
Functional Traits of a Rainforest Vascular Epiphyte Community: Trait Covariation and Indications for Host Specificity




Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Site and Focal Tree Species
2.2. Functional Traits of Epiphytes
2.3. Data Analysis
3. Results
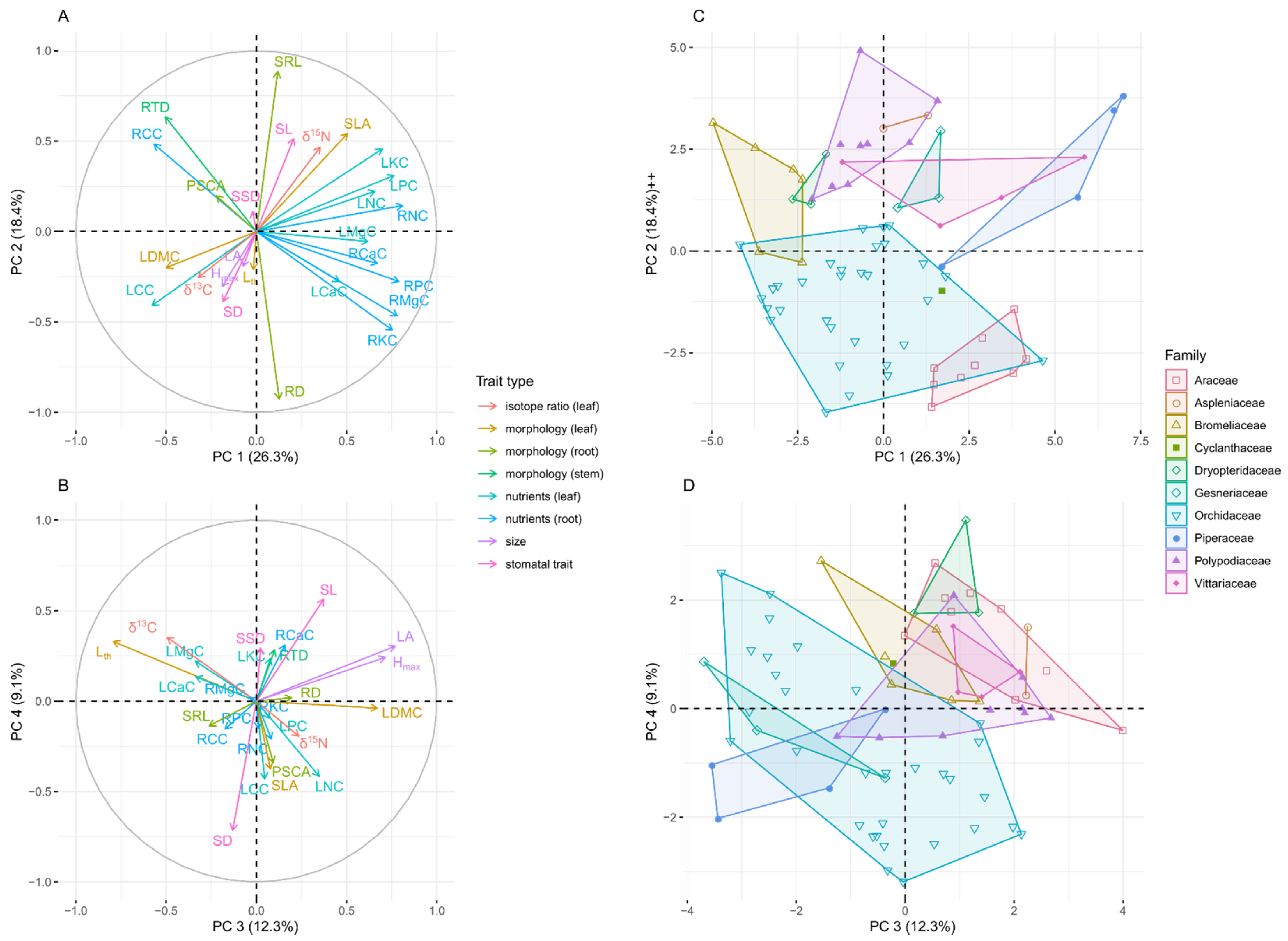
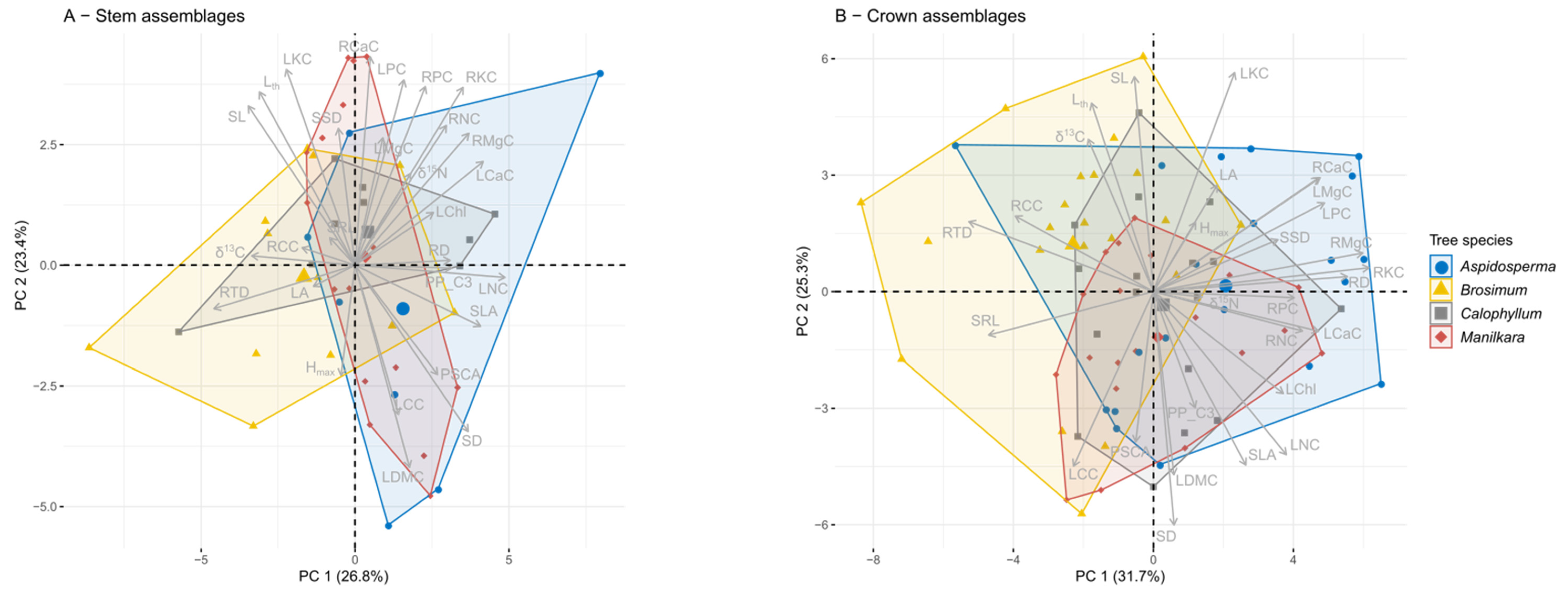
3.1. Epiphyte Trait Values and Trait Covariation
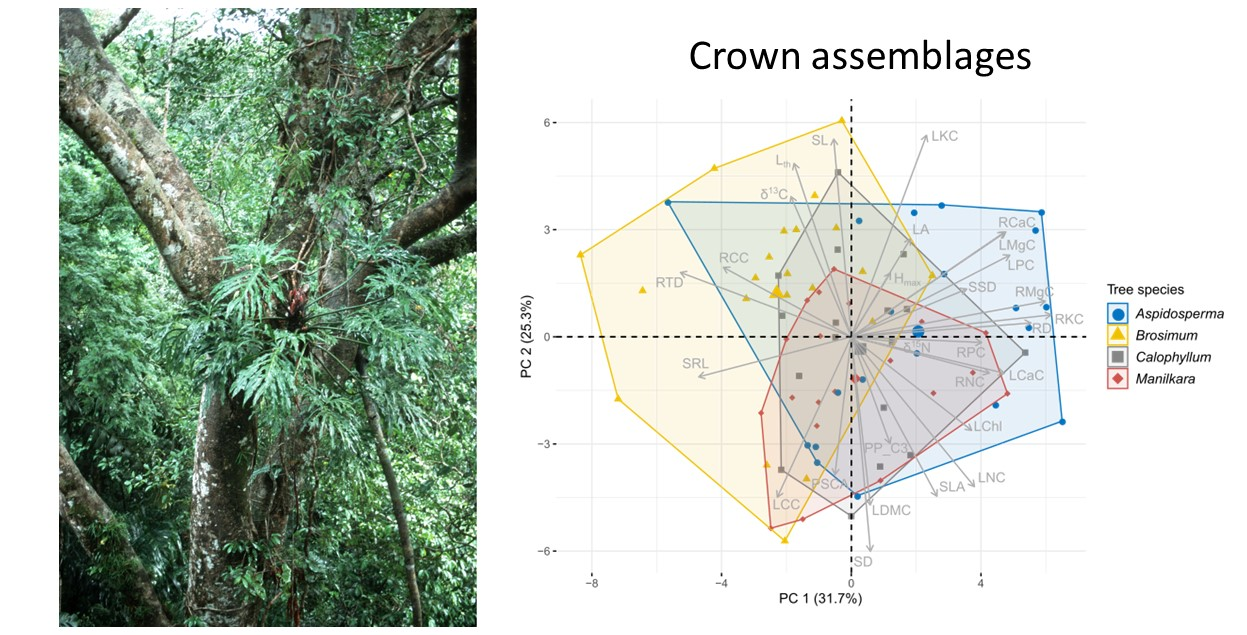
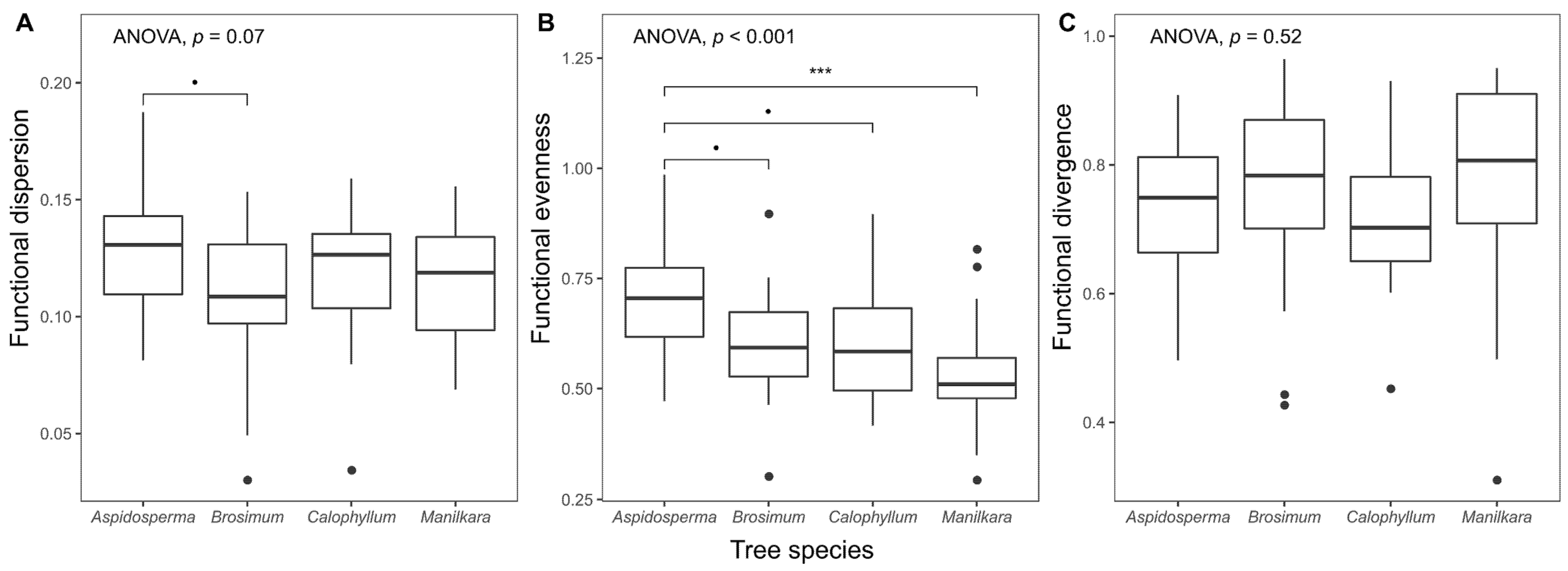
3.2. Differences among Tree Species
4. Discussion
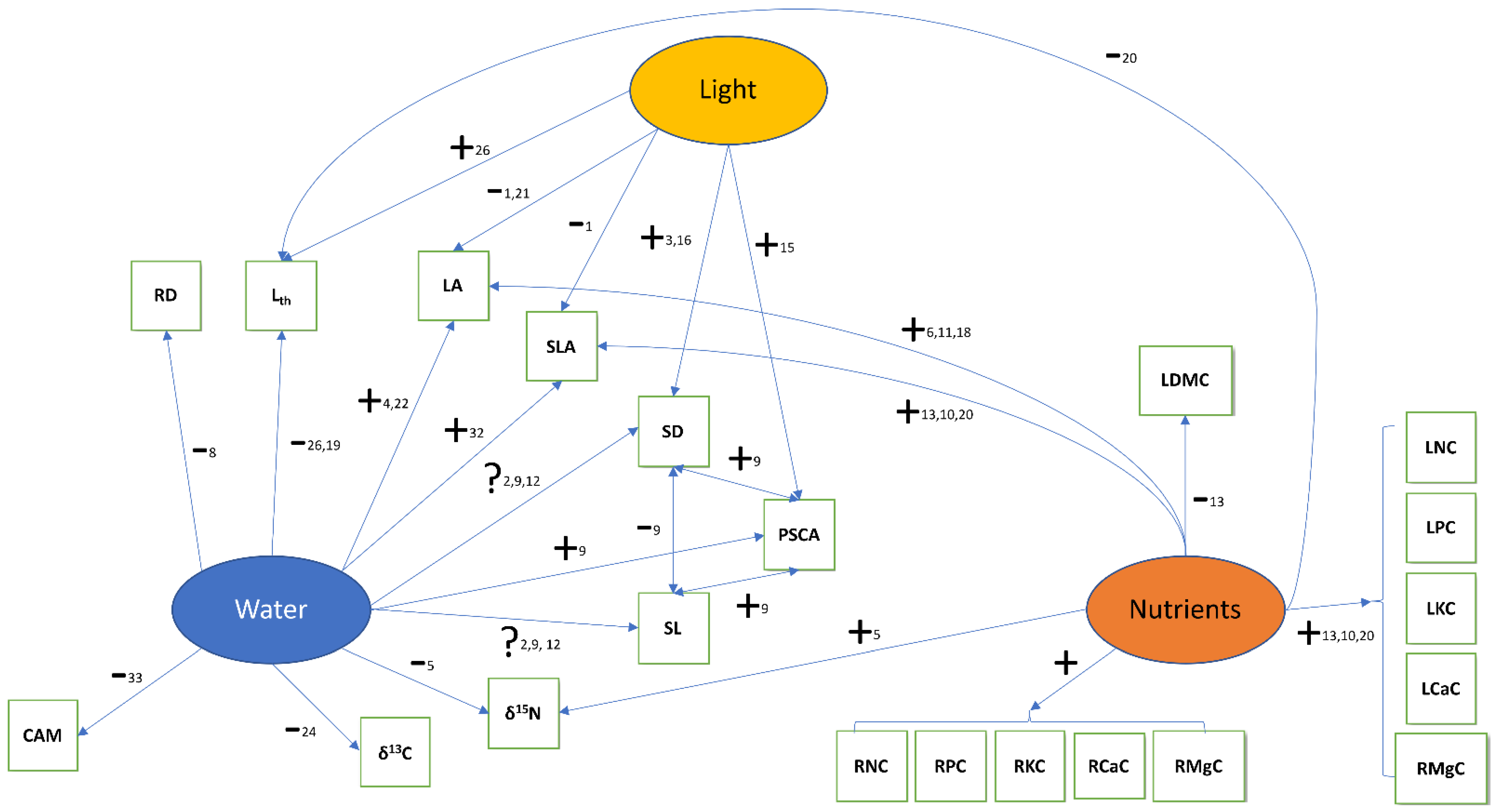
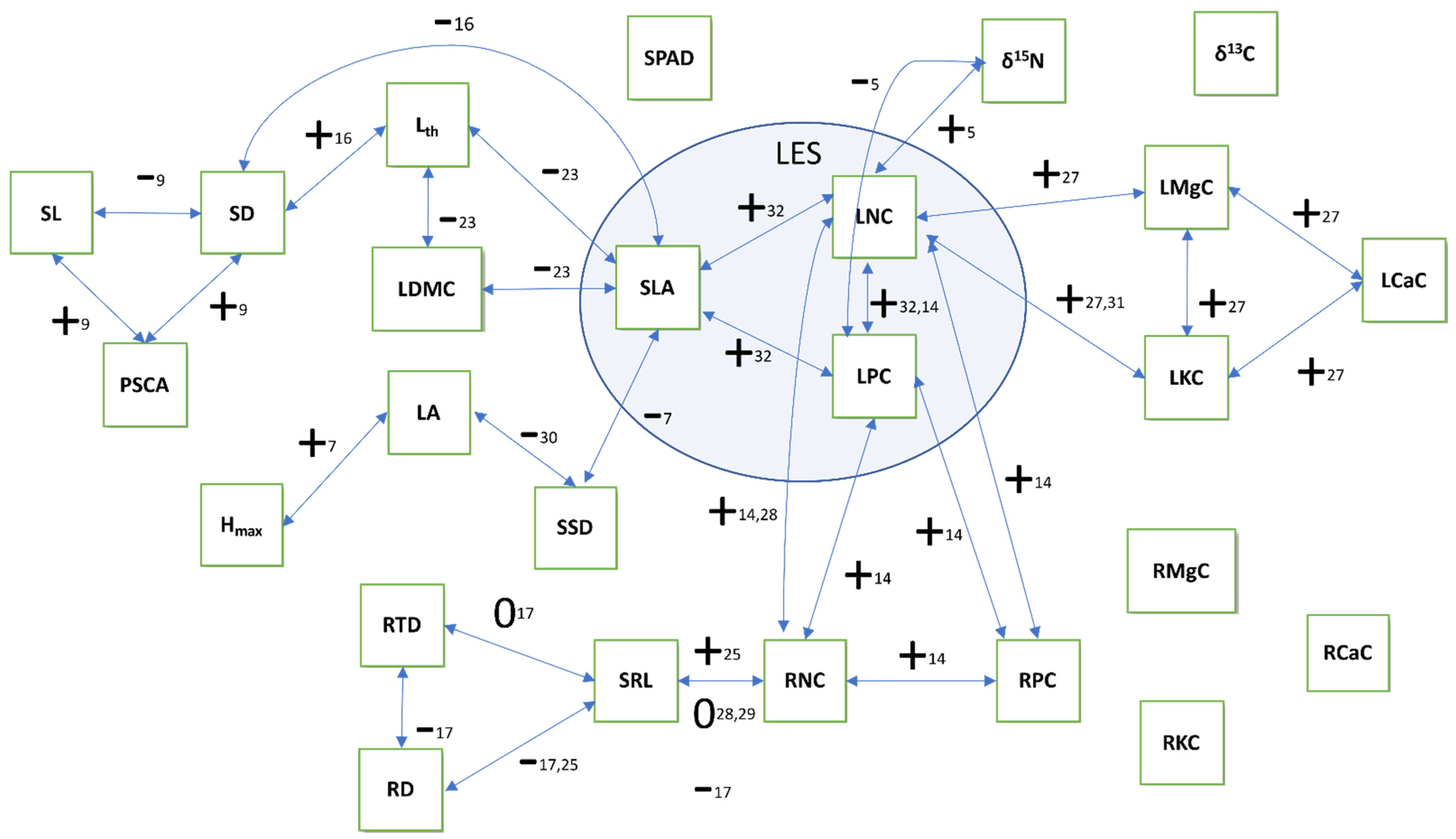
4.1. Trait Covariation
4.2. The Relationship of Epiphyte Trait Patterns and Host Tree Species
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zotz, G. Plants on Plants—The Biology of Vascular Epiphytes; Springer International Publishing: Cham, Switzerland, 2016. [Google Scholar] [CrossRef]
- Janzen, T.; Zotz, G.; Etienne, R.S. Community structure of vascular epiphytes: A neutral perspective. Oikos 2020, 129, 853–867. [Google Scholar] [CrossRef] [Green Version]
- Petter, G.; Wagner, K.; Wanek, W.; Sánchez Delgado, E.J.; Zotz, G.; Cabral, J.S.; Kreft, H. Functional leaf traits of vascular epiphytes: Vertical trends within the forest, intra- and interspecific trait variability, and taxonomic signals. Funct. Ecol. 2016, 30, 188–198. [Google Scholar] [CrossRef] [Green Version]
- Zotz, G. Johansson revisited: The spatial structure of epiphyte assemblages. J. Veg. Sci. 2007, 18, 123–130. [Google Scholar] [CrossRef]
- Krömer, T.; Kessler, M.; Gradstein, S.R. Vertical stratification of vascular epiphytes in submontane and montane forest of the Bolivian Andes: The importance of the understory. Plant Ecol. 2007, 189, 261–278. [Google Scholar] [CrossRef]
- Zotz, G. How prevalent is crassulacean acid metabolism among vascular epiphytes? Oecologia 2004, 138, 184–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, K.; Mendieta-Leiva, G.; Zotz, G. Host specificity in vascular epiphytes: A review of methodology, empirical evidence and potential mechanisms. AoB Plants 2015, 7, plu092. [Google Scholar] [CrossRef] [Green Version]
