
Increasing Survival of Wild Macaw Chicks Using Foster Parents and Supplemental Feeding



Abstract
:1. Introduction
2. Materials and Methods
2.1. Background Methodology
2.2. Chick Relocation Procedures
2.2.1. Criteria and Timing to Remove Chicks for Relocation Procedures
2.2.2. Removed Chicks’ Initial Conditions
2.2.3. Macaw Chick Rearing in the Nursery
2.2.4. Criteria to Assign Macaw Chicks to Wild Macaw Nests
2.2.5. Criteria to Select Foster Parents
2.2.6. Foster Chick Relocation Procedures and Timing
2.2.7. Foster Chick Relocation Schedule
2.2.8. Observations of Foster Parents/Foster Chick Interactions
2.2.9. Monitoring of Foster Chicks
2.2.10. Supplemental Feeding Plan after Relocation of Foster Chick
2.2.11. Foster Chick Acceptance Criteria
2.2.12. Foster Chick Acceptance Analysis
2.2.13. Chick Feeding Ratios
2.2.14. Foster Chick Growth
2.2.15. Foster Chick Influence on Breeding Success
3. Results
3.1. Foster Chick Acceptance
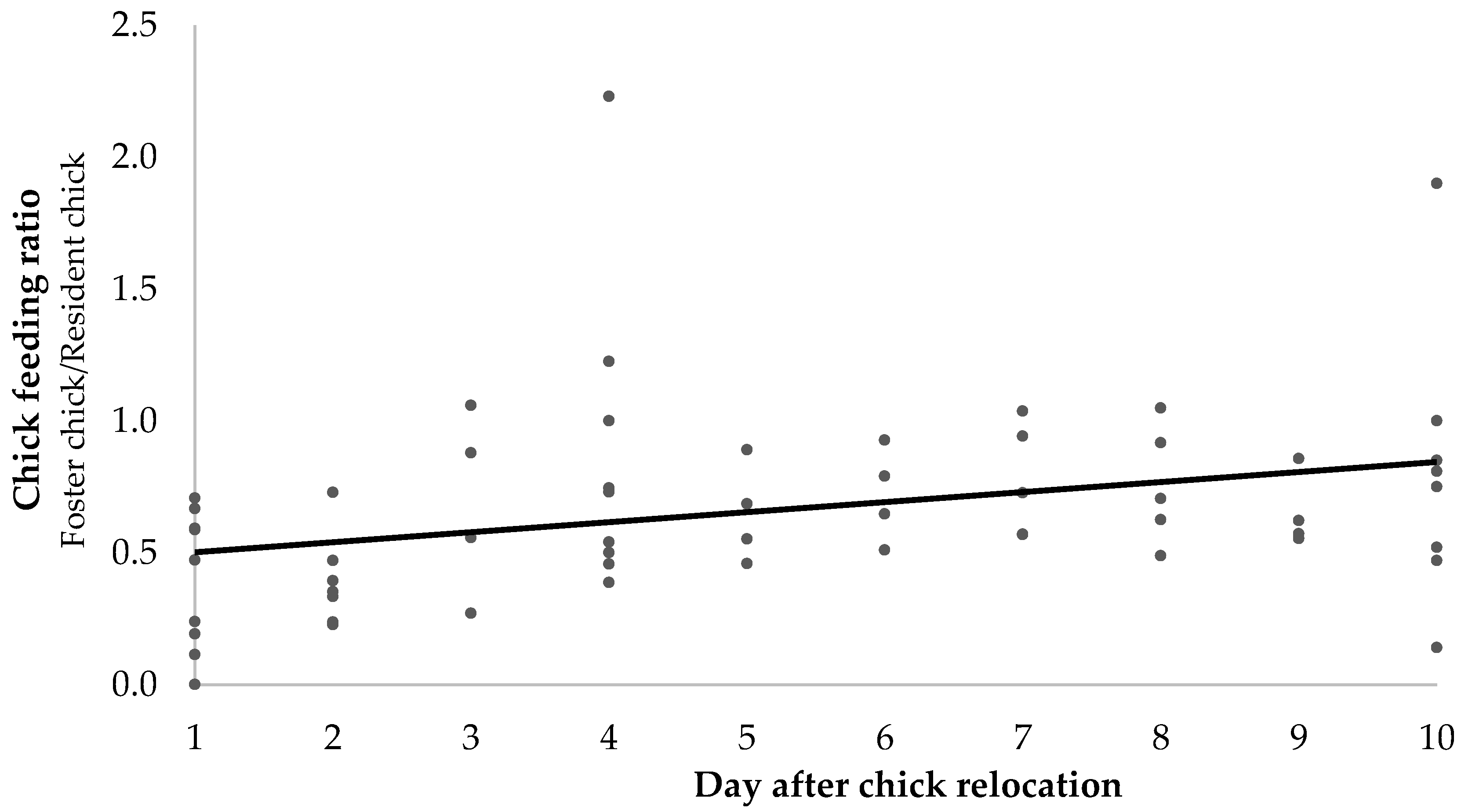
3.2. Foster Chick-Feeding Ratios
3.3. Foster Chick Growth
3.4. Foster Chick Influence on Breeding Success
4. Discussion
4.1. Scarlet Macaws as Foster Parents in the Wild
4.2. Foster Chick Acceptance and Rejection
4.3. Foster Chick Chick-Feeding Rates
4.4. Foster Chick Supplemental Feeding
4.5. Foster Chick Growth Rates
4.6. Testing Starvation Drivers
4.7. “Triple Brood” Chick Relocation Procedure
5. Conclusions
Interdisciplinary Collaboration in Parrot Conservation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Food Name | Age Range | Ingredients | Formula/Water Proportion | |||||
|---|---|---|---|---|---|---|---|---|
| Min (days) | Max (days) | Formula Zupreem Embrace Baby Bird | Formula One Avitech | Peanut Butter | Sheered Seeds | Water | ||
| Neonates Formula | 0 | 9 | Yes | No | No | No | Yes | 1 to 4 |
| Neonates Formula Plus | 2 | 20 | Yes | No | Yes | No | Yes | 1 to 4 |
| Regular Formula | 4 | 43 | Yes | No | No | No | Yes | 1 to 3 |
| Regular Formula Plus | 4 | 74 | Yes | No | Yes | No | Yes | 1 to 3 |
| Regular Formula Extra Plus | 12 | 28 | Yes | No | Yes | Yes | Yes | 1 to 3 |
