


Comparisons of Zooplankton Community Structure between with- and without- Pesticide Applications on Rice Fields



Abstract
:1. Introduction
2. Materials and Methods
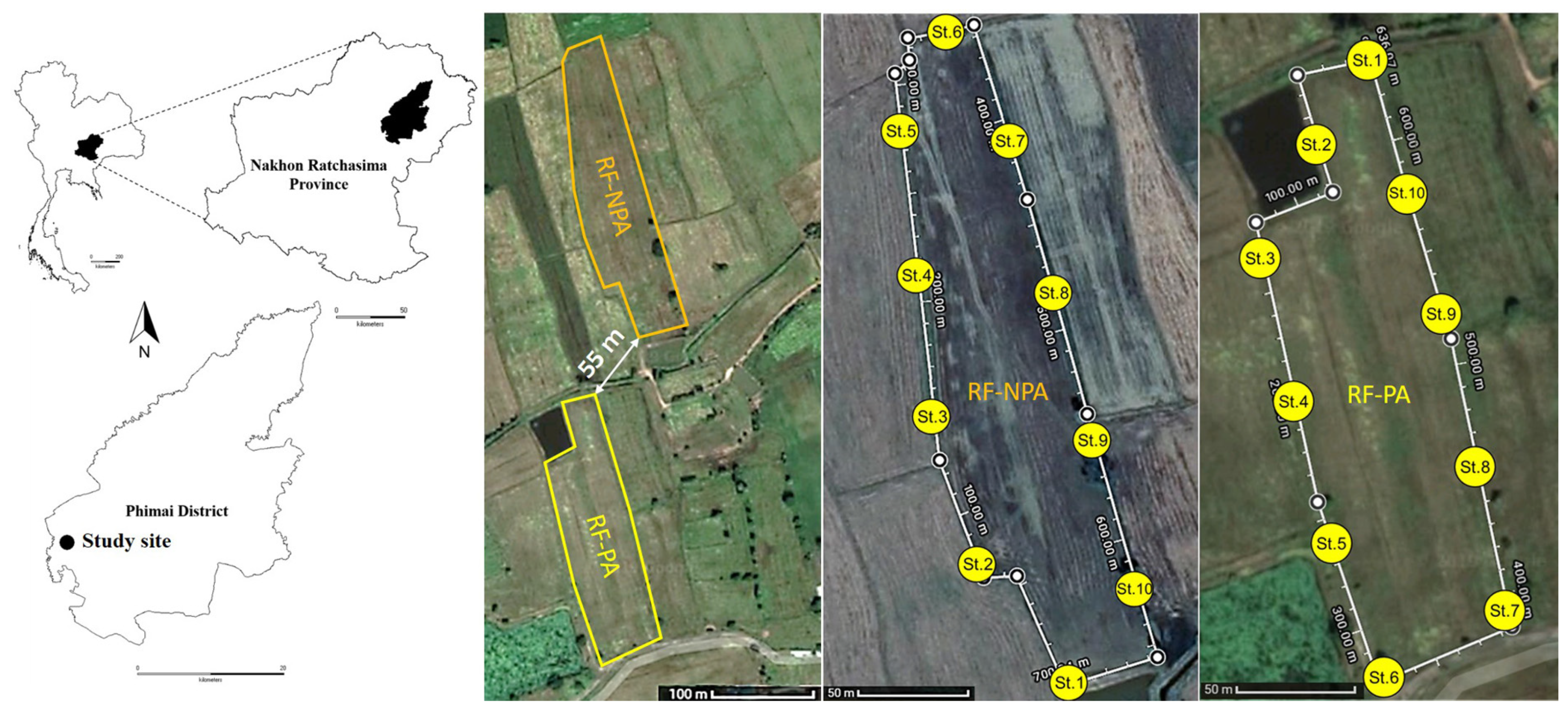
2.1. Study Sites
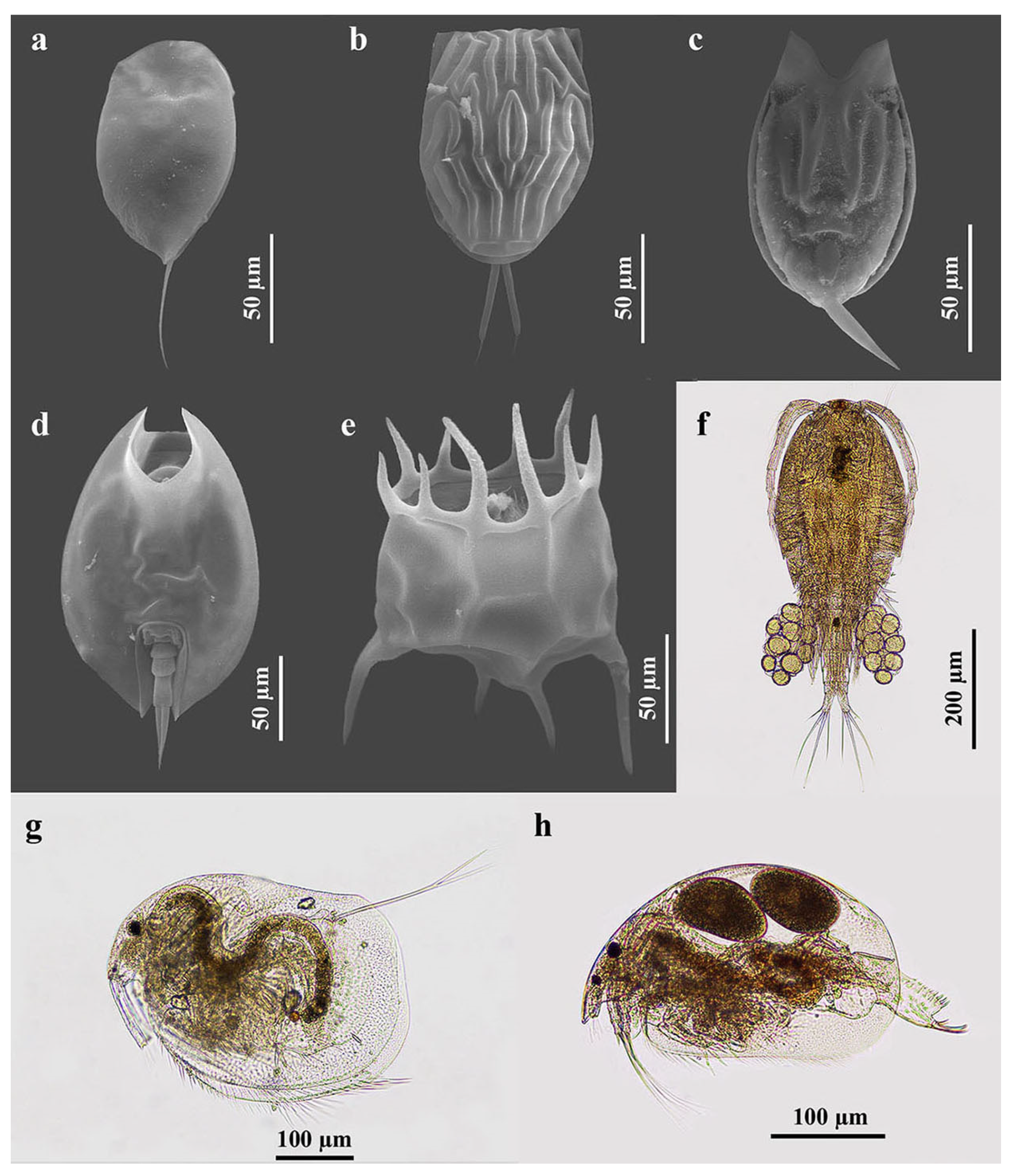
2.2. Zooplankton Sampling and Identification
2.3. Water Quality Measurement
2.4. Data Analysis
3. Results
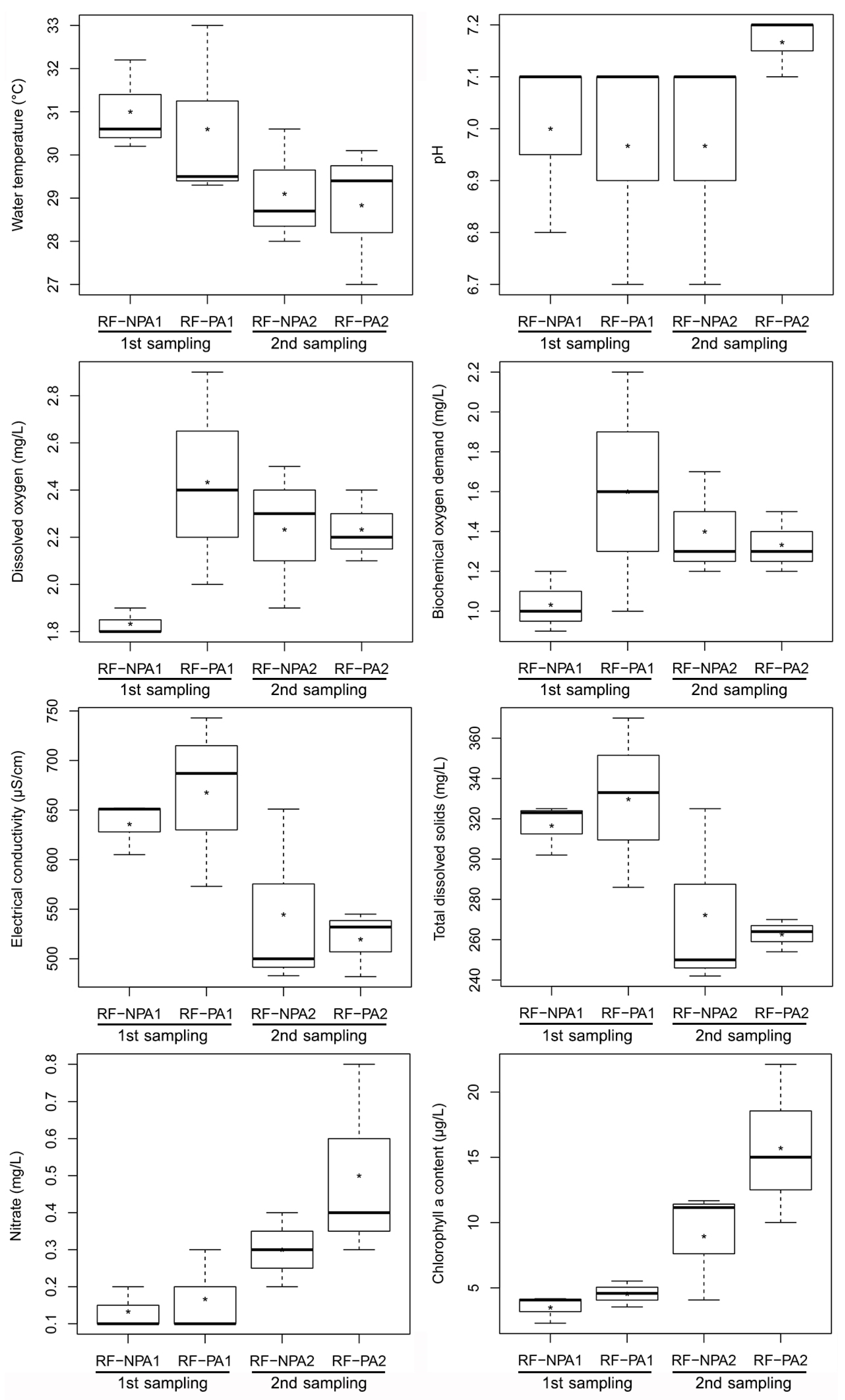
3.1. Environmental Factors of Rice Fields
3.2. Zooplankton Community Structure in RF-NPA and RF-PA
3.2.1. Species Composition
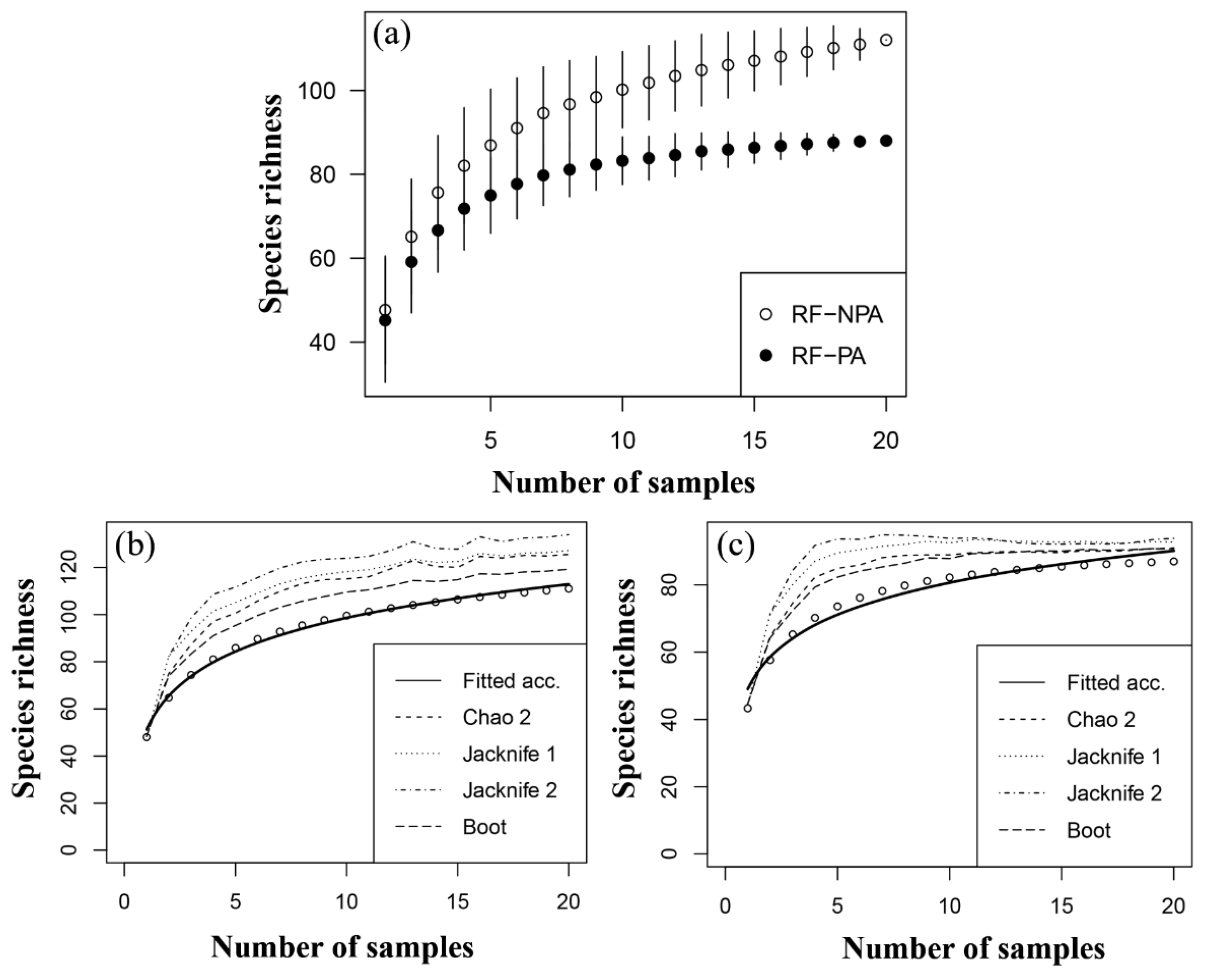
3.2.2. Species Accumulation and Estimate
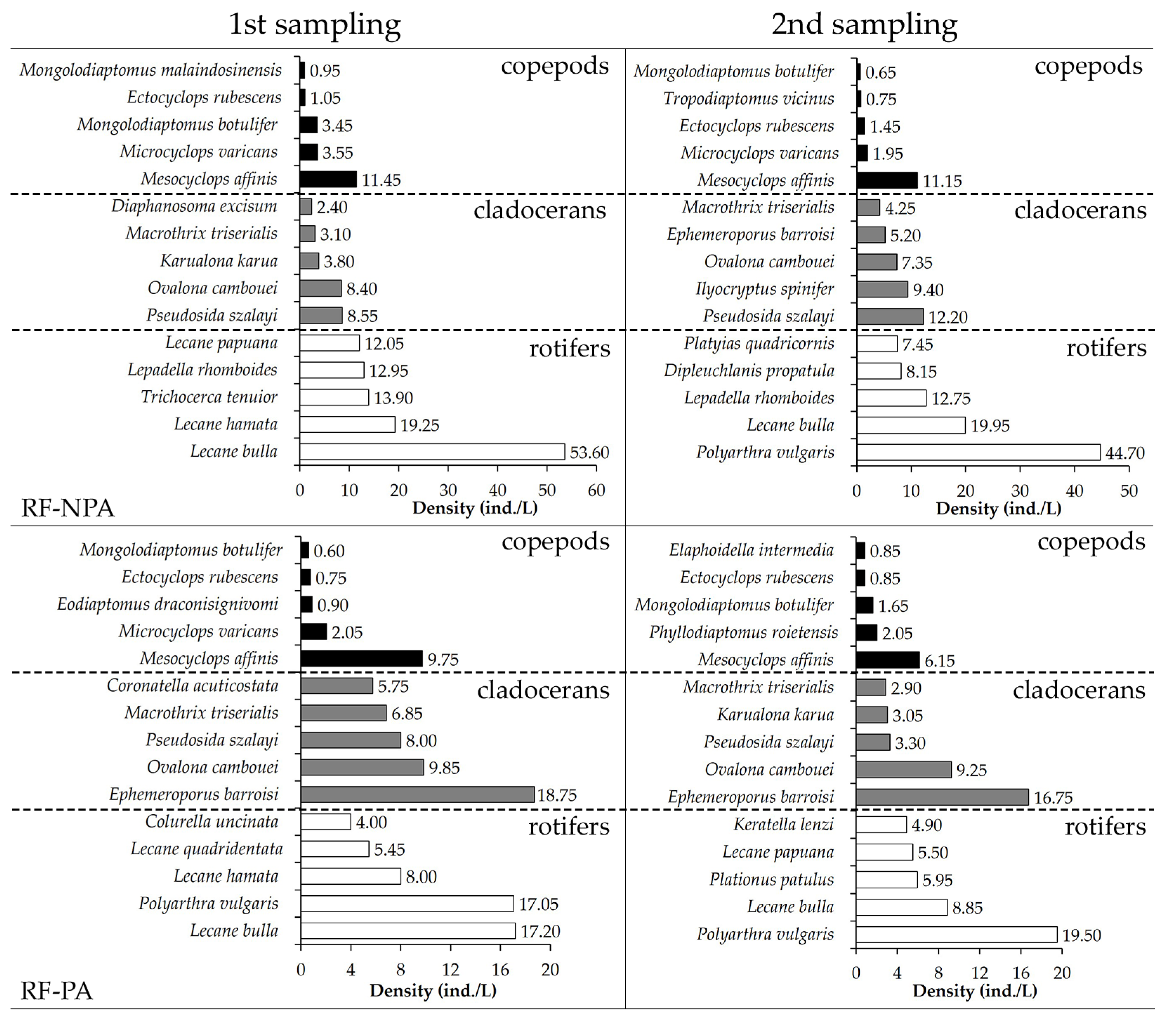
3.2.3. Abundance, Density, and Dominant Species
4. Discussion
4.1. Environmental Factors
4.2. Zooplankton Community Structure in RF-NPA and RF-PA
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salem, H.; Olajos, E.J. Review of pesticides: Chemistry, uses and toxicology. Toxicol. Ind. Health 1988, 4, 291–321. [Google Scholar] [CrossRef]
- Praneetvatakul, S.; Schreinemachers, P.; Pananurak, P.; Tipraqsa, P. Pesticides, external costs and policy options for Thai agriculture. Environ. Sci. Pol. 2013, 27, 103–113. [Google Scholar] [CrossRef]
- Laohaudomchok, W.; Nankongnab, N.; Siriruttanapruk, S.; Klaimala, P.; Lianchamroon, W.; Ousap, P.; Jatiket, M.; Kajitvichyanukul, P.; Kitana, N.; Siriwong, W.; et al. Pesticide use in Thailand: Current situation, health risks, and gaps in research and policy. Hum. Ecol. Risk Assess. 