
Diversity of Ostrya carpinifolia Forests in Ravine Habitats of Serbia (S-E Europe)

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
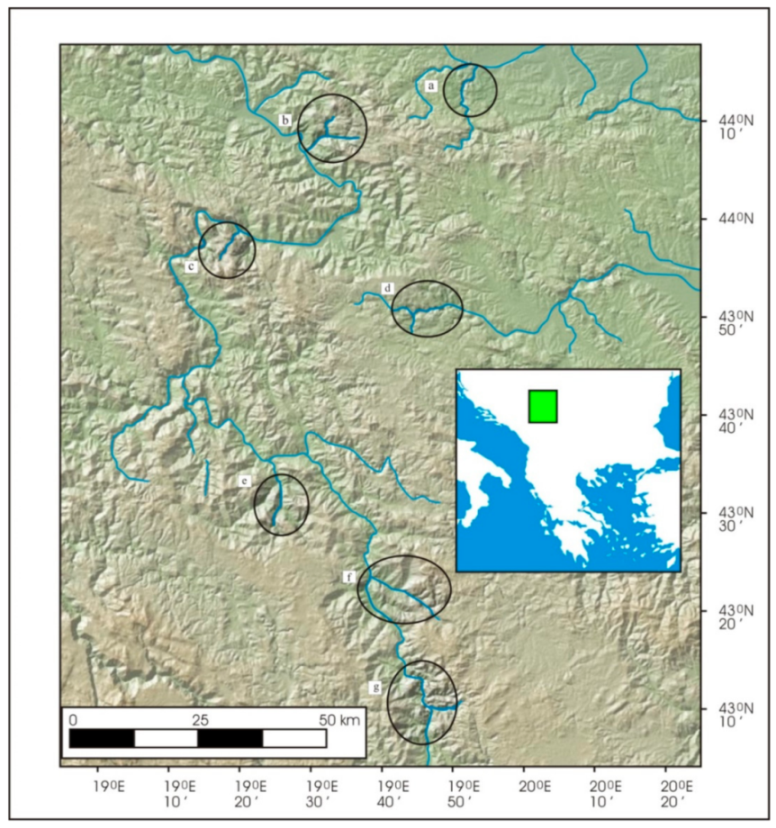
2.1. Study Area and Sampling Procedures
2.2. Statistical Analyses
3. Results
3.1. Classification of Communities
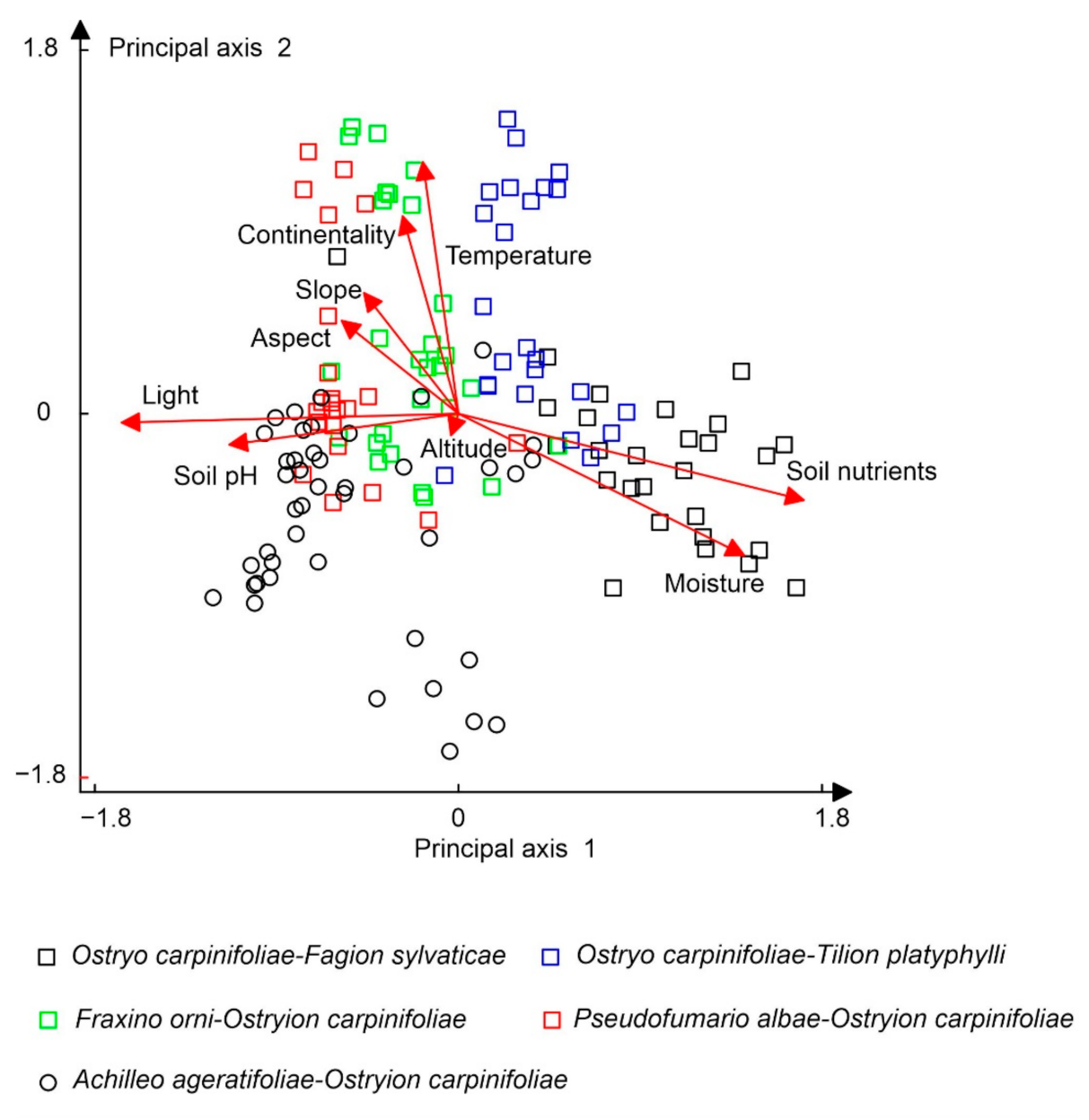
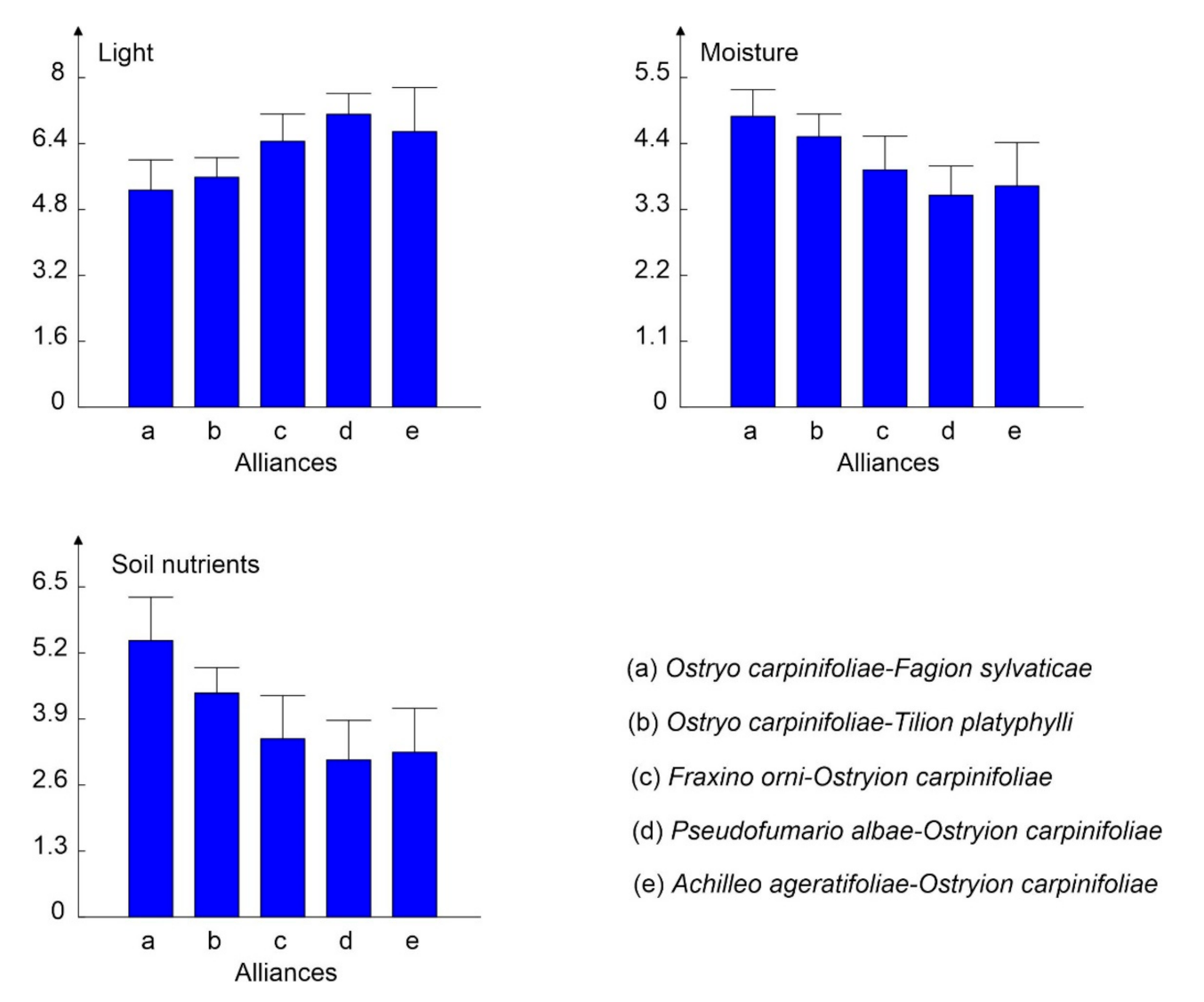
3.2. Patterns in the Vegetation–Environment Relationship
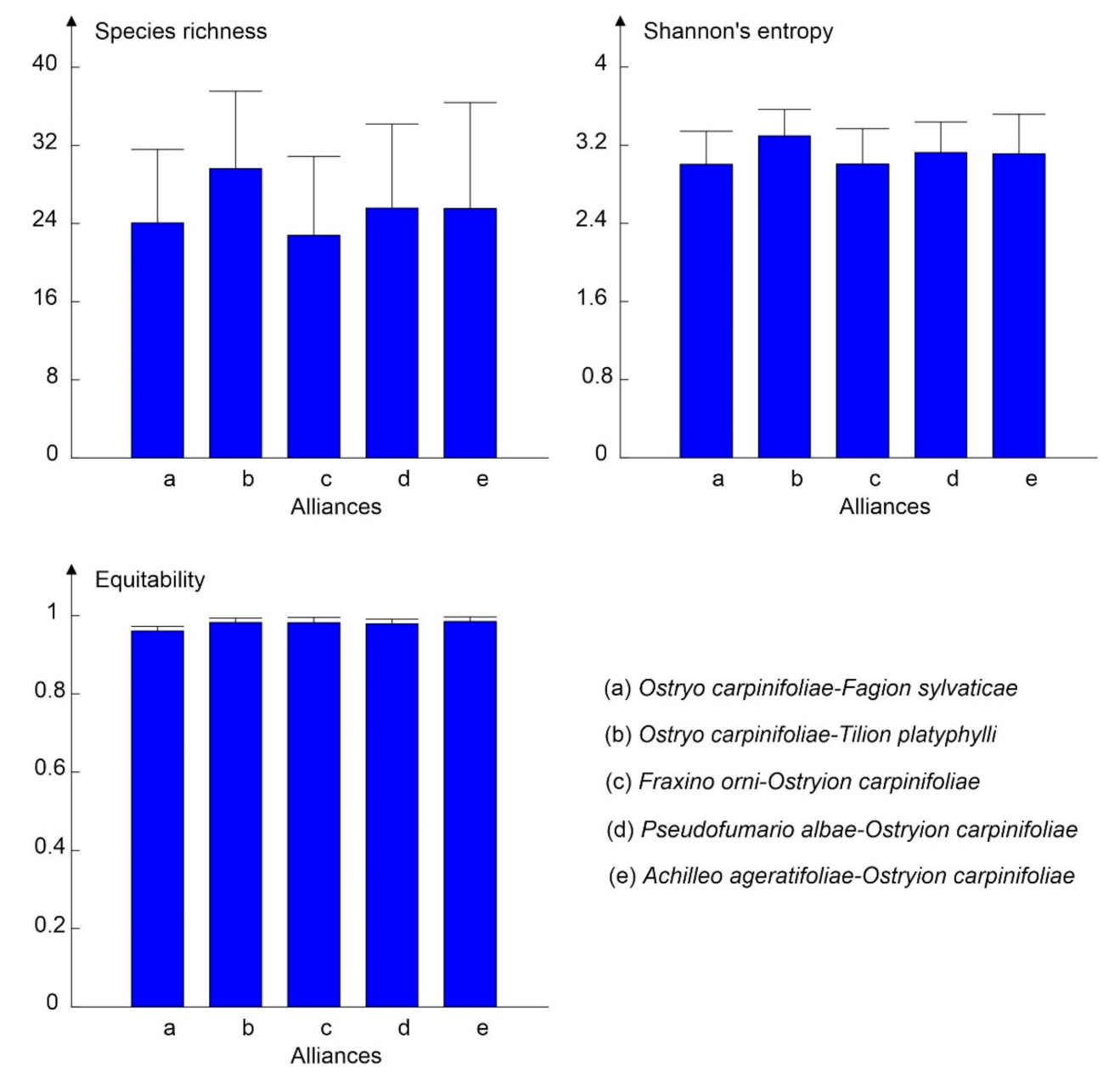
3.3. Alpha Diversity
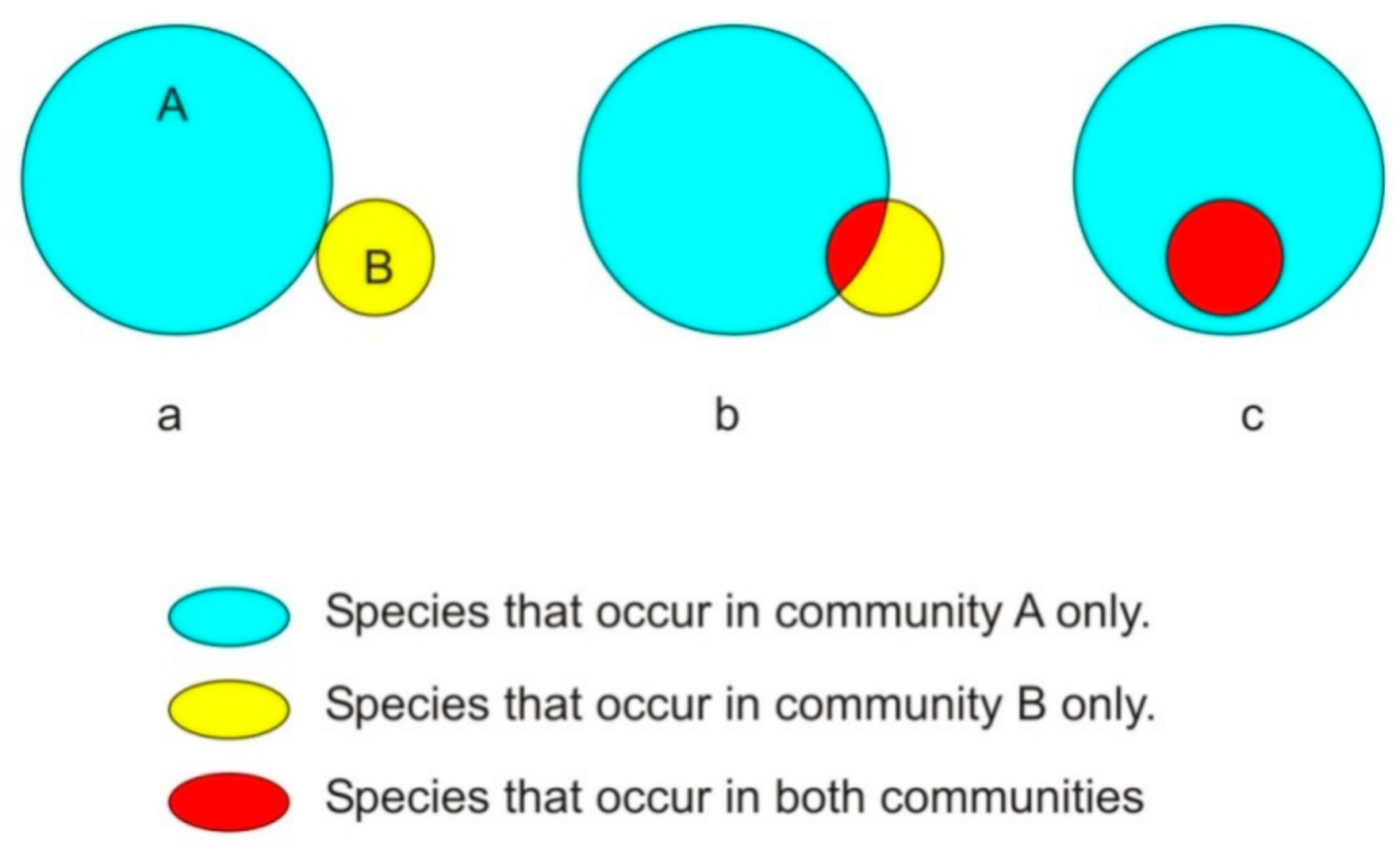
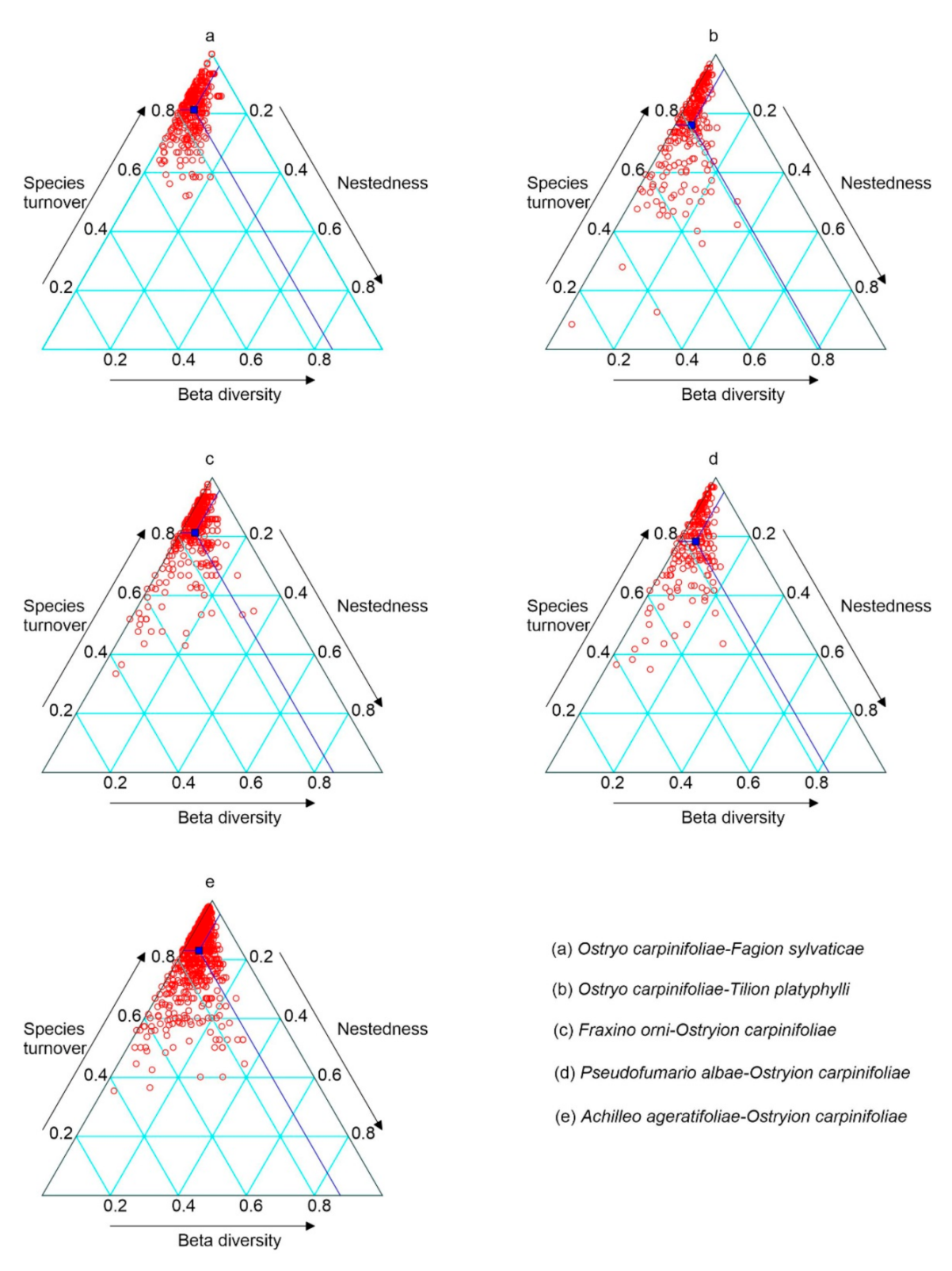
3.4. Beta Diversity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Alliances | ||||
|---|---|---|---|---|---|
| A | B | C | D | E | |
| Asperula taurina L. | 38 | . | . | . | . |
| Epimedium alpinum L. | 47 | . | . | . | . |
| Fraxinus excelsior L. | 39 | . | . | . | . |
| Evonymus europaeus L. | 39 | . | . | . | . |
| Alliaria petiolata (M.Bieb.) Cavara & Grande | 39 | . | . | . | . |
| Heracleum sphondylium L. | 39 | . | . | . | . |
| Primula veris L. | 30 | . | . | . | . |
| Pulmonaria officinalis L. | 39 | . | . | . | . |
| Melampyrum hoermannianum K. Malý | 44 | . | . | . | . |
| Angelica sylvestris L. | 35 | . | . | . | . |
| Galeopsis speciosa Mill. | 35 | . | . | . | . |
| Parietaria officinalis L. | 37 | 3 | . | . | . |
| Convallaria majalis L. | 35 | . | . | . | . |
| Athyrium filix-femina (L.) Roth | 43 | . | . | . | . |
| Cardamine bulbifera (L.) Crantz | 29 | 1 | . | . | . |
| Equisetum telmateia Ehrh. | 30 | . | . | . | . |
| Cyclamen purpurascens Mill. | 53 | . | . | . | . |
| Acer pseudoplatanus L. | 30 | . | . | . | . |
| Brachypodium sylvaticum (Huds.) P.Beauv. | 50 | 11 | . | . | . |
| Salvia glutinosa L. | 38 | 22 | . | . | . |
| Hedera helix L. | 45 | 37 | . | . | . |
| Asarum europaeum L. | 28 | 18 | . | . | . |
| Tilia platyphyllos Scop. | 10 | 22 | 7 | . | . |
| Helleborus odorus Waldst. & Kit. ex Willd. | 11 | 55 | . | . | . |
| Scutellaria altissima L. | 15 | 39 | . | . | . |
| Scrophularia nodosa L. | 4 | 47 | . | . | . |
| Aremonia agrimonoides (L.) DC. | 10 | 26 | 4 | . | . |
| Juglans regia L. | . | 41 | . | . | . |
| Cornus mas L. | . | 29 | 9 | 2 | . |
| Campanula rapunculoides L. | . | 27 | 6 | 4 | . |
| Digitalis grandiflora Mill. | . | 36 | 2 | . | . |
