
Sex and Age Effects on Monk Parakeet Home-Range Variation in the Urban Habitat



Abstract
:1. Introduction
2. Materials and Methods
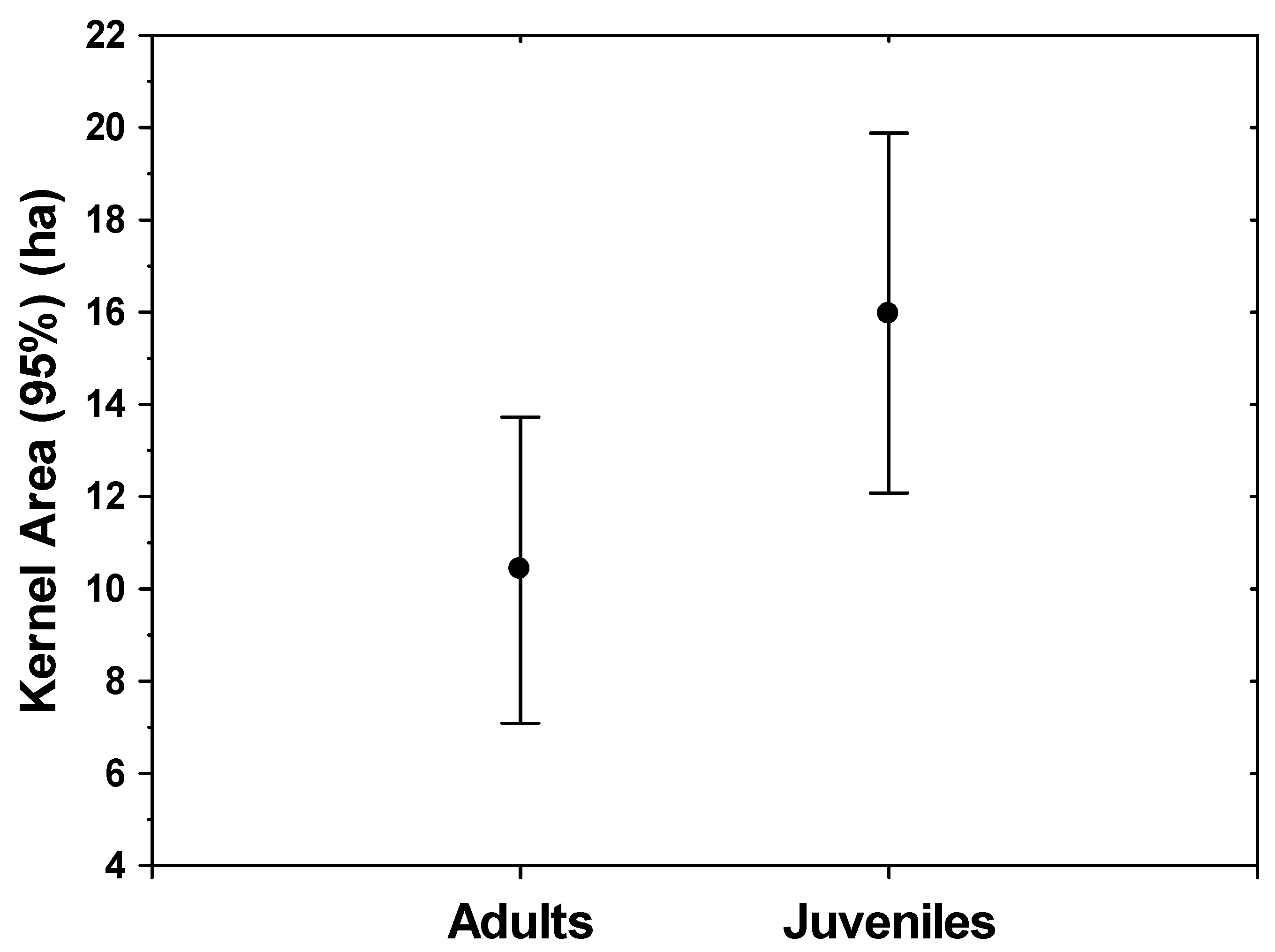
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nathan, R.; Getz, W.M.; Revilla, E.; Holyoak, M.; Kadmon, R.; Saltz, D.; Smouse, P.E. A movement ecology paradigm for unifying organismal movement research. Proc. Natl. Acad. Sci. USA 2008, 105, 19052–19059. [Google Scholar] [CrossRef] [Green Version]
- Holyoak, M.; Casagrandi, R.; Nathan, R.; Revilla, E.; Spiegel, O. Trends and missing parts in the study of movement ecology. Proc. Natl. Acad. Sci. USA 2008, 105, 19060–19065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellison, N.; Hatchwell, B.J.; Biddiscombe, S.J.; Napper, C.J.; Potts, J.R. Mechanistic home range analysis reveals drivers of space use patterns for a non-territorial passerine. J. Anim. Ecol. 2020, 89, 2763–2776. [Google Scholar] [CrossRef] [PubMed]
- Börger, L.; Dalziel, B.D.; Fryxell, J.M. Are there general mechanisms of animal home range behaviour? A review and prospects for future research. Ecol. Lett. 2008, 11, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Clobert, J.; Danchin, E.; Dhondt, A.A.; Nichols, J. Dispersal; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Hansson, L.-A.; Åkesson, S. Animal Movement Across Scales, 1st ed.; Oxford University Press: Oxford, UK, 2014; ISBN 9780199677184. [Google Scholar]
- Rolando, A. On the Ecology of Home Range in Birds. Revue d’écologie 2002, 57, 53–73. [Google Scholar]
- Adams, A.L.; Recio, M.R.; Robertson, B.C.; Dickinson, K.J.M.; van Heezik, Y. Understanding home range behaviour and resource selection of invasive common brushtail possums (Trichosurus vulpecula) in urban environments. Biol. Invasions 2014, 16, 1791–1804. [Google Scholar] [CrossRef]
- Cameron, B.G.; van Heezik, Y.; Maloney, R.F.; Seddon, P.J.; Harraway, J.A. Improving predator capture rates: Analysis of river margin trap site data in the Waitaki Basin, New Zealand. N. Z. J. Ecol. 2005, 29, 117–128. [Google Scholar]
- Norbury, G.L.; Norbury, D.C.; Heyward, R.P. Space use and denning behaviour of wild ferrets (Mustela furo) and cats (Felis catus). N. Z. J. Ecol. 1998, 22, 149–159. [Google Scholar]
- Bengsen, A.J.; Butler, J.A.; Masters, P. Applying home-range and landscape-use data to design effective feral-cat control programs. Wildl. Res. 2012, 39, 258. [Google Scholar] [CrossRef]
