
Microbiome Changes of Endemic Lake Baikal Sponges during Bleaching Syndrome Development

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Analysis of Essential Parameters and Taxonomic Hit Distribution
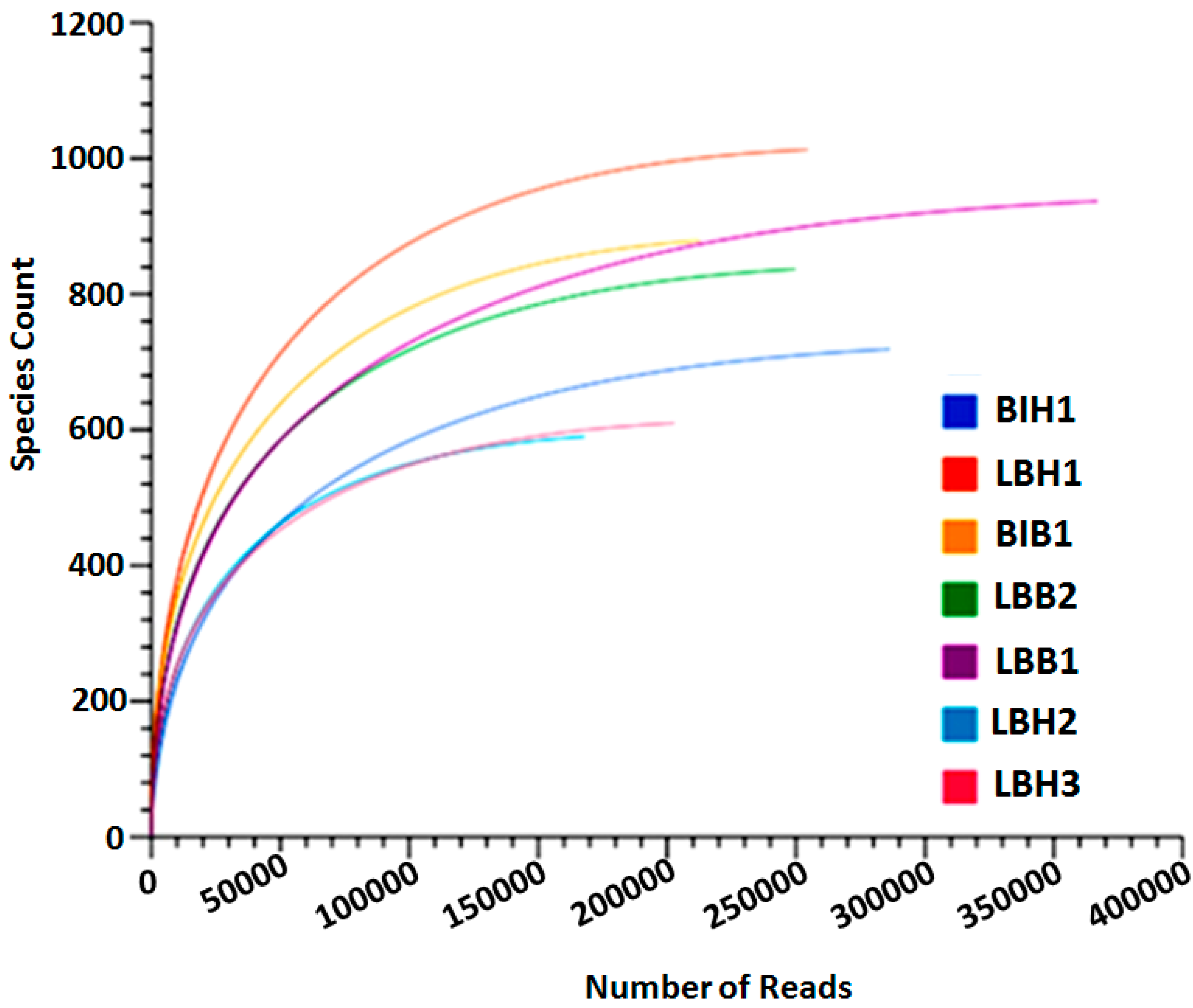
3.2. Analysis of Rarefaction Curve
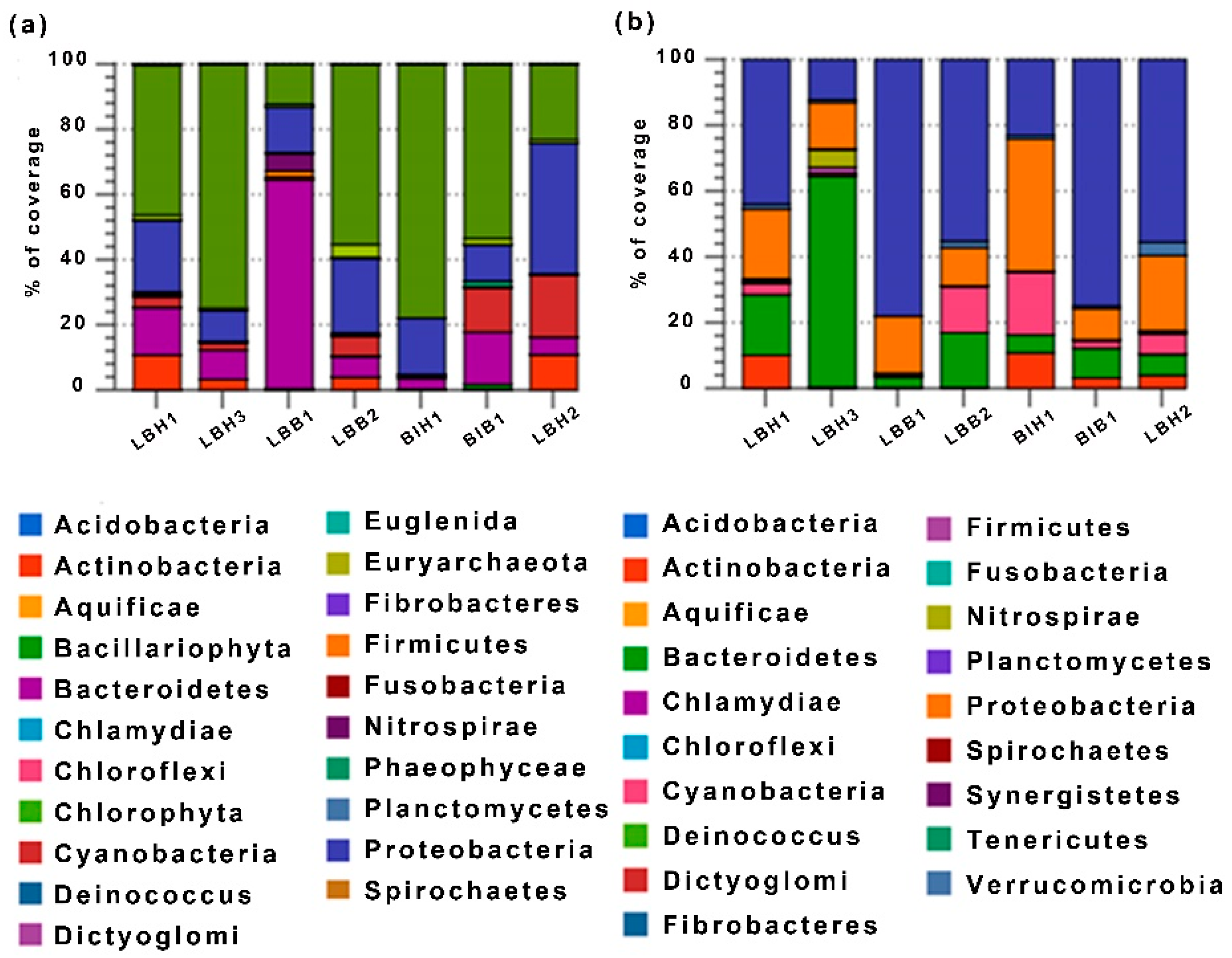
3.3. Analysis of Relative Abundances of OTUs at Genus Level
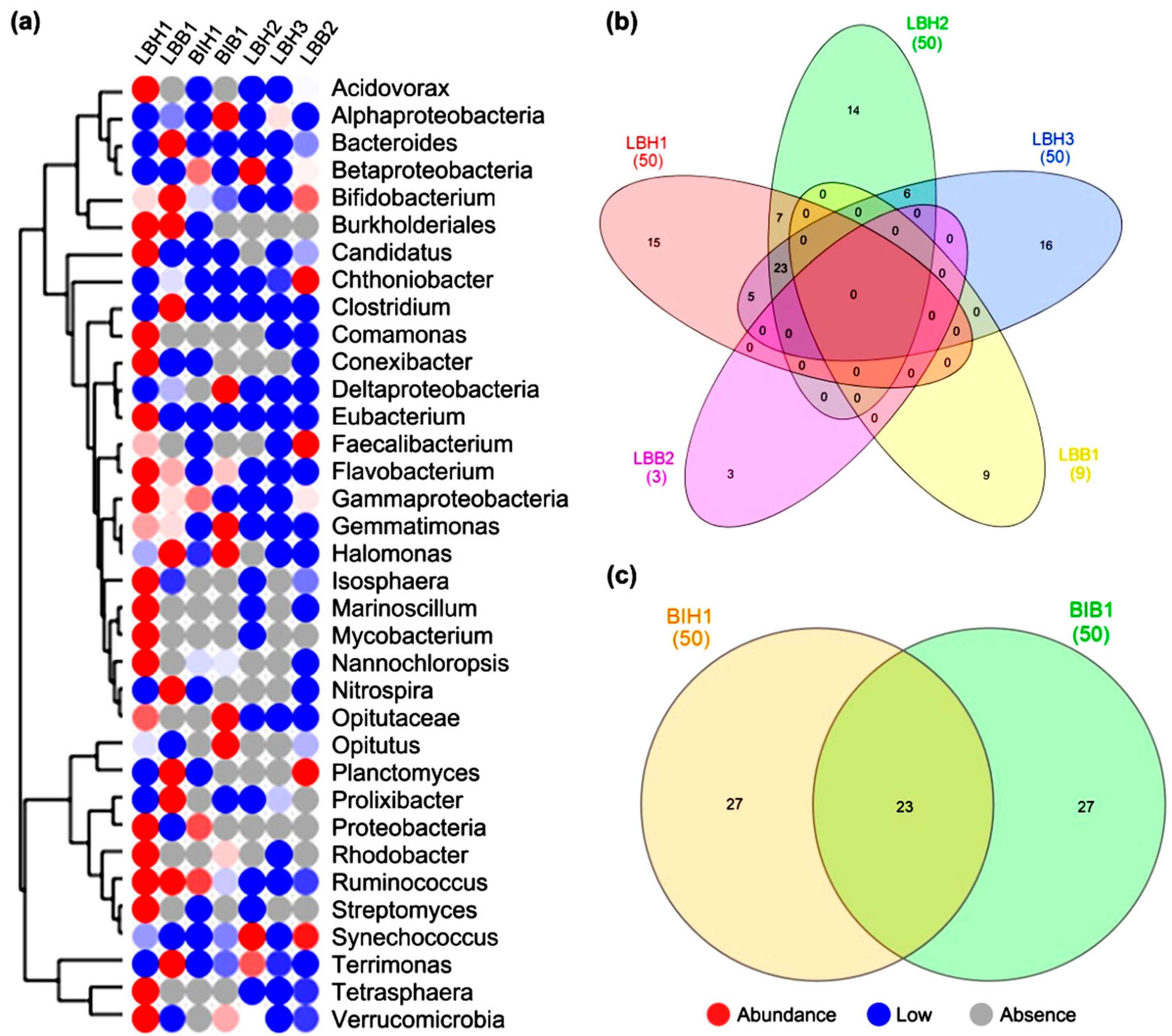
3.4. Analysis of OTUs within Genus Level
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Diaz, M.C.; Rützler, K. Sponges: An essential component of Caribbean coral reefs. Bull. Mar. Sci. 2001, 69, 535–546. [Google Scholar]
- Bell, J.J.; Biggerstaff, A.; Bates, T.; Bennett, H.; Marlow, J.; McGrath, E.; Shaffer, M. Sponge monitoring: Moving beyond diversityand abundance measures. Ecol. Indic. 2017, 78, 470–488. [Google Scholar] [CrossRef]
- Bell, J.J. The functional roles of marine sponges. Estuar. Coast. Shelf Sci. 2008, 79, 341–352. [Google Scholar] [CrossRef]
- Van Soest, R.W.M.; Nicole Boury-Esnault, J.V.; Dohrmann, M.; Erpenbeck, D.; De Voogd, N.J.; Santodomingo, N.; Vanhoorne, B.; Kelly, M.; Hooper, J.N.; Hooper, A. Global diversity of sponges (Porifera). PLoS ONE 2012, 7, e35105. [Google Scholar] [CrossRef]
- Hentschel, U.; Usher, K.M.; Taylor, M.W. Marine sponges as microbial fermenters. FEMS Microbiol. Ecol. 2006, 55, 167–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webster, N.S.; Blackall, L.L. What do we really know about sponge-microbial symbioses. ISME J. 2009, 3, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cervino, J.M.; Winiarski-Cervino, K.; Poison, S.W.; Goreau, T.; Smith, G.W. Identification of bacteria associated with a disease affecting the marine sponge Ianthella basta in New Britain, Papua New Guinea. Mar. Ecol. Prog. Ser. 2006, 324, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, J.D.; Pramanik, A.; Webster, N.; Llewellyn, L.E.; Gachhui, R.; Mukherjee, J. Assessing the complex sponge microbiota: Core, variable and species-specific bacterial communities in marine sponges. ISME J. 2012, 6, 564–576. [Google Scholar] [CrossRef] [Green Version]
