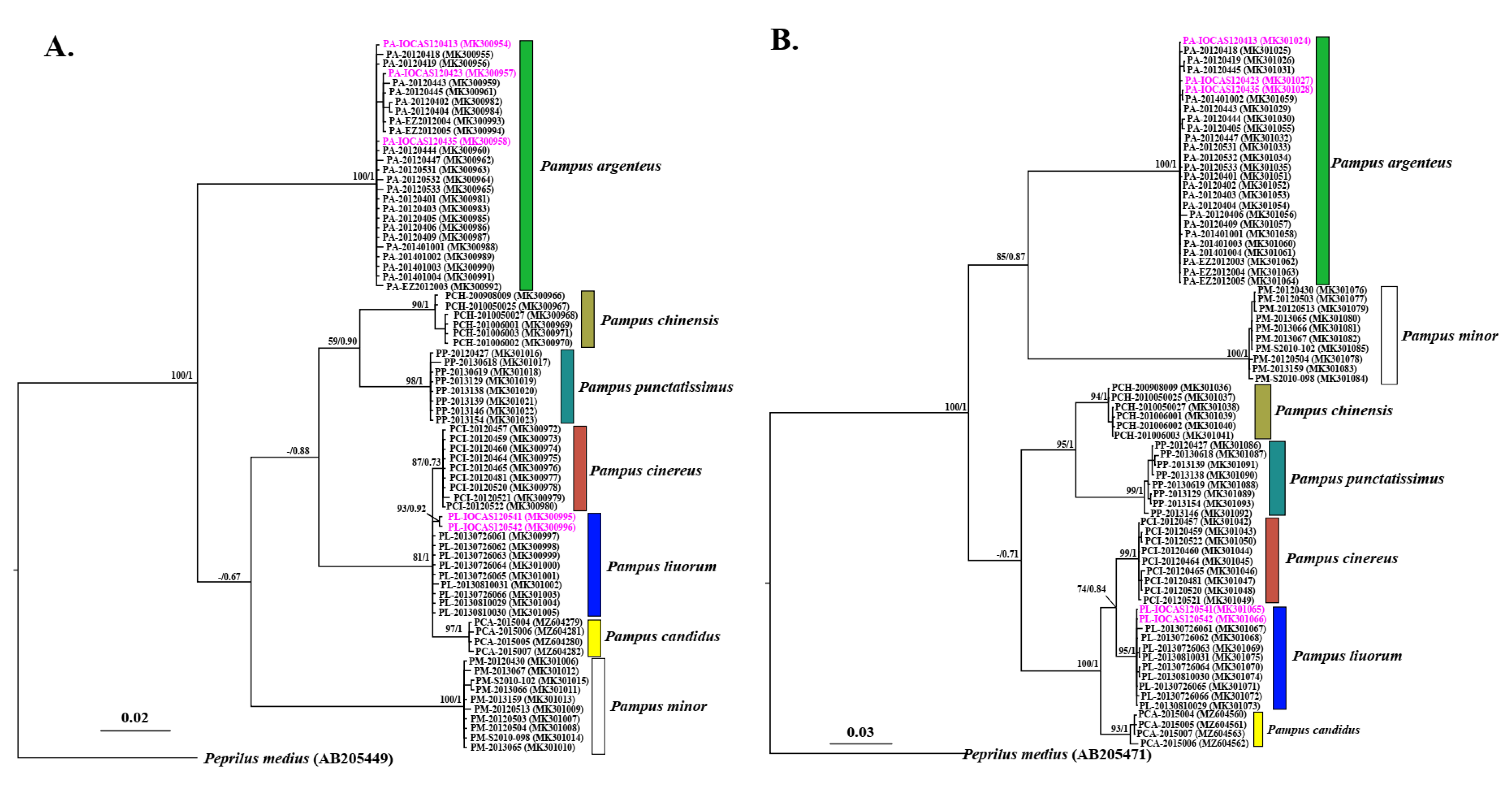
3.2. Phylogenetic Inference
3.3. Species Delimitation
4. Discussion
4.1. Pampus cinereus, Pampus liuorum, and Pampus candidus as Distinct Valid Species
4.2. Species Delimitation and Validity of Pampus argenteus
4.3. Verification of COI and Cytb as Potential DNA Barcodes for Pomfret Identification
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fricke, R.; Eschmeyer, W.N.; Van der Laan, R. Eschmeyer’s Catalog of Fishes: Genera, Species, References. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 17 July 2021).
