
Still Digging: Advances and Perspectives in the Study of the Diversity of Several Sedentarian Annelid Families


Abstract
:1. Introduction
2. Acrocirridae Banse, 1969
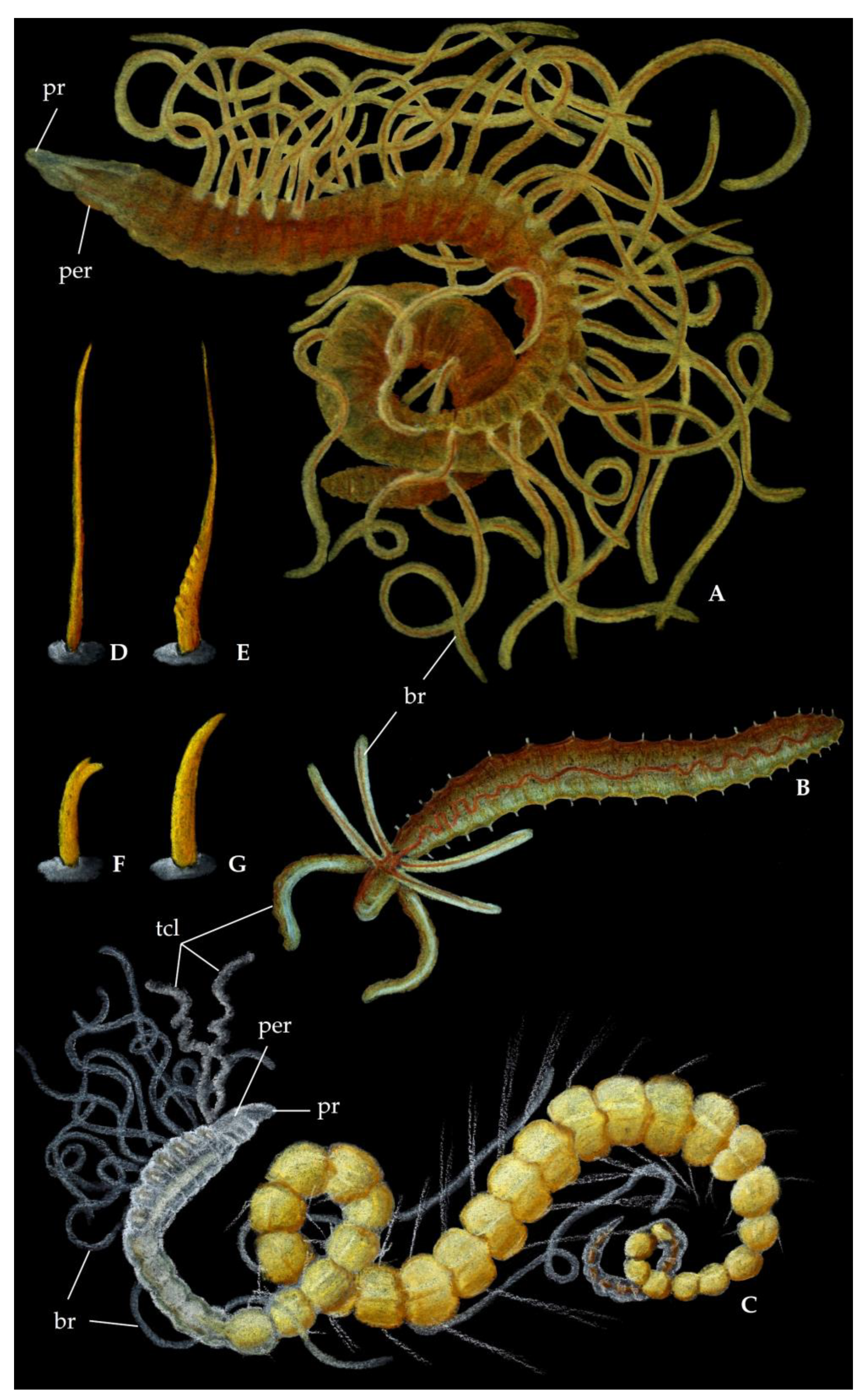
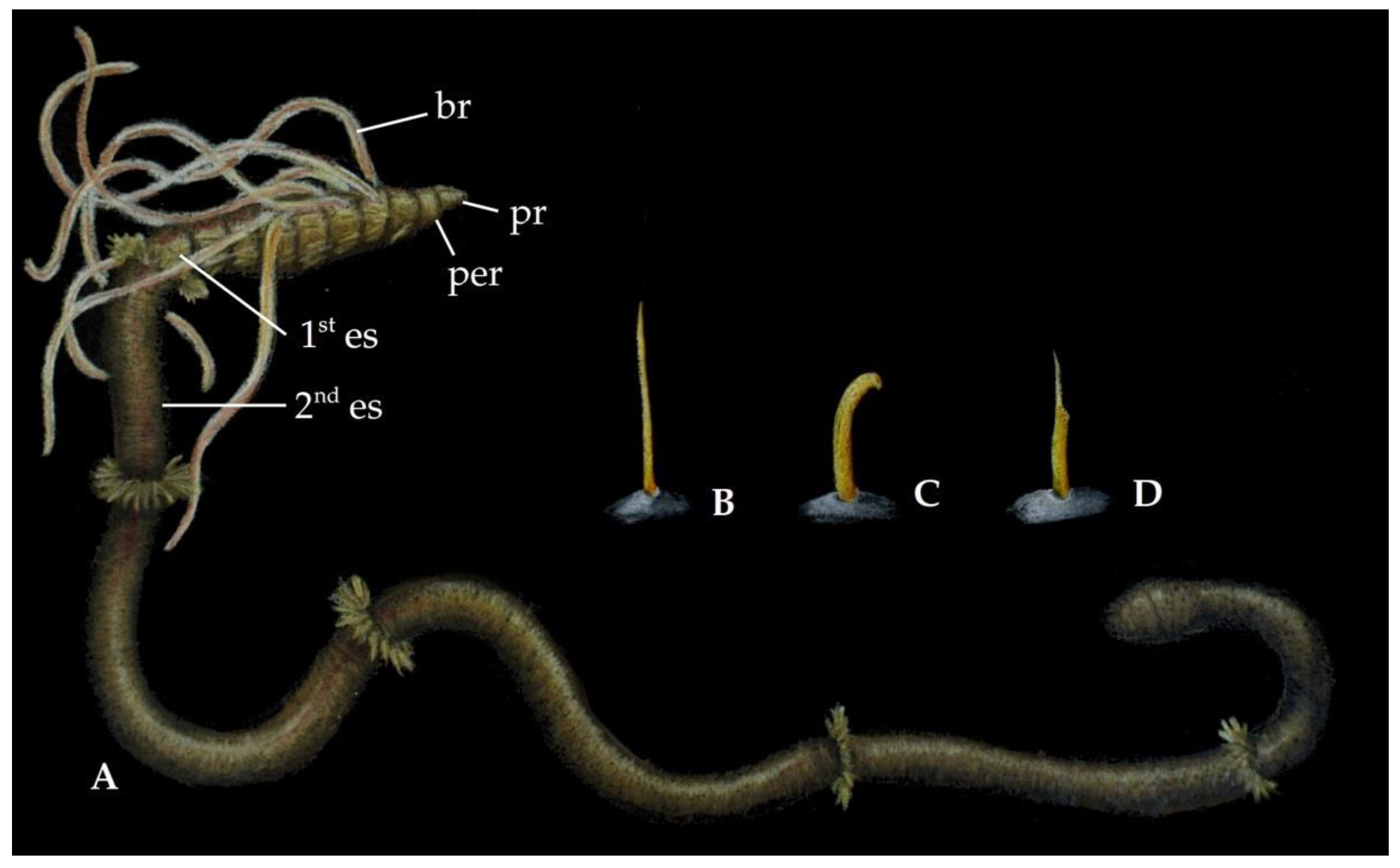
2.1. Morphology

2.2. Diversity and Phylogeny
2.3. Biogeography
2.4. Biology and Ecology
2.5. Conclusions
3. Cirratulidae (Ryckholt, 1851) Including Ctenodrilidae (Kennel, 1882)
3.1. Morphology
3.2. Diversity and Phylogeny
3.3. Biogeography
3.4. Biology and Ecology


3.5. Conclusions
4. Cossuridae Day, 1963
4.1. Morphology

4.2. Diversity and Phylogeny
4.3. Biogeography
4.4. Biology and Ecology
4.5. Conclusions
5. Longosomatidae (Hartman, 1944)
5.1. Morphology
5.2. Taxonomy and Phylogeny
5.3. Biogeography
5.4. Biology and Ecology
5.5. Conclusions
6. Paraonidae Cerruti, 1909
6.1. Morphology
6.2. Diversity and Phylogeny
6.3. Biogeography
6.4. Biology and Ecology
6.5. Conclusions
7. Sternaspidae (Carus, 1863)
7.1. Morphology

7.2. Taxonomy and Phylogeny
7.3. Biogeography

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Geographic Distribution | No. of Species | Species |
|---|---|---|
| Arctic Ocean | 2 | S. bushinskajae, S. fossor |
| Pacific | 29 | |
| NE Pacific | 5 | C. fauchaldi, C. nuda, S. affinis, S. uschakovi, S. williamsae |
| NW Pacific | 4 (+5) | S. annenkovae, S. bushinskajae, S. costata, S. uschakovi, (+ C. cf. nuda, S. cf. williamsae, and 3 unidentified species of Sternaspis; see [259] and [252] |
| CE Pacific | 9 | C. fauchaldi, C. villamari, P. deani, S. affinis, S. lindae, S. maior, S. maureri, S. uschakovi, S. williamsae |
| CW Pacific | 3 | P. palpallatoci, S. costata, S. piotrowskiae |
| W Pacific | ||
| E China Sea, Yellow Sea | 2 | S. chinensis, S. liui, (+? S. scutata in Jiaozhou Bay, see Wang et al. [268]) |
| South China Sea | 11 | C. laevis, P. salazari, S. andamanensis, S. britayevi, S. costata, S. nana, S. papillosa, S. radiata, S. spinosa, S. sunae, S. wui, |
| SW Pacific | 5 | C. laevis, S. annenkovae, S. maureri, S. princeps, S. spinosa |
| SE Pacific | 1 | S. chilensis |
| Atlantic | 9 | |
| NE Atlantic | 2 (+1) | C. gudmundssoni, S. islandica, (+ S. cf. scutata in English Channel; see [252]) |
| Mediterranean Sea | 2 | S. scutata, S. thalassemoides |
| NW Atlantic | 1 | S. fossor |
| CE Atlantic | 2 | M. longichaeta, S. africana |
| CW Atlantic | ||
| Gulf of Mexico | 1 | S. londognoi |
| SW Atlantic | 1 | P. capillata |
| SE Atlantic | 2 | M. longichaeta, S. africana |
| Indian Ocean | 8 | |
| Indian Ocean | 6 | C. laevis, P. palpallatoci, S. andamanensis, S. rietschi, S. spinosa, S. thorsoni, (+? S. scutata in Sundarban Mangroves, Gulf of Bengal, see [269], Cochin Estuary, SW India, see [270]; West coast of India, see [271]) |
| NW Indian Ocean | ||
| Arabian/Persian Gulf; Gulf of Oman | 1 | S. thorsoni |
| Red Sea | 1 | S. sherlockae, (+? S. scutata Gulf of Suez; see [272]) |
| SW Indian Ocean | 1 | P. harrisae |
| Antarctic Ocean | 1 | S. sendalli |
7.4. Biology and Ecology
8. Discussion: Methods and Perspectives
8.1. Washing Samples
8.2. Fixation
8.3. Species Description
8.3.1. Staining
8.3.2. Scanning Electron Microscopy
8.4. Molecular Genetic Investigations
8.5. Internal Morphology
9. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WoRMS—World Register of Marine Species. Available online: http://www.marinespecies.org/index.php (accessed on 10 March 2021).
- Struck, T.H.; Golombek, A.; Weigert, A.; Franke, F.A.; Westheide, W.; Purschke, G.; Bleidorn, C.; Halanych, K.M. The Evolution of Annelids Reveals Two Adaptive Routes to the Interstitial Realm. Curr. Biol. 2015, 25, 1993–1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weigert, A.; Helm, C.; Meyer, M.; Nickel, B.; Arendt, D.; Hausdorf, B.; Santos, S.R.; Halanych, K.M.; Purschke, G.; Bleidorn, C.; et al. Illuminating the Base of the Annelid Tree Using Transcriptomics. Mol. Biol. Evol. 2014, 31, 1391–1401. [Google Scholar] [CrossRef]
- Struck, T.H. Phylogeny. In Hanbook of Zoology, Annelida; Purschke, G., Böggemann, M., Westheide, W., Eds.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2019; Volume 1: Annelida Basal Groups and Pleistoannelida, Sedentaria I; pp. 37–68. [Google Scholar]
- Day, J.H. The polychaete fauna of South Africa. Part 8: New species and records from grab samples and dredgings. Bull. Br. Mus. (Nat. Hist.) Ser. Zool. 1963, 10, 381–445. [Google Scholar] [CrossRef]
- Cerruti, A. Contributo all’anatomia, biologia e sistematica delle Paraonidae (Levinsenidae) con particolare riguardo alle specie del golfo di Napoli. Mittheilungen aus der Zoologischen Station zu Neapel 1909, 19, 459–512. [Google Scholar]
- Caullery, M. Sur les Siboglinidae, type nouveau d’invertébrés receuillis par l’expédition du Siboga. Comptes Rendus Hebdomadaires des Séances de l’Académie des Sciences 1914, 158, 2014–2017. [Google Scholar]
- Pizl, V.; Chalupský, J. Hrabeiella periglandulata gen. et sp. n. (Annelida) a curious worm from Czechoslowakia. Vestn. Cesk. Spol. Zool. 1984, 48, 291–295. [Google Scholar]
- Ehrenberg, C.G. Animalia evertebrata exclusis Insectis. Series prima. In Symbolae Physicae, Seu Icones et Descriptiones Mammalium, Avium, Insectorum et Animalia Evertebra, Quae ex Itinere per Africam Borealem et Asiam Occidentalem Studio nova aut Illustrata Redierunt; Hemprich, F.G., Ehrenberg, C.G., Eds.; Officina Academica, ed.: Berlin, Germany, 1828; p. 126. [Google Scholar]