- Chaves, C.J.N.; Dyonisio, J.C.; Rossatto, D.R. Host trait combinations drive abundance and canopy distribution of atmospheric bromeliad assemblages. AoB Plants 2016, 8, plw010. [Google Scholar] [CrossRef] [Green Version]
- Laube, S.; Zotz, G. Neither host-specific nor random: Vascular epiphytes on three tree species in a Panamanian lowland forest. Ann. Bot. 2006, 97, 1103–1114. [Google Scholar] [CrossRef] [Green Version]
- Vergara-Torres, C.A.; Pacheco-Alvarez, M.C.; Flores-Palacios, A. Host preference and host limitation of vascular epiphytes in a tropical dry forest of central Mexico. J. Trop. Ecol. 2010, 26, 563–570. [Google Scholar] [CrossRef]
- Schleuning, M.; Fründ, J.; García, D. Predicting ecosystem functions from biodiversity and mutualistic networks: An extension of trait-based concepts to plant–animal interactions. Ecography 2015, 38, 380–392. [Google Scholar] [CrossRef]
- Callaway, R.M.; Reinhart, K.O.; Moore, G.W.; Moore, D.J.M.; Pennings, S.C. Epiphyte host preferences and host traits: Mechanisms for species-specific interactions. Oecologia 2002, 132, 221–230. [Google Scholar] [CrossRef]
- Mehltreter, K.; Flores-Palacios, A.; García-Franco, J.G. Host preferences of low-trunk vascular epiphytes in a cloud forest of Veracruz, Mexico. J. Trop. Ecol. 2005, 21, 651–660. [Google Scholar] [CrossRef]
- Garth, R.E. The ecology of spanish moss (Tillandsia usneoides): Its growth and distribution. Ecology 1964, 45, 470–481. [Google Scholar] [CrossRef]
- Cardelús, C.L.; Chazdon, R.L. Inner-crown microenvironments of two emergent tree species in a lowland wet forest. Biotropica 2005, 37, 238–244. [Google Scholar] [CrossRef]
- Einzmann, H.J.R.; Beyschlag, J.; Hofhansl, F.; Wanek, W.; Zotz, G. Host tree phenology affects vascular epiphytes at the physiological, demographic and community level. AoB Plants 2015, 7, plu73. [Google Scholar] [CrossRef] [Green Version]
- Sarmento Cabral, J.; Petter, G.; Mendieta-Leiva, G.; Wagner, K.; Zotz, G.; Kreft, H. Branchfall as a demographic filter for epiphyte communities: Lessons from forest floor-based sampling. PLoS ONE 2015, 10, e0128019. [Google Scholar] [CrossRef]