- Hentschel, U.; Piel, J.; Degnan, S.M.; Taylor, M.W. Genomic insights into the marine sponge microbiome. Nat. Rev. Microbiol. 2012, 10, 641–654. [Google Scholar] [CrossRef]
- Webster, N.S.; Thomas, T. The sponge hologenome. MBio 2016, 7, e00135-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, L.; Liu, M.; Simister, R.; Webster, N.S.; Thomas, T. Marine microbial symbiosis heats up: The phylogenetic and functional response of a sponge holobiont to thermal stress. ISME J. 2013, 7, 991–1002. [Google Scholar] [CrossRef]
- Webster, N.S.; Luter, H.M.; Soo, R.M.; Botte, E.S.; Simister, R.L.; Abdo, D.; Whalan, S. Same, same but different: Symbiotic bacterial associations in GBR sponges. Front. Microbiol. 2012, 3, 444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erwin, P.M.; Pita, L.; Lopez-Legentil, S.; Turon, X. Stability of sponge-associated bacteria over large seasonal shifts in temperature and irradiance. Appl. Environ. Microbiol. 2012, 78, 7358–7368. [Google Scholar] [CrossRef] [Green Version]
- Morrow, K.M.; Fiore, C.L.; Lesser, M.P. Environmental drivers of microbial community shifts in the giant barrel sponge, Xestospongia muta, over a shallow to mesophotic depth gradient. Environ. Microbiol. 2016, 18, 2025–2038. [Google Scholar] [CrossRef]
- Selvin, J.; Priya, S.; Seghal Kiran, G.; Thangavelu, T.; Sapna Bai, N. Sponge associated marine bacteria as indicators of heavy metal pollution. Microbiol. Res. 2009, 164, 352–363. [Google Scholar] [CrossRef] [PubMed]
- Bourne, D.G.; Dennis, P.G.; Uthicke, S.; Soo, R.M.; Tyson, G.W.; Webster, N. Coral reef invertebrate microbiomes correlate with the presence of photosymbionts. NPG 2013, 7, 1452–1458. [Google Scholar] [CrossRef]
- Bourne, D.G.; Morrow, K.M.; Webster, N.S. Insights into the coral microbiome: Underpinning the health and resilience of reef ecosystems. Annu. Rev. Microbiol. 2016, 70, 317–340. [Google Scholar] [CrossRef] [PubMed]
