
Production Performance of Moina macrocopa (Straus 1820) (Crustacea, Cladocera) Cultured in Different Salinities: The Effect on Growth, Survival, Reproduction, and Fatty Acid Composition of the Neonates




Abstract
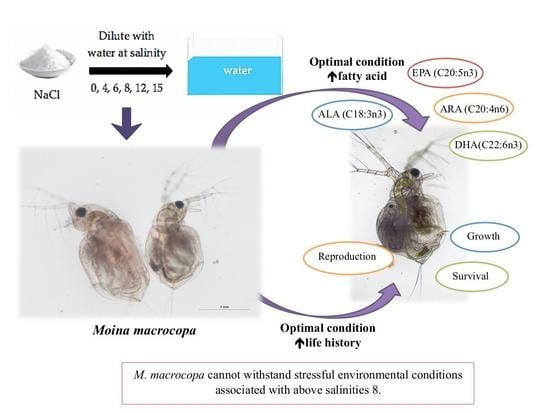
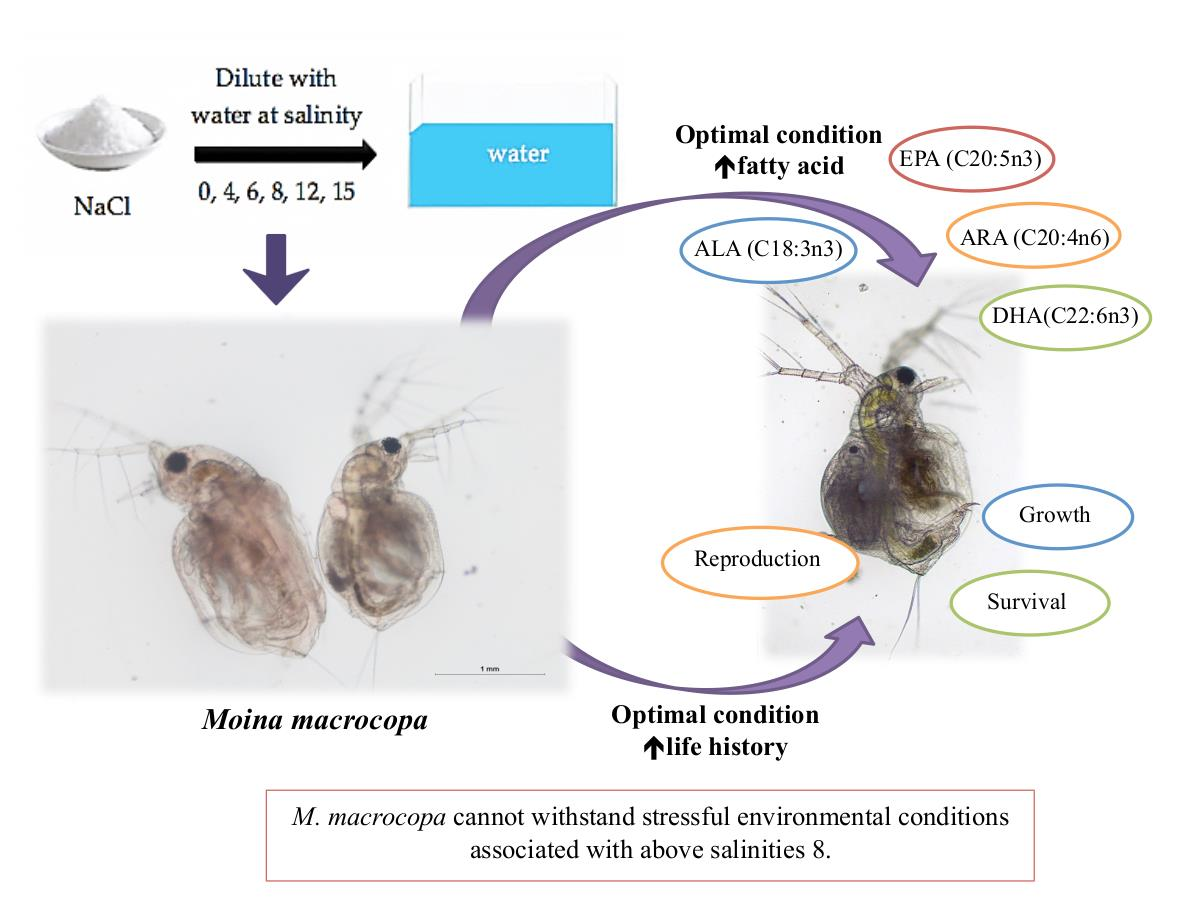
:
1. Introduction
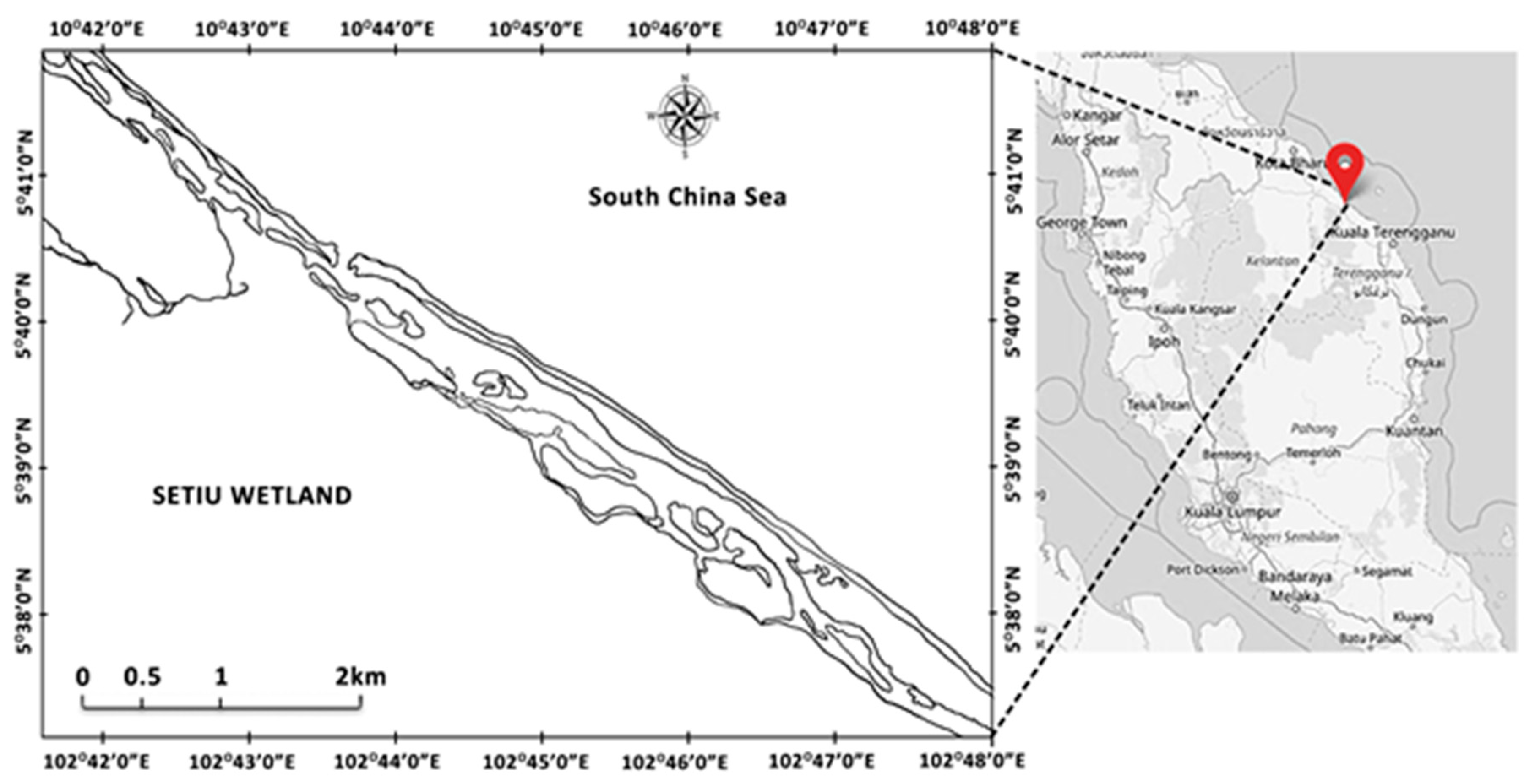
2. Materials and Methods
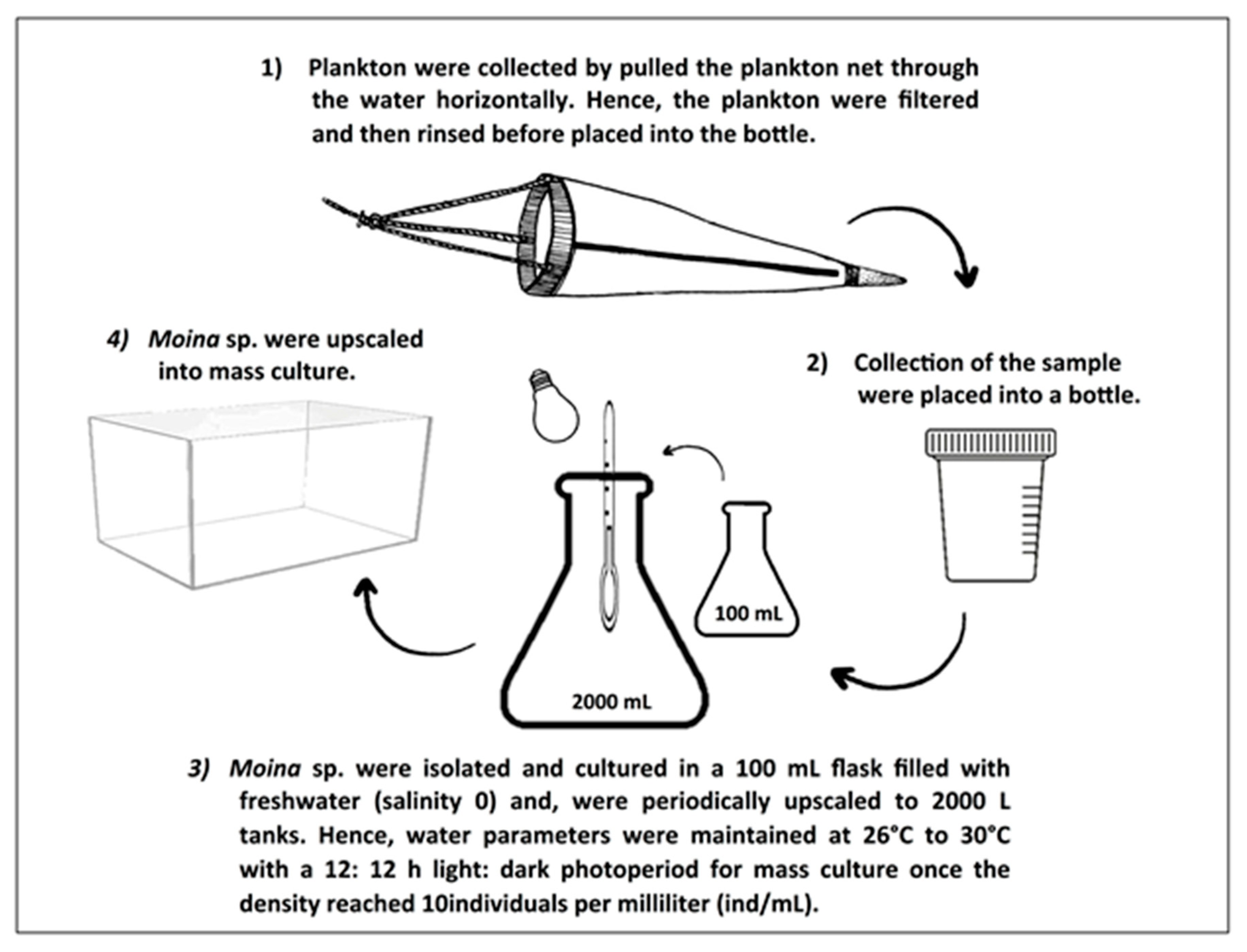
2.1. Cultivation of Moina Macrocopa
2.2. Analysis of Fatty Acid
2.3. Data Analysis
3. Results
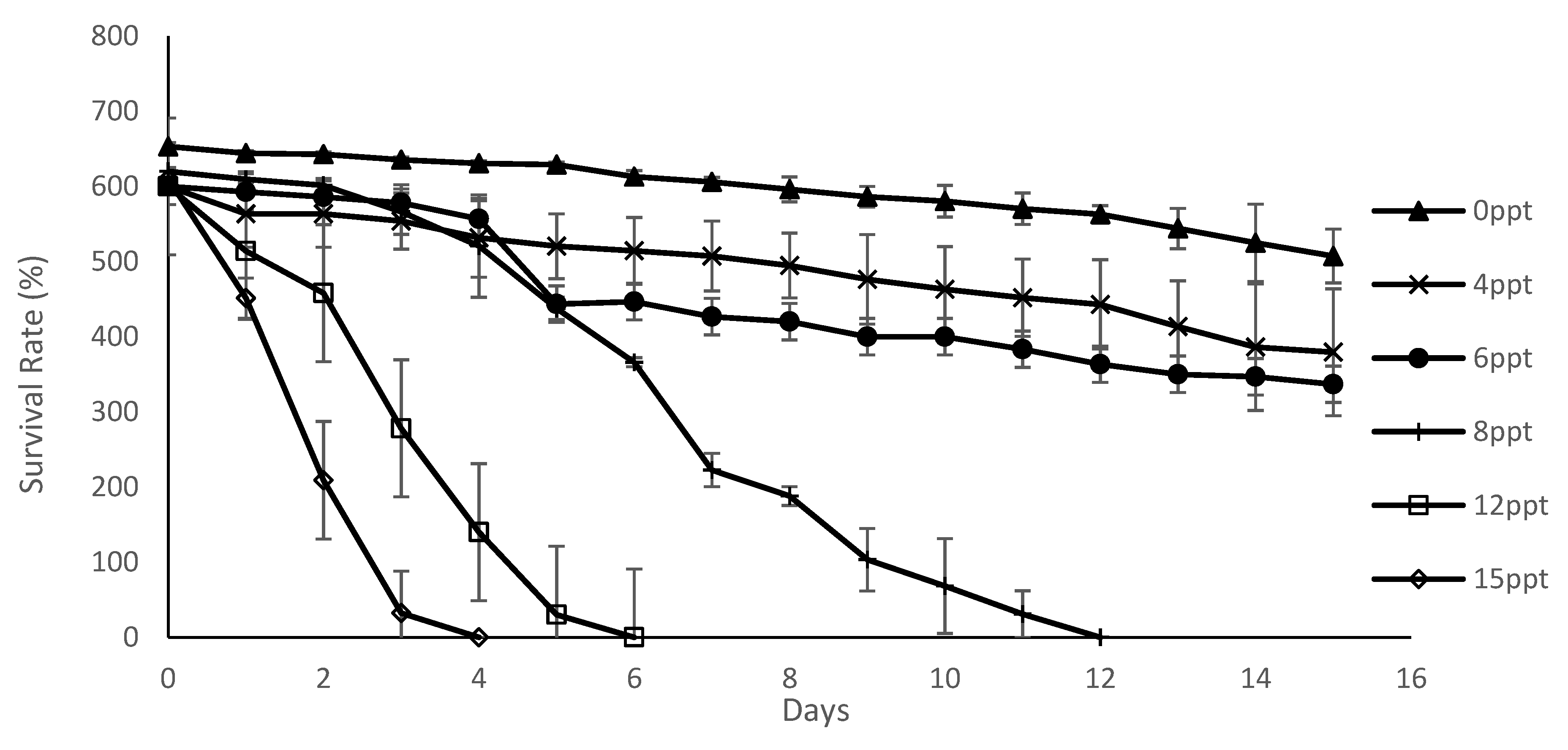
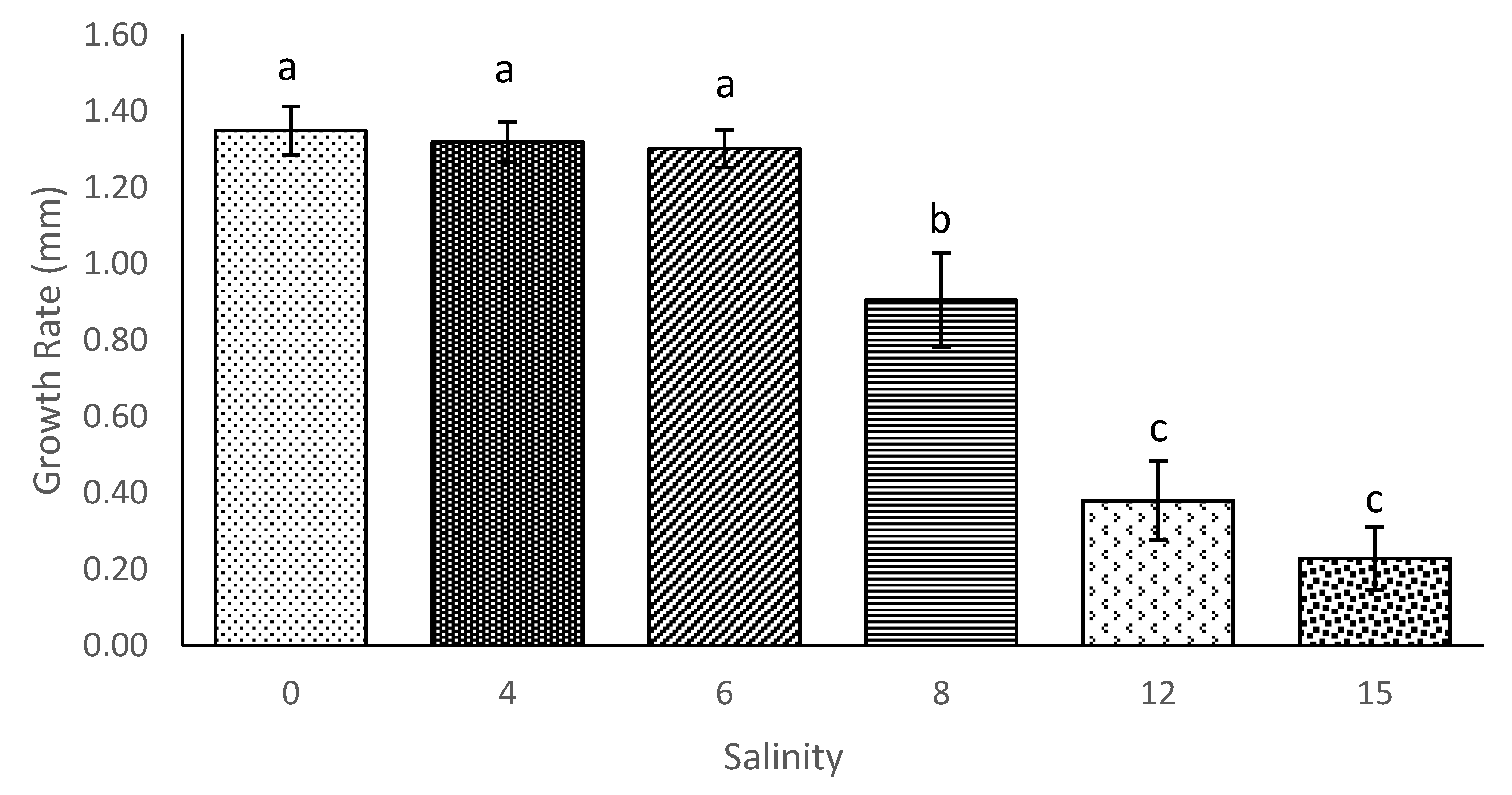
3.1. Survival and Growth of M. Macrocopa in Different Salinities
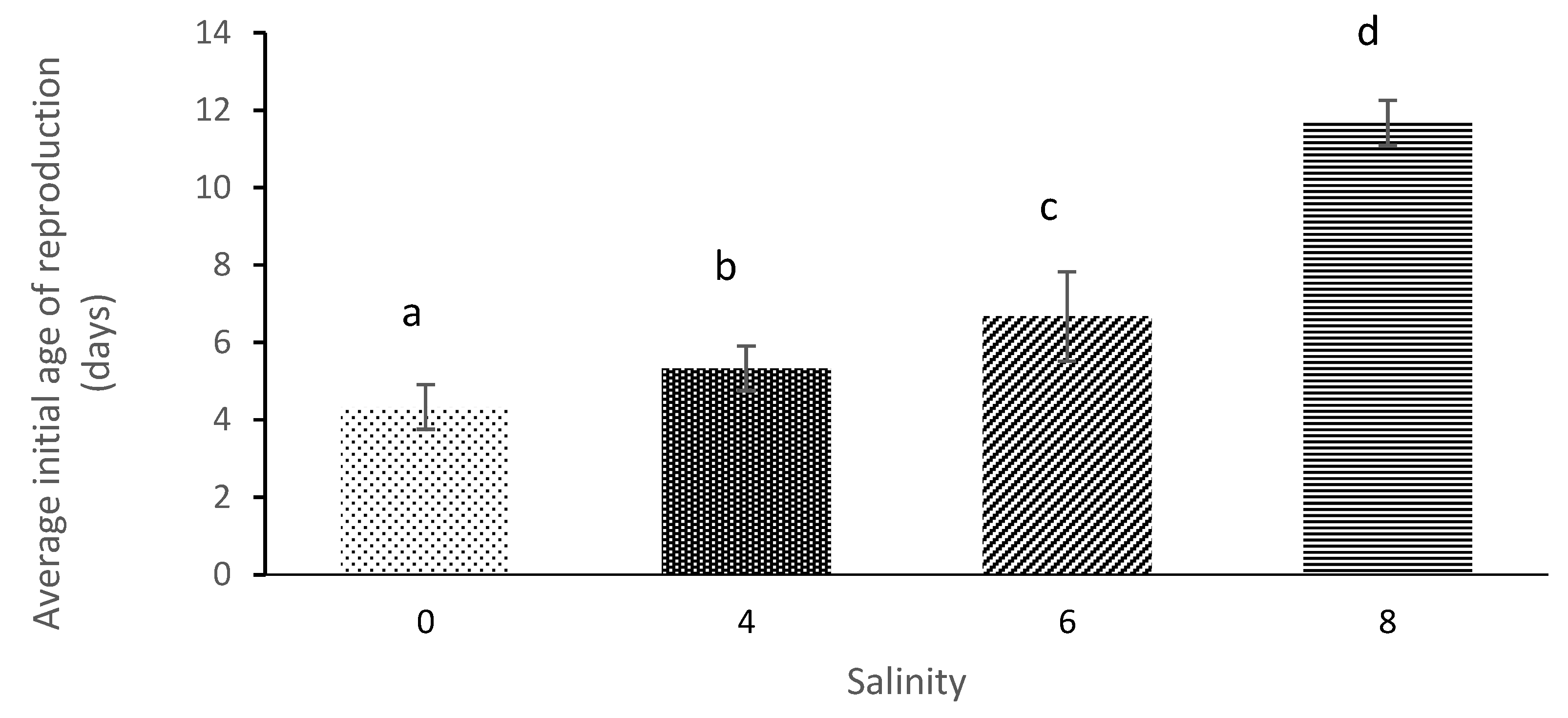
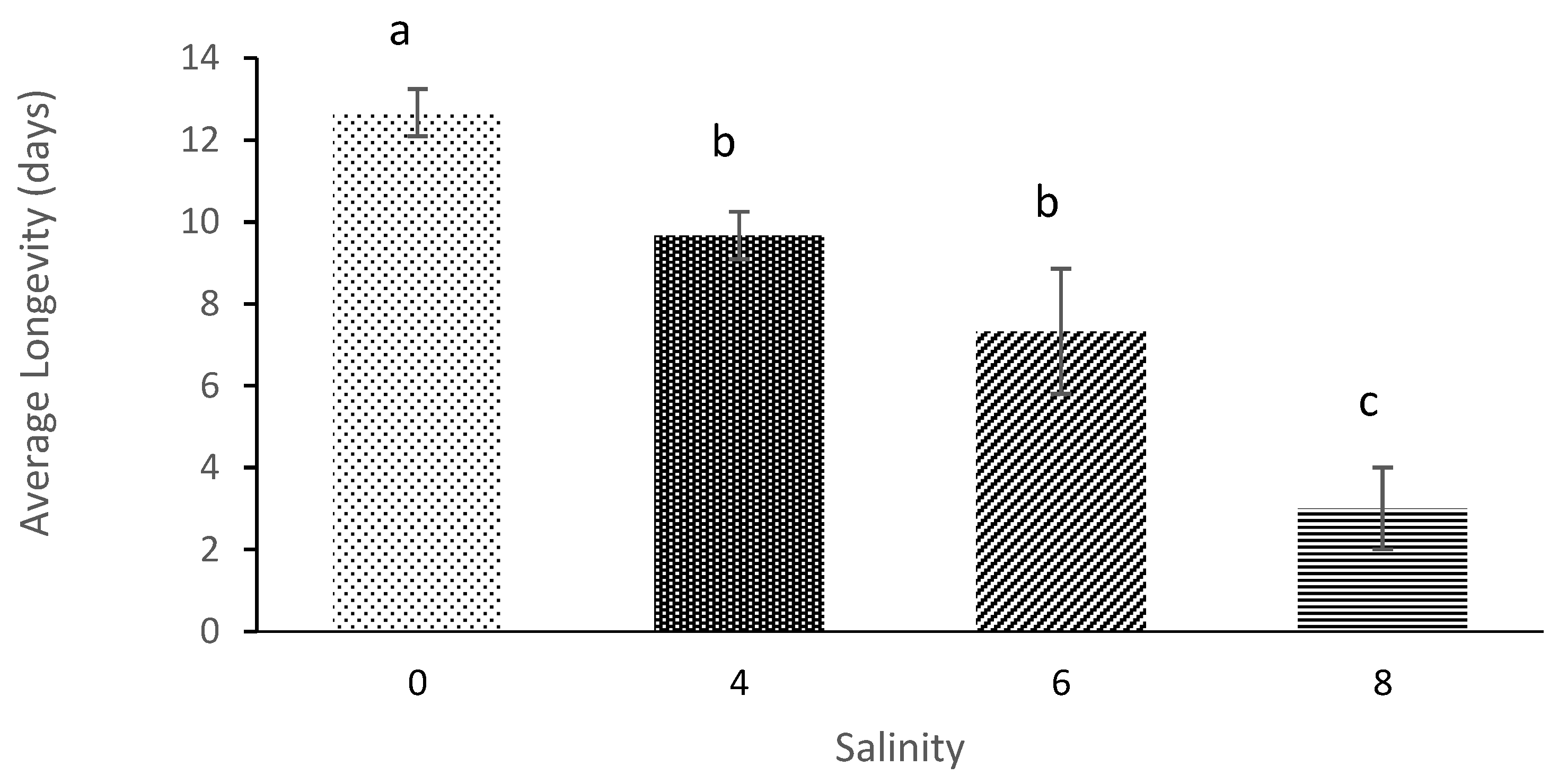
3.2. Age of First Maturation and Average Longevity
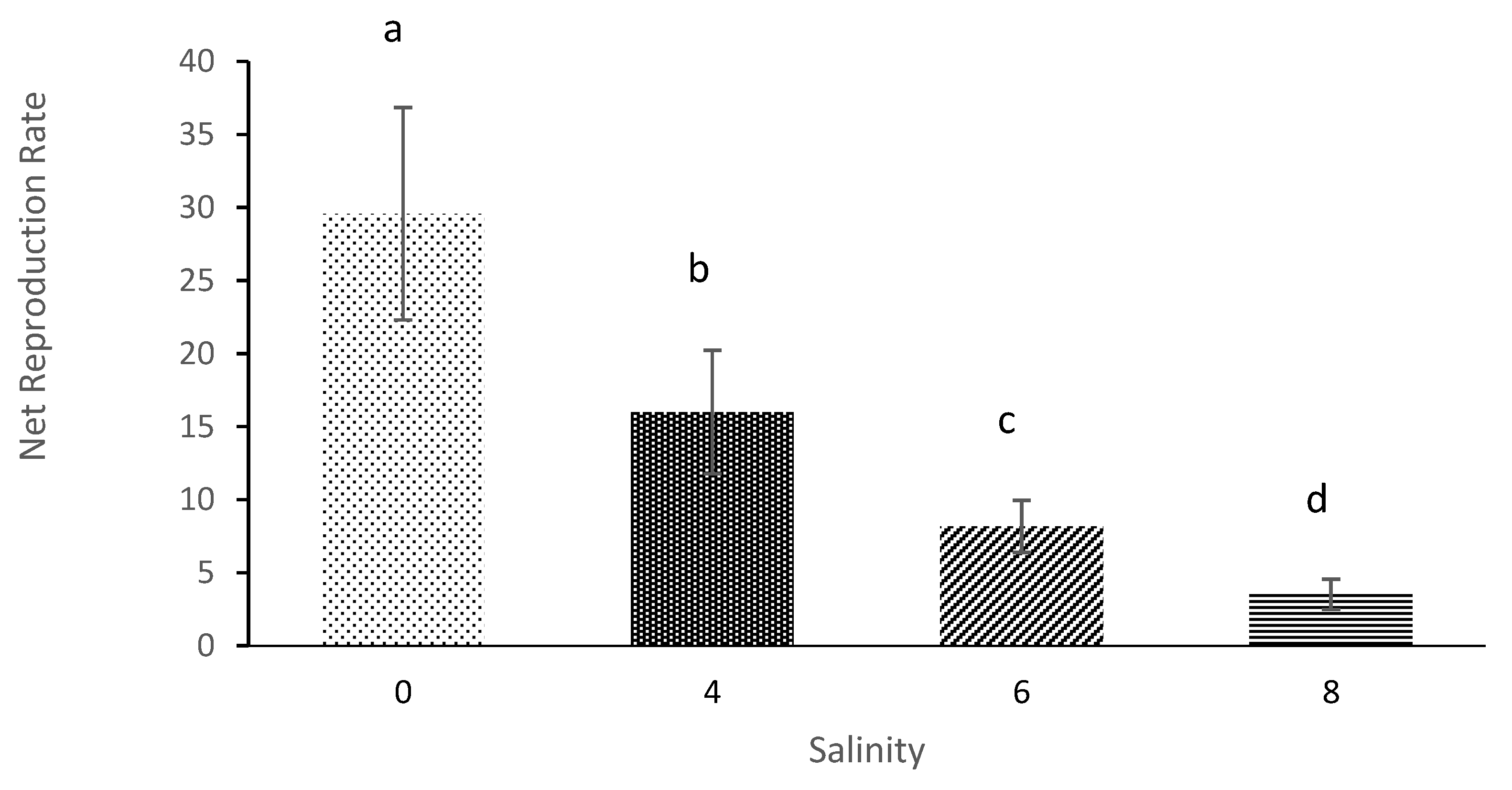
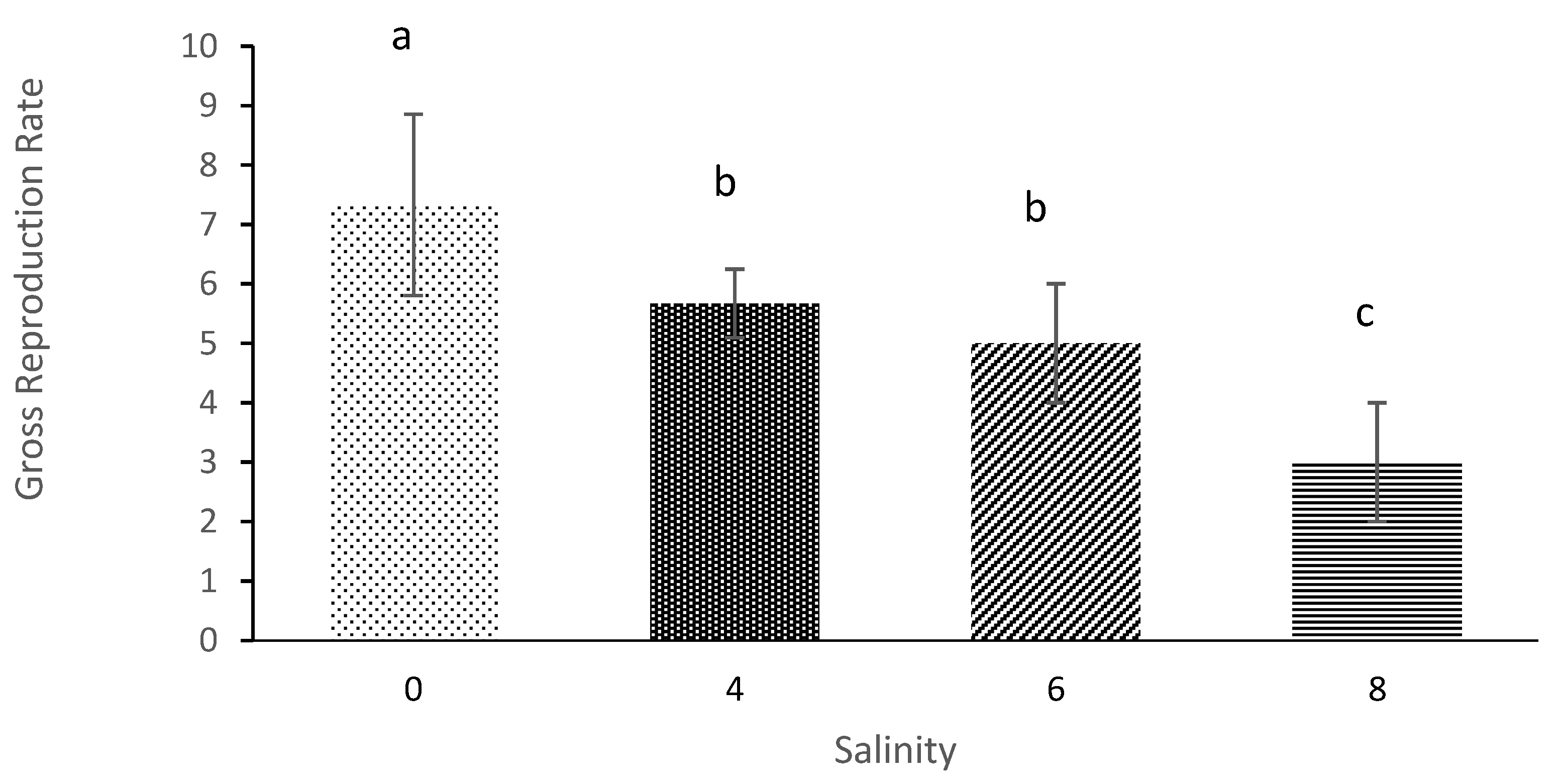
3.3. Net Reproduction Rate and Gross Reproduction Rate
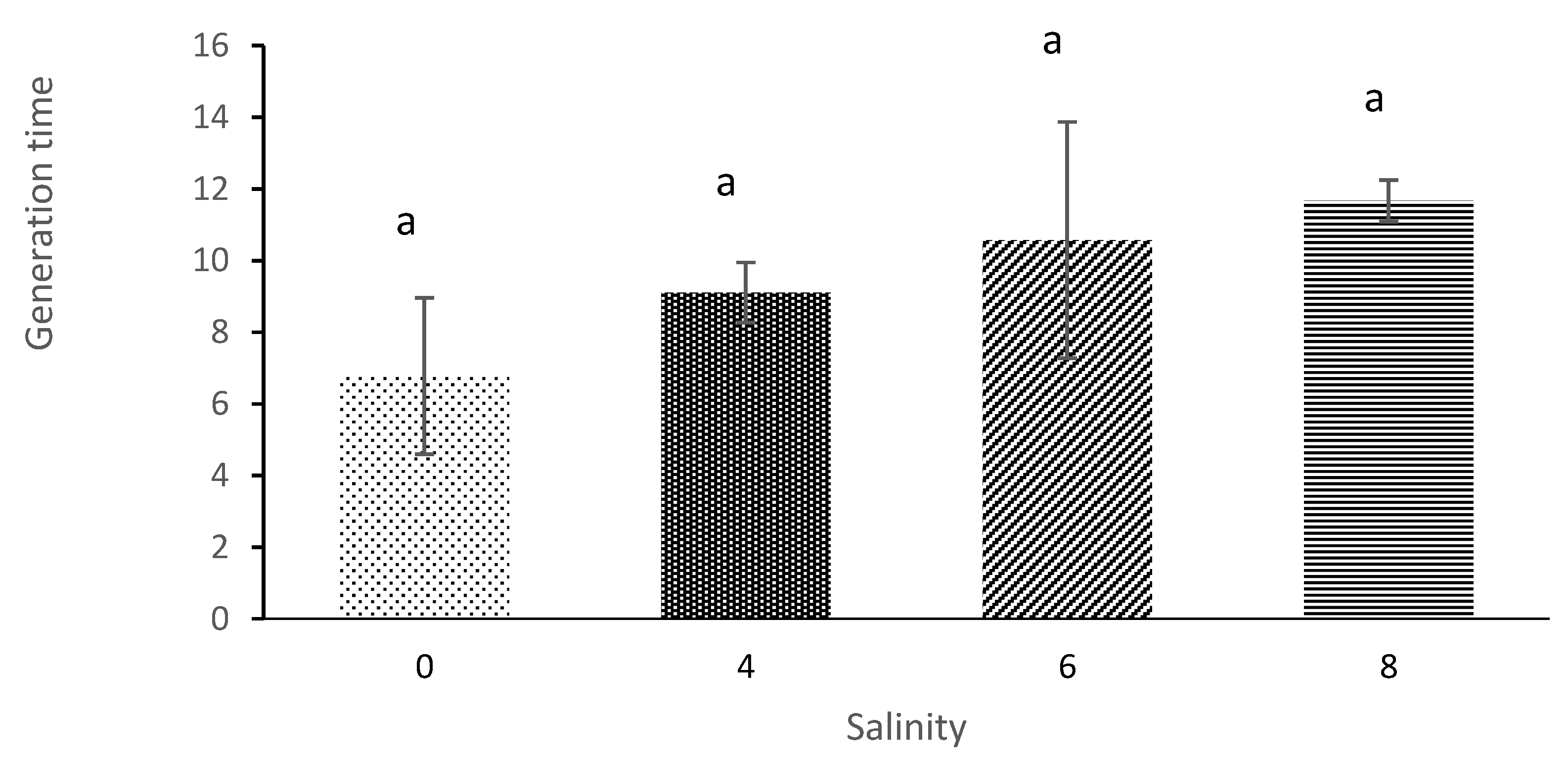
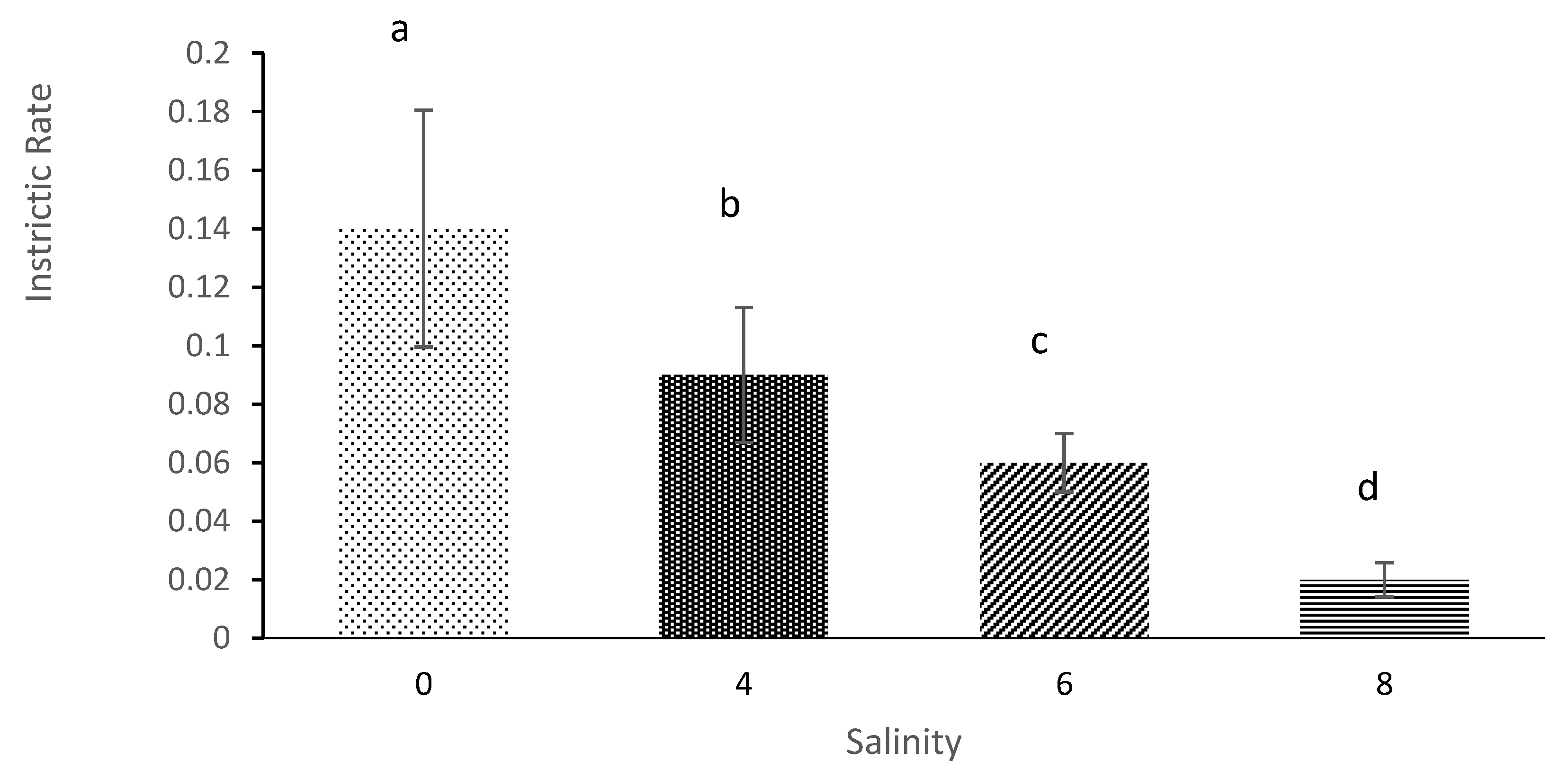
3.4. Generation Time and Intrinsic Rate of Population Increase
3.5. Fatty Acid Compositions of M. Macrocopa
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schallenberg, M.; Hall, C.J.; Burns, C.W. Consequences of climate-induced salinity increases on zooplankton abundance and diversity in coastal lakes. Mar. Ecol. Prog. Ser. 2003, 251, 181–189. [Google Scholar] [CrossRef]
- Suzuki, M.S.; Figueiredo, R.O.; Castro, S.C.; Silva, C.F.; Pereira, E.A.; Silva, J.A.; Aragon, G.T. Sand bar opening in a coastal lagoon (Iquipari) in the northern region of Rio de Janeiro State: Hydrological and hydrochemical changes. Braz. J. Biol. 2002, 62, 51–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santangelo, J.M.; Rocha, A.D.M.; Bozelli, R.L.; Carneiro, L.S.; Esteves, F.D.A. Zooplankton responses to sandbar opening in a tropical eutrophic coastal lagoon. Estuar. Coast. Shelf Sci. 2007, 71, 657–668. [Google Scholar] [CrossRef]
- Suhett, A.L.; Steinberg, C.E.W.; Santangelo, J.M.; Bozelli, R.L.; Farjalla, V.F. Natural dissolved humic subtances increases the lifespan and promote transgenerational resistance to salt stress in the cladoceran Moina macrocopa. Environ. Sci. Pollut. Res. 2011, 18, 1004–1014. [Google Scholar] [CrossRef]