- Lastochkin, D.A. New name [Potamodrilus] for the genus Stephensoniella. Ann. Mag. Nat. Hist. 1935, 16, 488. [Google Scholar] [CrossRef]
- Malmgren, A.J. Annulata Polychaeta Spetsbergiæ, Grœnlandiæ, Islandiæ et Scandinaviæ. Öfversigt af Kongliga Vetenskaps-Akademien Förhandlingar 1867, 24, 127–235. [Google Scholar]
- Hartmann-Schröder, G. Annelida, Borstenwürmer, Polychaeta. In Die Tierwelt Deutschlands 58; Dahl, M., Peus, F., Eds.; Gustav Fischer: Jena, Germany, 1971; p. 594. [Google Scholar]
- Struck, T.H.; Hessling, R.; Purschke, G. The phylogenetic position of the Aelosomatidae and Parergodrilidae, two enigmatic oligochaete-like taxa of the ‘Polychaeta’, based on molecular data from 18S rDNA sequences. J. Zool. Syst. Evol. Res. 2002, 40, 155–163. [Google Scholar] [CrossRef]
- Langeneck, J.; Barbieri, M.; Maltagliati, F.; Castelli, A. Molecular phylogeny of Paraonidae (Annelida). Mol. Phylogenet. Evol. 2019, 136, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Martínez, A.; Di Domenico, M.; Worsaae, K. Evolution of cave Axiokebuita and Speleobregma (Scalibregma, Annelida). Zool. Scr. 2013, 42, 623–636. [Google Scholar]
- Li, Y.; Kocot, K.M.; Whelan, N.W.; Santos, S.R.; Waits, D.S.; Thornhill, D.J.; Halanych, K.M. Phylogenomics of tube worms (Siboglinidae, Annelida) and comparative performance of different reconstruction methods. Zool. Scr. 2017, 46, 200–213. [Google Scholar] [CrossRef]
- Parapar, J.; Martínez, A.; Moreira, J. On the systematics and biodivesity of the Opheliidae and Scalibregmatidae. Diversity 2021, 13, 87. [Google Scholar] [CrossRef]
- Hutchings, P.; Carrerette, O.; Nogueira, J.M.M.; Hourdez, S.; Lavesque, N. The Terebelliformia-Recent Developments and Future Directions. Diversity 2021, 13, 60. [Google Scholar] [CrossRef]
- Worsaae, K.; Kerbl, A.; Di Domenico, M.; Gonzalez, B.C.; Bekkouche, N.; Martínez, A. Interstitial Annelida. Diversity 2021, 13, 77. [Google Scholar] [CrossRef]
- Martinsson, S.; Erséus, C. Cryptic Clitellata: Molecular Species Delimitation of Clitellate Worms (Annelida): An Overview. Diversity 2021, 13, 36. [Google Scholar] [CrossRef]
- Meca, M.A.; Zhadan, A.E.; Struck, T.H. The Early Branching Group of Orbiniida Sensu Struck et al., 2015: Parergodrilidae and Orbiniidae. Diversity 2021, 13, 29. [Google Scholar] [CrossRef]
- Banse, K. Acrocirridae n.fam. (Polychaeta Sedentaria). J. Fish. Res. Board Can. 1969, 26, 2595–2620. [Google Scholar] [CrossRef]
- Ryckholt [Baron de], P. Mélanges paléontologiques, première partie. Mémoires Couronnés des Savants Étrangers de l’Académie Royale des Sciences, des Lettres et des Beaux Arts de Belgique 1851, 24, 1–176. [Google Scholar]
- Carus, J.V. Vermes. In Handbuch der Zoologie; Kükenthal, W., Ed.; Wilhelm Engelmann: Leipzig, Germany, 1863; Volume 2, pp. 422–454. [Google Scholar]
- Hartman, O. Polychaetous annelids. Part 6. Paraonidae, Magelonidae, Longosomidae, Ctenodrilidae, and Sabellariidae. Allan Hancock Pac. Exped. 1944, 10, 311–389. [Google Scholar]
- Saint Joseph [Baron de], A.A. Les annélides polychètes des côtes de Dinard. Troisième partie. Ann. des Sci. Nat. Zool. et Paléontol. 1894, 17, 1–395. [Google Scholar]
- Hartman, O. Abyssal polychaetous annelids from the Mozambique Basin off southeast Africa, with a compendium of abyssal polychaetous annelids from world-wide areas. J. Fish. Res. Board Can. 1971, 28, 1407–1428. [Google Scholar] [CrossRef]
- Martínez, A.; Worsaae, K.; Núñez, J. Acocirridae Banse, 1968. In Handbook of Zoology. Annelida; Westheide, W., Böggemann, M., Purschke, G., Eds.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2019; Volume 1: Annelida Basal Groups and Pleistoannelida, Sedentaria I; pp. 422–439. [Google Scholar]
- Blake, J.A.; Magalhães, W. Cirratulidae Ryckholt, 1851. In Handbook of Zoology. Annelida; Purschke, G., Westheide, W., Böggemann, M., Eds.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2019; Volume 1: Annelida Basal Groups and Pleistoannelida, Sedentaria I; pp. 339–397. [Google Scholar]
- Zhadan, A.E.; Salazar-Vallejo, S. Fauveliopsidae Hartman, 1971. In Hanbook of Zoology. Annelida; Purschke, G., Böggemann, M., Westheide, W., Eds.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2019; Volume 1: Annelida Basal Groups and Pleistoannelida, Sedentaria I; pp. 317–328. [Google Scholar]
- Fiege, D. Sternaspidae Carus, 1863. In Handbook of Zoology. Annelida; Purschke, G., Böggemann, M., Westheide, W., Eds.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2019; Volume 1: Annelida Basal Groups and Pleistoannelida, Sedentaria I; pp. 440–457. [Google Scholar]
- Osborn, K.J.; Madin, L.P.; Rouse, G.W. The remarkable squidworm is an example of discoveries that await in deep-pelagic habitats. Biol. Lett. 2011, 7, 449–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salazar-Vallejo, S.I.; Carrera-Parra, L.F.; Fauchald, K. Phylogenetic affinities of the Flabelligeridae (Annelida, Polychaeta). J. Zool. Syst. Evol. Res. 2008, 46, 203–215. [Google Scholar] [CrossRef]
- Ebbe, B.; Blake, J.A. Cossuridae Day, 1963. In Handbook of Zoology. Annelida; Purschke, G., Böggemann, M., Westheide, W., Eds.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2019; Volume 1: Annelida Basal Groups and Pleistoannelida, Sedentaria I; pp. 309–317. [Google Scholar]