- Moseby, K.E.; Stott, J.; Crisp, H. Movement patterns of feral predators in an arid environment–implications for control through poison baiting. Wildl. Res. 2009, 36, 422. [Google Scholar] [CrossRef]
- Croft, S.; Aegerter, J.N.; Beatham, S.; Coats, J.; Massei, G. A spatially explicit population model to compare management using culling and fertility control to reduce numbers of grey squirrels. Ecol. Model. 2021, 440, 109386. [Google Scholar] [CrossRef]
- Suppo, C.; Naulin, J.M.; Langlais, M.; Artois, M. A modelling approach to vaccination and contraception programmes for rabies control in fox populations. Proc. Biol. Sci. 2000, 267, 1575–1582. [Google Scholar] [CrossRef]
- Olah, G.; Butchart, S.H.M.; Symes, A.; Guzmán, I.M.; Cunningham, R.; Brightsmith, D.J.; Heinsohn, R. Ecological and socio-economic factors affecting extinction risk in parrots. Biodivers. Conserv. 2016, 25, 205–223. [Google Scholar] [CrossRef]
- Bucher, E.H. Neotropical parrots as agricultural pests. In New World Parrots in Crisis, Solutions from Conservation Biology; Beissinger, S.R., Snyder, N., Eds.; Smithsonian Intitution Press: Washington, DC, USA, 1992; pp. 201–219. [Google Scholar]
- Menchetti, M.; Mori, E. Worldwide impact of alien parrots (Aves Psittaciformes) on native biodiversity and environment: A review. Ethol. Ecol. Evol. 2014, 26, 172–194. [Google Scholar] [CrossRef]
- Lindsey, G.D.; Arendt, W.J.; Kalina, J.; Pnedelton, G.W. Home ranges and movements of juvenile Puerto Rican Parrots. J. Wildl. Manag. 1991, 55, 318–322. [Google Scholar] [CrossRef]
- Strubbe, D.; Matthysen, E. A radiotelemetry study of habitat use by the exotic Ring-necked Parakeet Psittacula krameri in Belgium. Ibis 2011, 153, 180–184. [Google Scholar] [CrossRef]
- Brightsmith, D.J.; Boyd, J.D.; Hobson, E.A.; Randel, C.J. Satellite telemetry reveals complex migratory movement patterns of two large macaw species in the western Amazon basin. Avian Conserv. Ecol. 2021, 16, 14. [Google Scholar] [CrossRef]
- Ortiz-Maciel, S.G.; Hori-Ochoa, C.; Enkerlin-Hoeflich, E. Maroon-Fronted Parrot (Rhynchopsitta terrisi) Breeding Home Range and Habitat Selection in the Northern Sierra Madre Oriental, Mexico. Wilson J. Ornithol. 2010, 122, 513–517. [Google Scholar] [CrossRef]
- Stahala, C. Seasonal movements of the Bahama parrot (Amazona leucocephala bahamensis) between pine and hardwood forests: Implications for habitat conservation. Ornitol. Neotrop. 2008, 19, 165–171. [Google Scholar]
- Forshaw, J.M. Parrots of the World; Princeton University Press: Princeton, NJ, USA, 2010. [Google Scholar]