| Calamintha grandiflora (L.) Moench. | . | 26 | 16 | . | . |
| Arabis turrita L. | . | 27 | 1 | . | 1 |
| Lilium martagon L. | . | 37 | . | . | . |
| Rubus hirtus Wald. et Kit. | . | 42 | . | . | . |
| Campanula trachelium L. | . | 33 | 14 | . | . |
| Sanicula europaea L. | . | 37 | . | . | . |
| Arum maculatum L. | . | 32 | . | . | . |
| Sorbus torminalis (L.) Crantz. | . | 37 | . | . | . |
| Galium schultesii Vest | . | 40 | . | . | . |
| Cornus sanguinea L. | . | 35 | 10 | . | . |
| Potentilla micrantha Ram. ex DC. | . | 7 | 29 | . | . |
| Carex cariophyllea Latourr. | . | 5 | 26 | . | . |
| Glechoma hirsuta Waldst. et Kit. | . | 3 | 25 | 5 | . |
| Lathyrus sylvestris L. | . | . | 29 | 2 | . |
| Clinopodium nepeta (L.) Kuntze | . | . | 34 | . | . |
| Geranium sanguineum L. | . | 7 | 21 | . | . |
| Fraxinus ornus L. | . | 15 | 30 | 17 | . |
| Pseudofumaria alba (Mill.) Lidén | 5 | 30 | 10 | ||
| Stachys recta L. | . | . | 19 | 26 | . |
| Teucrium montanum L. | . | . | 11 | 29 | . |
| Prunus spinosa L. | . | . | 12 | 24 | . |
| Draba lasiocarpa Rochel | . | . | 2 | 28 | . |
| Allium flavum L. | . | . | 6 | 22 | . |
| Evonymus verrucosus Scop. | . | . | 3 | 25 | 18 |
| Melica ciliata L. | . | . | . | 32 | 12 |
| Campanula lingulata Waldst. & Kit. | . | 8 | . | 33 | . |
| Fritillaria montana Hoppe ex W.D.J.Koch | . | . | . | 27 | . |
| Minuartia bosniaca (Beck) K.Malý | . | . | . | 27 | . |
| Micromeria thymifolia (Scop.) Fritsch | . | . | . | 32 | 9 |
| Stipa calamagrostis (L.) P.Beauv. | . | . | . | 29 | . |
| Rhamnus saxatilis Jacq. | . | . | . | 34 | . |
| Galium purpureum L. | . | . | . | 24 | 24 |
| Globularia cordifolia L. | . | . | . | 13 | 21 |
| Hieracium waldsteinii Tausch | . | . | . | . | 50 |
| Geranium macrorrhizum L. | . | . | . | . | 48 |
| Campanula secundiflora Vis. & Pancic | . | . | . | . | 39 |
| Genista radiata (L.) Scop. | . | . | . | . | 37 |
| Saxifraga crustata Vest. | . | . | . | . | 34 |
| Valeriana montana L. | . | . | . | . | 34 |
| Cerastium decalvans Schloss. & Vuk. | . | . | . | . | 30 |
| Jurinea mollis (Torn.) Rchb. | . | . | . | . | 31 |
| Saxifraga tridactylites L. | . | . | . | 2 | 32 |
| Silene pusilla Waldst. et Kit. | . | . | . | . | 31 |
| Achillea ageratifolia (Sibth. & Sm.) Boiss | . | . | . | . | 38 |
| Seseli rigidum Waldst. & Kit. | . | . | . | . | 35 |