2020, 27, 1147–1169. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, A.R.; Bajet, C.M.; Matin, M.A.; Nhan, D.D.; Sulaiman, A.H. Ecotoxicology of pesticides in the tropical paddy field ecosystem. Environ. Toxicol. Chem. 1997, 16, 59–70. [Google Scholar] [CrossRef]
- Aktar, M.W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Iwai, C.B.; Sujira, H.; Somparn, A.; Komarova, T.; Mueller, J.; Noller, B. Monitoring pesticides in the paddy field ecosystem of North-eastern Thailand for environmental and health risks. In Rational Environmental Management of Agrochemicals: Risk Assessment, Monitoring, and Remedial Action; Kennedy, I., Solomon, K., Gee, S., Crossan, A., Wang, S., Sánchez-Bayo, F., Eds.; American Chemical Society: Washington, DC, USA, 2007; pp. 259–273. [Google Scholar]
- Keister, J.E.; Bonnet, D.; Chiba, S.; Johnson, C.L.; Mackas, D.L.; Escribano, R. Zooplankton population connections, community dynamics, and climate variability. ICES J. Mar. Sci. 2012, 63, 347–350. [Google Scholar] [CrossRef] [Green Version]
- Lawler, S.P. Rice fields as temporary wetlands: A review. Isr. J. Zool. 2001, 47, 513–528. [Google Scholar] [CrossRef]
- Segers, H.; Sanoamuang, L. Note on a highly diverse rotifer assemblage (Rotifera: Monogononta) in a Laotian rice paddy and adjacent pond. Int. Rev. Hydrobiol. 2007, 92, 640–646. [Google Scholar] [CrossRef]
- Ferdous, Z.; Muktadir, A. A review: Potentiality of zooplankton as bioindicator. Am. J. Appl. Sci. 2009, 6, 1815–1819. [Google Scholar] [CrossRef]