- Botté, E.S.; Nielsen, S.; Abdul Wahab, M.A.; Webster, J.; Robbins, S.; Thomas, T.; Webster, N.S. Changes in the metabolic potential of the sponge microbiome under ocean acidification. Nat. Commun. 2019, 10, 4134. [Google Scholar] [CrossRef] [Green Version]
- Kiran, G.S.; Sekar, S.; Ramasamy, P.; Thinesh, T.; Hassan, S.; Lipton, A.N.; Ninawe, A.S.; Selvin, J. Marine sponge microbial association: Towards disclosing unique symbiotic interactions. Mar. Environ. Res. 2018, 140, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Kaluzhnaya, O.V.; Krivich, A.A.; Itskovich, V.B. Diversity of 16S rRNA Genes in Metagenomic Community of the Freshwater Sponge Lubomirskia baicalensis. Russ. J. Genet. 2012, 48, 851–854. [Google Scholar] [CrossRef]
- Kaluzhnaya, O.V.; Itskovich, V.B. Phylogenetic Diversity of Microorganisms Associated with the Deep-Water Sponge Baikalospongia intermedia. Russ. J. Genet. 2014, 50, 667–676. [Google Scholar] [CrossRef]
- Kaluzhnaya, O.V.; Itskovich, V.B. Distinctive features of the microbial diversity and the polyketide synthase genes spectrum in the community of the endemic Baikal sponge Swartschewskia papyracea. Russ. J. Genet. 2016, 52, 38–48. [Google Scholar] [CrossRef]
- Costa, R.; Keller-Costa, T.; Gomes, N.C.; da Rocha, U.N.; van Overbeek, L.; van Elsas, J.D. Evidence for selective bacterial Communitystructuring in the freshwater sponge Ephydatia fluviatilis. Microb. Ecol. 2013, 65, 232–244. [Google Scholar] [CrossRef]
- Gladkikh, A.S.; Kaluyzhnaya, O.V.; Belykh, O.I.; Ahn, T.S.; Parfenova, V.V. Analysis of bacterial communities of two Lake Baikal endemic sponge species. Mikrobiologiia 2014, 83, 682–693. [Google Scholar] [CrossRef] [PubMed]
- Gaikwad, S.; Shouche, Y.S.; Gade, W.N. Microbial community structure of two freshwater sponges using Illumina MiSeq sequencing revealed high microbial diversity. AMB Expr. 2016, 6, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, E.Y.; Jung, D.; Belykh, O.I.; Bukshuk, N.A.; Parfenova, V.V.; Joung, Y.; Kim, I.C.; Yim, J.H.; Ahn, T.-S. Comparison of bacterial diversity and species composition in three endemic Baikalian sponges. Ann. Limnol. 2016, 52, 27–32. [Google Scholar] [CrossRef]