| Edraianthus graminifolius (L.) A.DC. | . | . | . | 4 | 37 |
| Euphorbia glabriflora Vis. | . | . | . | 7 | 34 |
| Frangula rupestris Schur | . | . | . | . | 34 |
| Amphoricarpos neumayeri Vis. | . | . | . | 2 | 21 |
References
- Mucina, L.; Bültmann, H.; Dierssen, K.; Theurillat, J.P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; García, R.G.; et al. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci. 2016, 19, 3–264. [Google Scholar] [CrossRef]
- Blasi, C.; Filibeck, G.; Rosati, L. Classification of southern Italy Ostrya carpinifolia woods. Fitosociologia 2006, 43, 3–23. [Google Scholar]
- Košir, P.; Čarni, A.; di Pietro, R. Classification and phytogeographical differentiation of broad-leaved ravine forests in southeastern Europe. J. Veg. Sci. 2008, 19, 331–342. [Google Scholar] [CrossRef]
- Čarni, A.; Košir, P.; Karadžić, B.; Matevski, V.; Redžić, S.; Škvorc, Ž. Thermophilous deciduous forests in Southeastern Europe. Plant Biosyst. 2009, 143, 1–13. [Google Scholar] [CrossRef]
- Stupar, V.; Brujić, J.; Škvorc, Ž.; Čarni, A. Vegetation types of thermophilous deciduous forests (Quercetea pubescentis) in the Western Balkans. Phytocoenologia 2016, 46, 49–68. [Google Scholar] [CrossRef]
- Karadžić, B.; Mijović, A.; Popović, R.; Perišić, S.; Marinković, S. Forest vegetation in West-Serbian canyons: Biodiversity hotspots. In Forest Research: A Challenge for an Integrated European Approach; Ragodlou, K., Ed.; NAGREF-Forest Research Institute: Thessaloniki, Greece, 2001; pp. 513–518. [Google Scholar]
- Karadžić, B. Chasmophytic forests of Ostrya carpinifolia in west-Serbian canyons. Biol. Nyssana 2017, 8, 73–81. [Google Scholar]
- Karadžić, B.; Bulić, Z.; Jarić, S.; Mitrović, M.; Pavlović, P. Vegetation in Ravine Habitats of Montenegro. In The Rivers of Montenegro; Pešić, V., Paunović, M., Kostianoy, A.G., Eds.; Springer Nature Switzerland AG: Cham, Switzerland, 2020; pp. 201–230. [Google Scholar]
- Popović, R.; Kojić, M.; Karadžić, B. Ecological characteristics of six important submediterranean tree species in Serbia. Bocconea 1996, 5, 431–438. [Google Scholar]
- Birks, H.J.B.; Willis, K.J. Alpines, trees, and refugia in Europe. Plant Ecol. Divers. 2008, 1, 147–160. [Google Scholar] [CrossRef]