- Liu, J.; Li, C. A new species of the genus Pampus (Perciformes, Stromateidae) from China. Acta Zootaxonomica Sin. 2013, 38, 885–890. [Google Scholar]
- Liu, J.; Li, C.S.; Niu, P. Identity of silver pomfret Pampus argenteus (Euphrasen, 1788) based on specimens from its type locality, with a neotype designation (Teleostei, Stromateidae). Acta Zootaxonomica Sin. 2013, 38, 171–177. [Google Scholar]
- Liu, J.; Li, C. A new pomfret species, Pampus minor sp. nov. (Stromateidae) from Chinese waters. Chin. J. Oceanol. Limnol. 1998, 16, 280–285. [Google Scholar]
- Liu, J.; Li, C. Redescription of a stromateid fish Pampus punctatissimus and comparison with Pampus argenteus from Chinese coastal waters. Chin. J. Oceanol. Limnol. 1998, 16, 161–166. [Google Scholar]
- Liu, J.; Li, C.; Ning, P. A redescription of grey pomfret Pampus cinereus (Bloch, 1795) with the designation of a neotype (Teleostei: Stromateidae). Chin. J. Oceanol. Limnol. 2013, 31, 140–145. [Google Scholar] [CrossRef]
- Yin, G.X.; Pan, Y.L.; Sarker, A.; Baki, M.A.; Kim, J.-K.; Wu, H.L.; Li, C.H. Molecular systematics of Pampus (Perciformes: Stromateidae) based on thousands of nuclear loci using target-gene enrichment. Mol. Phylogenet. Evol. 2019, 140, 106595. [Google Scholar] [CrossRef]
- Jawad, L.A.; Liu, J. Comparative osteology of the axial skeleton of the genus Pampus (Family: Stromateidae, Perciformes). J. Mar. Biol. Assoc. U. K. 2016, 97, 277–287. [Google Scholar] [CrossRef]
- Radhakrishnan, D.P.; Kumar, R.G.; Mohitha, C.; Rajool Shanis, C.; Kinattukara, B.K.; Saidumohammad, B.V.; Gopalakrishnan, A. Resurrection and Re-description of Pampus candidus (Cuvier), Silver Pomfret from the Northern Indian Ocean. Zool. Stud. 2019, 58, e7. [Google Scholar]
- Guo, Y.; Zhao, W.; Gao, H.; Wang, S.; Yu, P.; Yu, H.; Wang, D.; Wang, Q.; Wang, J.; Wang, Z. China Fishery Statistical Yearbook. In Bureau of Fisheries and Fishery Management; The Ministry of Agriculture of the PRC, Ed.; China Agriculture Press: Beijing, China, 2017; p. 143. [Google Scholar]
- Liu, J.; Li, C.; Li, X. Studies on Chinese pomfret fishes of the genus Pampus (Pisces: Stromateidae). Stud. Mar. Sin. 2002, 44, 240–252. [Google Scholar]
- Li, Y.; Zhang, Y.; Gao, T.; Han, Z.; Lin, L.; Zhang, X. Morphological characteristics and DNA barcoding of Pampus echinogaster (Basilewsky, 1855). Acta Oceanol. Sin. 2017, 36, 18–23. [Google Scholar] [CrossRef]
- Guo, E.; Liu, Y.; Liu, J.; Cui, Z. DNA barcoding discriminates Pampus minor (Liu et al., 1998) from Pampus species. Chin. J. Oceanol. Limnol. 2010, 28, 1266–1274. [Google Scholar] [CrossRef]
- Cui, Z.X.; Liu, Y.; Li, C.P.; Chu, K.H. Species delineation in Pampus (Perciformes) and the phylogenetic status of the Stromateoidei based on mitogenomics. Mol. Biol. Rep. 2011, 38, 1103–1114. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, Z.; Song, N.; Gao, T. Characteristics of complete mitogenome of Pampus sp. nov. (Perciformes: Stromateidae). Mitochondrial DNA Part A 2016, 27, 1640–1641. [Google Scholar]
- Radhakrishnan, P.D.; Mohitha, C.; Kumar Rahul, G.; Rajool Shanis, C.P.; Basheer, V.S.; Gopalakrishnan, A. Molecular based phylogenetic species recognition in the genus Pampus (Perciformes: Stromateidae) reveals hidden diversity in the Indian Ocean. Mol. Phylogenet. Evol. 2017, 109, 240–245. [Google Scholar]