- Vieira, A.C.B.; Medeiros, A.M.A.; Ribeiro, L.L.; Crispim, M.C. Population Dynamics of Moina minuta (Hansen, 1899), Ceriodaphnia cornuta (Sars, 1886), and Diaphanosoma spinulosum (Herbst, 1967) (Crustacean: Branchiopoda) in Different Nutrients (N and P) Concentration Ranges. Acta Limnol. Bras. 2011, 23, 1–9. [Google Scholar] [CrossRef]
- Bezirci, G.; Akkas, S.B.; Rinke, K.; Yildirim, F.; Kalaylioglu, Z.; Severcan, F.; Beklioglu, M. Impacts of salinity and fish-exuded kairomone on the survival and macromolecular profile of Daphnia pulex. Ecotoxicology 2012, 21, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Rasdi, N.W.; Arshad, A.; Ikhwanuddin, M.; Hagiwara, A.; Yusoff, F.M.; Azani, N. A review on the improvement of cladocera (Moina) nutrition as live food for aquaculture: Using valuable plankton fisheries resources. J. Environ. Biol. 2020, 41, 1239–1248. [Google Scholar] [CrossRef]
- Castro, B.B.; Loureiro, C.; Cuo, A.P.; Pedrosa, M.A.; Goncales, F. Life-history responses of salinity-tolerant and salinity-sensitive lineages of stenohalinecladocerans do not confirm clonal differentiation. Hydrobiologia 2012, 702, 73–82. [Google Scholar] [CrossRef]
- Gabriela, C.; Alicia, V.; Santiago, E. Acute effects of Nacl and Na2SO4 on Daphnia menucoensis Paggi, 1996 and Moina eugeniae Olivier, 1954 (Crustacea, Cladocera). Res. Zool. 2014, 4, 8–12. [Google Scholar] [CrossRef]
- Rasdi, N.W.; Qin, J.G. Copepod supplementation as a live food improved growth and survival of Asian seabass Lates calcarifer larvae. Aquac. Res. 2018, 49, 3606–3613. [Google Scholar] [CrossRef]
- Rasdi, N.W.; Qin, J.G. Impact of food type on growth, survival and reproduction of the cyclopoid copepod Cyclopina kasignete as a potential live food in aquaculture. Aquac. Int. 2018, 26, 1281–1295. [Google Scholar] [CrossRef]
- Samat, N.A.; Yusoff, F.M.; Rasdi, N.W.; Karim, M. Enhancement of live food nutritional status with essential nutrients for improving aquatic animal health: A review. Animals 2020, 10, 2457. [Google Scholar] [CrossRef]