- Efremova, S.M. New genus and new species of sponges from family Lubomirskiidae Rezvoj, 1936. In Index of Animal Species Inhabiting Lake Baikal and Its Catchment Area; Timoshkin, O.A., Ed.; Nauka: Lake Baikal, Russia, 2004; Volume 1, pp. 1261–1278. [Google Scholar]
- Bukshuk, N.A.; Maikova, O.O. A new species of Baikal endemic sponges (Porifera, Demospongiae, Spongillida, Lubomirskiidae). ZooKeys 2020, 906, 113–130. [Google Scholar] [CrossRef] [PubMed]
- Itskovich, V.; Kaluzhnaya, O.; Veynberg, Y.; Erpenbeck, D. Endemic Lake Baikal sponges from deep water. 2: Taxonomy and bathymetric distribution. Zootaxa 2017, 4236, 335–342. [Google Scholar] [CrossRef]
- Masuda, Y. Studies on the taxonomy and distribution of freshwater sponges in Lake Baikal. In Biosilica in Evolution, Morphogenesis and Nanobiotechnology; Müller, W.E.G., Grachev, M.A., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 81–110. [Google Scholar] [CrossRef]
- Timoshkin, O.A.; Samsonov, D.P.; Yamamuro, M.; Moore, M.V.; Belykh, O.I.; Malnik, V.V.; Sakirko, M.V.; Shirokaya, A.A.; Bondarenko, N.A.; Domysheva, V.M.; et al. Rapid ecological change in the coastal zone of Lake Baikal (East Siberia): Is the site of the world’s greatest freshwater biodiversity in danger? J. Great Lakes Res. 2016, 42, 487–497. [Google Scholar] [CrossRef] [Green Version]
- Kaluzhnaya, O.V.; Itskovich, V.B. Influence of Baikal sponge bleaching to taxonomic composition of symbiotic microorganisms. Russ. J. Genet. 2015, 51, 1335–1340. [Google Scholar]
- Itskovich, V.B.; Shigarova, A.M.; Glyzina, O.Y.; Kaluzhnaya, O.V.; Borovskii, G.B. Heat shock protein 70 (Hsp70) response to elevated temperatures in the endemic Baikal sponge Lubomirskia baicalensis. Ecol. Indic. 2018, 88, 1–7. [Google Scholar] [CrossRef]
- Khanaev, I.V.; Kravtsova, L.S.; Maikova, O.; Bukshuk, N.; Sakirko, M.V.; Kulakova, N.V.; Butina, T.V.; Nebesnykh, I.A.; Belikov, S.I. Current state of the sponge fauna (Porifera: Lubomirskiidae) of Lake Baikal: Sponge disease and the problem of conservation of diversity. J. Great Lakes Res. 2018, 44, 77–85. [Google Scholar] [CrossRef]