- Blake, J.A. Paraonidae Cerruti 1909. In Handbook of Zoology. Annelida; Purschke, G., Böggemann, M., Westheide, W., Eds.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2019; Volume 1: Annelida Basal Groups and Pleistoannelida, Sedentaria I; pp. 281–308. [Google Scholar]
- Sousa, L.K.S.; Júnior, M.N.; Cutrim, M.V.J.; De Oliveira, V.M. Cossura yacy sp. Nov. (cossuridae, annelida) from a tropical brazilian estuary. Iheringia Ser. Zool. 2019, 109. [Google Scholar] [CrossRef] [Green Version]
- Rouse, G.W.; Fauchald, K. Cladistics and polychaetes. Zool. Scr. 1997, 26, 139–204. [Google Scholar] [CrossRef]
- Zrzavý, J.; Říha, P.; Piálek, L.; Janouškovec, J. Phylogeny of Annelida (Lophotrochozoa): Total-evidence analysis of morphology and six genes. BMC Evol. Biol. 2009, 9, 189. [Google Scholar] [CrossRef] [Green Version]
- Strelzov, V.E. Polychaete Worms of the Family Paraonidae Cerruti, 1909 (Polychaeta, Sedentaria); Akademia Nauk SSSR: Leningrad, Russia, 1973; pp. 1–169. [Google Scholar]
- Bleidorn, C. The role of character loss in phylogenetic reconstruction as exemplified for the Annelida. J. Zool. Syst. Evol. Res. 2007, 45, 299–307. [Google Scholar] [CrossRef]
- Rousset, V.; Pleijel, F.; Rouse, G.W.; Erséus, C.; Siddall, M.E. A molecular phylogeny of annelids. Cladistics 2007, 23, 41–63. [Google Scholar] [CrossRef]
- Bleidorn, C. Phylogenetic relationships and evolution of Orbiniidae (Annelida, Polychaeta) based on molecular data. Zool. J. Linn. Soc. 2005, 144, 59–73. [Google Scholar] [CrossRef] [Green Version]
- Blake, J.; Maciolek, N.J. Longosomatidae Hartman, 1944. In Handbook of Zoology. Annelida; Purschke, G., Böggemann, M., Westheide, W., Eds.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2019; Volume 1: Annelida Basal Groups and Pleistoannelida, Sedentaria I; pp. 457–466. [Google Scholar]
- Ehlers, E. Annulata nova vel minus cognita in Expeditione ‘Porcupine’ capta. Ann. Mag. Nat. Hist. 1874, 13, 292–298. [Google Scholar] [CrossRef]
- Vijapure, T.; Sukumaran, S.; Manohar, C.S. Molecular characterization and phylogenetics of Indian polychaete fauna: Scope for implementation in ecological monitoring. Aquat. Ecol. 2019, 53, 665–677. [Google Scholar] [CrossRef]
- Salazar-Vallejo, S.I.; Zhadan, A.E.; Rizzo, A.E. Revision of Fauveliopsidae Hartman, 1971 (Annelida, Sedentaria). Zootaxa 2019, 4637, 1–67. [Google Scholar] [CrossRef]
- Grube, A.E. Die Familie der Cirratuliden. Jahres Bericht der Schlesischen Gesellschaft für Vaterländische Cultur Breslau 1873, 50, 59–66. [Google Scholar]
- Grube, A.E. Die Familien der Anneliden. Arch. für Nat. Berl. 1850, 16, 249–364. [Google Scholar]
- Gillet, P. Flabelligena amoureuxi new genus, new species (Polychaeta: Acrocirridae) from Crozet Islands (Indian Ocean). Bull. Mar. Sci. 2001, 68, 125–131. [Google Scholar]
- Jimi, N.; Fujimoto, S.; Imura, S. A new interstitial genus and species of Acrocirridae from Okinawa-jima Island, Japan. Zoosymposia 2020, 19, 164–172. [Google Scholar] [CrossRef]
- Hartman, O. Polychaeta from the Weddell Sea quadrant, Antarctica. Antarct. Res. Ser. 1978, 26, 125–223. [Google Scholar]
- Laubier, L. Chauvinelia biscayensis gen. sp. nov., un Flabelligeridae (annélide polychète sédentaire) aberrant de l’étage abyssal du Golfe de Gascogne. Bull. Soc. Zool. Fr. 1974, 99, 391–399. [Google Scholar]
- Osborn, K.J.; Haddock, S.H.D.; Pleijel, F.; Madin, L.P.; Rouse, G.W. Deep-Sea, Swimming Worms with Luminescent “Bombs”. Science 2009, 325, 964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salazar-Vallejo, S.; Gillet, P.; Carrera-Parra, L.F. Revision of Chauviniella, redescriptions of Flabelliseta incrusta, and Helmetophorus rankini, and their recognition as acrocirrids (Polychaeta: Acrocirridae). J. Mar. Biol. Assoc. U. K. 2007, 87, 465–477. [Google Scholar] [CrossRef]
- Kudenov, J.D. Polychaeta from southeaster Australia 1. Acrocirridae Banse, 1969, from Victoria and New South Wales. Rec. Aust. Mus. 1976, 30, 137–151. [Google Scholar] [CrossRef]
- Martínez, A.; Di Domenico, M.; Leasi, F.; Curini-Galletti, M.; Todaro, M.A.; Zotto, M.D.; Gobert, S.; Artois, T.; Norenburg, J.; Jörger, K.M.; et al. Patterns of diversity and endemism of soft-bodied meiofauna in an oceanic island, Lanzarote, Canary Islands. Mar. Biodivers. 2019, 49, 2033–2055. [Google Scholar] [CrossRef] [Green Version]
- Osborn, K.J.; Rouse, G.W. Phylogenetics of Acrocirridae and Flabelligeridae (Cirratuliformia, Annelida). Zool. Scr. 2011, 40, 204–219. [Google Scholar] [CrossRef]
- Hartman, O. Deep-Water Benthic Polychaetous Annelids off New England to Bermuda and Other North Atlantic Areas. Occas. Pap. Allan Hancock Found. 1965, 28, 1–378. [Google Scholar]
- Gonzalez, B.C.; Borda, E.; Carbalho, R.; Schulze, A. Polychaetes from the Mayan underworld: Phylogeny, evolution, and cryptic diversity. Nat. Croat. Period. Musei Hist. Nat. Croat. 2012, 21, 51–53. [Google Scholar]
- Nunez, J.; Ocana, O.; Brito, M.C. Two New Species (Polychaeta: Fauveliopsidae and Nerillidae) and other Polychaetes from the Marine Lagoon Cave of Jameos Del Agua, Lanzarote (Canary Islands). Bull. Mar. Sci. 1997, 60, 252–260. [Google Scholar]
- Riera, R.; Monterroso, Ó.; Núñez, J.; Martínez, A. Distribution of meiofaunal abundances in a marine cave complex with secondary openings and freshwater filtrations. Mar. Biodivers. 2018, 48, 203–215. [Google Scholar] [CrossRef]