- Schlesinger, W.J.; Marks, P.L. Mineral cycling and the niche of Spanish moss, Tillandsia usneoides L. Am. J. Bot. 1977, 64, 1254–1262. [Google Scholar] [CrossRef]
- Martin, C.E.; Lin, T.-C.; Hsu, C.-C.; Lin, S.-H. No effect of host tree species on the physiology of the epiphytic orchid Bulbophyllum japonicum in a subtropical rainforest in Northeastern Taiwan. Taiwan J. For. Sci. 2007, 22, 241–251. [Google Scholar]
- Kattge, J.; Bönisch, G.; Díaz, S.; Lavorel, S.; Prentice, I.C.; Leadley, P.; Tautenhahn, S.; Werner, G.D.A.; Aakala, T.; Abedi, M.; et al. TRY plant trait database—Enhanced coverage and open access. Glob. Chang. Biol. 2020, 26, 119–188. [Google Scholar] [CrossRef] [Green Version]
- Violle, C.; Navas, M.-L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Schellenberger Costa, D.; Zotz, G.; Hemp, A.; Kleyer, M. Trait patterns of epiphytes compared to other plant life forms along a tropical elevation gradient. Funct. Ecol. 2018, 32, 2073–2084. [Google Scholar] [CrossRef]
- Agudelo, C.M.; Benavides, A.M.; Taylor, T.; Feeley, K.J.; Duque, A. Functional composition of epiphyte communities in the Colombian Andes. Ecology 2019, 100, e02858. [Google Scholar] [CrossRef] [PubMed]
- Einzmann, H.J.R.; Schickenberg, N.; Zotz, G. Variation in root morphology of epiphytic orchids along small-scale and large-scale moisture gradients. Acta Bot. Bras. 2020, 34, 66–73. [Google Scholar] [CrossRef]
- Cach-Pérez, M.J.; Andrade, J.L.; Cetzal-Ix, W.; Reyes-García, C. Environmental influence on the inter- and intraspecific variation in the density and morphology of stomata and trichomes of epiphytic bromeliads of the Yucatan Peninsula. Bot. J. Linn. Soc. 2016, 181, 441–458. [Google Scholar] [CrossRef]
- Susan-Tepetlan, T.M.; Velázquez-Rosas, N.; Krömer, T. Cambios en las características funcionales de epífitas vasculares de bosque mesófilo de montana y vegetación secundaria en la región central de Veracruz, México. Bot. Sci. 2015, 93, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Paula Oliveira, R.; Zotz, G.; Wanek, W.; Franco, A.C. Leaf trait co-variation and trade-offs in gallery forest C3 and CAM epiphytes. Biotropica 2021, in press. [Google Scholar] [CrossRef]