| Special Emergency Formula | 7 | 13 | No | Yes | No | No | Yes | 1 to 4 |
Appendix B
| Type of Observation | Seasons | # Total Chicks | # Observers | Total Hours Observed | Hours Observed Per Day | |||
|---|---|---|---|---|---|---|---|---|
| Max | Min | Average | St Dev | |||||
| Field Observations | 2018 | 7 | 20 | 417.9 | 12.0 | 4.3 | 8.4 | 2.8 |
| Recorded Observations | 2017 and 2018 | 10 | 1 | 573.4 | 23.6 | 3.7 | 9.0 | 4.2 |
Appendix C
References
- Cade, T.J. The husbandry of falcons for return to the wild. Int. Zoo Yearbook 1980, 20, 3. [Google Scholar] [CrossRef]
- Saint Jalme, M. Endangered avian species captive propagation: An overview of functions and techniques. Avian Poult. Biol. Rev. 1999, 13, 16. [Google Scholar] [CrossRef]
- Fentzloff, C. Breeding, artificial incubation and release of White-tailed sea eagles. Int. Zoo Yearb. 1984, 23, 17. [Google Scholar] [CrossRef]
- Romer, L. Management of the Double-eyed or Red-Browed fig parrot. Zool. Soc. Lond. 2000, 37, 6. [Google Scholar]
- Snyder, N.R.R.; Wiley, J.W.; Kepler, C.B. The Parrots of Luquillo: Natural History and Conservation of the Puerto Rican Parrot; Western Foundation of Vertebrate Zoology: Los Angeles, CA, USA, 1987. [Google Scholar]
- Beissinger, S.R.; Snyder, N.F.R. New World Parrots In Crisis; Smithsonian Institution Press: Washington, DC, USA, 1992. [Google Scholar]
- Beissinger, S.R.; Butcher, E.H. Sustainable harvesting of parrots. In New World Parrots in Crisis; Beissinger, S.R., Snyder, N.F.R., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1992; pp. 73–115. [Google Scholar]
- Rowley, I. Parent-offspring Recognition in a Cockatoo, the Galah Cacatua roseicapilla. Aust. J. Zool. 1980, 28, 9. [Google Scholar] [CrossRef]
- Beissinger, S.R.; Waltman, J.R. Extraordinary clutch size and hatching asynchrony of a Neotropical parrot. Auk 1991, 108, 863–871. [Google Scholar]
- Sanz, V.; Rodriguez-Ferraro, A. Reproductive parameters and productivity of the Yellow-shouldered Parrot on Margarita Island, Venezuela: A long-term study. Condor 2006, 108, 178–192. [Google Scholar] [CrossRef]