- Hartman, O. Polychaetous annelids of the Indian Ocean including an account of species collected by members of the International Indian Ocean Expeditions, 1963-1964 and a catalogue and bibliography of the species from India. J. Mar. Biol. Assoc. India 1976, 16, 191–252. [Google Scholar]
- Santos, P.J.P.; Silva, V.M.A.P. Macrochaeta westheidi n. sp.: First record of Acrocirridae for the Brazilian coast (annelida Polychaeta). Nerítica Curitiba 1993, 7, 7–12. [Google Scholar]
- Magalhães, W.; Bailey-Brock, J.H. A new species of Acrocirrus (Polychaeta: Acrocirridae) from Coconut Island, Oahu, Hawaii. J. Mar. Biol. Assoc. U. K. 2012, 92, 1019–1102. [Google Scholar] [CrossRef]
- Friedrich, H. Polychaetenstudien, I-III. Kiel. Meeresforsch. 1937, 1, 343–351. [Google Scholar]
- Fauvel, P. Polychètes Sédentaires. Addenda aux Errantes, Archiannélides, Myzostomaires; Paul Chevalier: Paris, France, 1927; Volume 16, 494p. [Google Scholar]
- Westheide, W. Interstitielle Fauna von Galapagos. XXVI. Questidae, Cirratulidae, Acrocirridae, Ctenodrillidae (Polychaeta). Mikrofauna des Meeresbod. 1981, 82, 1–23. [Google Scholar]
- Parapar, J.; Moreira, J. Polychaeta of the ‘DIVA-Artabria I’ project (cruise 2002) in the continental shelf and upper slope off Galicia (NW Spain). Cah. de Biol. Mar. 2009, 50, 57–78. [Google Scholar]
- Martínez, A.; Gonzalez, B.C. Volcaline anchialine habitats of Lanzarote. In Cave Ecology; Moldovan, O.T., Kováč, L., Halse, S., Eds.; Ecological Studies; Springer: Cham, Switzerland, 2018; Volume 235, pp. 399–414. [Google Scholar]
- Martínez, A.; Kvindebjerg, K.; Lliffe, T.; Worsaae, K. Evolution of cave suspension feeding in Protodrilidae (Annelida). Zool. Scr. 2017, 46, 214–226. [Google Scholar] [CrossRef]
- Worsaae, K.; Gonzalez, B.C.; Kerbl, A.; Holdfold Nielsen, S.; Terp Jørgensen, J.; Armenteros, M.; Iliffe, T.M.; Martínez, A. Diversity and evolution of the stygobitic Speleonerilla nom. nov. (Nerillidae, Annelida) with description of three new species from anchialine caves in the Caribbean and Lanzarote. Mar. Biodivers. 2019, 49, 2167–2192. [Google Scholar] [CrossRef]
- Gonzalez, B.C.; Worsaae, K.; Fontaneto, D.; Martínez, A. Anophtalmia and elongation of body appendages in cave scale worms (Annelida: Aphroditiformia). Zool. Scr. 2018, 47, 106–121. [Google Scholar] [CrossRef]
- Jumars, P.A.; Dorgan, K.M.; Lindsay, S.M. Diet of Worms Emended: An Update of Polychaete Feeding Guilds. Annu. Rev. Mar. Sci. 2015, 7, 497–520. [Google Scholar] [CrossRef] [Green Version]
- Sars, M. Beskrivelser og Iagttagelser over nogle Moerkelige eller nye i Havet ved den Bergenske Kyst Levende Dyr af Polypernes, Acalephernes, Radiaternes, Annelidernes og Molluskernes Classer, med en kort Oversigt over de Hidtil af Forfatteren Sammesteds Fundne Arter og deres Forekommen; Thorstein Hallagers: Bergen, Norway, 1835; 81p. [Google Scholar]
- Lamarck, J.B. Histoire Naturelle des Animaux sans Vertèbres, Préséntant les Caractères Généraux et Particuliers de ces Animaux, leur Distribution, leurs Classes, leurs Familles, leurs Genres, et la Citation des Principales Espèces qui s’y Rapportent; Précedés d’une Introduction Offrant la Determination des Caracteres Essentiels de l’Animal, sa Distinction du Vegetal et des Autres Corps Naturels, Enfin, l’Exposition des Principes Fondamentaux de la Zoologie; Deterville: Paris, France, 1818. [Google Scholar]
- Hartman, O. Nomenclatural changes involving California polychaete worms. J. Wash. Acad. Sci. 1936, 26, 31–32. [Google Scholar]
- Kinberg, J.G.H. Annulata Nova. Öfversigt af Königlich Vetenskapsakademiens Förhandlingar Stockholm 1866, 22, 239–258. [Google Scholar]
- Çinar, M.E.; Petersen, M.E. Re-description of Cirratulus dollfusi (Polychaeta: Cirratulidae), and Fauvelicirratulus as a new genus. J. Mar. Biol. Assoc. U. K. 2011, 91, 415–418. [Google Scholar] [CrossRef]
- Czerniavski, V. Materialia ad zoographiam Ponticam comparatam. Bull. de la Société Impériale des Naturalistes de Moscou 1881, 56, 338–420. [Google Scholar]
- Blake, J. Revision of some genera and species of Cirratulidae from the Western North Atlantic. Ophelia Suppl. 1991, 5, 17–30. [Google Scholar]
- Chamberlin, R.V. The Annelida Polychaeta [Albatross Expeditions]. Mem. Mus. Comp. Zool. Harv. Coll. 1919, 48, 1–514. [Google Scholar]
- Blake, J.A. Bitentaculate Cirratulidae (Annelida, Polychaeta) collected chiefly during cruises of the R/V Anton Bruun, USNS Eltanin, USCG Glacier, R/V Hero, RVIB Nathaniel B. Palmer, and R/V Polarstern from the Southern Ocean, Antarctica, and off Western South America. Zootaxa 2018, 4537, 1–130. [Google Scholar] [CrossRef] [Green Version]
- Blake, J.A. Kirkegaardia (Polychaeta, Cirratulidae), new name for Monticellina Laubier, preoccupied in the Rhabdocoela, together with new records and descriptions of eight previously known and sixteen new species from the Atlantic, Pacific, and Southern Oceans. Zootaxa 2016, 4166, 1–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webster, H.E.; Benedict, J.E. The Annelida Chaetopoda from Eastport, Maine; Report of the United States Commissioner of Fisheries; U.S. Commission of Fish & Fisheries: Washington, DC, USA, 1887; pp. 707–758. [Google Scholar]