- Hewitt, G.M. Some genetic consequences of ice ages, and their role in divergence and speciation. Biol. J. Linn. Soc. 1996, 58, 247–276. [Google Scholar] [CrossRef]
- Hewitt, G.M. Postglacial recolonization of European Biota. Biol. J. Linn. Soc. 1999, 68, 87–112. [Google Scholar] [CrossRef]
- Hewitt, G.M. The genetic legacy of the quaternary ice ages. Nature 2000, 405, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, G.M. Mediterranean peninsulas: The evolution of hotspots. In Biodiversity Hotspots. Distribution and Protection of Conservation Priority Areas; Zachos, F.E., Habel, J.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 123–147. [Google Scholar]
- Puncer, I.; Zupančič, M. Die ökologische und wirtschaftliche Bedeutung der Ostrya carpinifolia Scop. in Slowenien. Studia Geobot. 1982, 2, 25–32. [Google Scholar]
- Tomić, Z. Cenoareal crnog graba (Ostrya carpinifolia Scop.) u Srbiji. Bot. Serbica 1994, 28, 173–182. [Google Scholar]
- Jovanović, B. Šuma crnog graba u okolini Titovog Užica (Sesleria variae-Ostryetum ass. n.). Šumarstvo 1972, 25, 3–9. [Google Scholar]
- Trinajstić, I.; Cerovečki, Z. O cenoarealu crnoga graba, Ostrya carpinifolia Scop. (Corylaceae) u Hrvatskoj [On coenoareal of hop-hornbeam, Ostrya carpinifolia Scop. (Corylaceae) in Croatia]. Biosistematika 1978, 4, 57–65. [Google Scholar]
- Tomić, Z. Fitocenoze crnoga graba (Ostrya carpinifolia Scop.) u Srbiji [Phytocenoses of Hop hornbeam (Ostrya carpinifolia Scop.) in Serbia]. Ph.D. Thesis, University in Belgrade, Belgrade, Serbia, 1980. [Google Scholar]
- Lakušić, R.; Pavlović, D.; Redžić, S. Horološko-ekološka i floristička diferencijacija šuma i šikara sa bjelograbićem (Carpinus orientalis Mill.) i crnim grabom (Ostrya carpinifolia Scop.) na prostoru Jugoslavije. Glas. Republičkog Zavoda za Zaštitu Prir. Prir. Muz. Titogr. 1982, 15, 103–116. [Google Scholar]
- Tomić, Z. Specijski diverzitet u crnograbovim šumama sveze Orno-Ostryon Tomž. 1940. i njegove karakteristike. Zaštita Prir. 1998, 50, 57–61. [Google Scholar]
- Tomić, Z. Zajednica Orno-Ostryetum Aich. 1933 u refugijumima jugozapadne Srbije. Glas. Šumarskog Fak. 2000, 82, 177–189. [Google Scholar]
- Tomić, Z. Šume crnog graba (Hop hornbeam forests). In Vegetacija Srbije 2—Šumske zajednice 2; Škorić, D.M., Ed.; Srpska akademija nauka i umetnosti, Odeljenje hemijskih i bioloških nauka: Beograd, Srbija, 2006; pp. 29–68. [Google Scholar]