- Xiong, W.; Huang, X.; Chen, Y.; Fu, R.; Du, X.; Chen, X.; Zhan, A. Zooplankton biodiversity monitoring in polluted freshwater ecosystems: A technical review. Environ. Sci. Ecotech. 2020, 1, 100008. [Google Scholar] [CrossRef]
- Snell, T.W.; Joaquim-Justo, C. Workshop on rotifers in ecotoxicology. Hydrobiologia 2007, 593, 227–232. [Google Scholar] [CrossRef]
- Vera, M.S.; Di Fiori, E.; Lagomarsino, L.; Sinistro, R.; Escaray, R.; Iummato, M.M.; Juárez, A.; de Molina, M.d.C.R.; Tell, G.; Pizarro, H. Direct and indirect effects of the glyphosate formulation Glifosato Atanor® on freshwater microbial communities. Ecotoxicology 2012, 21, 1805–1816. [Google Scholar] [CrossRef] [PubMed]
- Xiao, P.; Liu, F.; Liu, Y.; Yao, S.; Zhu, G. Effects of pesticide mixtures on zooplankton assemblages in aquatic microcosms simulating rice paddy fields. Bull. Environ. Contam. Toxicol. 2017, 99, 27–32. [Google Scholar] [CrossRef]
- Lim, X.; Koksong, L.; Koksong, L.; Liew, H.; Loh, J.; Loh, J. Acute toxicity of glyphosate on various life stages of calanoid copepod, Pseudodiaptomus annandalei. Asia Pac. J. Mol. Biol. Biotechnol. 2019, 27, 24–31. [Google Scholar] [CrossRef]
- Reimche, G.B.; Machado, S.L.; Oliveira, M.A.; Zanella, R.; Dressler, V.L.; Flores, E.M.; Gonçalves, F.F.; Donato, F.F.; Nunes, M.A. Imazethapyr and imazapic, bispyribac-sodium and penoxsulam: Zooplankton and dissipation in subtropical rice paddy water. Sci. Total Environ. 