- Canedo-Arguelles, M.; Hawkins, C.P.; Kefford, B.J.; Schafer, R.B.; Dyack, B.J.; Brucet, S.; Buchwalter, D.; Dunlop, J.; Fror, O.; Lazorchak, J.; et al. Saving freshwater from salts. Science 2016, 351, 914–916. [Google Scholar] [CrossRef] [PubMed]
- Gokce, D.; Turhan, D.O. Effects of salinity tolerance on survival and life history of 2 cladocerans. Turk. J. Zool. 2014, 38, 347–353. [Google Scholar] [CrossRef]
- Afonina, E.Y.; Tashlykova, N.A. Plankton community and the relationship with the environment in slaine lakes of Onon-Torey plain, Northeastern Mongolia. Saudi J. Biol. Sci. 2017. [Google Scholar] [CrossRef]
- El-Gamal, M.M.; Mona, M.H.; Abdel, R.F.; Salim, H.K.; Nour Eldeen, M.F. Salinity and temperature effect on survival and life history of freshwater cladoceran Daphnia longispina inhabiting Egyptian water. Sci. Afric J. Sci. Issues Res. Essays 2014, 2, 365–374. [Google Scholar]
- Teschner, M. Effects of salinity on the life history and fitness of Daphnia magna: Variability within and between populations. In Cladocera as Model Organisms in Biology; Springer: Dordrecht, The Netherlands, 1995; pp. 33–41. [Google Scholar]
- Freitas, E.C.; Rocha, O. Acute and chronic effects of sodium and potassium on the tropical freshwater cladoceran Pseudosida ramosa. Ecotoxicology 2010, 20, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Sarma, S.S.S.; Nandini, S.; Morales-Ventura, J.; Delgado-Martínez, I.; González-Valverde, L. Effects of NaCl salinity on the population dynamics of freshwater zooplankton (rotifers and cladocerans). Aquat. Ecol. 2006, 40, 349. [Google Scholar] [CrossRef]
- Ismail, S.S.; Ramli, N.H.; Semawi, N.M.; Kadir, M.A.A.; Ali, A.N. Variations in physico-chemical parameters and Chl-a concentration in Setiu Wetlands lagoon during the northeast and inter-monsoon seasons 2018. In Proceedings of the IOP Conference Series: Earth and Environmental Science, Changsha, China, 18–20 September 2020; IOP Publishing: Bristol, UK, 2020; Volume 494, p. 012014. [Google Scholar]
- Martinez-Jeronimo, F.; Martinez-Jeronimo, L. Chronic effect of NaCl salinity on a freshwater strain of Daphnia magna Straus (Crustacea: Cladocera): A demographic study. Ecotoxicol. Environ. Saf. 2007, 67, 411–416. [Google Scholar] [CrossRef]