- Braun-Blanquet, J. Plant Sociology: The Study of Plant Communities; Hafner: London, UK, 1965. [Google Scholar]
- Van der Maarel, E. Transformation of coverabundance values in phytosociology and its effects on community similarity. Vegetatio 1979, 39, 97–114. [Google Scholar]
- MacQueen, J. Some Methods for Classification and Analysis of Multivariate Observations, Proceedings of the Fifth Berkeley Symposium on Mathematical Statistics and Probability, Oakland, CA, USA, 27 December 1965–7 January 1966; University of California Press: Los Angeles, CA, USA, 1967; pp. 281–297. [Google Scholar]
- James, G.; Witten, D.; Hastie, T.; Tibshirani, R. An Introduction to Statistical Learning: With Applications in R; Springer: New York, NY, USA, 2013. [Google Scholar]
- Marinković, N.; Karadžić, B.; Pešić, V.; Gligorović, B.; Grosser, C.; Paunović, M.; Nikolić, V.; Raković, M. Faunistic patterns and diversity components of leech assemblages in karst springs of Montenegro. Knowl. Manag. Aquat. Ecosyst. 2019, 420, 26. [Google Scholar]
- Chytrý, M.; Tichý, L.; Holt, J.; Botta-Dukát, Z. Determination of diagnostic species with statistical fidelity measures. J. Veg. Sci. 2002, 13, 79–90. [Google Scholar] [CrossRef]
- Ter Braak, C.J. Canonical correspondence analysis: A new eigenvector technique for multivariate direct gradient analysis. Ecology 1986, 67, 1167–1179. [Google Scholar] [CrossRef] [Green Version]
- Ellenberg, H.; Leuschner, C. Vegetation Mitteleuropas mit den Alpen, 6th ed.; Ulmer: Stuttgart, Germany, 2010. [Google Scholar]
- Pielou, E.C. Population and Community Ecology: Principles and Methods; Gordon and Breach: New York, NY, USA, 1974. [Google Scholar]
- Whittaker, R.H. Evolution and measurement of species diversity. Taxon 1972, 21, 213–251. [Google Scholar] [CrossRef] [Green Version]
- Anderson, M.J.; Crist, T.O.; Chase, J.M.; Vellend, M.; Inouye, B.D.; Freestone, A.L.; Sanders, N.J.; Cornell, H.V.; Comita, L.S.; Davies, K.F.; et al. Navigating the multiple meanings of beta diversity: A roadmap for the practicing ecologist. Ecol. Lett. 2011, 14, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Baselga, A. The relationship between species replacement, dissimilarity derived from nestedness, and nestedness. Glob. Ecol. Biogeogr. 2012, 21, 1223–1232. [Google Scholar] [CrossRef]