- Kulakova, N.V.; Sakirko, M.V.; Adelshin, R.V.; Khanaev, I.V.; Nebesnykh, I.A.; Perez, T. Brown Rot Syndrome and Changes in the Bacterial community of the Baikal Sponge Lubomirskia baicalensis. Microb. Ecol. 2018, 75, 1024–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belikov, S.; Belkova, N.; Butina, T.; Chernogor, L.; Martynova-Van Kley, A.; Nalian, A.; Rorex, C.; Khanaev, I.I.; Maikova, O.; Feranchuk, S. Diversity and shifts of the bacterial community associated with Baikal sponge mass mortalities. PLoS ONE 2019, 14, e0213926. [Google Scholar] [CrossRef]
- Cebrian, E.; Uriz, M.J.; Garrabou, E.; Ballesteros, E. Sponge mass mortalities in a warming Mediterranean. Are cyanobacteria harboring species worse off? PLoS ONE 2011, 6, e20211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokolow, S. Effects of a changing climate on the dynamics of coral infectious disease: A review of the evidence. Dis. Aquat. Org. 2009, 87, 5–18. [Google Scholar] [CrossRef] [Green Version]
- Ramsby, B.D.; Hoogenboom, M.O.; Whalan, S.; Webster, N.S. Elevated seawater temperature disrupts the microbiome of an ecologically important bioeroding sponge. Mol. Ecol. 2018, 27, 2124–2137. [Google Scholar] [CrossRef] [PubMed]
- Vicente, V.P. Regional commercial sponge extinction in the West Indies: Are recent climatic changes responsible? Mar. Ecol. Prog. Ser. 1989, 10, 179–191. [Google Scholar] [CrossRef]
- Cerrano, C.; Bavestrello, G.; Bianchi, C.N.; Cattaneovietti, R.; Bava, S.; Morganti, C.; Morri, C.; Picco, P.; Sara, G.; Schiaparelli, S.; et al. A catastrophic mass-mortality episode of gorgonians and other organisms in the Ligurian Sea (North-Western Mediterranean), summer 1999. Ecol. Lett. 2000, 3, 284–293. [Google Scholar] [CrossRef]
- Kaluzhnaya, O.; Itskovich, V. Molecular identification of filamentous cyanobacteria overgrowing the endemic sponge Lubomirskia baicalensis. Inland Waters 2017, 7, 267–271. [Google Scholar] [CrossRef]