- Li, Y.; Zhou, Y.D.; Li, P.F.; Gao, T.X.; Lin, L.S. Species identification and cryptic diversity in Pampus species as inferred from morphological and molecular characteristics. Mar. Biodivers. 2019, 49, 2521–2534. [Google Scholar] [CrossRef]
- Doiuchi, R.; Nakabo, T. Molecular phylogeny of the stromateoid fishes (Teleostei: Perciformes) inferred from mitochondrial DNA sequences and compared with morphology-based hypotheses. Mol. Phylogenet. Evol. 2006, 39, 111–123. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001; ISBN 978-0-87969-576-7. [Google Scholar]
- Ward, R.D.; Zemlak, T.S.; Innes, B.H.; Last, P.R.; Hebert, P.D. DNA barcoding Australia’s fish species. Philos. Trans. R. Soc. B 2005, 360, 1847–1857. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Meier, R.; Zhang, G.Y.; Ali, F. The use of mean instead of smallest interspecific distances exaggerates the size of the "barcoding gap" and leads to misidentification. Syst. Biol. 2008, 57, 809–813. [Google Scholar] [CrossRef]
- Cardoni, S.; Tenchini, R.; Ficulle, I.; Piredda, R.; Simeone, M.C.; Belfiore, C. DNA barcode assessment of Mediterranean mayflies (Ephemeroptera), benchmark data for a regional reference library for rapid biomonitoring of freshwaters. Biochem. Syst. Ecol. 2015, 62, 36–50. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control. 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Hasegawa, M.; Kishino, H.; Yano, T.A. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef]
- Tavare, S. Some probabilistic and statistical problems in the analysis of DNA sequences. Lect. Math. Life Sci. 1986, 17, 57–86. [Google Scholar]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef]
- Pons, J.; Barraclough, T.G.; Gomez-Zurita, J.; Cardoso, A.; Duran, D.P.; Hazell, S.; Kamoun, S.; Sumlin, W.D.; Vogler, A.P. Sequence-based species delimitation for the DNA taxonomy of undescribed insects. Syst. Biol. 2006, 55, 595–609. [Google Scholar] [CrossRef] [Green Version]
- Fontaneto, D.; Herniou, E.A.; Boschetti, C.; Caprioli, M.; Melone, G.; Ricci, C.; Barraclough, T.G. Independently evolving species in asexual bdelloid rotifers. PLoS Biol. 2007, 5, e87. [Google Scholar] [CrossRef]
- Fujisawa, T.; Barraclough, T.G. Delimiting species using single-locus data and the Generalized Mixed Yule Coalescent approach: A revised method and evaluation on simulated data sets. Syst. Biol. 2013, 62, 707–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jukes, T.H.; Cantor, C.R. Evolution of protein molecules. Mamm. Protein Metab. 1969, 3, 21–132. [Google Scholar]
- Bouckaert, R.; Heled, J.; Kuhnert, D.; Vaughan, T.; Wu, C.H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahrens, D.; Fujisawa, T.; Krammer, H.J.; Eberle, J.; Fabrizi, S.; Vogler, A.P. Rarity and Incomplete Sampling in DNA-Based Species Delimitation. Syst. Biol. 2016, 65, 478–494. [Google Scholar] [CrossRef] [Green Version]
- Le Ru, B.P.; Capdevielle-Dulac, C.; Toussaint, E.F.A.; Conlong, D.; Van den Berg, J.; Pallangyo, B.; Ong’amo, G.; Chipabika, G.; Molo, R.; Overholt, W.A.; et al. Integrative taxonomy of Acrapex stem borers (Lepidoptera: Noctuidae: Apameini): Combining morphology and Poisson Tree Process analyses. Invertebr. Syst. 2014, 28, 451–475. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, N.V.; Zemlak, T.S.; Hanner, R.H.; Hebert, P.D. Universal primer cocktails for fish DNA barcoding. Mol. Ecol. Notes 2007, 7, 544–548. [Google Scholar] [CrossRef]
