
Variation in Assemblages of Freshwater Mussels Downstream of Dams and Dam Removals in the Lake Michigan Basin, Michigan, USA


Abstract
:1. Introduction
2. Materials and Methods
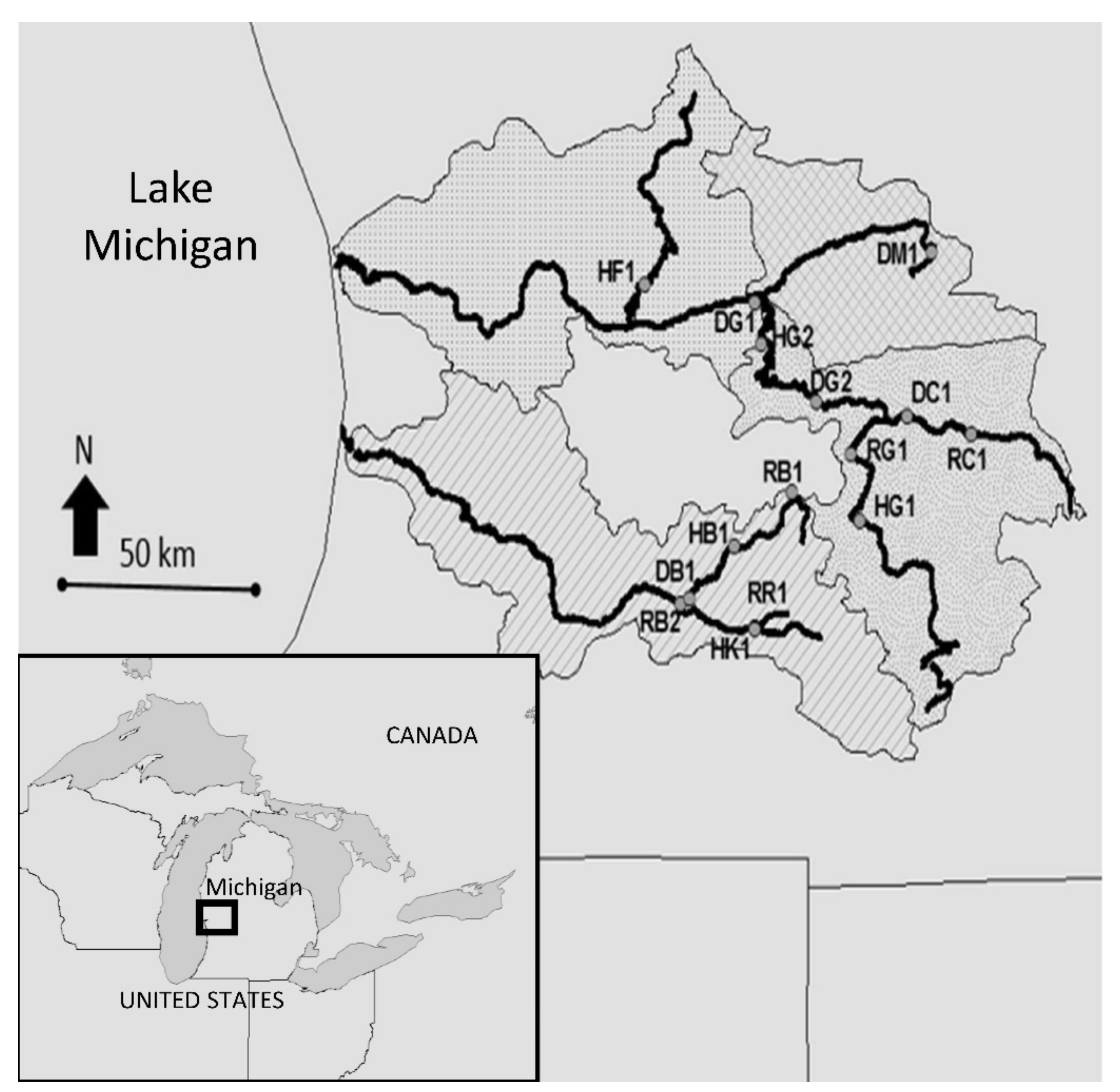
2.1. Study Sites
2.2. Mussel Sampling
2.3. Microhabitat Parameters
2.4. Data Analyses: Comparison of Mussel Assemblages Downstream of Three Anthropogenic Barrier Types
2.5. Data Analyses: Variation of Mussel Assemblages Along a Downstream Gradient from Anthropogenic Barriers
2.6. Data Analyses: Microhabitat Parameters
3. Results
3.1. Comparison of Mussel Assemblages Downstream of Three Anthropogenic Barrier Types
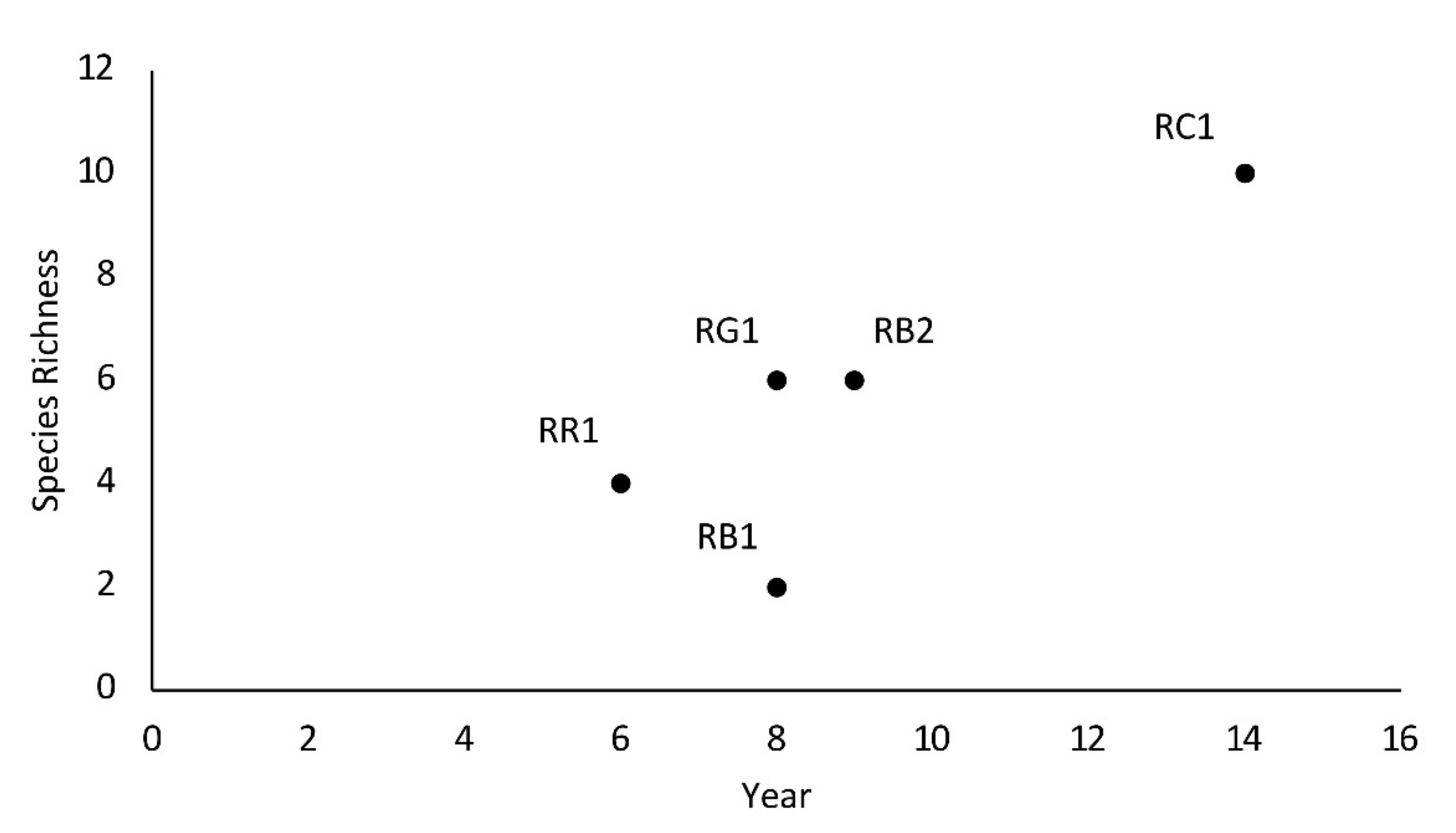
3.2. Variation of Mussel Assemblages Along a Downstream Gradient from Anthropogenic Barriers
3.3. Microhabitat Parameters
4. Discussion
4.1. Mussel Assemblages Downstream of Three Anthropogenic Barrier Types
4.2. Variation of Mussel Assemblages Along a Downstream Gradient from Anthropogenic Barriers
4.3. Management Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baxter, R.M. Environmental effects of dams and impoundments. Annu. Rev. Ecol. Syst. 1977, 8, 255–283. [Google Scholar] [CrossRef]
- Benke, A.C. A perspective on America’s vanishing streams. J. N. Am. Benthol. Soc. 1990, 9, 77–88. [Google Scholar] [CrossRef]
- Dynesius, M.; Nilsson, C. Fragmentation and flow regulation of river systems in the northern third of the world. Science 1994, 266, 753. [Google Scholar] [CrossRef] [PubMed]
- Graf, W.L. Dam nation: A geographic census of American dams and their large-scale hydrologic impacts. Water Resour. Res. 1999, 35, 1305–1311. [Google Scholar] [CrossRef]
- WCD (World Commission on Dams). Dams and Development: A New Framework for Decision-Making; Earthscan Publications: Sterling, VA, USA, 2000. [Google Scholar]
- Anderson, E.P.; Pringle, C.M.; Freeman, M.C. Quantifying the extent of river fragmentation by hydropower dams in the Sarapiqui River Basin, Costa Rica. Aquat. Conserv. 2008, 18, 408–417. [Google Scholar] [CrossRef]
- Bednarek, A.T. Undamming rivers: A review of the ecological impacts of dam removal. Environ. Manag. 2001, 27, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Stanley, E.H.; Doyle, M.W. A geomorphic perspective on nutrient retention following dam removal. Bioscience 2002, 52, 693–701. [Google Scholar] [CrossRef] [Green Version]
- Haslam, S.M. The Riverscape and the River; Cambridge University Press: New York, NY, USA, 2008. [Google Scholar]