2015, 514, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Segers, H. Rotifera 2. The Lecanidae (Monogononta). In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; Dumont, H.J., Nogrady, T., Eds.; SPB Academic Publishing: The Hague, The Netherlands, 1995; Volume 6, pp. 1–226. [Google Scholar]
- Smirnov, N.N. Cladocera: The Chydorinae and Sayciinae (Chydoridae) of the world. In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; Dumont, H., Ed.; SPB Academic Publishing: Amsterdam, The Netherlands, 1996; pp. 1–197. [Google Scholar]
- Nogrady, T.; Segers, H. Rotifera 6: Asplanchnidae, Gastropodidae, Lindiidae, Microcodidae, Synchaetidae, Trochosphaeridae and Filinia. In Guides to the Identification of the Microinvertebrates of the Continental Water of the World; Dumont, H., Ed.; Backhuys Publishers: Leiden, The Netherlands, 2002; Volume 18, pp. 1–264. [Google Scholar]
- Sanoamuang, L. Freshwater Zooplankton: Calanoid Copepods in Thailand; Klungnana Vitthaya Press: Khon Kaen, Thailand, 2002; pp. 1–159. [Google Scholar]
- Hołyńska, M. Copepoda: Cyclopoida: Genera Mesocyclops and Thermocyclops. In Guides to the Identification of the Microinvertebrates of the Continental Water of the World; Dumont, H., Ed.; Backhuys Publishers: Leiden, The Netherlands, 2003; Volume 20, pp. 1–318. [Google Scholar]
- Lim, R.; Fernando, C. A review of Malaysian freshwater Copepoda with notes on new records and little known species. Hydrobiologia 1985, 128, 71–89. [Google Scholar] [CrossRef]