- Smith, W.K.; Bell, D.T.; Shepherd, K.A. Associations between leaf structure, orientation, and sunlight exposure in five Western Australian communities. Am. J. Bot. 1998, 85, 56–63. [Google Scholar] [CrossRef]
- Niinemets, Ü. Global-scale climatic controls of leaf dry mass per area, density, and thickness in trees and shrubs. Ecology 2001, 82, 453–469. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Ackerly, D.D. Community assembly and shifts in plant trait distributions across an environmental gradient in coastal California. Ecol. Monogr. 2009, 79, 109–126. [Google Scholar] [CrossRef] [Green Version]
- Peppe, D.J.; Royer, D.L.; Cariglino, B.; Oliver, S.Y.; Newman, S.; Leight, E.; Enikolopov, G.; Fernandez-Burgos, M.; Herrera, F.; Adams, J.M.; et al. Sensitivity of leaf size and shape to climate: Global patterns and paleoclimatic applications. New Phytol. 2011, 190, 724–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Zotz, G.; Schickenberg, N.; Albach, D. The velamen radicum is common among terrestrial monocotyledons. Ann. Bot. 2017, 120, 625–632. [Google Scholar] [CrossRef] [Green Version]
- Sanford, W.W.; Adanlawo, F.L.S.; Adanlawo, I. Velamen and exodermis characters of West African epiphytic orchids in relation to taxonomic grouping and habitat tolerance. Bot. J. Linn. Soc. 1973, 66, 307–321. [Google Scholar] [CrossRef]
- Franks, P.J.; Beerling, D.J. Maximum leaf conductance driven by CO2 effects on stomatal size and density over geologic time. Proc. Natl. Acad. Sci. USA 2009, 106, 10343–10347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hietz, P.; Briones, O. Correlation between water relations and within-canopy distribution of epiphytic ferns in a Mexican cloud forest. Oecologia 1998, 114, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhou, G. Responses of leaf stomatal density to water status and its relationship with photosynthesis in a grass. J. Exp. Bot. 2008, 59, 3317–3325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Körner, C.; Scheel, J.A.; Bauer, H. Maximum leaf diffusive conductance in vascular plants. Photosynthetica 1979, 13, 45–82. [Google Scholar]