- Poff, N.L.; Hart, D.D. How dams vary and why it matters for the emerging science of dam removal. Bioscience 2002, 52, 659–668. [Google Scholar] [CrossRef] [Green Version]
- Kingsford, R.T. Ecological impacts of dams, water diversions and river management on floodplain wetlands in Australia. Austral. Ecol. 2000, 25, 109–127. [Google Scholar] [CrossRef]
- Gangloff, M.M.; Hartfield, E.E.; Werneke, D.C.; Feminella, J.W. Associations between small dams and mollusk assemblages in Alabama streams. J. N. Am. Benthol. Soc. 2011, 30, 1107–1116. [Google Scholar] [CrossRef]
- Singer, E.E.; Gangloff, M.M. Effects of a small dam on freshwater mussel growth in an Alabama (USA) stream. Freshwater Biol. 2011, 56, 1904–1915. [Google Scholar] [CrossRef]
- Haag, W.R. North American Freshwater Mussels: Natural History, Ecology, and Conservation; Cambridge University Press: New York, NY, USA, 2012. [Google Scholar]
- Gangloff, M.M. Taxonomic and ecological tradeoffs associated with small dam removals. Aquat. Conserv. 2013, 23, 475–480. [Google Scholar] [CrossRef]
- Poff, N.L.; Allan, J.D.; Bain, M.B.; Karr, J.R.; Prestegaard, K.L.; Richter, B.D.; Stromberg, J.C. The natural flow regime. Bioscience 1997, 47, 769–784. [Google Scholar] [CrossRef]
- Hoch, R.A. Beaver and Mill Dams Alter Freshwater Mussel Habitat, Growth, and Survival in North Carolina Piedmont Streams. Master’s Thesis, Appalachian State University, Boone, NC, USA, 2012. [Google Scholar]
- McCormick, M.A. Effects of Small Dams on Freshwater Bivalve Assemblages in North Carolina Piedmont and Coastal Plain Streams. Master’s Thesis, Appalachian State University, Boone, NC, USA, 2012. [Google Scholar]
- ICF Consulting. A Summary of Existing Research on Low-Head Dam Removal Projects; American Association of State Highway and Transportation Officials: Washington, DC, USA, 2005. [Google Scholar]
- Csiki, S.J.; Rhoads, B.L. Influence of four run-of-river dams on channel morphology and sediment characteristics in Illinois, USA. Geomorphology 2014, 206, 215–229. [Google Scholar] [CrossRef]
- Wilson, J.B. Mechanisms of species coexistence: Twelve explanations for Hutchinson’s paradox of the plankton: Evidence from New Zealand plant communities. N. Z. J. Ecol. 1990, 13, 17–42. [Google Scholar]
- Cushing, C.E., Jr. Filter-feeding insect distribution and planktonic food in the Montreal River. Trans. Am. Fish. Soc. 1963, 92, 216–219. [Google Scholar] [CrossRef]
- Simmons, G.M., Jr.; Voshell, J.R., Jr. Pre-and post-impoundment benthic macroinvertebrate communities of the North Anna River. In Current Perspectives on River-Reservoir Ecosystems; Cairns, J., Benfield, E.F., Webster, J.R., Eds.; North American Benthological Society: Havana, IL, USA, 1978; pp. 45–61. [Google Scholar]