- Sinev, A.Y. Key for identification of Cladocera of the subfamily Aloninae (Anomopoda: Chydoridae) from South-East Asia. Zootaxa 2016, 4200, 451–486. [Google Scholar] [CrossRef]
- Winkler, L.W. Die bestimmung des im wasser gelösten sauerstoffes. Ber. Dtsch. Chem. Ges. 1888, 21, 2843–2854. [Google Scholar] [CrossRef] [Green Version]
- APHA-AWWA-WPCF. Standard Methods for Examination of Water and Waste Water; American Public Health Association, American Water Work Association and Water Pollution Control Federation: Washington, DC, USA, 1998; pp. 1–2671. [Google Scholar]
- Chao, A.; Chazdon, R.L.; Colwell, R.K.; Shen, T.J. Abundance-based similarity indices and their estimation when there are unseen species in samples. Biometrics 2006, 62, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Shannon, C.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949; pp. 1–125. [Google Scholar]
- Krebs, C. Ecological Methodology; Addison-Wesley Educational Publishers, Inc.: Menlo Park, CA, USA, 1999; pp. 1–631. [Google Scholar]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Publishing: Oxford, UK, 2004; pp. 1–215. [Google Scholar]
- Avolio, M.L.; Forrestel, E.J.; Chang, C.C.; La Pierre, K.J.; Burghardt, K.T.; Smith, M.D. Demystifying dominant species. New Phytol. 2019, 223, 1106–1126. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.; O’Hara, R.; Simpson, G.; Solymos, P.; et al. Vegan: Community Ecology Package. Available online: https://CRAN.R-project.org/package=vegan (accessed on 6 April 2020).