- Podani, J.; Ricotta, C.; Schmera, D. A general framework for analyzing beta diversity, nestedness and related community-level phenomena based on abundance data. Ecol. Complex. 2013, 15, 52–61. [Google Scholar] [CrossRef] [Green Version]
- Schmera, D.; Podani, J. Comments on separating components of beta diversity. Community Ecol. 2011, 12, 153–160. [Google Scholar] [CrossRef]
- Cardoso, P.; Rigal, F.; Carvalho, J.C.; Fortelius, M.; Borges, P.A.V.; Podani, J.; Schmera, D. Partitioning taxon, phylogenetic and functional beta diversity into replacement and richness difference components. J. Biogeogr. 2014, 41, 749–761. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, J.C.; Cardoso, P.; Gomes, P. Determining the relative roles of species replacement and species richness differences in generating beta-diversity patterns. Glob. Ecol. Biogeogr. 2012, 21, 760–771. [Google Scholar] [CrossRef]
- Legendre, P. Interpreting the replacement and richness difference components of beta diversity. Glob. Ecol. Biogeogr. 2014, 23, 1324–1334. [Google Scholar] [CrossRef]
- Karadžić, B. FLORA: A software package for statistical analysis of ecological data. Water Res. Manag. 2013, 3, 45–54. [Google Scholar]
- Pielou, E.C. The Interpretation of Ecological Data: A Primer on Classification and Ordination; Wiley: New York, NY, USA, 1984. [Google Scholar]
- Jongman, R.H.G.; ter Braak, C.J.F.; van Tongeren, O.F.R. Data Analysis in Community and Landscape Ecology; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical Ecology; Elsevier: New York, NY, USA, 2012. [Google Scholar]
- Hill, M.O.; Bunce, R.G.H.; Shaw, M.W. Indicator species analysis, a divisive polythetic method of classification and its application to a survey of native pinewoods in Scotland. J. Ecol. 1975, 63, 597–613. [Google Scholar] [CrossRef]
- Van Groenewoud, H. The robustness of Correspondence, Detrended Correspondence, and TWINSPAN Analysis. J. Veg. Sci. 1992, 3, 239–246. [Google Scholar] [CrossRef]
- Belbin, L.; McDonald, C. Comparing Three Classification Strategies for Use in Ecology. J. Veg. Sci. 1993, 4, 341–348. [Google Scholar] [CrossRef] [Green Version]
- Austin, M.P.; Nicholls, A.O.; Doherty, M.D.; Meyers, J.A. Determining species response functions to an environmental gradient by means of a β-function. J. Veg. Sci. 1994, 5, 215–228. [Google Scholar] [CrossRef]