- Ørsted, A.S. Annulatorum Danicorum Conspectus. Auctore A.S. Örsted. Fasc. I. Maricolæ. (Quæstio ab Universitate Hafniensi ad Solvendum Proposita et Proemio Ornata); Sumtibus Librariæ Wahlianæ: Copenhagen, Denmark, 1843. [Google Scholar]
- Claparède, A.R.E. Beobachtungen über Anatomie und Entwicklungsgeschichte Wirbelloser Thiere: An der Küste von Normandie Angestellt; W. Engelmann: Leipzig, Germany, 1863; p. 120, 118 plates. [Google Scholar]
- Wilfert, M. Aphropharynx heterochaeta nov. gen. nov. spec., ein neuer polychaet aus der familie Ctenodrilidae Kennel 1882. Cah. de Biol. Mar. 1974, 15, 495–504. [Google Scholar]
- Monticelli, F.S. Di un nuovo Ctenodrilide del Golfo di Napoli. Nota preliminare riassuntiva del socio ordinario. Rend. dell’Accad. delle Sci. Fische e Mat. 1910, 16, 61–64. [Google Scholar]
- Hartman, O. Polychaetous annelids from California. Allan Hancock Pac. Exped. 1961, 25, 1–226. [Google Scholar]
- Kennel, J. Über Ctenodrilus pardalis Clpd. Ein Beitrag zur Kenntniss der Anatomie und Knospung der Anneliden. Arb. aus dem Zool. Zootomischen Inst. Würzbg. 1882, 5, 373–429. [Google Scholar]
- Magalhães, W.F.; Linse, K.; Wiklund, H. A new species of Raricirrus (Annelida: Cirratuliformia) from deep-water sunken wood off California. Zootaxa 2017, 4353, 51–68. [Google Scholar] [CrossRef] [PubMed]
- Weidhase, M.; Bleidorn, C.; Simon, C.A. On the taxonomy and phylogeny of Ctenodrilus (Annelida: Cirratulidae) with a first report from South Africa. Mar. Biodivers. 2016, 46, 243–252. [Google Scholar] [CrossRef]
- Grosse, M.; Bakken, T.; Nygren, A.; Kongsrud, J.A.; Capa, M. Species delimitation analyses of NE Atlantic Chaetozone (Annelida, Cirratulidae) reveals hidden diversity among a common and abundant marine annelid. Mol. Phylogenet. Evol. 2020, 149, 106852. [Google Scholar] [CrossRef]
- Elias, R.; Rivero, M.S.; Orensanz, J.M.L. New species of Monticellina and Chaetozone (Polychaeta: Cirratulidae) in the SW Atlantic, and a review of Monticellina species. J. Mar. Biol. Assoc. U. K. 2017, 97, 1553–1563. [Google Scholar] [CrossRef]
- Blake, J. Family Cirratulidae Ryckholdt, 1851. Including a revision of the Genera and Species from the Eastern North Pacific. In Taxonomic Atlas of the Benthic Fauna of the Santa Maria Bassin and Western Santa Barbara Channel; Blake, J., Hilbig, B., Scott, P.H., Eds.; Santa Barbara Museum of Natural History: Santa Barbara, CA, USA, 1996; Volume 6—The Annelida Part 3. Polychaeta: Orbiniidae to Cossuridae; pp. 27–70. [Google Scholar]
- Magalhães, W.F.; Bailey-Brock, J.H.; Davenport, J.F. On the genus Raphidrilus Monticelli, 1910 (Polychaeta: Ctenodrilidae) with description of two new species. Zootaxa 2011, 2804, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Elías, R.; Rivero, M.S. Two new species of Cirratulidae (Annelida: Polychaeta) from Mar del Plata, Argentina (SW Atlantic). Zoosymposia 2009, 2, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Dean, H.K.; Blake, J. Monticellina (Polychaeta: Cirratulidae) from the Pacific coast of Costa ica with descriptions of six new species. Zoosymposia 2009, 2, 105–126. [Google Scholar] [CrossRef] [Green Version]
- Blake, J.A.; Lavesque, N. A new species of Chaetozone (Polychaeta, Cirratulidae) from the Bay of Biscay offshore France, together with a review of Chaetozone species from the North Atlantic and adjacent waters. Zootaxa 2017, 4312, 565. [Google Scholar] [CrossRef] [Green Version]
- Doner, S.A.; Blake, J.A. New species of Cirratulidae (Polychaeta) from the northeastern United States. Sci. Mar. 2006, 70, 65–73. [Google Scholar] [CrossRef]
- Dean, H.K.; Blake, J.A. Ctenodrilidae Kennel, 1882. In Handbook of Zoology. Annelida; Westheide, W., Purschke, G., Böggemann, M., Eds.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2019; Volume 1: Annelida Basal Groups and Pleistoannelida, Sedentaria I; pp. 328–338. [Google Scholar]
- Blake, J.A. New species of Cirratulidae (Annelida, Polychaeta) from abyssal depths of the Clarion-Clipperton Fracture Zone, North Equatorial Pacific Ocean. Zootaxa 2019, 4629, 151–187. [Google Scholar] [CrossRef] [Green Version]
- Magalhães, W.F.; Bailey-Brock, J.H. Bitentaculate Cirratulidae (Annelida: Polychaeta) from the northwestern Pacific Islands with description of nine new species. Zootaxa 2013, 3630, 80–116. [Google Scholar] [CrossRef] [Green Version]
- Magalhães, W.F.; Seixas, V.C.; Paiva, P.C.; Elias, R. The Multitentaculate Cirratulidae of the Genera Cirriformia and Timarete (Annelida: Polychaeta) from Shallow Waters of Brazil. PLoS ONE 2014, 9, e112727. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.K.; Kim, H.; Yoon, S.M. Timarete posteria, a new cirratulid species from Korea (Annelida, Polychaeta, Cirratulidae). ZooKeys 2018, 806, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blake, J.A.; Dean, H.K. New Species of Cirratulidae (Annelida, Polychaeta) from the Caribbean Sea. Zootaxa 2019, 4671, 301–338. [Google Scholar] [CrossRef]
- Elías, R.; Saracho-Bottero, M.A.; Simon, C.A. Protocirrineris (Polychaeta:Cirratulidae) in South Africa and description of two new species. Rev. de Biol. Trop. 2019, 67, S70–S80. [Google Scholar] [CrossRef]
- Elías, R.; Saracho-Bottero, M.A.; Magalhães, W.F. Two new species of Protocirrineris (Polychaeta: Cirratulidae) from Brazil. Rev. de Biol. Trop. 2019, 67, S81–S91. [Google Scholar] [CrossRef]