- Vaughan, C.; Nemeth, N.; Marineros, L. Ecology and management of natural and artificial scarlet macaw (Ara macao) nest cavities in Costa Rica. Ornithol. Neotrop. 2003, 14, 381–396. [Google Scholar]
- Marineros, L.; Vaughan, C. Scarlet macaws in Carara. In The Large Macaws: Their Care, Breeding and Conservation; Abramson, J., Spear, B.L., Thomsen, J.B., Eds.; Raintree Publications: Ft. Bragg, CA, USA, 1995; pp. 445–468. [Google Scholar]
- Krebs, E. Breeding biology of crimson rosellas (Platycercus elegans) on Black Mountain Australian Capital Territory. Aust. J. Zool. 1998, 46, 17. [Google Scholar] [CrossRef]
- Vigo, G.; Williams, M.; Brightsmith, D.J. Growth of Scarlet Macaw (Ara macao) chicks in southeastern Peru. Neotrop. Ornithol. 2011, 22, 143–153. [Google Scholar]
- Vigo Trauco, G. Crecimiento de Pichones de Guacamayo Escarlata, Ara macao (Linneus: 1758) en la Reserva Nacional Tambopata—Madre de Dios—Perú. Bachelor’s Thesis, Universidad Nacional Agraria La Molina, Lima, Peru, 2007. [Google Scholar]
- Smith, G.A. Systematics of parrots. Ibis 1975, 117, 99. [Google Scholar] [CrossRef]
- Raso, T.D.; Seixas, G.H.F.; Guedes, N.M.R.; Pinto, A.A. Chlamydophila psittaci in free-living Blue-fronted Amazon parrots (Amazona aestiva) and Hyacinth macaws (Anodorhynchus hyacinthinus) in the Pantanal of Mato Grosso do Sul, Brazil. Vet. Microbiol. 2006, 117, 235–241. [Google Scholar] [CrossRef]
- Sanz, V.; Rojas-Suárez, F. Los nidos nodriza como técnica para incrementar el reclutamiento de la cotorra cabeciamarilla (Amazona barbadensis, Aves: Psittacidae). Vida Silv. Neotrop. 1997, 6, 8–14. [Google Scholar]
- Stoodley, A.A. Surrogate Parents. In AFA Watchbid; American Federation of Aviculture: Austin, TX, USA, 1986; Volume 13, p. 6. Available online: https://journals.tdl.org/watchbird/index.php/watchbird/article/view/2455 (accessed on 1 October 2016).
- Dingle, S. Olde Tymer Bill Rattray: King of the King Parrots. In AFA Wathcbird; American Federation of Aviculture: Austin, TX, USA, 1998; Volume 25, Available online: https://journals.tdl.org/watchbird/index.php/watchbird/article/view/1407 (accessed on 1 October 2016).