- Karadžić, B.; Marinković, S.; Kataranovski, D. Use of the β-function to estimate the skewness of species responses. J. Veg. Sci. 2003, 14, 799–805. [Google Scholar] [CrossRef]
- Karadžić, B. Beech forests (order Fagetalia sylvaticae Pawlowski 1928) in Serbia. Bot. Serbica 2018, 42, 91–107. [Google Scholar]
- Whittaker, R.H. Communities and Ecosystems; MacMillan: New York, NY, USA, 1970. [Google Scholar]
- Lloyd, M.; Ghelardi, R.J. A Table for Calculating the Equitability Component of Species Diversity. J. Anim. Ecol. 1964, 33, 217–225. [Google Scholar] [CrossRef]
- Stevanović, V. Analysis of the Central European and Mediterranean orophytic element on the mountains of the West and Central Balkan Peninsula, with special reference to endemics. Bocconea 1996, 5, 77–97. [Google Scholar]
- Stevanović, V.; Tan, K.; Petrova, A. Mapping the endemic flora of the Balkans—A progress report. Bocconea 2007, 21, 131–137. [Google Scholar]
- Tomović, G.; Niketić, M.; Lakušić, D.; Ranđelović, V.; Stevanović, V. Balkan endemic plants in Central Serbia and Kosovo regions: Distribution patterns, ecological characteristics, and centres of diversity. Bot. J. Linn. Soc. 2014, 176, 173–202. [Google Scholar] [CrossRef] [Green Version]
- Velčev, V.; Kožuharov, S.; Ančev, M. Atlas of the Endemic Plants in Bulgaria; Publishing House of the Bulgarian Academy of Sciences: Sofia, Bulgaria, 1992. [Google Scholar]
- Davis, H.P. Cliff vegetation in the Eastern Mediterranean. J. Ecol. 1951, 39, 63–93. [Google Scholar] [CrossRef]
- Surina, B.; Schönswetter, P.; Schneeweiss, G.M. Quaternary range dynamics of ecologically divergent species (Edraianthus serpyllifolius and E. tenuifolius, Campanulaceae) within the Balkan refugium. J. Biogeogr. 2011, 38, 1381–1393. [Google Scholar] [CrossRef]
- Caković, D.; Stešević, D.; Schönswetter, P.; Frajman, B. How many taxa? Spatiotemporal evolution and taxonomy of Amphoricarpus (Asteraceae, Carduoideae) on the Balkan Peninsula. Org. Divers. Evol. 2015, 15, 429–455. [Google Scholar] [CrossRef]
- Council Directive 92/43/EEC of 21 May 1992 on the Conservation of Natural Habitats and of Wild Fauna and Flora. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:31992L0043&from=EN (accessed on 1 February 2021).