- Forro Korovchinsky, N.M.; Kotov, A.A.; Petrusek, A. Global diversty of cladoceran (Cladocera; crustacean) in freshwater animal diversity assessment. Hydrobiologia 2008, 595, 177–184. [Google Scholar] [CrossRef]
- Gama-Flores, J.L.; Huidobro-Salas, M.E.; Sarma, S.S.S.; Nandini, S.; Zepeda-Mejia, R.; Gulati, R.D. Temperature and age affect the life history characteristics and fatty acid profiles of Moina macrocopa (Cladocera). J. Therm. Biol. 2015, 53, 135–142. [Google Scholar] [CrossRef]
- Gani, P.; Sunar, N.M.; Matias-Peralta, H.; Abdul Latiff, A.A.; Abdul Razak, A.R. Influence of Initial Cell Concentrations on the Growth Rate and Biomass Productivity of Microalgae in Domestic Wastewater. Appl. Ecol. Environ. Res. 2016, 14, 399–409. [Google Scholar] [CrossRef]
- El-Deeb, G.M.M.; Habashy, M.M.; Kossa, F.I.; Mohammady, E.Y. Effects of salinity on survival, growth and reproduction of the water flea, Daphnia magna. Nat. Sci. 2009, 7, 28–42. [Google Scholar]
- Goncalves AM, M.; Castro, B.B.; Pardal, M.A.; Goncalves, F. Salinity effects on survival and life history of two freshwater cladocerans (Daphnia magna and Daphnia longispina). Ann. Limnol. Int. J. Limnol. 2007, 43, 13–20. [Google Scholar] [CrossRef]
- Ismail, H.N.; Qin, J.G.; Seuront, L. Dietary responses of the brackish cladoceran Daphniopsis australis fed on different algal species. J. Exp. Mar. Biol. Ecol. 2011, 409, 275–282. [Google Scholar] [CrossRef]
- Grzesiuk, M.; Mikulski, A. The effect of salinity on freshwater crustaceans. Pol. J. Ecol. 2006, 54, 669–674. [Google Scholar]
- Haridevan, G.; Jyothibabu, R.; Arunpandi, N.; Jagadesan, L.; Biju, L. Influence of salinity on the life table demography of a rare cladoceran. Environ. Monit. Assess. 2015, 187, 643. [Google Scholar] [CrossRef]
- Kumar, K.H.; Kiran, B.R. A report on diversity of cladocera in sewage fed tank of Bhadravathi taluk, Karnataka. Int. J. Fauna Biol. Stud. 2016, 3, 18–20. [Google Scholar]
- Hardy Elsa, R.; Duncan, A. Food Concentration and Temperature Effects on Life Cycle Characteristics of Tropical Cladocera (Daphnia gessneri Herbst, Diaphanosoma sarsi Richard, Moina reticulate (Daday)): I. Development Time. Acta Amaz. 1994, 24, 119–134. [Google Scholar] [CrossRef] [Green Version]