- Chernogor, L.; Klimenko, E.; Khanaev, I.; Belikov, S. Microbiome analysis of healthy and diseased sponges Lubomirskia Baicalensis by using cell cultures of primmorphs. PeerJ 2019, 7, e27851v1. [Google Scholar] [CrossRef] [PubMed]
- Sorokovikova, E.; Belykh, O.; Krasnopeev, A.; Potapov, S.; Tikhonova, I.; Khanaev, I.; Kabilov, M.; Baturina, O.; Podlesnaya, G.; Timoshkin, O. First data on cyanobacterial biodiversity in benthic biofilms during mass mortality of endemic sponges in Lake Baikal. J. Great Lakes Res. 2020, 46, 75–84. [Google Scholar] [CrossRef]
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T.; Nishijima, M. Development of a prokaryotic universal primer for simultaneous analysis of Bacteria and Archaea using next-generation sequencing. PLoS ONE 2014, 9, e105592. [Google Scholar] [CrossRef] [Green Version]
- Meyer, F.; Paarmann, D.; D’Souza, M.; Olson, R.; Glass, E.M.; Kubal, M.; Paczian, T.; Rodriguez, A.; Stevens, R.; Wilke, A.; et al. The metagenomics RAST server—A public resource for the automatic phylogenetic and functional analysis of metagenomes. BMC Bioinform. 2008, 9, 386. [Google Scholar] [CrossRef] [Green Version]
- Hughes, J.B.; Hellmann, J.J.; Ricketts, T.H.; Bohannan, B.J.M. Counting the uncountable: Statistical approaches to estimating microbial diversity. Appl. Environ. Microbiol. 2001, 67, 4399–4406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schloss, P.D.; Handelsman, J. Introducing DOTUR, a computer program for defining operational taxonomic units and estimating species richness. Appl. Environ. Microbiol. 2005, 71, 1501–1506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prathiviraj, R.; Rajeev, R.; Fernandes, H.; Rathna, K.; Lipton, A.N.; Selvin, J.; Kiran, G.S. A gelatinized lipopeptide diet effectively modulates immune response, disease resistance and gut microbiome in Penaeus vannamei challenged with Vibrio parahaemolyticus. Fish. Shellfish Immunol. 2021, 112, 92–107. [Google Scholar] [CrossRef]
- Schloss, P.D.; Handelsman, J. Toward a census of bacteria in soil. PLoS Comput. Biol. 2006, 2, e92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dattorro, J. Convex Optimization, Euclidean Distance Geometry 2ε; Mεβoo Publishing: Palo Alto, CA, USA, 2015. [Google Scholar]