- Yantz, J. Notes on fostering finches. In AFA Watchbird Magazine; American Federationof Aviculture: Austin, TX, USA, 1986; Available online: https://journals.tdl.org/watchbird/index.php/watchbird/article/view/2299 (accessed on 1 October 2016).
- Krebs, E.A.; Cunningham, R.B.; Donnelly, C.F. Complex patterns of food allocation in asynchronously hatching broods of Crimson Rosellas. Anim. Behavior 1999, 57, 753–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forshaw, J.M. Parrots of the World, 3rd ed.; Landsdowne Editions: Melbourne, Australia, 1989; p. 672. [Google Scholar]
- Iñigo-Elias, E.E. Ecology and Breeding Biology of the Scarlet Macaw (Ara macao) in the Usumacinta Drainage Basin of Mexico and Guatemala. Ph.D. Dissertation, University of Florida, Gainesville, FL, USA, 1996. [Google Scholar]
- Wiedenfeld, D.A. A new subspecies of Scarlet Macaw and its status and conservation. Ornitol. Neotrop. 1994, 5, 99–104. [Google Scholar]
- Portillo Reyes, H. Distribucion actual de la guara (lapa) roja (Ara macao) en Honduras. Zeledonia 2005, 9, 3. [Google Scholar]
- Tella, J.L.; Hiraldo, F. Illegal and legal parrot trade shows a long-term, cross-cultural preference for the most attractive species increasing their risk of extinction. PLoS ONE 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero-Vidal, P.; Hidalgo, F.; Rosetto, F.; Blanco, G.; Carrete, M.; Tella, J.L. Opportunistic or Non-random Wildlife Crime? Attractiveness rather than Abundances in the Wild Leads to Selective Parrot Poaching. Diversity 2020, 12, 314. [Google Scholar] [CrossRef]
- Vigo, G. Scarlet Macaw Nesting Ecology and Behavior: Implications for Conservation Management. Ph.D.Thesis, Texas A&M University, College Station, TX, USA, 2020. [Google Scholar]
- Bentin, M.; Leyva, J.C. Model of assisted breeding of two Primolious couloni chicks by Ara macao adults. AFA Watchbird Magazine, 5 September 2018; Volume XLV, 23–29. [Google Scholar]
- Brightsmith, D.J. Effects of diet, migration, and breeding on clay lick use by parrots in Southeastern Peru. In Proceedings of the Annual Convention 2004, American Federation of Aviculture, San Francisco, CA, USA, 31 March–3 April 2004; pp. 13–14. [Google Scholar]
- Tosi, J.A. Zonas de Vida Natural en el Perú. Memoria Explicativa Sobre el Mapa Ecológico del Perú; Instituto Interamericano de las Ciencias Agricolas de la Organización de los Estados Americanos: Lima, Peru, 1960; p. 271. [Google Scholar]
- Olah, G.; Vigo, G.; Heinsohn, R.; Brightsmith, D.J. Nest site selection and efficacy of artificial nests for breeding success of Scarlet Macaws (Ara macao) in lowland Peru. J. Nat. Conserv. 2014, 22, 176–185. [Google Scholar] [CrossRef]
- Brightsmith, D.J.; Holle, K.M.; Stronza, A. Ecotourism, conservation biology, and volunteer tourism: A mutually beneficial triumvirate. Biol. Conserv. 2008, 141, 2832–2842. [Google Scholar] [CrossRef]
- Gish, E. The Case for Bio-Centric Development: An Ethnographic Study of the Tambopata Macaw Project, Ecotourism and Volunteer Tourism in the Peruvian Amazon. Master’s Dissertation, Wageningen University, Wageningen, The Netherlands, 2009. [Google Scholar]
- Brightsmith, D.J. The Tambopata Macaw Project: Developing techniques to increase reproductive success of large macaws. AFA Watchbird 2001, 28, 24–32. [Google Scholar]