- Pianka, E.R. Evolutionary Ecology, 4th ed.; Harper and Row: New York, NY, USA, 1988; 468p. [Google Scholar]
- Ichihara, K.; Fukubayashi, Y. Preparation of fatty acid methyl esters for gas-liquid chromatography. J. Lipid Res. 2010, 51, 3635–3640. [Google Scholar] [CrossRef] [Green Version]
- Fuster, I.H.; Retter, C.V.; Sabat, P.; Jiliberto, R.R. Osmoregulatory and demographic responses to salinity of the exotic cladoceran Daphnia exilis. J. Plankton Res. 2010, 32, 1405–1411. [Google Scholar] [CrossRef] [Green Version]
- Iannacone, O.; Alvariño, F. Evaluation of the environmental risk of cartep insecticide in bioassays with three invertebrates. Tech. Agric. 2002, 62, 366–374. [Google Scholar] [CrossRef]
- Coldsnow, K.D.; Matters, B.M.; Hintz, W.D.; Relyea, R.A. Rapid evolution of tolerance to road salt in zooplankton. Environ. Pollut. 2017, 222, 367–373. [Google Scholar] [CrossRef]
- Kefford, B.J.; Palmer, C.G.; Pakhomova, L.; Nugegoda, D. Comparing test systems to measure the salinity tolerance of freshwater invertebrates. Water SA 2004, 30, 499–506. [Google Scholar] [CrossRef] [Green Version]
- Anton-Pardo, M.; Armengol, Z. Effects of salinity and water temporality on zooplankton community in coastal Mediterranean ponds. Estuar. Coast. Shelf Sci. 2012, 114, 93–99. [Google Scholar] [CrossRef]
- Lani, M.N.; Rivan, N.F.M.; Ismail, A.; Omar, W.B.W.; Hassan, Z.; Suhaili, Z. Comparative Study of Physico-Chemical Analyses of Different Water Resources in Setiu Wetland, Terengganu. Food Sci. Technol. 2018, 25–37. [Google Scholar] [CrossRef]
- Patil, V.; Källqvist, T.; Olsen, E.; Vogt, G.; Gislerød, H. Fatty acid composition of 12 microalgae for possible use in aquaculture feed. Aquac. Int. 2007, 15, 1–9. [Google Scholar] [CrossRef]
- Sarma, S.S.S.; Nandini, S. Review of recent ecotoxicological studies on cladocerans. J. Environ. Sci. Health Part B 2006, 41, 1417–1430. [Google Scholar] [CrossRef] [PubMed]
- Püttmann, M.; Krug, H.; von Ochsenstein, E.; Kattermann, R. Fast HPLC Determination of Serum Free Fatty Acids in the Picomole Range. Clin. Chem. 1993, 39, 825–832. [Google Scholar] [CrossRef] [PubMed]