| River | Geographic Coordinates | Number of Relevés |
|---|---|---|
| Gradac | 44.188436 N, 19.868344 E–44.233333 N, 19.867778 E | 23 |
| Trešnjica | 44.141667 N, 19.531667 E–44.150278 N, 19.540278 E | 28 |
| Derventa | 43.954167 N, 19.356944 E–43.963056 N, 19.358333 E | 21 |
| Djetinja | 43.848611 N, 19.808611 E–43.844444 N, 19.772778 E | 18 |
| Ljutina | 43.475833 N, 19.405556 E–43.501389 N, 19.422500 E | 20 |
| Mileševka | 43.364444 N, 19.728889 E–43.358889 N, 19.739722 E | 15 |
| Lim | 43.190278 N, 19.766111 E–43.213056 N, 19.764167 E | 19 |
| Classification Method | ||||||
|---|---|---|---|---|---|---|
| K-Means Clustering | Bayesian Classification | |||||
| Number of Groups | B | W | B/W | B | W | B/W |
| 2 | 0.0359 | 1.4760 | 0.0243 | 0.0344 | 1.4721 | 0.0234 |
| 3 | 0.0534 | 2.1473 | 0.0249 | 0.0513 | 2.1435 | 0.0239 |
| 4 | 0.0714 | 2.7889 | 0.0256 | 0.0721 | 2.7865 | 0.0259 |
| 5 | 0.1026 | 3.2876 | 0.0312 | 0.0890 | 3.4257 | 0.0260 |
| 6 | 0.1097 | 3.8599 | 0.0284 | 0.0866 | 4.0811 | 0.0212 |
| 7 | 0.1094 | 4.5725 | 0.0239 | 0.0934 | 4.7242 | 0.0198 |
| 8 | 0.1115 | 5.2554 | 0.0212 | 0.1089 | 5.3205 | 0.0205 |
| 9 | 0.1123 | 5.9542 | 0.0189 | 0.1112 | 5.9612 | 0.0187 |
| 10 | 0.1221 | 6.5167 | 0.0187 | 0.1201 | 6.5223 | 0.0184 |
| Variable | Eigenvalue | F Statistic | Probability |
|---|---|---|---|
| Soil nutrients | 0.4214 | 5.101 | 0.000 * |
| Light | 0.2365 | 2.818 | 0.000 * |
| Temperature | 0.2252 | 2.682 | 0.000 * |
| Continentality | 0.1730 | 2.051 | 0.000 * |
| Soil acidity | 0.1324 | 1.564 | 0.000 * |
| Moisture | 0.1234 | 1.457 | 0.000 * |
| Aspect | 0.1087 | 1.282 | 0.002 * |
| Slope | 0.1025 | 1.208 | 0.006 * |
| Altitude | 0.0799 | 0.939 | 0.326 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sekulić, D.; Karadžić, B.; Kuzmanović, N.; Jarić, S.; Mitrović, M.; Pavlović, P. Diversity of Ostrya carpinifolia Forests in Ravine Habitats of Serbia (S-E Europe). Diversity 2021, 13, 59. https://doi.org/10.3390/d13020059
Sekulić D, Karadžić B, Kuzmanović N, Jarić S, Mitrović M, Pavlović P. Diversity of Ostrya carpinifolia Forests in Ravine Habitats of Serbia (S-E Europe). Diversity. 2021; 13(2):59. https://doi.org/10.3390/d13020059
Chicago/Turabian StyleSekulić, Dimitrije, Branko Karadžić, Nevena Kuzmanović, Snežana Jarić, Miroslava Mitrović, and Pavle Pavlović. 2021. "Diversity of Ostrya carpinifolia Forests in Ravine Habitats of Serbia (S-E Europe)" Diversity 13, no. 2: 59. https://doi.org/10.3390/d13020059
APA StyleSekulić, D., Karadžić, B., Kuzmanović, N., Jarić, S., Mitrović, M., & Pavlović, P. (2021). Diversity of Ostrya carpinifolia Forests in Ravine Habitats of Serbia (S-E Europe). Diversity, 13(2), 59. https://doi.org/10.3390/d13020059