- Perry, D.R.; Williams, J. The tropical rain forest canopy: A method providing total access. Biotropica 1981, 13, 283–285. [Google Scholar] [CrossRef]
- Perry, D.R. A method of access into the crowns of emergent and canopy trees. Biotropica 1978, 10, 155–157. [Google Scholar] [CrossRef]
- Voren, H.; Jordan, R. Parrots: Hand Feeding & Nursery Management; Mattachione, Silvio, & Company: Toronto, ON, Cananda, 1992. [Google Scholar]
- Brightsmith, D.J.; Hilburn, J.; Del Campo, A.; Boyd, J.; Frisius, M.; Frisius, R.; Janik, D.; Guillén, F. The use of hand-raised Psittacines for reintroduction: A case study of Scarlet Macaws (Ara macao) in Peru and Costa Rica. Biol. Conserv. 2005, 121, 465–472. [Google Scholar] [CrossRef]
- Ricklefs, R.E. Patterns of growth in birds. Ibis 1968, 419–451. [Google Scholar] [CrossRef]
- Krebs, E. Begging and food distribution in Crimson Rosella (Platycercus elegans) broods: Why dont hungry chicks beg more? Behav. Ecol. Sociobiol. 2001, 50, 10. [Google Scholar] [CrossRef]
- Renton, K.; Brightsmith, D.J. Cavity use and reproductive success of nesting macaws in lowland forest of southeast Peru. J. Field Ornithol. 2009, 80, 1–8. [Google Scholar] [CrossRef]
- Renton, K. Agonistic interactions of nesting and nonbreeding macaws. Condor 2004, 106, 354–362. [Google Scholar] [CrossRef]
- Stoleson, S.H.; Beissinger, S.R. Hatching asynchrony, brood reduction, and food limitation in a Neotropical parrot. Ecol. Monogr. 1997, 76, 131–154. [Google Scholar] [CrossRef]
- Schew, W.A.; Ricklefs, R.E. Developmental plasticity. In Avian Growth and Development-Evolution within the Altricial-Precocial Spectrum; Starck, M., Ricklefs, R.E., Eds.; Oxford University Press: Oxford, UK, 1998; pp. 288–303. [Google Scholar]
- Brightsmith, D.J.; Matsufuji, D.; McDonald, D.; Bailey, C.A. Nutritional content of free-living Scarlet Macaw chick diets in southeastern Peru. J. Avian Med. Surg. 2010, 24, 9–23. [Google Scholar] [CrossRef]
- Cornejo, J.; Dierenfeld, E.S.; Bailey, C.A.; Brightsmith, D.J. Nutritional and physical characteristics of commercial hand-feeding formulas for parrots. Zoo Biol. 2013. [Google Scholar] [CrossRef]
- Clubb, S.L.; Clubb, K.J.; Skidmore, D.; Wolf, S.; Phillips, A. Psittacine neonatal care and hand-feeding. In Psittacine Aviculture: Perspectives, TECHNIQUES and Research; Shubot, R.M., Clubb, K.J., Clubb, S.L., Eds.; Avicultural Breeding and Research Center: Loxahatchee, FL, USA, 1992. [Google Scholar]
- Myers, M.; Vaughan, C. Movement and behavior of Scarlet Macaws (Ara macao) during the post-fledging dependence period: Implications for in situ versus ex situ management. Biol. Conserv. 2004, 118, 9. [Google Scholar] [CrossRef]
- Boyd, J.; McNab, R.B. The Scarlet Macaw in Guatemala and El Salvador: 2008 status and future possibilities. Findings and Recommendations from a Species Recovery Workshop 9–15 March 2008; Wildlife Conservation Society-Guatemala Program: Guatemala City and Flores City, Peten, Guatemala, 2008; p. 178. [Google Scholar]
- Clubb, S.L. The role of private avicultre in the conservation of Neotropical psittacines. In New World Parrots in Crisis; Snyder, N.F.R., Beissinger, S.R., Eds.; Smithsonian: Washington, DC, USA, 1992; pp. 117–132. [Google Scholar]