- Tabet, R.; Ayadi, H.; Koken, M.; Leignel, V. Homeostatic responses of crustaceans to salinity changes. Hydrobiologia 2017, 799, 1–20. [Google Scholar] [CrossRef]
- Loureiro, C.; Pereira, J.L.; Pedrosa, M.A.; Gonçalves, F.; Castro, B.B. Competitive outcome of Daphnia-Simocephalus experimental microcosms: Salinity versus priority effects. PLoS ONE 2013, 8, e70572. [Google Scholar] [CrossRef]
- Toruan, R.L. Zooplankton community emerging from fresh and saline wetlands. Ecohydrol. Hydrobiol. 2012, 12, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Bouchnak, R.; Steinberg, C.E.W. Algal diets and natural xenobiotics impact energy allocation in cladocerans. II. Moina macrocopa and Moina micrura. Limnologica 2013, 44, 23–31. [Google Scholar] [CrossRef]
- Rottmann, R.W.; Scott Graves, J.; Watson, C.; Yanong, R.P.E. Culture Techniques of Moina: The Ideal Daphnia for Feeding Freshwater Fish Fry. In UF/IFAS Tropical Aquaculture Laboratory 813; The Institute of Food and Agricultural Sciences IFAS Publishing: Ocala, FL, USA, 1992; pp. 671–5230. [Google Scholar]
- Scharf, F.S.; Manderson, J.P.; Fabriziob, M.C. The effects of seafloor habitat complexity on survival of juvenile fishes: Species-specific interactions with structural refuge. J. Exp. Mar. Biol. Ecol. 2006, 335, 167–176. [Google Scholar] [CrossRef]
- Soni, H.B.; Thomas, S. Occurrence of zooplanktons at sacred palustrine habitat, Central Gujarat, India, with conservation and management strategies. Int. J. Environ. 2014, 3, 111–121. [Google Scholar] [CrossRef] [Green Version]
- Souza, A.T.; Dias, E.; Nogueira, A.; Campos, J.; Marques, J.C.; Martin, I. Population ecology and habitat preferences of juvenile flounder Platichthys flesus (Actinopterygii: Pleuronectidae) in a temperate estuary. J. Sea Res. 2013, 79, 60–69. [Google Scholar] [CrossRef]
- Herbst, D.B. Gradients of salinity stress, environmental stability and water chemistry as a templet for defining habitat types and physiological strategies in inland salt waters. In Saline Lakes; Springer: Dordrecht, The Netherlands, 2001; pp. 209–219. [Google Scholar]