- Ward, R.D. FISH-BOL, a case study for DNA barcodes. In DNA Barcodes; Kress, W.J., Erickson, D.L., Eds.; Humana Press: New York, NY, USA, 2012; pp. 423–439. [Google Scholar]
- Wu, R.; Zhang, H.; Liu, J.; Niu, S.; Xiao, Y.; Chen, Y. DNA barcoding of the family Sparidae along the coast of China and revelation of potential cryptic diversity in the Indo-West Pacific oceans based on COI and 16S rRNA genes. J. Oceanol. Limnol. 2018, 36, 1753–1770. [Google Scholar] [CrossRef]
- Becker, S.; Hanner, R.; Steinke, D. Five years of FISH-BOL: Brief status report. Mitochondrial DNA Part A 2011, 22, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Rathipriya, A.; Karal Marx, K.; Jeyashakila, R. Molecular identification and phylogenetic relationship of flying fishes of Tamil Nadu coast for fishery management purposes. Mitochondrial DNA Part A 2019, 30, 500–510. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, D.C.; Palhares, R.M.; Drummond, M.G.; Frigo, T.B. DNA Barcoding identification of commercialized seafood in South Brazil: A governmental regulatory forensic program. Food Control 2015, 50, 784–788. [Google Scholar] [CrossRef]
- Zhang, J.B. Species identification of marine fishes in china with DNA barcoding. Evid. Based Complement. Altern. Med. 2011, 2011, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-H.; Qin, G.; Zhang, H.-X.; Wang, X.; Lin, Q. DNA barcoding reflects the diversity and variety of brooding traits of fish species in the family Syngnathidae along China’s coast. Fish. Res. 2017, 185, 137–144. [Google Scholar] [CrossRef]
- Xu, L.; Wang, X.; Van Damme, K.; Huang, D.; Li, Y.; Wang, L.; Ning, J.; Du, F. Assessment of fish diversity in the South China Sea using DNA taxonomy. Fish. Res. 2021, 233, 105771. [Google Scholar] [CrossRef]
- Shen, Y.; Guan, L.; Wang, D.; Gan, X. DNA barcoding and evaluation of genetic diversity in Cyprinidae fish in the midstream of the Yangtze River. Ecol. Evol. 2016, 6, 2702–2713. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.Y.; Zhao, S.; Yu, Z.C.; Zhou, Y.J.; Yang, J.Y.; Zhao, R.; Yang, C.X.; Ma, W.W.; Wang, X.; Feng, M.X.; et al. Application of DNA barcoding in fish identification of supermarkets in Henan province, China: More and longer COI gene sequences were obtained by designing new primers. Food Res. Int. 2020, 136, 109516. [Google Scholar] [CrossRef] [PubMed]
- Lemer, S.; Aurelle, D.; Vigliola, L.; Durand, J.D.; Borsa, P. Cytochrome b barcoding, molecular systematics and geographic differentiation in rabbitfishes (Siganidae). Comptes Rendus Biol. 2007, 330, 86–94. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Hanner, R. Molecular approach to the identification of fish in the South China Sea. PLoS ONE 2012, 7, e30621. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Sampling Date | Sampling Location (Number of Specimens) | BOLD Specimen Voucher | GenBank Accession Number | ||
|---|---|---|---|---|---|---|
| Museum ID | Photo Reference | COI | Cytb | |||