- Pejchar, L.; Warner, K. A river might run through it again: Criteria for consideration of dam removal and interim lessons from California. Environ. Manag. 2001, 28, 561–575. [Google Scholar]
- Grant, G. Dam removal: Panacea or pandora for rivers? Hydrol. Process. 2001, 15, 1531–1532. [Google Scholar] [CrossRef]
- Hart, D.D.; Johnson, T.E.; Bushaw-Newton, K.L.; Horwitz, R.J.; Bednarek, A.T.; Charles, D.F.; Velinsky, D.J. Dam Removal: Challenges and Opportunities for Ecological Research and River Restoration. Bioscience 2002, 52, 669–682. [Google Scholar] [CrossRef]
- Palmer, M.A.; Bernhardt, E.S.; Allan, J.D.; Lake, P.S.; Alexander, G.; Brooks, S.; Sudduth, E. Standards for ecologically successful river restoration. J. Appl. Ecol. 2005, 42, 208–217. [Google Scholar] [CrossRef]
- Shuman, J.R. Environmental considerations for assessing dam removal alternatives for river restoration. Regul. Rivers Res. Manag. 1995, 11, 249–261. [Google Scholar] [CrossRef]
- Perrin, C.J.; Ashley, K.I.; Larkin, G.A. Effect of drawdown on ammonium and iron concentrations in a coastal mountain reservoir. Water Qual. Res. J. Can. 2000, 35, 231–244. [Google Scholar] [CrossRef]
- Pizzuto, J. Effects of Dam Removal on River Form and Process. Bioscience 2002, 52, 683–691. [Google Scholar] [CrossRef] [Green Version]
- Heinz Center. Dam Removal: Science and Decision Making; The H. John Heinz III Center for Science, Economics, and the Environment: Washington, DC, USA, 2002. [Google Scholar]
- Doyle, M.W.; Harbor, J.M.; Stanley, E.H. Toward policies and decision-making for dam removal. Environ. Manag. 2003, 31, 453–465. [Google Scholar] [CrossRef]
- Kanehl, P.D.; Lyons, J.; Nelson, J.E. Changes in the habitat and fish community of the Milwaukee River, Wisconsin, following removal of the Woolen Mills Dam. N. Am. J. Fish. Manag. 1997, 17, 387–400. [Google Scholar] [CrossRef]
- Smith, L.W.; Dittmer, E.; Prevost, M.; Burt, D.R. Breaching of a small irrigation dam in Oregon: A case history. N. Am. J. Fish. Manag. 2000, 20, 205–219. [Google Scholar] [CrossRef]
- Stanley, E.H.; Doyle, M.W. Trading off: The ecological effects of dam removal. Front. Ecol. Environ. 2003, 1, 15–22. [Google Scholar] [CrossRef]
- Heise, R.J.; Cope, W.G.; Kwak, T.J.; Eads, C.B. Short-term effects of small dam removal on a freshwater mussel assemblage. Walkerana 2013, 16, 41–52. [Google Scholar] [CrossRef]