- Hall, C.J.; Burns, C.W. Mortality and growth responses of Daphnia carinata to increases in temperature and salinity. Freshw. Biol. 2001, 47, 451–458. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Temperature (°C) | Dissolved Oxygen (mg/L) | pH | Salinity |
|---|---|---|---|---|
| 1 March 2020 | 28.32 | 4.21 | 7.16 | 6.23 |
| 1 April 2020 | 29.67 | 4.45 | 7.50 | 2.11 |
| 2 May 2020 | 29.33 | 3.89 | 7.24 | 2.08 |
| Salinity | Survival Rate (%) (Mean ± SD) |
|---|---|
| 0 | 91.16 ± 1.67 a |
| 4 | 81.89 ± 8.08 b |
| 6 | 75.31 ± 7.93 c |
| 8 | 43.69 ± 2.20 d |
| 12 | 21.04 ± 1.13 e |
| 15 | 13.35 ± 1.25 e |
| Salinity | Growth Rate (mm) (Mean ± SD) |
|---|---|
| 0 | 1.35 ± 0.28 a |
| 4 | 1.32 ± 0.25 a |
| 6 | 1.30 ± 0.25 a |
| 8 | 0.72 ± 0.61 b |
| 12 | 0.39 ± 0.54 c |
| 15 | 0.23 ± 0.43 c |
| Salinity | Average Initial Age of Reproduction (Days) | Average Longevity (Days) | Net Reproduction Rate | Gross Reproduction Rate | Generation Time | Intrinsic Rate of Population Increases |
|---|---|---|---|---|---|---|
| 0 | 4.33 ± 0.58 a | 12.67 ± 0.58 a | 29.58 ± 7.27 a | 7.33 ± 1.53 a | 6.78 ± 2.18 a | 0.14 ± 0.04 a |
| 4 | 5.33 ± 0.58 b | 9.67 ± 0.58 b | 16.001 ± 4.23 b | 5.67 ± 0.57 b | 9.11 ± 0.84 a | 0.087 ± 0.023 b |
| 6 | 6.67 ± 1.15 c | 7.33 ± 1.53 b | 8.17 ± 1.78 c | 5.00 ± 1.00 c | 10.56 ± 3.29 a | 0.060 ± 0.01 c |
| 8 | 11.67 ± 0.58 d | 3.00 ± 01.00 c | 3.52 ± 1.04 d | 3.00 ± 1.00 d | 11.67 ± 0.577 a | 0.012 ± 0.006 d |
| 12 * | - | - | - | - | - | - |
| 15 * | - | - | - | - | - | - |
| Fatty Acid | Salinities | |||||
|---|---|---|---|---|---|---|
| 0 | 4 | 6 | 8 | 12 | 15 | |
| C14:0 | 3.407 ± 0.189 a | 2.977 ± 0.157 a | 1.789 ± 0.65 b | 0.511 ± 0.168 c | 0.183 ± 0.002 c | 0.008 ± 0.006 c |
| C16:0 | 27.746 ± 2.585 a | 26.220 ± 1.926 a | 19.451 ± 1.534 b | 4.427 ± 1.366 c | 0.017 ± 0.001 d | 0.007 ± 0.004 d |
| C16:1 | 1.200 ± 0.290 a | 0.563 ± 0.235 b | 0.037 ± 0.023 c | 0.012 ± 0.002 c | - | - |
| C18:1 (n9) | 8.867 ± 0.252 a | 5.923 ± 0.484 b | 3.380 ± 1.124 c | 0.920 ± 0.459 d | 0.1037 ± 0.031 d | 0.047 ± 0.014 d |
| C22:1 (n9) | 0.229 ± 0.0325 a | 0.062 ± 0.009 b | 0.007 ± 0.002 c | 0.002 ± 0.002 c | 0.001 ± 0.001 c | - |
| C18:2 (n6) | 24.033 ± 1.952 a | 13.269 ± 2.376 b | 5.070 ± 0.354 c | 1.116 ± 0.703 d | 0.368 ± 0.319 d | 0.055 ± 0.044 d |