- Murray, M.; Soh, W.K.; Yiotis, C.; Batke, S.; Parnell, A.C.; Spicer, R.A.; Lawson, T.; Caballero, R.; Wright, I.J.; Purcell, C.; et al. Convergence in maximum stomatal conductance of C3 woody angiosperms in natural ecosystems across bioclimatic zones. Front. Plant Sci. 2019, 10, 558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruelheide, H.; Dengler, J.; Purschke, O.; Lenoir, J.; Jiménez-Alfaro, B.; Hennekens, S.M.; Botta-Dukát, Z.; Chytrý, M.; Field, R.; Jansen, F.; et al. Global trait–environment relationships of plant communities. Nat. Ecol. Evol. 2018, 2, 1906–1917. [Google Scholar] [CrossRef]
- Marks, C.; Lechowicz, M. Alternative designs and the evolution of functional diversity. Am. Nat. 2006, 167, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Jager, M.M.; Richardson, S.J.; Bellingham, P.J.; Clearwater, M.J.; Laughlin, D.C. Soil fertility induces coordinated responses of multiple independent functional traits. J. Ecol. 2015, 103, 374–385. [Google Scholar] [CrossRef]
- Schellenberger Costa, D.; Gerschlauer, F.; Pabst, H.; Kühnel, A.; Huwe, B.; Kiese, R.; Kuzyakov, Y.; Kleyer, M. Community-weighted means and functional dispersion of plant functional traits along environmental gradients on Mount Kilimanjaro. J. Veg. Sci. 2017, 28, 684–695. [Google Scholar] [CrossRef]
- Grime, J.P. Trait convergence and trait divergence in herbaceous plant communities: Mechanisms and consequences. J. Veg. Sci. 2006, 17, 255–260. [Google Scholar] [CrossRef]
- Laliberté, E.; Norton, D.A.; Scott, D. Contrasting effects of productivity and disturbance on plant functional diversity at local and metacommunity scales. J. Veg. Sci. 2013, 24, 834–842. [Google Scholar] [CrossRef]
- Johansson, D. Ecology of vascular epiphytes in West African rain forests. Acta Phytogeogr. Suec. 1974, 59, 1–129. [Google Scholar]
- Wagner, K.; Zotz, G. Including dynamics in the equation: Tree growth rates and host specificity of vascular epiphytes. J. Ecol. 2020, 108, 761–773. [Google Scholar] [CrossRef]
- Mendieta-Leiva, G.; Zotz, G. A conceptual framework for the analysis of vascular epiphyte assemblages. Perspect. Plant Ecol. Evol. Syst. 2015, 17, 510–521. [Google Scholar] [CrossRef]