- Christian, A.D.; Smith, B.N.; Berg, D.J.; Smoot, J.C.; Findlay, R.H. Trophic position and potential food sources of 2 species of unionid bivalves (Mollusca: Unionidae) in 2 small Ohio streams. J. N. Am. Benthol. Soc. 2004, 23, 101–113. [Google Scholar] [CrossRef]
- Vaughn, C.C.; Hakenkamp, C.C. The functional role of burrowing bivalves in freshwater ecosystems. Freshwater Biol. 2001, 46, 1431–1446. [Google Scholar] [CrossRef] [Green Version]
- Spooner, D.E.; Vaughn, C.C. A trait-based approach to species’ roles in stream ecosystems: Climate change, community structure, and material cycling. Oecologia 2008, 158, 307–317. [Google Scholar] [CrossRef]
- Atkinson, C.L.; First, M.R.; Covich, A.P.; Opsahl, S.P.; Golladay, S.W. Suspended material availability and filtration–biodeposition processes performed by a native and invasive bivalve species in streams. Hydrobiologia 2011, 667, 191–204. [Google Scholar] [CrossRef]
- McCall, P.L.; Tevesz, M.J.; Schwelgien, S.F. Sediment mixing by Lampsilis radiata siliquoidea (Mollusca) from western Lake Erie. J. Great Lakes Res. 1979, 5, 105–111. [Google Scholar] [CrossRef]
- Strayer, D.L.; Hunter, D.C.; Smith, L.C.; Borg, C.K. Distribution, abundance, and roles of freshwater clams (Bivalvia, Unionidae) in the freshwater tidal Hudson River. Freshw. Biol. 1994, 31, 239–248. [Google Scholar] [CrossRef]
- Williams, J.D.; Warren, M.L., Jr.; Cummings, K.S.; Harris, J.L.; Neves, R.J. Conservation status of freshwater mussels of the United States and Canada. Fisheries 1993, 18, 6–22. [Google Scholar] [CrossRef]
- Ferreira-Rodriguez, N.; Akiyama, Y.; Aksenova, O.A.; Araujo, R.; Barnhart, C.; Bespalaya, Y.; Bogan, A.; Bolotov, I.; Budha, P.B.; Clavijo, C. Research priorities for freshwater mussel conservation assessment. Biol. Conserv. 2019, 231, 77–87. [Google Scholar] [CrossRef] [Green Version]
- MDNR (Michigan Department of Natural Resources). Michigan’s Official List of Endangered and Threatened Species. 2009. Available online: http://www.michigan.gov/documents/dnr/2007-007_NR_Threatened_Endangered_Species__nonstrike__9-12._274586_7.pdf (accessed on 31 January 2021).
- USFWS (United States Fish and Wildlife Service). Endangered and threatened wildlife and plants; determination of endangered status for the rayed bean and snuffbox mussels throughout their ranges. Fed. Regist. 2012, 77, 8632–8665. [Google Scholar]
- Mulcrone, R.S.; Rathbun, J.E. Pocket Field Guide to the Freshwater Mussels of Michigan, 2nd ed.; Michigan Department of Natural Resources: Lansing, MI, USA, 2020; pp. 1–78.