- Schmidt, O. Zur Kenntnis der Turbellaria, Rhabdocoela und einiger anderer Wuermer des Mittelmeeres. Sitzungsberichte der Kaiserliche Akademie der Wissenschaften Wien Mathematisch Naturwissenschaftliche Klasse 1857, 23, 347–366. [Google Scholar]
- Grube, A.E. Annulata Örstediana. Enumeratio Annulatorum, quae in itinere Indiam Occidentalem et Americam Centralem annis 1845-1848 suscepto legit cl. A.S. Oersted, adjectis speciebus nonnullis a cl. H. Kroyero in itinere ad Americam meridionalem collectis. Videnskabelige Meddelelser fra Dansk naturhistorisk Forening i Köbenhavn 1858, 3, 105–120. [Google Scholar]
- Blake, J.A. New species of Chaetozone and Tharyx (Polychaeta: Cirratulidae) from the Alaskan and Canadian Arctic and the Northeastern Pacific, including a description of the lectotype of Chaetozone setosa Malmgren from Spitsbergen in the Norwegian Arctic. Zootaxa 2015, 3919, 501. [Google Scholar] [CrossRef]
- Seixas, V.C.; Zanol, J.; Magalhães, W.F.; Paiva, P.C. Genetic diversity of Timarete punctata (Annelida: Cirratulidae): Detection of pseudo-cryptic species and a potential biological invader. Estuar. Coast. Shelf Sci. 2017, 197, 214–220. [Google Scholar] [CrossRef]
- Westheide, W.; Haß-Cordes, E.; Krabusch, M. Ctenodrilus serratus (Polychaeta: Ctenodrilidae) is a truly amphi-Atlantic meiofauna species—Evidence from molecular data. Mar. Biol. 2003, 142, 637–642. [Google Scholar] [CrossRef]
- Berkeley, E.; Berkeley, C. On a collection of Polychaeta from Southern California. Bull. South. Calif. Acad. Sci. 1941, 40, 16–60. [Google Scholar]
- Omena, E.; Creed, J.C. Polychaete fauna of seagrass beds (Halodule wrightii Ascherson) along the coast of Rio de Janeiro (southeast Brazil). P. S. Z. N. Marine Ecology 2004, 25, 273–288. [Google Scholar] [CrossRef]
- Çinar, M.E.; Ergen, Z. The presence of Chaetozone corona (Polychaeta: Cirratulidae) in the Mediterranean Sea: An alien or a native species? Cah. de Biol. Mar. 2007, 48, 339–346. [Google Scholar]
- Le Garrec, V.; Grall, J.; Chevalier, C.; Guyonnet, B.; Jourde, J.; Lavesque, N.; Bonifácio, P.; Blake, J.A. Chaetozone corona (Polychaeta, Cirratulidae) in the Bay of Biscay: A new alien species for the North-east Atlantic waters? J. Mar. Biol. Assoc. U. K. 2017, 97, 433–445. [Google Scholar] [CrossRef]
- Chambers, S.J. A redescription of Chaetozone setosa Malmgren, 1867 including a definition of the genus, and a description of a new species of Chaetozone (Polychaeta: Cirratulidae) from the Northeast Atlantic. Bull. Mar. Sci. 2000, 67, 10. [Google Scholar]
- Langerhans, P. Die Wurmfauna von Madeira. III. Z. für Wiss. Zool. 1880, 34, 87–143. [Google Scholar]
- Montagu, G. Description of several marine animals found on the south coast of Devonshire. Trans. Linn. Soc. Lond. 1808, 9, 81–114. [Google Scholar] [CrossRef] [Green Version]
- Berkeley, E.; Berkeley, C. Notes on the life-history of the polychaete Dodecaceria fewkesii (nom.n.). J. Fish. Res. Board Can. 1954, 11, 326–334. [Google Scholar] [CrossRef]
- Dean, H.K. A new species of Raricirrus (Polychaeta: Ctenodrilidae) from wood collected in the Tongue of the Ocean, Virgin Islands. Proc. Biol. Soc. Wash. 1995, 108, 169–179. [Google Scholar]
- Hily, C. Spatio-temporal variability of Chaetozone setosa (Malmgren) populations on an organic gradient in the Bay of Brest, France. J. Exp. Marine Biol. Ecol. 1987, 112, 201–216. [Google Scholar] [CrossRef]
- Petersen, M.E.; George, J.D. A new species of Raricirrus from Northern Europe, with notes on its biology and a discussion of the affinities of the genus (polychaeta: Ctenodrilidae). Ophelia Suppl. 1991, 5, 185–208. [Google Scholar]
- Pearson, T.H.; Rosenberg, R. Macrobenthic succession in relation to organic enrichment and pollution of the marine environment. Oceanogr. Mar. Biol. Annu. Rev. 1978, 16, 229–311. [Google Scholar]
- Elías, R.; Rivero, M.S.; Palacios, J.R.; Vallarino, E.A. Sewage-induced disturbance on polychaetes inhabiting intertidal mussel beds of Brachidontes rodriguezii off Mar del Plata (SW Atlantic, Argentina). Sci. Mar. 2006, 70, 187–196. [Google Scholar] [CrossRef] [Green Version]
- Elías, R.; Rivero, M.S.; Vallarino, E.A. Sewage impact on the composition and distribution of Polychaeta associated to intertidal mussel beds of the Mar del Plata rocky shore, Argentina. Iheringia Ser. Zool. 2003, 93, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, P.; Langston, W.; Burt, G.; Pascoe, P. Tharyx marioni (Polychaeta): A remarkable accumulator of arsenic. J. Mar. Biol. Assoc. U. K. 1983, 63, 313–325. [Google Scholar] [CrossRef]
- Milanovich, F.P.; Spies, R.; Guram, M.S.; Sykes, E.E. Uptake of copper by the polychaete Cirriformia spirabrancha in the presence of dissolved yellow organic matter of natural origin. Estuar. Coast. Mar. Sci. 1976, 4, 585–588. [Google Scholar] [CrossRef]
- Moore, D.C. Raricirrus beryli Petersen and George (Ctenodrilidae): A new polychaete indicator species for hydrocarbon-polluted sediments. Ophelia Suppl. 1991, 5, 477–486. [Google Scholar]
- Reish, D.J.; Gerlinger, T.V. A Review of the Toxicological Studies with Polychaetous Annelids. Bull. Mar. Sci. 1997, 60, 584–607. [Google Scholar]
- Petersen, M.E. Reproduction and development in Cirratulidae (Annelida: Polychaeta). In Reproductive Strategies and Developmental Patterns in Annelids; Dorresteijn, A.W.C., Westheide, W., Eds.; Springer: Dordrecht, The Netherlands, 1999; pp. 107–128. [Google Scholar]