| C18:3 (n3)(ALA) | 0.474 ± 0.047 a | 0.135 ± 0.047 b | 0.014 ± 0.020 c | 0.001 ± 0.001 c | - | - |
| C20:4 (n6)(ARA) | 0.616 ± 0.082 a | 0.145 ± 0.364 b | 0.041 ± 0.442 c | 0.006 ± 0.001 d | 0.002 ± 0.002 d | 0.001 ± 0.001 d |
| C20:5 (n3)(EPA) | 0.725 ± 0.047 a | 0.420 ± 0.318 a | 0.004 ± 0.004 b | - | - | - |
| C22:6 (n3)(DHA) | 1.151 ± 0.133 a | 1.173 ± 0.356 a | 0.427 ± 0.300 b | 0.002 ± 0.002 b | 0.001 ± 0.001 b | - |
| Fatty Acid | Nanochloropsis sp. |
|---|---|
| C14:0 | 3.77 ± 1.60 |
| C16:0 | 28.14 ± 2.15 |
| C16:1 | 18.17 ± 2.68 |
| C18:1(n9) | 8.02 ± 1.63 |
| C18:2(n6) | 4.25 ± 0.73 |
| C18:3(n3)(ALA) | 0.84 ± 0.17 |
| C20:4(n6)(ARA) | 5.28 ± 1.05 |
| C20:5(n3)(EPA) | 25.88 ± 3.79 |
| C22:6(n3)(DHA) | 0.49 ± 0.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuslan, A.; Najuwa, S.; Hagiwara, A.; Ghaffar, M.A.; Suhaimi, H.; Rasdi, N.W. Production Performance of Moina macrocopa (Straus 1820) (Crustacea, Cladocera) Cultured in Different Salinities: The Effect on Growth, Survival, Reproduction, and Fatty Acid Composition of the Neonates. Diversity 2021, 13, 105. https://doi.org/10.3390/d13030105
Yuslan A, Najuwa S, Hagiwara A, Ghaffar MA, Suhaimi H, Rasdi NW. Production Performance of Moina macrocopa (Straus 1820) (Crustacea, Cladocera) Cultured in Different Salinities: The Effect on Growth, Survival, Reproduction, and Fatty Acid Composition of the Neonates. Diversity. 2021; 13(3):105. https://doi.org/10.3390/d13030105
Chicago/Turabian StyleYuslan, Amirah, Sharifah Najuwa, Atsushi Hagiwara, Mazlan A. Ghaffar, Hidayu Suhaimi, and Nadiah W. Rasdi. 2021. "Production Performance of Moina macrocopa (Straus 1820) (Crustacea, Cladocera) Cultured in Different Salinities: The Effect on Growth, Survival, Reproduction, and Fatty Acid Composition of the Neonates" Diversity 13, no. 3: 105. https://doi.org/10.3390/d13030105
APA StyleYuslan, A., Najuwa, S., Hagiwara, A., Ghaffar, M. A., Suhaimi, H., & Rasdi, N. W. (2021). Production Performance of Moina macrocopa (Straus 1820) (Crustacea, Cladocera) Cultured in Different Salinities: The Effect on Growth, Survival, Reproduction, and Fatty Acid Composition of the Neonates. Diversity, 13(3), 105. https://doi.org/10.3390/d13030105