- Ricciardi, A.; Rasmussen, J.B. Extinction rates of North American freshwater fauna. Conserv. Biol. 1999, 13, 1220–1222. [Google Scholar] [CrossRef]
- Zanatta, D.T.; Mackie, G.L.; Metcalfe-Smith, J.L.; Woolnough, D.A. A refuge for native freshwater mussels (Bivalvia: Unionidae) from impacts of the exotic zebra mussel (Dreissena polymorpha) in Lake St. Clair. J. Great Lakes Res. 2002, 28, 479–489. [Google Scholar] [CrossRef]
- Downing, J.A.; Van Meter, P.; Woolnough, D.A. Suspects and evidence: A review of the causes of extirpation and decline in freshwater mussels. Anim. Biodivers. Conserv. 2010, 33, 151–185. [Google Scholar]
- Walter, W.M. Mollusks of the upper Neuse River basin, North Carolina. J. Elisha Mitchell Sci. Soc. 1956, 72, 262–274. [Google Scholar]
- Hornbach, D.J.; Hove, M.C.; Liu, H.T.; Schenck, F.R.; Rubin, D.; Sansom, B.J. The influence of two differently sized dams on mussel assemblages and growth. Hydrobiologia 2014, 724, 279–291. [Google Scholar] [CrossRef]
- Tiemann, J.S.; Dodd, H.R.; Owens, N.; Wahl, D.H. Effects of lowhead dams on unionids in the Fox River, Illinois. Northeast. Nat. 2007, 14, 125–138. [Google Scholar] [CrossRef]
- Sethi, S.A.; Selle, A.R.; Doyle, M.W.; Stanley, E.H.; Kitchel, H.E. Response of unionid mussels to dam removal in Koshkonong Creek, Wisconsin (USA). Hydrobiologia 2004, 525, 157–165. [Google Scholar] [CrossRef]
- American Rivers. Comprehensive List of Dams Removed: 1999–2019. 2019. Available online: https://www.americanrivers.org/wp-content/uploads/2020/02/DamsRemoved_1999-2019.pdf (accessed on 31 January 2021).
- MDNR (Michigan Department of Natural Resources). Michigan Dam Removal Case Studies. 1999. Available online: https://www.michigan.gov/documents/dnr/damsuccess_513764_7.pdf (accessed on 1 February 2021).
- Metcalfe-Smith, J.L.; Di Maio, J.; Staton, S.K.; Mackie, G.L. Effect of sampling effort on the efficiency of the timed search method for sampling freshwater mussel communities. J. N. Am. Benthol. Soc. 2000, 19, 725–732. [Google Scholar] [CrossRef]
- Strayer, D.L.; Smith, D.R. A Guide to Sampling Freshwater Mussel Populations, 8th ed.; American Fisheries Society: Bethesda, MD, USA, 2003. [Google Scholar]
- Wolman, M.G. A method of sampling coarse river-bed material. EOS 1954, 35, 951–956. [Google Scholar]
- American Public Health Association; American Water Works Association; Water Pollution Control Federation; Water Environment Federation. Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 1995. [Google Scholar]
- Shannon, C.E.; Weaver, W. A mathematical theory of communication. Bell. Syst. Tech. J. 1948, 27, 379–423, 623–656. [Google Scholar] [CrossRef] [Green Version]
- Jost, L. Entropy and diversity. Oikos 2006, 113, 363–375. [Google Scholar] [CrossRef]
- Caceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L.F. Numerical Ecology; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; Ohara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.; Wagner, H. Package ‘Vegan’; R Package: New York, NY, USA, 2015. [Google Scholar]
- Minchin, P.R. An evaluation of the relative robustness of techniques for ecological ordination. Plant Ecol. 1987, 69, 89–107. [Google Scholar] [CrossRef]
- Faith, D.P.; Minchin, P.R.; Belbin, L. Compositional dissimilarity as a robust measure of ecological distance. Vegetatio 1987, 69, 57–68. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Dinno, A. Package ‘Dunn.test’; R Package: New York, NY, USA, 2015. [Google Scholar]
- Zar, J.H. Biostatistical Analysis; Pearson Education: Delhi, India, 1999. [Google Scholar]
- Peterman, R.M. Statistical power analysis can improve fisheries research and management. Can. J. Fish. Aquat. Sci. 1990, 47, 2–15. [Google Scholar] [CrossRef]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S; Springer: New York, NY, USA, 2002. [Google Scholar]
- Zuur, A.; Leno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression; Sage: Newbury Park, CA, USA, 2011. [Google Scholar]
- Myers, R.H. Classical and Modern Regression with Applications; Duxbury Press: Pacific Grove, CA, USA, 1990. [Google Scholar]
- Helms, B.S.; Feminella, J.W.; Pan, S. Detection of biotic responses to urbanization using fish assemblages from small streams of western Georgia, USA. Urban Ecosyst. 2005, 8, 39–57. [Google Scholar] [CrossRef]
- Anderson, D.R.; Burnham, K.P.; Thompson, W.L. Null hypothesis testing: Problems, prevalence, and an alternative. J. Wildl. Manag. 2000, 64, 912–923. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Multimodel inference understanding AIC and BIC in model selection. Soc. Method. Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- Symonds, M.R.; Moussalli, A. A brief guide to model selection, multimodel inference and model averaging in behavioural ecology using Akaike’s information criterion. Behav. Ecol. Sociobiol. 2011, 65, 13–21. [Google Scholar] [CrossRef]
- Strayer, D.L.; Ralley, J. Microhabitat use by an assemblage of stream-dwelling unionaceans (Bivalvia), including two rare species of Alasmidonta. J. N. Am. Benthol. Soc. 1993, 12, 247–258. [Google Scholar] [CrossRef]