- Clubb, K.J.; Clubb, S.L. Status of macaws in aviculture. In Psittacine Aviculture: Perspectives, Techniques and Research; Shubot, R.M., Clubb, K.J., Clubb, S.L., Eds.; Avicultural Breeding and Research Center: Loxahatchee, FL, USA, 1992. [Google Scholar]
- Abramson, J.; Spear, B.L.; Thomsen, J.B. The Large Macaws: Their Care, Breeding and Conservation; Raintree Publications: Fort Bragg, CA, USA, 1995. [Google Scholar]
- Brightsmith, D.J. Parrot nesting in southeastern Peru: Seasonal patterns and keystone trees. Wilson Bull. 2005, 117, 296–305. [Google Scholar] [CrossRef]
- Olah, G.; Vigo, G.; Ortiz, L.; Rozsa, L.; Brightsmith, D.J. Philornis sp. bot fly larvae in free living Scarlet macaw nestlings and a new technique for their extraction. Vet. Parasitol. 2013. [Google Scholar] [CrossRef]
- Juniper, T. Spix’s Macaw: The Race to Save the World’s Rarest Bird; Fourth Estate: London, UK, 2003; p. 296. [Google Scholar]
- Schischakin, N. Special Report: The Spix’s Macaw Conservation Program. AFA Watchb. 1999, 26, 46–55. Available online: https://journals.tdl.org/watchbird/index.php/watchbird/article/view/1398 (accessed on 1 October 2016).



| Hatch Order of Removed Chicks and Criteria for When to Remove Chicks | Timing to Remove Chick from Original Nest | # Chicks Removed |
|---|---|---|
| First Chicks (11 chicks removed) | ||
| When second chick hatched > 3 days after first chick | As soon as second chick hatched | 7 |
| If chick showed signs of life-threatening botfly related infection | When infection was clearly getting worse but still localized in one area | 2 |
| To create conditions for a triple brood by adding a third chick as the younger brood member | When third chick was placed in foster nest | 2 |
| Second Chicks (7 chicks removed) | ||
| If chick showed signs of starvation | When chick was still active and begging. Usually within 3 days of not gaining weight as expected, before they started to lose weight | 5 |
| If chick showed signs of life-threatening botfly related infection | When infection was clearly getting worse but still localized in one area | 1 |
| If chick was needed for relocation to another nest where the clutch or brood was lost due to damage to the nest | As soon as damaged nest was fixed | 1 |
| Third Chicks (12 chicks removed) | ||
| All third chicks were removed | <24 h after hatching | 12 |
| Fourth Chick (2 chicks removed) | ||
| All fourth chicks were removed | <24 h after hatching | 2 |
| Chicks removed from their original nests | 32 | |
| Chicks that perished in our nursery before relocation | 4 | |
| Total chicks relocated | 28 | |
| Developmental Stages in Wild Scarlet Macaw Chicks | ||
|---|---|---|
| Stage | Age Range | Description |
| Stage 1 | 0 to 2 | Hatchling |
| Stage 2A | 3 to 18 | Naked to light pinfeathers and eyes closed |
| Stage 2B | 19 to 33 | Light pinfeathers to heavy pinfeathers and eyes open |
| Stage 3 | 34 to 65 | Heavy pinfeathers to mostly feathered |
| Stage 4 | 66 to fledged | Mostly feathered to fully feathered |
| Macaw Chick Relocation Procedures | Hypothesis | Foster Nest Requirements | Foster Chick Age at Relocation Criterion | Timing to Place Foster Chick | Foster Chick Age at Relocation | # Foster Broods | # Foster Chicks | # Foster Chicks that Fledged | |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Acceptance | Wild macaws will foster unrelated chicks and fledge them as their own | Nest with single resident chick | ≤2 days older or younger than resident chick | Within 24 h of both foster and resident chicks, opening their eyes | ~18 days old | 9 | 11 5 w/video | 9 |