- Chave, J.; Coomes, D.; Jansen, S.; Lewis, S.L.; Swenson, N.G.; Zanne, A.E. Towards a worldwide wood economics spectrum. Ecol. Lett. 2009, 12, 351–366. [Google Scholar] [CrossRef]
- Roumet, C.; Birouste, M.; Picon-Cochard, C.; Ghestem, M.; Osman, N.; Vrignon-Brenas, S.; Cao, K.-F.; Stokes, A. Root structure–function relationships in 74 species: Evidence of a root economics spectrum related to carbon economy. New Phytol. 2016, 210, 815–826. [Google Scholar] [CrossRef]
- Ma, Z.; Guo, D.; Xu, X.; Lu, M.; Bardgett, R.D.; Eissenstat, D.M.; McCormack, M.L.; Hedin, L.O. Evolutionary history resolves global organization of root functional traits. Nature 2018, 555, 94. [Google Scholar] [CrossRef] [Green Version]
- Díaz, S.; Kattge, J.; Cornelissen, J.H.C.; Wright, I.J.; Lavorel, S.; Dray, S.; Reu, B.; Kleyer, M.; Wirth, C.; Colin Prentice, I.; et al. The global spectrum of plant form and function. Nature 2016, 529, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B. The world-wide ‘fast–slow’ plant economics spectrum: A traits manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- Westoby, M.; Wright, I.J. Land-plant ecology on the basis of functional traits. Trends Ecol. Evol. 2006, 21, 261–268. [Google Scholar] [CrossRef]
- Weemstra, M.; Mommer, L.; Visser, E.J.W.; Ruijven, J.; Kuyper, T.W.; Mohren, G.M.J.; Sterck, F.J. Towards a multidimensional root trait framework: A tree root review. New Phytol. 2016, 211, 1159–1169. [Google Scholar] [CrossRef] [Green Version]
- Valverde-Barrantes, O.J.; Smemo, K.A.; Blackwood, C.B. Fine root morphology is phylogenetically structured, but nitrogen is related to the plant economics spectrum in temperate trees. Funct. Ecol. 2015, 29, 796–807. [Google Scholar] [CrossRef]
- Westoby, M.; Falster, D.S.; Moles, A.T.; Vesk, P.A.; Wright, I.J. Plant ecological strategies: Some leading dimensions of variation between species. Annu. Rev. Ecol. Syst. 2002, 33, 125–159. [Google Scholar] [CrossRef] [Green Version]
- Paton, S. Meteorological Summary for the San Lorenzo/Fort Sherman Canopy Crane; Smithsonian Tropical Research Institute: Panama City, Panama, 2017; Available online: https://biogeodb.stri.si.edu/physical_monitoring/ (accessed on 1 December 2020).