| Pampus argenteus | April 2012 | Zhuhai, Guangdong, China (3) | PA-IOCAS120413 | ✓ | MK300954 | MK301024 |
| PA-IOCAS120423 | ✓ | MK300957 | MK301027 | |||
| PA-IOCAS120435 | ✓ | MK300958 | MK301028 | |||
| April–May 2012 | Shenzhen, Guangdong, China (6) | PA-20120418 | ✓ | MK300955 | MK301025 | |
| PA-20120419 | ✓ | MK300956 | MK301026 | |||
| PA-20120443 | ✓ | MK300959 | MK301029 | |||
| PA-20120444 | ✓ | MK300960 | MK301030 | |||
| PA-20120445 | ✓ | MK300961 | MK301031 | |||
| PA-20120447 | ✓ | MK300962 | MK301032 | |||
| May 2012 | Zhanjiang, Guangdong, China (3) | PA-20120531 | ✓ | MK300963 | MK301033 | |
| PA-20120532 | ✓ | MK300964 | MK301034 | |||
| PA-20120533 | ✓ | MK300965 | MK301035 | |||
| January 2014 | Weihai, Shandong, China (4) | PA-201401001 | MK300988 | MK301058 | ||
| PA-201401002 | MK300989 | MK301059 | ||||
| PA-201401003 | MK300990 | MK301060 | ||||
| PA-201401004 | MK300991 | MK301061 | ||||
| April 2012 | Qingdao, Shandong, China (7) | PA-20120401 | ✓ | MK300981 | MK301051 | |
| PA-20120402 | ✓ | MK300982 | MK301052 | |||
| PA-20120403 | ✓ | MK300983 | MK301053 | |||
| PA-20120404 | ✓ | MK300984 | MK301054 | |||
| PA-20120405 | MK300985 | MK301055 | ||||
| PA-20120406 | MK300986 | MK301056 | ||||
| PA-20120409 | MK300987 | MK301057 | ||||
| May 2012 | Zhoushan, Zhejiang, China (3) | PA-EZ2012003 | ✓ | MK300992 | MK301062 | |
| PA-EZ2012004 | ✓ | MK300993 | MK301063 | |||
| PA-EZ2012005 | ✓ | MK300994 | MK301064 | |||
| Pampus candidus | January 2015 | Iraq (4) | PCA-2015004 | ✓ | MZ604279 | MZ604560 |
| PCA-2015005 | MZ604280 | MZ604561 | ||||
| PCA-2015006 | MZ604281 | MZ604562 | ||||
| PCA-2015007 | MZ604282 | MZ604563 | ||||
| Pampus chinensis | August 2009 | Xiamen, Fujian, China (1) | PCH-200908009 | ✓ | MK300966 | MK301036 |
| May 2010 | Zhuhai, Guangdong, China (5) | PCH-2010050025 | ✓ | MK301037 | MK300967 | |
| PCH-2010050027 | ✓ | MK301038 | MK300968 | |||
| PCH-201006001 | ✓ | MK301039 | MK300969 | |||
| PCH-201006002 | ✓ | MK301040 | MK300970 | |||
| PCH-201006003 | ✓ | MK301041 | MK300971 | |||
| Pampus cinereus | April–May 2012 | Shenzhen, Guangdong, China (3) | PCI-20120457 | ✓ | MK300972 | MK301042 |
| PCI-20120459 | ✓ | MK300973 | MK301043 | |||
| PCI-20120460 | ✓ | MK300974 | MK301044 | |||
| April–May 2012 | Zhuhai, Guangdong, China (3) | PCI-20120464 | ✓ | MK300975 | MK301045 | |
| PCI-20120465 | ✓ | MK300976 | MK301046 | |||
| PCI-20120481 | ✓ | MK300977 | MK301047 | |||
| May 2012 | Zhanjiang, Guangdong, China (3) | PCI-20120520 | ✓ | MK300978 | MK301048 | |
| PCI-20120521 | ✓ | MK300979 | MK301049 | |||
| PCI-20120522 | ✓ | MK300980 | MK301050 | |||
| Pampus liuorum | May 2012 | Zhuhai, Guangdong, China (2) | PL-IOCAS120541 | ✓ | MK300995 | MK301065 |
| PL-IOCAS120542 | ✓ | MK300996 | MK301066 | |||
| July–August 2013 | Dongshan, Fujian, China (9) | PL-20130726061 | MK300997 | MK301067 | ||
| PL-20130726062 | MK300998 | MK301068 | ||||
| PL-20130726063 | MK300999 | MK301069 | ||||
| PL-20130726064 | MK301000 | MK301070 | ||||
| PL-20130726065 | MK301001 | MK301071 | ||||
| PL-20130810031 | MK301002 | MK301072 | ||||
| PL-20130726066 | MK301003 | MK301073 | ||||
| PL-20130810029 | MK301004 | MK301074 | ||||
| PL-20130810030 | MK301005 | MK301075 | ||||
| Pampus minor | October 2013 | Zhoushan, Zhejiang, China (1) | PM-2013159 | MK301013 | MK301083 | |