- Pruett-Jones, S. Naturalized Parrots of the World; Princeton University Press: Princeton, NJ, USA, 2021. [Google Scholar]
- Senar, J.C.; Carrillo-Ortiz, J.; Arroyo, L. Numbered neck collars for long-distance identification of parakeets. J. Field Ornithol. 2012, 83, 180–185. [Google Scholar] [CrossRef]
- Postigo, J.L.; Strubbe, D.; Mori, E.; Ancillotto, L.; Carneiro, I.; Latsoudis, P.; Menchetti, M.; Pârâu, L.G.; Parrott, D.; Reino, L.; et al. Mediterranean versus Atlantic monk parakeets Myiopsitta monachus: Towards differentiated management at the European scale. Pest Manag. Sci. 2019, 75, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Dawson-Pell, F.S.E.; Senar, J.C.; Hatchwell, B.J. Fine-scale genetic structure reflects limited and coordinated dispersal in the colonial monk parakeet, Myiopsitta monachus. Mol. Ecol. 2021, 30, 1531–1544. [Google Scholar] [CrossRef] [PubMed]
- Dawson-Pell, F.S.E.; Hatchwell, B.J.; Ortega-Segalerva, A.; Dawson, D.A.; Horsburgh, G.J.; Senar, J.C. Microsatellite characterisation and sex-typing in two invasive parakeet species, the monk parakeet Myiopsitta monachus and ring-necked parakeet Psittacula krameri. Mol. Biol. Rep. 2020, 47, 1543–1550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Pastor, R.; Senar, J.C.; Ortega, A.; Faus, J.; Uribe, F.; Montalvo, T. Distribution patterns of invasive Monk parakeets (Myiopsitta monachus) in an urban habitat. Anim. Biodiv. Conserv. 2012, 35, 107–117. [Google Scholar] [CrossRef]
- Carrillo-Ortiz, J. Dinámica de poblaciones de la cotorra de pecho gris (Myiopsitta monachus) en la ciudad de Barcelona. Ph.D. Thesis, University of Barcelona, Barcelona, Spain, 2009. [Google Scholar]
- Conroy, M.J.; Senar, J.C. Integration of demographic analyses and decision modeling in support of management of invasive monk parakeets, an urban and agricultural pest. Environ. Ecol. Stat. 2009, 3, 491–510. [Google Scholar]
- Navarro, J.L.; Martín, L.F.; Bucher, E.H. El uso de la muda de remiges para determinar clases de edad en la cotorra (Myiopsitta monachus). El Hornero 1992, 13, 261–262. [Google Scholar]
- Senar, J.C.; Carrillo-Ortiz, J.; Ortega-Segalerva, A.; Dawson-Pell, F.S.E.; Pascual, J.; Arroyo, L.; Mazzoni, D.; Montalvo, T.; Hatchwell, B.J. The reproductive capacity of monk parakeets Myiopsitta monachus is higher in their invasive range. Bird Study 2019, 66, 136–140. [Google Scholar] [CrossRef]