- Colwell, R.K.; Coddington, J.A. Estimating terrestrial biodiversity through extrapolation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1994, 345, 101–118. [Google Scholar] [PubMed] [Green Version]
- Lautenschlager, R.; Schaertl, G.R. Electrical conductivity of five concentrations of two glyphosate-containing herbicides. S. J. Appl. Forest. 1991, 15, 85–88. [Google Scholar] [CrossRef]
- Giesy, J.P.; Dobson, S.; Solomon, K.R. Ecotoxicological risk assessment for Roundup® herbicide. Rev. Environ. Contam. Toxicol. 2000, 167, 35–120. [Google Scholar]
- Khrueakham, A.; Anurugsa, B.; Hungspreug, N. Influence of chemical fertilizer applications on water quality in paddy fields in Nong Harn, Sakon Nakhon Province, Thailand. Agr. Nat. Resour. 2015, 49, 868–879. [Google Scholar]
- Liu, Y.; Zou, G.; Yuan, Q.; Huang, W.; Zhou, W. Phytoplankton community characteristics in rice paddy fields under different nitrogen fertiliser applications. Acta Physiol. Plant. 2020, 42, 33. [Google Scholar] [CrossRef]
- Zhang, X.; Shen, Y.; Yu, X.-y.; Liu, X.-j. Dissipation of chlorpyrifos and residue analysis in rice, soil and water under paddy field conditions. Ecotoxicol. Environ. Saf. 2012, 78, 276–280. [Google Scholar] [CrossRef]
- Fu, Y.; Liu, F.; Zhao, C.; Zhao, Y.; Liu, Y.; Zhu, G. Distribution of chlorpyrifos in rice paddy environment and its potential dietary risk. J. Environ. Sci. 2015, 35, 101–107. [Google Scholar] [CrossRef]
- Plangklang, N.; Athibai, S. Species diversity and abundance of rotifer fauna in a conventional rice field in Nakhon Ratchasima Province. Khon Kaen Agr. J. 2019, 47, 651–656. [Google Scholar]
- Kotov, A.A.; Dumont, H.J. Analysis of the Ilyocryptus spinifer-species group (Anomopoda, Branchiopoda), with description of a new species. Hydrobiologia 2000, 428, 85–113. [Google Scholar] [CrossRef]
- Korovchinsky, N.M.; Sanoamuang, L. Overview of Sididae (Crustacea: Cladocera: Ctenopoda) of Northeast and East Thailand, with description of a new species of the genus Diaphanosoma. Zootaxa 2008, 1682, 45–61. [Google Scholar] [CrossRef]
- Korovchinsky, N.M. Cladocera (Crustacea: Branchiopoda) of South East Asia: History of exploration, taxon richness and notes on zoogeography. J. Limnol. 2013, 72, 109–124. [Google Scholar] [CrossRef] [Green Version]
- Sanoamuang, L.; Faitakum, S. Species diversity of cladocerans and copepods in the floodplain of the River Mun, Northeast Thailand. Asia-Pac. J. Sci. Technol. 2005, 10, 106–113. [Google Scholar]
- Chittapun, S.; Pholpunthin, P.; Sanoamuang, L. Diversity and composition of zooplankton in rice fields during a crop cycle at Pathum Thani province, Thailand. Songklanakarin J. Sci. Technol. 2009, 31, 261–267. [Google Scholar]
- Sanoamuang, L. Species composition and distribution of freshwater Calanoida and Cyclopoida (Copepoda) of north-east Thailand. In Crustaceans and Biodiversity Crisis; Schram, F., Klein, J., Eds.; Brill Academic Publishers: Leiden, The Netherlands, 1999; pp. 217–230. [Google Scholar]
- Fernando, C.; Furtado, J.; Lim, R. The Aquatic Fauna of the World’s Rice Fields; Department of Zoology, University of Malaya: Kuala Lumpur, Malaysia, 1979; pp. 1–105. [Google Scholar]
- Romero, N.; Attademo, A.M.; Reno, U.; Regaldo, L.; Repetti, M.R.; Lajmanovich, R.; Gagneten, A.M. Analysis of the zooplanktonic community in rice fields during a crop cycle in agroecological versus conventional management. Limnetica 2022, 41. [Google Scholar] [CrossRef]
- Hanazato, T. Pesticide effects on freshwater zooplankton: An ecological perspective. Environ. Pollut. 2001, 112, 1–10. [Google Scholar] [CrossRef]
- Baker, L.F.; Mudge, J.F.; Thompson, D.G.; Houlahan, J.E.; Kidd, K.A. The combined influence of two agricultural contaminants on natural communities of phytoplankton and zooplankton. Ecotoxicology 2016, 25, 1021–1032. [Google Scholar] [CrossRef]
- Degenhardt, D.; Humphries, D.; Cessna, A.J.; Messing, P.; Badiou, P.H.; Raina, R.; Farenhorst, A.; Pennock, D.J. Dissipation of glyphosate and aminomethylphosphonic acid in water and sediment of two Canadian prairie wetlands. J. Environ. Sci. Health B 2012, 47, 631–639. [Google Scholar] [CrossRef]
- Goldsborough, L.G.; Brown, D.J. Dissipation of glyphosate and aminomethylphosphonic acid in water and sediments of boreal forest ponds. Environ. Toxicol. Chem. 1993, 12, 1139–1147. [Google Scholar] [CrossRef]