- Strayer, D.L. Macrohabitats of freshwater mussels (Bivalvia: Unionacea) in streams of the northern Atlantic Slope. J. N. Am. Benthol. Soc. 1993, 12, 236–246. [Google Scholar] [CrossRef]
- Layzer, J.B.; Madison, L.M. Microhabitat use by freshwater mussels and recommendations for determining their instream flow needs. Regul. River. 1995, 10, 329–345. [Google Scholar] [CrossRef]
- Haag, W.R.; Warren, M.L., Jr. Role of ecological factors and reproductive strategies in structuring freshwater mussel communities. Can. J. Fish. Aquat. Sci. 1998, 55, 297–306. [Google Scholar] [CrossRef]
- McRae, S.E.; Allan, J.D.; Burch, J.B. Reach-and catchment-scale determinants of the distribution of freshwater mussels (Bivalvia: Unionidae) in south-eastern Michigan, USA. Freshw. Biol. 2004, 49, 127–142. [Google Scholar] [CrossRef] [Green Version]
- Strayer, D.L. Freshwater Mussel Ecology: A Multifactor Approach to Distribution and Abundance, 1st ed.; University of California Press: Oakland, CA, USA, 2008. [Google Scholar]
- Petts, G.E. Impounded Rivers: Perspectives for Ecological Management; John Wiley and Sons: Hoboken, NJ, USA, 1985. [Google Scholar]
- Lessard, J.L.; Hayes, D.B. Effects of elevated water temperature on fish and macroinvertebrate communities below small dams. River Res. Appl. 2003, 19, 721–732. [Google Scholar] [CrossRef]
- Merkley, W.B. Impact of red rock reservoir on the Des Moines River. In Current Perspective on River-Reservoir Ecosystems; Cairns, J., Benefield, E.F., Webster, J.R., Eds.; North American Benthological Society: Havana, IL, USA, 1978; pp. 62–76. [Google Scholar]
- Tiemann, J.S.; Douglass, S.A.; Stodola, A.P.; Cummings, K.C. Effects of lowhead dams on freshwater mussels in the Vermilion River Basin, Illinois, with comments on a natural dam removal. Trans. Ill. State Acad. Sci. 2016, 109, 1–7. [Google Scholar]
- Augspurger, T.; Keller, A.E.; Black, M.C.; Cope, W.G.; Dwyer, F.J. Water quality guidance for protection of freshwater mussels (Unionidae) from ammonia exposure. Environ. Toxicol. Chem. 2003, 22, 2569–2575. [Google Scholar] [CrossRef]
- Mummert, A.K.; Neves, R.J.; Newcomb, T.J.; Cherry, D.S. Sensitivity of juvenile freshwater mussels (Lampsilis fasciola, Villosa iris) to total and un-ionized ammonia. Environ. Toxicol. Chem. 2003, 22, 2545–2553. [Google Scholar] [CrossRef]
- Newton, T.J.; Allran, J.W.; O’Donnell, J.A.; Bartsch, M.R.; Richardson, W.B. Effects of ammonia on juvenile unionid mussels (Lampsilis cardium) in laboratory sediment toxicity tests. Environ. Toxicol. Chem. 2003, 22, 2554–2560. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.D.; Wolman, M.G. Downstream Effects of Dams on Alluvial Rivers; Geological Survey Paper 1286, United States Government Printing Office: Washington, DC, USA, 1984. [Google Scholar]
- Vaughn, C.C.; Taylor, C. Impoundments and the decline of freshwater mussels: A case study of an extinction gradient. Con Biol. 1999, 13, 912–920. [Google Scholar] [CrossRef]
- Ligon, F.K.; Dietrich, W.E.; Trush, W.J. Downstream ecological effects of dams. BioScience 1995, 45, 183–192. [Google Scholar] [CrossRef]
- Burroughs, B.A.; Hayes, D.B.; Klomp, K.D.; Hansen, J.F.; Mistak, J. Effects of Strobach Dam removal on fluvial geomorphology in the Pine River, Michigan, United States. Geomorphology 2009, 110, 96–107. [Google Scholar] [CrossRef]
- Simons, R.K.; Simons, D.B. Sediment problems associated with dam removal, Muskegon River, Michigan; Hydraulic Engineering. In Proceedings of the 1991 National Conference of the American Society of Civil Engineers, Nashville, TN, USA, 29 July–2 August 1991; American Society of Civil Engineers: New York, NY, USA, 1991; pp. 680–685. [Google Scholar]
- McLaughlin, R.L.; Smyth, E.R.; Castro-Santos, T.; Jones, M.L.; Koops, M.A.; Pratt, T.C.; Vélez-Espino, L.A. Unintended consequences and trade-offs of fish passage. Fish Fish. 2013, 14, 580–604. [Google Scholar] [CrossRef]
- Jackson, C.R.; Pringle, C.M. Ecological benefits of reduced hydrologic connectivity in intensively developed landscapes. Bioscience 2010, 60, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Nalepa, T.F.; Schloesser, D.W. (Eds.) Quagga and Zebra Mussels: Biology, Impacts, and Control, 2nd ed.; CRC Press: New York, NY, USA, 2014. [Google Scholar]
- Wang, N.; Ingersoll, C.G.; Greer, I.E.; Hardesty, D.K.; Ivey, C.D.; Kunz, J.L.; Barnhart, M.C. Chronic toxicity of copper and ammonia to juvenile freshwater mussels (Unionidae). Environ. Toxicol. Chem. 2007, 26, 2048–2056. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Consbrock, R.A.; Ingersoll, C.G.; Barnhart, M.C. Evaluation of influence of sediment on the sensitivity of a unionid mussel (Lampsilis siliquoidea) to ammonia in 28-day water exposures. Environ. Toxicol. Chem. 2011, 30, 2270–2276. [Google Scholar] [CrossRef]
- USEPA (United States Environmental Protection Agency). United States EPA Aquatic Life Water Quality Criteria for Ammonia-Freshwater 2013; EPA Report 822-R-13-001; US Environmental Protection Agency: Washington, DC, USA, 2013.