- Mori, E.; Pascual, J.; Fattorini, N.; Menchetti, M.; Montalvo, T.; Senar, J.C. Ectoparasite sharing among native and invasive birds in a metropolitan area. Parasitol. Res. 2019, 118, 399–409. [Google Scholar] [CrossRef]
- Kenward, R.E.; Casey, N.M.; Walls, S.S.; South, A.B. Ranges 9: For the Analysis of Tracking and Location Data, Online Manual; Anatrack Ltd.: Wareham, UK, 2014. [Google Scholar]
- Worton, B.J. Kernel methods for estimating the utilization distribution in home-range studies. Ecology 1989, 70, 164–168. [Google Scholar] [CrossRef]
- White, G.C.; Garrott, R.A. Analysis of Wildlife Radio-Tracking Data; Elsevier: New York, NY, USA, 1990. [Google Scholar]
- Osborne, J.W. Best Practices in Data Cleaning: A Complete Guide to Everything You Need to Do before and after Collecting Your Data; SAGE: Thousand Oaks, CA, USA, 2013; ISBN 1412988012. [Google Scholar]
- Erickson, B.H.; Nosanchuk, T. Understanding Data, 2nd ed.; Open University Press: Buckingham, UK, 2009; ISBN 9780335096626. [Google Scholar]
- Warnock, S.E.; Takekawa, J.Y. Wintering site fidelity and movement patterns of Western Sandpipers Calidris mauri in the San Francisco Bay estuary. Ibis 1996, 138, 160–167. [Google Scholar] [CrossRef]
- Adams, E.S. Approaches to the study of territory size and shape. Annu. Rev. Ecol. Syst. 2001, 32, 277–303. [Google Scholar] [CrossRef]
- Lameris, T.K.; Brown, J.S.; Kleyheeg, E.; Jansen, P.A.; van Langevelde, F. Nest defensibility decreases home-range size in central place foragers. Behav. Ecol. 2018, 29, 1038–1045. [Google Scholar] [CrossRef]
- White, T.H.; Collazo, J.A.; Vilella, F.J.; Guerrer, S.A. Effects of Hurricane Georges on habitat use by captive-reared Hispaniolan Parrots (Amazona ventralis) released in the Dominican Republic. Ornitol. Neotrop. 2005, 16, 405–417. [Google Scholar]
- Borray-Escalante, N.A.; Mazzoni, D.; Ortega-Segalerva, A.; Arroyo, L.; Morera-Pujol, V.; González-Solís, J.; Senar, J.C. Diet assessments as a tool to control invasive species: Comparison between Monk and Rose-ringed parakeets with stable isotopes. J. Urban Ecol. 2020, 6, juaa005. [Google Scholar] [CrossRef] [Green Version]
- Spreyer, M.F.; Bucher, E.H. Monk Parakeet (Myiopsitta monachus). In The Birds of North America; Poole, A., Gill, F., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 1998; pp. 1–23. [Google Scholar]
- Larson, G.E. The Monk Parakeet in Illinois: New Views of Alarm. Ill. Audubon Bull. 1973, 166, 29–30. [Google Scholar]