- Wright, S.J.; Horlyck, V.; Basset, Y.; Barrios, H.; Bethancourt, A.; Bohlman, S.A.; Gilbert, G.S.; Goldstein, G.; Graham, E.A.; Kitajima, K.; et al. Tropical canopy biology program, Republic of Panama. In Studying Forest Canopies from Above: The International Canopy Crane Network; Basset, Y., Horlyck, V., Wright, S.J., Eds.; Smithsonian Tropical Research Institute: Ancon, Panama; United Nations Environmental Programme: New York, NY, USA, 2003; pp. 137–155. [Google Scholar]
- Zotz, G.; Schultz, S. The vascular epiphytes of a lowland forest in Panama-species composition and spatial structure. Plant Ecol. 2008, 195, 131–141. [Google Scholar] [CrossRef]
- Condit, R. Tropical Forest Census Plots; Springer: Berlin, Germany, 1998. [Google Scholar]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- James, S.A.; Bell, D.T. Leaf morphological and anatomical characteristics of heteroblastic Eucalyptus globulus ssp. globulus (Myrtaceae). Aust. J. Bot. 2001, 49, 259–269. [Google Scholar] [CrossRef]
- Zotz, G.; Winkler, U. Aerial roots of epiphytic orchids: The velamen radicum and its role in water and nutrient uptake. Oecologia 2013, 171, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Göbel, C.Y.; Schlumpberger, B.O.; Zotz, G. What Is a pseudobulb? Towards a quantitative definition. Int. J. Plant Sci. 2020, 181, 686–696. [Google Scholar] [CrossRef]
- Luh Huang, C.Y.; Schulte, E.E. Digestion of plant tissue for analysis by ICP emission spectroscopy. Commun. Soil Sci. Plant Anal. 1985, 16, 943–958. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A single-solution method for the determination of soluble phosphate in sea water. J. Mar. Biol. Assoc. UK 1958, 37, 9–14. [Google Scholar] [CrossRef] [Green Version]
- R_Core_Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018.
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Wei, T.; Simko, V. R package corrplot: Visualization of a Correlation Matrix. Statistician 2017, 56, e24. [Google Scholar]
- Le, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Kassambra, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses, R Package Version 1.0.6. Available online: https://rpkgs.datanovia.com/factoextra/index.html (accessed on 7 November 2020).
- Lavorel, S.; Grigulis, K.; McIntyre, S.; Williams, N.S.G.; Garden, D.; Dorrough, J.; Berman, S.; Quetier, F.; Thebault, A.; Bonis, A. Assessing functional diversity in the field—Methodology matters! Funct. Ecol. 2008, 22, 134–147. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P.; Shipley, B. FD: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology. Available online: https://rdrr.io/cran/FD/ (accessed on 1 December 2020).
- Villéger, S.; Mason, N.W.H.; Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herberich, E.; Sikorski, J.; Hothorn, T. A robust procedure for comparing multiple means under heteroscedasticity in unbalanced designs. PLoS ONE 2010, 5, e9788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loranger, J.; Shipley, B. Interspecific covariation between stomatal density and other functional leaf traits in a local flora. Botany 2010, 88, 30–38. [Google Scholar] [CrossRef]
- Kramer-Walter, K.R.; Bellingham, P.J.; Millar, T.R.; Smissen, R.D.; Richardson, S.J.; Laughlin, D.C. Root traits are multidimensional: Specific root length is independent from root tissue density and the plant economic spectrum. J. Ecol. 2016, 104, 1299–1310. [Google Scholar] [CrossRef]
- Zotz, G. The systematic distribution of vascular epiphytes—A critical update. Bot. J. Linn. Soc. 2013, 171, 453–481. [Google Scholar] [CrossRef] [Green Version]
- Hofhansl, F.; Wanek, W.; Drage, S.; Huber, W.; Weissenhofer, A.; Richter, A. Topography strongly affects atmospheric deposition and canopy exchange processes in different types of wet lowland rainforest, Southwest Costa Rica. Biogeochemistry 2011, 106, 371–396. [Google Scholar] [CrossRef]
- Hofhansl, F.; Wanek, W.; Drage, S.; Huber, W.; Weissenhofer, A.; Richter, A. Controls of hydrochemical fluxes via stemflow in tropical lowland rainforests: Effects of meteorology and vegetation characteristics. J. Hydrol. 2012, 452–453, 247–258. [Google Scholar] [CrossRef]