- Christie, G. A new species of Tharyx (Polychaeta: Cirratulidae) from the estuaries in north-east England. Sarsia 1984, 69, 69–73. [Google Scholar]
- Hilbig, B. Family Cossuridae Day, 1963. In Taxonomic Atlas of the Benthic Fauna of the Santa Maria Basin and Western Santa Barbara Channel; Blake, J.A., Hilbig, B., Scott, P.H., Eds.; Santa Barbara Museum of Natural History: Santa Barbara, CA, USA, 1996; Volume 6—The Annelida Part 3. Polychaeta: Orbiniidae to Cossuridae; pp. 385–402. [Google Scholar]
- Liñero-Arana, I.; Díaz-Díaz, Ó. A new species of Cossuridae (Annelida: Polychaeta) from Venezuela. Interciencia 2010, 35, 789–792. [Google Scholar]
- Zhadan, A. Cossuridae (Annelida: Polychaeta: Sedentaria) from australian and adjacent waters: The first faunistic survey. Rec. Aust. Mus. 2015, 67, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Fournier, J.A.; Petersen, M.E. Cossura longocirrata: Redescription and distribution, with notes on reproductive biology and a comparison of described species of Cossura (Polychaeta: Cossuridae). Ophelia Suppl. 1991, 5, 63–80. [Google Scholar]
- Read, G.B. Taxonomy and distribution of a new Cossura species (Annelida: Polychaeta: Cossuridae) from New Zealand. Proc. Biol. Soc. Wash. 2000, 113, 1096–1110. [Google Scholar]
- Zhadan, A.E.; Vortsepneva, E.V.; Tsetlin, A.B. Redescription and biology of Cossura pygodactylata Jones, 1956 (Polychaeta: Cossuridae) in the White Sea. Invertebr. Zool. 2012, 9, 115–125. [Google Scholar] [CrossRef]
- Reish, D.J. Description of a new species of Cossura (Annelida: Polychaeta) from the Mississippi Delta. J. Wash. Acad. Sci. 1958, 48, 53–55. [Google Scholar]
- Bachelet, G.; Laubier, L. Morphology, ecology and juvenile development of Cossura pygodactylata Jones (Polychaeta, Cossuridae) in Arcachon Bay, SW France, with a reassessment of the geographical distribution of C. pygodactylata and C. soyeri Laubier. Mem. Mus. Natl. D’histoire Nat. 1994, 162, 355–369. [Google Scholar]
- Ewing, R.M. Review of the genus Cossurella (Polychaeta: Cossuridae) including descriptions of two new species and a key to the species of the world. Bull. Biol. Soc. Wash. 1987, 7, 3–10. [Google Scholar]
- Kitamori, R. Two new species of cirratulid and Nephthydidae (Annelida: Polychaeta. Bull. Jpn. Soc. Sci. Fish. 1960, 26, 1082–1085. [Google Scholar] [CrossRef]
- Górska, B.; Gromisz, S.; Włodarska-Kowalczuk, M. Size assessment in polychaete worms—application of morphometric correlations for common North Atlantic taxa. Limnol. Oceanogr. Methods 2019, 17, 254–265. [Google Scholar] [CrossRef]
- Zhadan, A. Two new species of Cossura (Cossuridae, Annelida) from the terminal lobes of the Congo River deep-sea fan. J. Mar. Biol. Assoc. U. K. 2017, 97, 881–888. [Google Scholar] [CrossRef]
- Laubier, L. Découverte du genre Cossura (polychète, Cossuridae) en Méditerranée: Cossura soyeri sp. n. Vie et Milieu 1963, 14, 833–842. [Google Scholar]
- Callier, M.D.; Fletcher, R.L.; Thorp, C.H.; Fichet, D. Macrofaunal community responses to marina-related pollution on the south coast of England and west coast of France. J. Mar. Biol. Assoc. U. K. 2009, 89, 19–29. [Google Scholar] [CrossRef]
- Bogdanos, C.; Fredj, G. Sur la présence de Cossura coasta Kitamori, 1960 (Polychaeta, Cossuridae) dans les eaux cotieres Grecques (Mer Égée, Mer Ionienne). Thalassographica 1983, 6, 5–15. [Google Scholar]
- Zenetos, A.; Gofas, S.; Verlaque, M.; Cinar, M.E.; Garcia Raso, J.E.; Bianchi, C.N.; Morri, C.; Azzurro, E.; Bilecenoglu, M.; Froglia, C.; et al. Alien species in the Mediterranean Sea by 2010. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part I. Spatial distribution. Mediterr. Mar. Sci. 2010, 11, 381. [Google Scholar] [CrossRef] [Green Version]
- Faulwetter, S.; Simboura, N.; Katsiaras, N.; Chatzigeorgiou, G.; Arvanitidis, C. Polychaetes of Greece: An updated and annotated checklist. Biodivers. Data J. 2017, 22, e20997. [Google Scholar] [CrossRef] [Green Version]
- Raghunathan, C.; Gopi, K.C.; Chandra, K. Invasive alien faunal species in India. J. Gov. 2019, 18, 124–140. [Google Scholar]
- Gaonkar, C.A.; Sawant, S.S.; Anil, A.C.; Venkat, K.; Harkantra, S.N. Mumbai harbour, India: Gateway for introduction of marine organisms. Environ. Monit. Assess. 2010, 163, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, D.; Gallardo, V.A.; Mayor, S.; Neira, C.; Vásquez, C.; Sellanes, J.; Rivas, M.; Soto, A.; Carrasco, F.; Baltazar, M. Effects of dissolved oxygen and fresh organic matter on the bioturbation potential of macrofauna in sublittoral sediments off Central Chile during the 1997/1998 El Niño. Mar. Ecol. Prog. Ser. 2000, 202, 81–99. [Google Scholar] [CrossRef] [Green Version]
- Ryu, J.; Khim, J.S.; Kang, S.-G.; Kang, D.; Lee, C.-h.; Koh, C.-h. The impact of heavy metal pollution gradients in sediments on benthic macrofauna at population and community levels. Environ. Pollut. 2011, 159, 2622–2629. [Google Scholar] [CrossRef] [PubMed]
- Moreira, J.; Lourido, A.; Troncoso, J.S. Temporal dynamics of the benthic assemblage in the muddy sediments of the harbour of Baiona (Galicia, NW Iberian Peninsula). Thalassas 2010, 26, 9–22. [Google Scholar]
- Balachandar, K.; Sundaramanickam, A.; Kumaresan, S. Spatial and seasonal variation of macrobenthos from Puducherry Coast, Southeast Coast of India. Int. J. Curr. Microbiol. Appl. Sci. 2016, 5, 33–49. [Google Scholar] [CrossRef] [Green Version]