- Brooks, D.M. Behavioral ecology of a blue-crowned parakeet (Aratinga acuticaudata) in a subtropical urban landscape far from its natural range. Bull. Texas Ornithol. Soc. 2009, 42, 78–82. [Google Scholar]
- White, J.G.; Gubiani, R.; Smallman, N.; Snell, K.; Morton, A. Home range, habitat selection and diet of foxes (Vulpes vulpes) in a semi-urban riparian environment. Wildl. Res. 2006, 33, 175. [Google Scholar] [CrossRef] [Green Version]
- Prange, S.; Gehrt, S.D.; Wiggers, E.P. Influences of anthropogenic resources on raccoon (Procyon lotor) movements and spatial distribution. J. Mammal. 2004, 85, 483–490. [Google Scholar] [CrossRef]
- Senar, J.C.; Conroy, M.J.; Montalvo, T. Decision-making Models and Management of the Monk Parakeet. In Naturalized Parrots of the World; Pruett-Jones, S., Ed.; Princeton University Press: Princeton, NJ, USA, 2021; pp. 102–122. [Google Scholar]
- Bomford, M.; O’Brien, P. Eradication or control for vertebrate pests? Wildl. Soc. Bull. 1995, 23, 249–255. [Google Scholar]
- Rainbolt, R.E.; Coblentz, B.E. A different perspective on eradication of vertebrate pests. Wildl. Soc. Bull. 1997, 25, 189–191. [Google Scholar]
- Murton, R.K.; Thearle, R.J.P.; Thompson, J. Ecological studies of the feral pigeon Columba livia var.: I. population, breeding biology and methods of control. J. Appl. Ecol. 1972, 9, 835–874. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| F | Df | P | |
|---|---|---|---|
| Sex | 0.005 | 1.69 | 0.95 |
| Age | 4.50 | 1.69 | 0.04 |
| Sex x Age | 0.008 | 1.69 | 0.93 |
| F | Df | P | |
|---|---|---|---|
| Sex | 0.65 | 1.69 | 0.42 |
| Age | 1.12 | 1.69 | 0.29 |
| Sex x Age | 0.11 | 1.69 | 0.74 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Senar, J.C.; Moyà, A.; Pujol, J.; Tomas, X.; Hatchwell, B.J. Sex and Age Effects on Monk Parakeet Home-Range Variation in the Urban Habitat. Diversity 2021, 13, 648. https://doi.org/10.3390/d13120648
Senar JC, Moyà A, Pujol J, Tomas X, Hatchwell BJ. Sex and Age Effects on Monk Parakeet Home-Range Variation in the Urban Habitat. Diversity. 2021; 13(12):648. https://doi.org/10.3390/d13120648
Chicago/Turabian StyleSenar, Juan Carlos, Aura Moyà, Jorge Pujol, Xavier Tomas, and Ben J. Hatchwell. 2021. "Sex and Age Effects on Monk Parakeet Home-Range Variation in the Urban Habitat" Diversity 13, no. 12: 648. https://doi.org/10.3390/d13120648
APA StyleSenar, J. C., Moyà, A., Pujol, J., Tomas, X., & Hatchwell, B. J. (2021). Sex and Age Effects on Monk Parakeet Home-Range Variation in the Urban Habitat. Diversity, 13(12), 648. https://doi.org/10.3390/d13120648