- Reno, U.; Doyle, S.R.; Momo, F.R.; Regaldo, L.; Gagneten, A.M. Effects of glyphosate formulations on the population dynamics of two freshwater cladoceran species. Ecotoxicology 2018, 27, 784–793. [Google Scholar] [CrossRef]
- Portinho, J.L.; Nielsen, D.L.; Daré, L.; Henry, R.; Oliveira, R.C.; Branco, C.C. Mixture of commercial herbicides based on 2, 4-D and glyphosate mixture can suppress the emergence of zooplankton from sediments. Chemosphere 2018, 203, 151–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, R.; Abdullah, M.; Fernando, C. Ecological studies of Cladocera in the ricefields of Tanjung Karang, Malaysia, subjected to pesticide treatment. Hydrobiologia 1984, 113, 99–103. [Google Scholar] [CrossRef]
- Fantón, N.; Bacchetta, C.; Rossi, A.; Gutierrez, M.F. Effects of a glyphosate-based herbicide on the development and biochemical biomarkers of the freshwater copepod Notodiaptomus carteri (Lowndes, 1934). Ecotoxicol. Environ. Saf. 2020, 196, 110501. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Name | ||
|---|---|---|
| Rotifers | L. pyriformis (Daday, 1905) 2 | Tripleuchlanis plicata (Levander, 1894) 1,2 |
| Anuraeopsis fissa Gosse, 1851 2 | L. quadridentata (Ehrenberg, 1830) 1,2,3,4 | Cladocerans |
| Asplanchnopus multiceps (Schrank, 1793) 1,2,3,4 | L. signifera (Jennings, 1896) 2,3 | Anthalona harti van Damme, Sinev & Dumont 2011 1,2,3,4 |
| Beauchampiella eudactylota (Gosse, 1886) 1,2,3,4 | L. stenroosi (Meissner, 1908) 3 | |
| Brachionus angularis Gosse, 1851 2 | L. tenuiseta Harring, 1914 1,2,3,4 | Chydorus eurynotus Sars, 1901 2,3,4 |
| B. quadridentatus Hermann, 1783 1,2,3,4 | L. thienemanni (Hauer, 1938) 1,2 | Coronatella acuticostata (Sars, 1903) 1,2,3,4 |
| B. quadridentatus mirabilis Daday, 1897 1,2,3 | L. unguitata (Fadeev, 1926) 1,2,3,4 | C. rectangular (Sars, 1861) 3 |
| Cephalodella forficula (Ehrenberg, 1830) 1,2,4 | Lepadella akrobeles Myers, 1934 1 | Dadaya macrops (Daday, 1898) 3 |
| C. gibba (Ehrenberg, 1830) 1,2,3,4 | L. apsicora Myers, 1834 1 | Diaphanosoma excisum Sars, 1885 1 |
| Colurella obtusa (Gosse, 1886) 1,2,3 | L. apsida Harring, 1916 1 | D. senegal Gauthier, 1951 4 |
| C. sanoamuangae Chittapun, Pholpunthin & Segers, 1999 1,2 | L. costatoides Segers, 1992 1,2,3 | Dunhevedia crassa King, 1853 1 |
| L. discoidea Segers, 1993 1,2,3,4 | Ephemeroporus barroisi (Richard, 1894) 1,2,3,4 | |
| C. uncinata (Müller, 1773) 1,2,3,4 | L. ovalis (Müller, 1786) 1,2,4 | Euryalona orientalis (Daday, 1898) 2 |
| Dipleuchlanis propatula (Gosse, 1886) 1,2,3,4 | L. patella (Müller, 1773) 1,2,3,4 | Grimaldina brazzai Richard, 1892 1,2 |
| Euchlanis incisa Carlin, 1939 2,3,4 | L. rhomboides (Gosse, 1886) 1,2,3,4 | Ilyocryptus spinifer Herrick, 1882 1,2,3,4 |
| Hexarthra mira (Hudson, 1871) 2 | L. triba Myers, 1934 1 | Karualona karua (King, 1853) 1,2,3,4 |
| Keratella cochlearis (Gosse, 1851) 2,3 | L. triptera (Ehrenberg, 1832) 1,2 | Kurzia brevilabris (Rajapaksa & Fernando 1986) 1,2,3 |
| K. lenzi Hauer, 1953 2,3,4 | L. voigti Hauer, 1931 1,2,4 | Latonopsis australis Sars, 1888 3 |
| K. tecta (Gosse, 1851) 2 | Lophocharis salpina (Ehrenberg, 1834) 1,2,3,4 | Leberis diaphanus (King, 1853) 1,2 |
| K. tropica (Apstein, 1907) 2,3 | Macrochaetus danneelae Koste & Shiel, 1983 3,4 | Leydigia acanthocercoides (Fischer, 1854) 1 |
| Lecane arcula Harring, 1914 1,2 | Monommata grandis Tessin, 1890 1,2 | Macrothrix spinosa King, 1853 1,2 |
| L. baimaii Sanoamuang & Savatenalinton, 1999 1,2 | M. longiseta (Müller, 1786) 1,2,3,4 | M. triserialis Brady, 1886 1,2,3,4 |
| Mytilina bisulcata (Lucks, 1912) 1,2 | Moina micrura Kurz, 1874 1 | |
| L. bulla (Gosse, 1851) 1,2,3,4 | M. trigona (Gosse, 1851) 1,2,4 | Moinodaphnia macleayi (King, 1853) 1,2,3,4 |
| L. closterocerca (Schmarda, 1859) 1,2,3,4 | M. unguipes (Lucks, 1912) 1,2 | Ovalona cambouei (Guerne & Richard, 1983) 1,2,3,4 |
| L. crepida Harring, 1914 1,2,3,4 | M. ventralis (Ehrenberg, 1830) 1,2,3,4 | Pseudosida szalayi Daday, 1898 1,2,3,4 |
| L. curvicornis (Murray, 1913) 1,2,3,4 | Plationus patulus (Müller, 1786) 1,2,3,4 | Scapholeberis kingi Sars, 1903 1,2,3,4 |
| L. elegans Harring, 1914 1,2 | Platyias leloupi Gillard, 1957 1,2,3 | Simocephalus serrulatus (Koch, 1841) 1,2,3,4 |
| L. haliclysta Harring & Myers, 1926 3 | P. quadricornis (Ehrenberg, 1832) 1,2,3,4 | Copepods |