- Craine, J.M.; Elmore, A.J.; Aidar, M.P.M.; Bustamante, M.; Dawson, T.E.; Hobbie, E.A.; Kahmen, A.; Mack, M.C.; McLauchlan, K.K.; Michelsen, A.; et al. Global patterns of foliar nitrogen isotopes and their relationships with climate, mycorrhizal fungi, foliar nutrient concentrations, and nitrogen availability. New Phytol. 2009, 183, 980–992. [Google Scholar] [CrossRef]
- Coste, S.; Baraloto, C.; Leroy, C.; Marcon, E.; Renaud, A.; Richardson, A.D.; Roggy, J.C.; Schimann, H.; Uddling, J.; Herault, B. Assessing foliar chlorophyll contents with the SPAD-502 chlorophyll meter: A calibration test with thirteen tree species of tropical rainforest in French Guiana. Ann. For. Sci. 2010, 67, 607. [Google Scholar] [CrossRef] [Green Version]
- Lambers, H.; Chapin, F.S., III; Pons, T.L. Plant Physiological Ecology, 2nd ed.; Springer: New York, NY, USA, 2008; p. 540. [Google Scholar]
- Kichenin, E.; Wardle, D.A.; Peltzer, D.A.; Morse, C.W.; Freschet, G.T. Contrasting effects of plant inter- and intraspecific variation on community-level trait measures along an environmental gradient. Funct. Ecol. 2013, 27, 1254–1261. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait Type | Abbreviation | Trait | Unit | n | Minimum | Mean | SD | Maximum |
|---|---|---|---|---|---|---|---|---|
| whole plant | Hmax | maximum plant height | cm | 89 | 1 | 46 | 39 | 230 |
| leaf morphological | Lth | leaf thickness | mm | 99 | 0.05 | 0.69 | 0.67 | 4.04 |
| SLA | specific leaf area | mm2 mg−1 | 99 | 2.8 | 18.4 | 13.5 | 66.9 | |
| LDMC | leaf dry matter content | g g−1 | 99 | 0.03 | 0.18 | 0.10 | 0.59 | |
| LAmax | maximum leaf area | cm2 | 94 | 0.1 | 223.6 | 805.7 | 6534.1 | |
| SD | stomatal density | mm−2 | 88 | 12 | 52 | 36 | 193 | |
| SL | stomatal guard cell length | um | 88 | 19 | 38 | 12 | 76 | |
| PSCA | percent stomatal complex area | % | 88 | 1.0 | 3.8 | 1.8 | 8.6 | |
| root morphological | RD | root average diameter | mm | 92 | 0.3 | 1.2 | 1.0 | 6.0 |
| SRL | specific root length | m g−1 | 92 | 0.5 | 18.2 | 27.2 | 193.3 | |
| RTD | root tissue density | g cm−3 | 92 | 0.05 | 0.22 | 0.14 | 0.75 | |
| stem morphological | SSD | stem specific density | mg mm−3 | 87 | 0.05 | 0.22 | 0.12 | 0.63 |
| isotope ratios | δ13C | leaf carbon isotope ratio | ‰ | 95 | −36 | −28 | 6 | −12 |
| δ15N | leaf nitrogen isotope ratio | ‰ | 95 | −5 | −2 | 1 | 2 | |
| leaf chemical | LChl | relative chlorophyll concentration per leaf area | SPAD unit | 87 | 12 | 41 | 12 | 68 |
| LCC | leaf carbon concentration | % | 95 | 34 | 46 | 4 | 62 | |
| LNC | leaf nitrogen concentration | % | 95 | 0.6 | 1.2 | 0.5 | 2.7 | |
| LPC | leaf phosphorus concentration | % | 96 | 0.03 | 0.12 | 0.08 | 0.50 | |
| LKC | leaf potassium concentration | % | 96 | 0.23 | 2.16 | 1.75 | 12.21 | |
| LMgC | leaf magnesium concentration | % | 96 | 0.14 | 0.66 | 0.43 | 2.14 | |
| LCaC | leaf calcium concentration | % | 96 | 0.24 | 1.66 | 1.10 | 4.80 | |
| root chemical | RCC | root carbon concentration | % | 90 | 44 | 51 | 4 | 72 |
| RNC | root nitrogen concentration | % | 90 | 0.19 | 0.87 | 0.55 | 3.53 | |
| RPCs | root phosphorus concentration | % | 89 | 0.01 | 0.07 | 0.04 | 0.18 | |
| RKC | root potassium concentration | % | 89 | 0.09 | 0.99 | 0.87 | 4.34 | |
| RMgC | root magnesium concentration | % | 89 | 0.05 | 0.41 | 0.30 | 1.25 | |
| RCaC | root calcium concentration | % | 89 | 0.04 | 0.87 | 0.83 | 4.45 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wagner, K.; Wanek, W.; Zotz, G. Functional Traits of a Rainforest Vascular Epiphyte Community: Trait Covariation and Indications for Host Specificity. Diversity 2021, 13, 97. https://doi.org/10.3390/d13020097
Wagner K, Wanek W, Zotz G. Functional Traits of a Rainforest Vascular Epiphyte Community: Trait Covariation and Indications for Host Specificity. Diversity. 2021; 13(2):97. https://doi.org/10.3390/d13020097
Chicago/Turabian StyleWagner, Katrin, Wolfgang Wanek, and Gerhard Zotz. 2021. "Functional Traits of a Rainforest Vascular Epiphyte Community: Trait Covariation and Indications for Host Specificity" Diversity 13, no. 2: 97. https://doi.org/10.3390/d13020097
APA StyleWagner, K., Wanek, W., & Zotz, G. (2021). Functional Traits of a Rainforest Vascular Epiphyte Community: Trait Covariation and Indications for Host Specificity. Diversity, 13(2), 97. https://doi.org/10.3390/d13020097