- Wagner, J.G.; Aghajanian, G.K.; Bing, O.H. Correlation of performance test scores with tissue concentration of lysergic aciddiethylamide in human subjects. CPT 1968, 9, 635–638. [Google Scholar]
- Woebken, D.; Teeling, H.; Wecker, P.; Dumitriu, A.; Kostadinov, I.; Delong, E.F.; Amann, R.; Glцckner, F.O. Fosmids of novel marine Planctomycetes from the Namibian and Oregon coast upwelling systems and their cross_comparison with planctomycete genomes. ISME J. 2007, 1, 419–435. [Google Scholar] [CrossRef] [Green Version]
- Lemoine, N.; Buell, N.; Hill, A.L.; Hill, M.S. Assessing the utility of sponge microbial symbiont communities as models to study global climate change: A case study with Halichondria bowerbanki. In Porifera Research Biodiveristy, Innovation and Sustainability; Custodio, M.R., Lobo-Hajdu, G., Hajdu, E., Muricy, G., Eds.; Museu Nacional: Rio De Janeiro, Brazil, 2007; pp. 419–425. [Google Scholar]
- Webster, N.S.; Soo, R.; Cobb, R.; Negri, A.P. Elevated seawater temperature causes a microbial shift on crustose coralline algae with implications for the recruitment of coral larvae. ISME J. 2011, 5, 759–770. [Google Scholar] [CrossRef] [Green Version]
- Sweet, M.J.; Bulling, M.T. On the importance of the microbiome and pathobiome in coral health and disease. Front. Mar. Sci. 2017, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Legentil, S.; Song, B.; McMurray, S.E.; Pawlik, J.R. Bleaching and stress in coral reef ecosystems: Hsp70 expression by the giant barrel sponge Xestospongia muta. Mol. Ecol. 2008, 17, 1840–1849. [Google Scholar] [CrossRef] [PubMed]
- Bourne, D.; Iida, Y.; Uthicke, S.; Smith-Keune, C. Changes in coral-associated microbial communities during a bleaching event. ISME J. 2008, 2, 350–363. [Google Scholar] [CrossRef]
- Lee, S.T.; Davy, S.K.; Tang, S.L.; Fan, T.-Y.; Kench, P.S. Successive shifts in the microbial community of the surface mucus layer and tissues of the coral Acropora muricata under thermal stress. FEMS Microbiol. Ecol. 2015, 91, fiv142. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | BioProject ID | BioSample ID | Sample Name | Species | Nature of the Sample | Base Pair Count | Seq. Count | Location |