| L. hamata (Stokes, 1896) 1,2,3,4 | Polyarthra vulgaris Carlin, 1943 1,2,3,4 | Ectocyclops rubescens (Brady, 1904) 1,2,3,4 |
| L. hastata (Murray, 1913) 1,2,3 | Scaridium longicaudum (Müller, 1786) 1,2,3,4 | Elaphoidella intermedia Chappuis, 1931 1,2,3,4 |
| L. hornemanni (Ehrenberg, 1834) 1,3 | Sinantherina spinosa (Thorpe, 1893) 1,2,3,4 | Eodiaptomus draconisignivomi Brehm, 1952 1,2,3 |
| L. inopinata Harring & Myers, 1926 1,2,3,4 | Squatinella lamellaris (Müller, 1786) 2 | Heliodiaptomus elegans Kiefer, 1935 1,2,3 |
| L. lateralis Sharma, 1978 1,2,3,4 | Testudinella ahlstromi Hauer, 1956 1,3 | Mesocyclops affinis Van de Velde, 1987 1,2,3,4 |
| L. leontina (Turner, 1892) 1,2,3,4 | T. greeni Koste, 1981 1,2,3,4 | Microcyclops varicans (Sars, 1863) 1,2,3,4 |
| L. ludwigii (Eckstein, 1883) 1,2 | T. patina (Hermann, 1783) 1,2,3,4 | Mongolodiaptomus botulifer (Kiefer, 1974) 1,2 |
| L. luna (Müller, 1776) 1,2,3,4 | Trichocerca bicristata (Gosse, 1887) 1,2,3,4 | M. malaindosinensis (Lai & Fernando, 1978) 1,2,3,4 |
| L. lunaris (Ehrenberg, 1832) 1,2,4 | T. capucina (Wierzejski & Zacharias, 1893) 1 | Neodiaptomus yangtsekiangensis Mashiko, 1951 1,2 |
| L. monostyla (Daday, 1897) 3 | T. pusilla (Jennings, 1903) 2 | Phyllodiaptomus praedictus Dumont & Ranga Reddy, 1994) 1,2,3,4 |
| L. nitida (Murray, 1913) 3 | T. similis (Wierzejski, 1893) 1,4 | |
| L. obtusa (Murray, 1913) 2,3 | T. tenuior (Gosse, 1886) 1,2,3,4 | P. roietensis Sanoamuang & Watiroyram, 2020 1,2,3,4 |
| L. papuana (Murray, 1913) 1,2,3,4 | T. weberi (Jennings, 1903) 1,3 | Tropodiaptomus vicinus (Kiefer, 1930) 1,2,3,4 |
| L. pusilla Harring, 1914 1,4 | Trichotria tetractis (Ehrenberg, 1830) 1,2,3,4 | |
| Zooplankton | RF | Species Richness | Total Abundance | H′ | J |
|---|---|---|---|---|---|
| Rotifers | 1st NPA | 66 | 3874 | 2.87 | 0.68 |
| 2nd NPA | 68 | 3481 | 3.01 | 0.72 | |
| 1st PA | 55 | 1996 | 2.74 | 0.68 | |
| 2nd PA | 45 | 1952 | 2.72 | 0.71 | |
| Cladocerans | 1st NPA | 19 | 675 | 2.18 | 0.74 |
| 2nd NPA | 17 | 930 | 2.07 | 0.73 | |
| 1st PA | 16 | 1277 | 2.12 | 0.76 | |
| 2nd PA | 13 | 859 | 1.88 | 0.73 | |
| Copepods | 1st NPA | 12 | 457 | 1.62 | 0.65 |
| 2nd NPA | 12 | 352 | 1.38 | 0.55 | |
| 1st PA | 10 | 314 | 1.39 | 0.60 | |
| 2nd PA | 8 | 251 | 1.56 | 0.75 | |
| Total taxa | 1st NPA | 97 | 5006 | 3.35 | 0.73 |
| 2nd NPA | 97 | 4763 | 3.45 | 0.75 | |
| 1st PA | 81 | 3587 | 3.15 | 0.71 | |
| 2nd PA | 66 | 3062 | 3.09 | 0.70 |
| Zooplankton | RF-NPA | p Value | RF-PA | p Value | ||
|---|---|---|---|---|---|---|
| 1st | 2nd | 1st | 2nd | |||
| Rotifers | ||||||
| Average density (ind./L) | 19.37 ± 4.17 | 17.40 ± 8.15 | 0.333 | 9.98 ± 2.28 | 9.76 ± 3.03 | 0.878 |
| Relative density (%) | 63.98−84.63 | 51.30−89.51 | 49.47−62.82 | 52.60−77.32 | ||
| Cladocerans | ||||||
| Average density (ind./L) | 3.38 ± 1.09 | 4.65 ± 1.03 | 0.053 | 6.39 ± 17 | 4.30 ± 1.36 | 0.001 * |
| Relative density (%) | 10.00−17.93 | 8.05−31.61 | 32.79−37.22 | 15.46−38.06 | ||
| Copepods Adult | ||||||
| Average density (ind./L) | 2.29 ± 1.62 | 1.76 ± 1.25 | 0.202 | 1.57 ± 0.58 | 1.26 ± 0.56 | 0.126 |
| Relative density (%) | 3.90−24.36 | 2.44−17.10 | 4.39−14.21 | 4.95−15.85 | ||
| Copepodite Average density (ind./L) Nauplii | 2.41 ± 1.09 | 2.48 ± 1.24 | 0.374 | 2.05 ± 0.71 | 1.73 ± 0.52 | 0.137 |
| Average density (ind./L) | 7.44 ± 1.85 | 6.52 ± 2.59 | 0.386 | 14.26 ± 4.18 | 10.74 ± 2.65 | 0.043 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Plangklang, N.; Athibai, S. Comparisons of Zooplankton Community Structure between with- and without- Pesticide Applications on Rice Fields. Diversity 2021, 13, 644. https://doi.org/10.3390/d13120644
Plangklang N, Athibai S. Comparisons of Zooplankton Community Structure between with- and without- Pesticide Applications on Rice Fields. Diversity. 2021; 13(12):644. https://doi.org/10.3390/d13120644
Chicago/Turabian StylePlangklang, Nattaporn, and Sujeephon Athibai. 2021. "Comparisons of Zooplankton Community Structure between with- and without- Pesticide Applications on Rice Fields" Diversity 13, no. 12: 644. https://doi.org/10.3390/d13120644
APA StylePlangklang, N., & Athibai, S. (2021). Comparisons of Zooplankton Community Structure between with- and without- Pesticide Applications on Rice Fields. Diversity, 13(12), 644. https://doi.org/10.3390/d13120644