| 2 | Age difference | Age difference is the absolute driver of death by starvation | Nest with single resident chick | From 4 to 9 days older or younger than the resident chick. | Within 48 h of the resident chick opening its eyes | ~18 days old | 9 | 10 7 w/video | 10 |
| 3 | Empty nest | Wild macaws will foster and fledge chicks after losing their own brood or clutch | Nest that lost eggs due to depredation but pair was still incubating an artificial egg | Foster chick was a solo chick | At expected hatching day | <3 days old | 2 | 2 1 w/video | 4 |
| Nest that lost chicks due to depredation or accident (lighting) | Similar developmental stage as lost chicks | <36 h of original brood disappearance | <46 days old | 2 | 3 No video | ||||
| 4 | Triple brood | Wild macaws will accept an additional chick in a brood of two chicks and fledge three chicks. | Nest that hatched three chicks | <5 days between first and third chick | Third chick swapped for first chick when eyes fully opened. | 18 days old | 1 | 2 No video | 2 |
| First chick was placed back in foster nest 5 days after. | 24 days old | ||||||||
| Chick Type | Number of Individuals | Maximum Growth | Growth | Age at Maximum Growth * | |
|---|---|---|---|---|---|
| Rate | |||||
| A | B | C | |||
| (Mean) | (Mean) | (Mean) | |||
| Wild chicks | Solo chick | 17 | 1028 ± 83.2 A | 0.126 ± 0.013 B | 24.7 ± 2.3 D |
| First chick | 38 | 1022 ± 75.4 A | 0.115 ± 0.014 B | 25.7 ± 2.4 E | |
| Second chick | 26 | 993 ± 77.7 A | 0.111 ± 0.015 C | 28.2 ± 3.2 F | |
| All combined | 81 | 1014 ± 79.7 | 0.116 ± 0.016 | 26.3 ± 3 | |
| Foster chicks | 23 | 1020 ± 81.1 A | 0.121 ± 0.014 B | 23.9 ±1.7 D | |
| Breeding Success Parameters | Seasons with Foster Nests (n = 3) | Seasons without Foster Nests (n = 17) | χ2 | p-Value |
|---|---|---|---|---|
| Chicks that fledged per available cavity | 0.43 ± 0.05 | 0.23 ± 0.86 | 7.2 | 0.036 |
| Chicks that fledged per nest with eggs | 1.13 ± 0.15 | 0.56 ± 0.21 | 7.1 | 0.007 |
| Chicks that fledged per nest where at least one chick hatched | 1.5 ± 0.3 | 0.86 ± 0.24 | 7.1 | 0.01 |
| Percentage of younger chicks that starved | 0.06 ± 0.03 | 0.35 ± 0.22 | 5.5 | 0.02 |
| Percentage of chicks that fledged | 0.7 ± 0.05 | 0.46 ± 0.14 | 5.5 | 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vigo-Trauco, G.; Garcia-Anleu, R.; Brightsmith, D.J. Increasing Survival of Wild Macaw Chicks Using Foster Parents and Supplemental Feeding. Diversity 2021, 13, 121. https://doi.org/10.3390/d13030121
Vigo-Trauco G, Garcia-Anleu R, Brightsmith DJ. Increasing Survival of Wild Macaw Chicks Using Foster Parents and Supplemental Feeding. Diversity. 2021; 13(3):121. https://doi.org/10.3390/d13030121
Chicago/Turabian StyleVigo-Trauco, Gabriela, Rony Garcia-Anleu, and Donald J. Brightsmith. 2021. "Increasing Survival of Wild Macaw Chicks Using Foster Parents and Supplemental Feeding" Diversity 13, no. 3: 121. https://doi.org/10.3390/d13030121
APA StyleVigo-Trauco, G., Garcia-Anleu, R., & Brightsmith, D. J. (2021). Increasing Survival of Wild Macaw Chicks Using Foster Parents and Supplemental Feeding. Diversity, 13(3), 121. https://doi.org/10.3390/d13030121