|---|---|---|---|---|---|---|---|---|
| 1 | PRJNA665339 | SAMN16252099 | LBH1 | L. baikalensis | Healthy | 64,423,962 | 254,909 | Bolshie Koty, Baikal, Russia |
| 2 | SAMN16252100 | LBB1 | L. baikalensis | Bleached | 92,784,188 | 367,304 | Bolshie Koty, Baikal, Russia | |
| 3 | SAMN16252101 | BIH1 | B. intermedia | Healthy | 72,441,375 | 286,657 | North Baikal, Russia | |
| 4 | SAMN16252102 | BIB1 | B. intermedia | Bleached | 53,745,195 | 212,772 | North Baikal, Russia | |
| 5 | SAMN16252103 | LBH2 | L. baikalensis | Healthy | 42,470,245 | 168,170 | Bolshie Koty, Baikal, Russia | |
| 6 | SAMN16252104 | LBH3 | L. baikalensis | Healthy | 51,347,284 | 203,100 | Chertov most, South Baikal, Russia | |
| 7 | SAMN16252105 | LBB2 | L. baikalensis | Bleached | 63,233,204 | 250,215 | Chertov most, South Baikal, Russia |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Itskovich, V.; Kaluzhnaya, O.; Glyzina, O.; Prathiviraj, R.; Seghal Kiran, G.; Selvin, J. Microbiome Changes of Endemic Lake Baikal Sponges during Bleaching Syndrome Development. Diversity 2021, 13, 653. https://doi.org/10.3390/d13120653
Itskovich V, Kaluzhnaya O, Glyzina O, Prathiviraj R, Seghal Kiran G, Selvin J. Microbiome Changes of Endemic Lake Baikal Sponges during Bleaching Syndrome Development. Diversity. 2021; 13(12):653. https://doi.org/10.3390/d13120653
Chicago/Turabian StyleItskovich, Valeria, Oxana Kaluzhnaya, Olga Glyzina, Ragothaman Prathiviraj, George Seghal Kiran, and Joseph Selvin. 2021. "Microbiome Changes of Endemic Lake Baikal Sponges during Bleaching Syndrome Development" Diversity 13, no. 12: 653. https://doi.org/10.3390/d13120653
APA StyleItskovich, V., Kaluzhnaya, O., Glyzina, O., Prathiviraj, R., Seghal Kiran, G., & Selvin, J. (2021). Microbiome Changes of Endemic Lake Baikal Sponges during Bleaching Syndrome Development. Diversity, 13(12), 653. https://doi.org/10.3390/d13120653