- Woolnough, D.A.; Bellamy, A.; Longstaff Hummel, S.; Annis, M. Environmental exposure of freshwater mussels to contaminants of emerging concern: Implications for species conservation. J. Great Lakes Res. 2020, 46, 1625–1638. [Google Scholar] [CrossRef]
- Evans, J.E.; Mackey, S.D.; Gottgens, J.F.; Gill, W.M. Lessons from a dam failure. Ohio J. Sci. 2000, 100, 121–131. [Google Scholar]
- Wang, J.; Ding, L.; Tao, J.; Ding, C.; He, D. The effects of dams on macroinvertebrates: Global trends and insights. River Res. Appl. 2019, 35, 702–713. [Google Scholar]
- Boon, P.J. The impact of river regulation on invertebrate communities in the U.K. River Res. Appl. 1998, 2, 389–409. [Google Scholar] [CrossRef]
- Barbarossa, V.; Schmitt, R.J.P.; Huijbregts, M.A.J.; Zarfl, C.; King, H.; Schipper, A.M. Impacts of current and future large dams on the geographic range connectivity of freshwater fish worldwide. PNAS 2020, 117, 3648–3655. [Google Scholar] [CrossRef] [Green Version]
- Sietman, B.E.; Whitney, S.D.; Kelner, D.E.; Douglas Blodgett, K.; Dunn, H.E. Post-extirpation recovery of the freshwater mussel (Bivalvia: Unionidae) fauna in the Upper Illinois River. J. Freshw. Ecol. 2001, 16, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Morley, S.A.; Foley, M.M.; Duda, J.J.; Beirne, M.M.; Paradis, R.L.; Johnson, R.C.; McHenry, M.L.; Elofson, M.; Sampson, E.M.; McCoy, R.E.; et al. Shifting food web structure during dam removal—Disturbance and recovery during a major restoration action. PLoS ONE 2020, 15, e0239198. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mussel Taxa | L | DG1 | DG2 | DB1 | DM1 | DC1 | HG1 | HG2 | HB1 | HF1 | HK1 | RG1 | RB1 | RB2 | RR1 | RC1 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Actinonaias ligamentina | E | 1 | 1 | 142 | 2 | 2 | 64 | |||||||||
| Alasmidonta marginata (SC) | P | 7 | 3 | 10 | 4 | 5 | 2 | |||||||||
| Alasmidonta viridis (T) | P | 14 | ||||||||||||||
| Amblema plicata | E | 43 | 51 | |||||||||||||
| Cambarunio iris (SC) | P | 8 | 12 | 7 | ||||||||||||
| Cyclonaias pustulosa | E | 5 | 70 | |||||||||||||
| Eurynia dilatata | P | 160 | 11 | 211 | 144 | 29 | 2 | 2 | 3 | 2 | 25 | 943 | ||||
| Epioblasma triquetra (F) | P | 23 | 6 | |||||||||||||
| Fusconaia flava | E | 3 | 86 | 79 | 16 | 17 | 62 | 4 | 2 | 6 | 1 | 21 | ||||
| Lampsilis cardium | P | 29 | 85 | 123 | 15 | 4 | 72 | 51 | 3 | 5 | 2 | 16 | ||||
| Lampsilis siliquoidea | P | 1 | 11 | 44 | 13 | 1 | 10 | 19 | ||||||||
| Lasmigona complanata | O | 264 | 4 | 2 | ||||||||||||
| Lasmigona costata | P | 1 | 2 | 25 | 79 | 45 | 15 | 33 | 9 | 1 | 1 | 25 | ||||
| Ligumia recta (E) | P | 2 | 6 | |||||||||||||
| Ptychobranchus fasciolaris (SC) | E | 12 | ||||||||||||||
| Pyganodon grandis | O | 3 | 1 | 5 | 4 | 1 | 1 | 45 | ||||||||
| Quadrula quadrula | E | 10 | 1 | |||||||||||||
| Strophitus undulatus | P | 10 | 6 | 3 | 3 | 3 | 10 | 9 | 1 | 23 | ||||||
| Utterbackia imbecillis (SC) | O | 2 | ||||||||||||||
| Venustaconcha ellipsiformis (SC) | P | 2 | 22 | 43 | 21 | 54 | 2 | 1 | 58 | |||||||
| Total Species Richness | 3 | 14 | 11 | 15 | 11 | 1 | 0 | 9 | 7 | 7 | 6 | 2 | 6 | 4 | 10 | |