| April 2012 | Shenzhen, Guangdong, China (1) | PM-20120430 | ✓ | MK301006 | MK301076 | |
| May 2010 | Zhuhai, Guangdong, China (2) | PM-S20-098 | MK301014 | MK301084 | ||
| PM-S20-102 | ✓ | MK301015 | MK301085 | |||
| May 2012 | Zhanjiang, Guangdong, China (3) | PM-20120503 | ✓ | MK301007 | MK301077 | |
| PM-20120504 | ✓ | MK301008 | MK301078 | |||
| PM-20120513 | ✓ | MK301009 | MK301079 | |||
| April 2013 | Beihai, Guangxi, China (3) | PM-2013065 | MK301010 | MK301080 | ||
| PM-2013066 | MK301011 | MK301081 | ||||
| PM-2013067 | MK301012 | MK301082 | ||||
| Pampus punctatissimus | June 2013 | Zhoushan, Zhejiang, China (2) | PP-20130618 | ✓ | MK301017 | MK301087 |
| PP-20130619 | ✓ | MK301018 | MK301088 | |||
| October 2013 | Xiamen, Fujian, China (5) | PP-2013129 | ✓ | MK301019 | MK301089 | |
| PP-2013138 | MK301020 | MK301090 | ||||
| PP-2013139 | MK301021 | MK301091 | ||||
| PP-2013146 | MK301022 | MK301092 | ||||
| PP-2013154 | MK301023 | MK301093 | ||||
| April 2012 | Zhuhai, Guangdong, China (1) | PP-20120427 | ✓ | MK301016 | MK301086 | |
| Species | COI | Cytb | ||||
|---|---|---|---|---|---|---|
| Interspecific | Intraspecific | Barcoding Gap | Interspecific | Intraspecific | Barcoding Gap | |
| Pampus argenteus | 0.1273–0.1572 | 0.0000–0.0052 | 0.1221 | 0.1508–0.1809 | 0.0000–0.0056 | 0.1452 |
| Pampus candidus | 0.0139–0.1556 | 0.0000–0.0034 | 0.0105 | 0.0355–0.1849 | 0.0009–0.0065 | 0.0290 |
| Pampus chinensis | 0.0580–0.1823 | 0.0000–0.0034 | 0.0545 | 0.0555–0.1790 | 0.0000–0.0028 | 0.0527 |
| Pampus cinereus | 0.0034–0.1799 | 0.0000–0.0017 | 0.0017 | 0.0237–0.1850 | 0.0000–0.0037 | 0.0200 |
| Pampus liuorum | 0.0034–0.1572 | 0.0000–0.0034 | 0.0000 | 0.0237–0.1811 | 0.0000–0.0037 | 0.0200 |
| Pampus minor | 0.1318–0.1572 | 0.0000–0.0034 | 0.1283 | 0.1698–0.1850 | 0.0000–0.0047 | 0.1651 |
| Pampus punctatissimus | 0.0580–0.1427 | 0.0000–0.0034 | 0.0545 | 0.0555–0.1777 | 0.0000–0.0056 | 0.0499 |
| Overall | 0.0034–0.1823 | 0.0000–0.0052 | −0.0018 | 0.0237–0.1850 | 0.0000–0.0065 | 0.0172 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, J.; Wu, R.; Xiao, Y.; Zhang, H.; Jawad, L.A.; Wang, Y.; Liu, J.; Al-Mukhtar, M.A. Validity of Pampus liuorum Liu & Li, 2013, Revealed by the DNA Barcoding of Pampus Fishes (Perciformes, Stromateidae). Diversity 2021, 13, 618. https://doi.org/10.3390/d13120618
Wei J, Wu R, Xiao Y, Zhang H, Jawad LA, Wang Y, Liu J, Al-Mukhtar MA. Validity of Pampus liuorum Liu & Li, 2013, Revealed by the DNA Barcoding of Pampus Fishes (Perciformes, Stromateidae). Diversity. 2021; 13(12):618. https://doi.org/10.3390/d13120618
Chicago/Turabian StyleWei, Jiehong, Renxie Wu, Yongshuang Xiao, Haoran Zhang, Laith A. Jawad, Yajun Wang, Jing Liu, and Mustafa A. Al-Mukhtar. 2021. "Validity of Pampus liuorum Liu & Li, 2013, Revealed by the DNA Barcoding of Pampus Fishes (Perciformes, Stromateidae)" Diversity 13, no. 12: 618. https://doi.org/10.3390/d13120618
APA StyleWei, J., Wu, R., Xiao, Y., Zhang, H., Jawad, L. A., Wang, Y., Liu, J., & Al-Mukhtar, M. A. (2021). Validity of Pampus liuorum Liu & Li, 2013, Revealed by the DNA Barcoding of Pampus Fishes (Perciformes, Stromateidae). Diversity, 13(12), 618. https://doi.org/10.3390/d13120618