| Total Mussels | 5 | 139 | 785 | 523 | 366 | 1 | 0 | 227 | 265 | 79 | 11 | 4 | 80 | 29 | 1171 | |
| Mean CPUE | 0.4 | 10.3 | 61.9 | 39.5 | 37.3 | NA | NA | 17.5 | 20.2 | 6.3 | 1.5 | 0.3 | 20.0 | 3.7 | 74.3 | |
| Mean Shannon’s Diversity (EXP(H’)) | 2.59 | 7.45 | 5.67 | 8.94 | 4.41 | NA | NA | 3.77 | 5.57 | 3.35 | 3.33 | 1.00 | 2.38 | 1.78 | 2.60 | |
| * see footnote | * | * | * | * | * | * | ** | * |
| Response Variable | Parameters in Model (B) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| SC | SA | GR | CO | BO | BE | DEP | TSS | AIC | ∆AIC | wm | |
| Presence | −1.236 | 0.179 | −0.684 | 0.359 | 590.6 | 0.000 | 0.242 | ||||
| −1.214 | 0.206 | 0.364 | 591.5 | 0.900 | 0.154 | ||||||
| −1.171 | 0.172 | −0.704 | 0.007 | 0.371 | 591.7 | 1.100 | 0.140 | ||||
| −1.136 | 0.137 | 0.239 | −0.625 | 0.363 | 591.7 | 1.100 | 0.140 | ||||
| −1.087 | 0.180 | 0.280 | 0.369 | 592.0 | 1.400 | 0.120 | |||||
| −1.272 | 0.149 | −0.113 | −0.715 | 0.358 | 592.1 | 1.500 | 0.114 | ||||
| −1.241 | 0.175 | −0.133 | −0.692 | 0.360 | 592.6 | 2.000 | 0.089 | ||||
| 628.2 | 37.600 | 0.000 | |||||||||
| Richness | −0.683 | 0.111 | −0.543 | 0.205 | 530.0 | 0.000 | 0.264 | ||||
| −0.707 | 0.093 | −0.073 | −0.563 | 0.205 | 530.1 | 0.100 | 0.251 | ||||
| −0.643 | 0.069 | 0.143 | −0.512 | 0.207 | 531.2 | 1.200 | 0.145 | ||||
| −0.670 | 0.125 | 0.208 | 531.5 | 1.500 | 0.125 | ||||||
| −0.679 | 0.115 | 0.119 | −0.538 | 0.204 | 531.6 | 1.600 | 0.119 | ||||
| −0.687 | 0.111 | −0.542 | 0.000 | 0.204 | 532.0 | 2.000 | 0.097 | ||||
| 560.5 | 30.500 | 0.000 | |||||||||
| Density | −0.725 | 0.120 | 0.114 | −0.625 | 0.261 | 754.8 | 0.000 | 0.317 | |||
| −0.752 | 0.121 | 0.112 | −0.616 | −0.003 | 0.257 | 756.2 | 1.400 | 0.158 | |||
| −0.868 | −0.133 | −0.740 | 0.259 | 756.3 | 1.500 | 0.150 | |||||
| −0.753 | −0.034 | 0.098 | 0.096 | −0.648 | 0.260 | 756.6 | 1.800 | 0.129 | |||
| −0.728 | 0.118 | 0.108 | −0.049 | −0.629 | 0.261 | 756.6 | 1.800 | 0.129 | |||
| −0.869 | −0.132 | −0.136 | −0.746 | 0.259 | 756.8 | 2.000 | 0.117 | ||||
| 804.9 | 0.000 | 0.000 | |||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barnett, S.E.; Woolnough, D.A. Variation in Assemblages of Freshwater Mussels Downstream of Dams and Dam Removals in the Lake Michigan Basin, Michigan, USA. Diversity 2021, 13, 119. https://doi.org/10.3390/d13030119
Barnett SE, Woolnough DA. Variation in Assemblages of Freshwater Mussels Downstream of Dams and Dam Removals in the Lake Michigan Basin, Michigan, USA. Diversity. 2021; 13(3):119. https://doi.org/10.3390/d13030119
Chicago/Turabian StyleBarnett, Shaughn E., and Daelyn A. Woolnough. 2021. "Variation in Assemblages of Freshwater Mussels Downstream of Dams and Dam Removals in the Lake Michigan Basin, Michigan, USA" Diversity 13, no. 3: 119. https://doi.org/10.3390/d13030119
APA StyleBarnett, S. E., & Woolnough, D. A. (2021). Variation in Assemblages of Freshwater Mussels Downstream of Dams and Dam Removals in the Lake Michigan Basin, Michigan, USA. Diversity, 13(3), 119. https://doi.org/10.3390/d13030119