- Chainho, P.; Chaves, M.L.; Costa, J.L.; Costa, M.J.; Dauer, D.M. Use of multimetric indices to classify estuaries with different hydromorphological characteristics and different levels of human pressure. Mar. Pollut. Bull. 2008, 56, 1128–1137. [Google Scholar] [CrossRef]
- Khan, S.A.; Murugesan, P. Polychaete diversity in Indian estuaries. Indian J. Geo-Mar. Sci. 2005, 34, 114–119. [Google Scholar] [CrossRef]
- Murugesan, P.; Ajmal Khan, S.; Ajithkumar, T.T. Temporal changes in the benthic community structure of the marine zone of Vellar estuary southeast coast of India. J. Mar. Biol. Assoc. India 2007, 49, 154–158. [Google Scholar]
- Rakocinski, C.F.; Brown, S.S.; Gaston, G.R.; Heard, R.W.; Walker, W.W.; Summers, J.K. Macrobenthic responses to natural and contaminant-related gradients in northern Gulf of Mexico estuaries. Ecol. Appl. 1997, 7, 1278–1298. [Google Scholar] [CrossRef]
- Méndez, N. Annelid assemblages in soft bottoms subjected to human impact in the Urías estuary (Sinaloa, Mexico). Oceanol. Acta 2002, 25, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Lohrer, A.M.; Chiaroni, L.D.; Hewitt, J.E.; Thrush, S.F. Biogenic Disturbance Determines Invasion Success In A Subtidal Soft-Sediment System. Ecology 2008, 89, 1299–1307. [Google Scholar] [CrossRef]
- Widdicombe, S.; Austen, M.C. Mesocosm investigation into the effects of bioturbation on the diversity and structure of a subtidal macrobenthic community. Mar. Ecol. Prog. Ser. 1999, 189, 181–193. [Google Scholar] [CrossRef]
- Rygg, B. Effect of sediment copper on benthic fauna. Mar. Ecol. Prog. Ser. 1985, 25, 83–89. [Google Scholar] [CrossRef]
- Olsgard, F.; Hasle, J.R. Impact of waste from titanium mining on benthic fauna. J. Exp. Mar. Biol. Ecol. 1993, 172, 185–213. [Google Scholar] [CrossRef]
- Burd, B.J. Evaluation of mine tailings effects on a benthic marine infaunal community over 29 years. Mar. Environ. Res. 2002, 53, 481–519. [Google Scholar] [CrossRef]
- Ellis, D.V. The concept of “sustainable ecological succession”; and its value in assessing the recovery of sediment seabed biodiversity from environmental impact. Mar. Pollut. Bull. 2003, 46, 39–41. [Google Scholar] [CrossRef]
- Ellis, D.V. Biodiversity reduction on contaminated sediments in the sea: Contamination or sediment mobility? Mar. Pollut. Bull. 2005, 50, 244–246. [Google Scholar] [CrossRef]
- Lancellotti, D.A.; Stotz, W.B. Effects of shoreline discharge of iron mine tailings on a marine soft-bottom community in northern Chile. Mar. Pollut. Bull. 2004, 48, 303–312. [Google Scholar] [CrossRef]
- Raman, A.V.; Ganapati, P.N. Pollution effects on ecobiology of benthic polychaetes in Visakhapatnam Harbour (Bay of Bengal). Mar. Pollut. Bull. 1983, 14, 46–52. [Google Scholar] [CrossRef]
- Varadharajan, D.; Soundarapandian, P.; Gunalan, B.; Babu, R. Seasonal abundance of macro benthic composition and diversity along the south east coast of India. Eur. J. Appl. Sci. 2010, 2, 1–5. [Google Scholar]
- Natesan, U.; Kalaivani, S.; Kalpana, G. Pollution assessment of Ennore (India) creek using macrobenthos. J. Environ. Geol. 2017, 1, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Shokat, P.; Nabavi, S.M.B.; Savari, A.; Kochanian, P. Ecological quality of Bahrekan coast, by using biotic indices and benthic communities. Transit. Waters Bull. 2010, 4, 25–34. [Google Scholar]
- Grall, J.; Glémarec, M. Using biotic indices to estimate macrobenthic community perturbations in the Bay of Brest. Estuar. Coast. Shelf Sci. 1997, 44, 43–53. [Google Scholar] [CrossRef]
- Borja, A.; Franco, J.; Pérez, V. A Marine Biotic Index to Establish the Ecological Quality of Soft-Bottom Benthos Within European Estuarine and Coastal Environments. Mar. Pollut. Bull. 2000, 40, 1100–1114. [Google Scholar] [CrossRef]
- Teixeira, H.; Weisberg, S.B.; Borja, A.; Ranasinghe, J.A.; Cadien, D.B.; Velarde, R.G.; Lovell, L.L.; Pasko, D.; Phillips, C.A.; Montagne, D.E.; et al. Calibration and validation of the AZTI’s Marine Biotic Index (AMBI) for Southern California marine bays. Ecol. Indic. 2012, 12, 84–95. [Google Scholar] [CrossRef]
- Bigot, L.; Grémare, A.; Amoureux, J.-M.; Frouin, P.; Maire, O.; Gaertner, J.C. Assessment of the ecological quality status of soft-bottoms in Reunion Island (tropical Southwest Indian Ocean) using AZTI marine biotic indices. Mar. Pollut. Bull. 2008, 56, 704–722. [Google Scholar] [CrossRef] [PubMed]
- Reish, D. Benthic Invertebrates as Indicators of Marine Pollution: 35 Years of Study. In Proceedings of the OCEANS’86, Washington, DC, USA, 23–25 September 1986; pp. 885–888. [Google Scholar] [CrossRef]
- Reish, D.J. The relation of polychaetous Annelids to harbor pollution. Public Health Rep. 1955, 70, 1168–1174. [Google Scholar] [CrossRef]
- Keeley, N.B.; Macleod, C.K.; Forrest, B.M. Combining best professional judgement and quantile regression splines to improve characterisation of macrofaunal responses to enrichment. Ecol. Indic. 2012, 12, 154–166. [Google Scholar] [CrossRef]
- Blake, J.A. Life history analysis of five dominant infaunal polychaete species from the continental slope off North Carolina. J. Mar. Biol. Assoc. U. K. 1993, 73, 123–141. [Google Scholar] [CrossRef]
- Berkeley, E.; Berkeley, C. Some Further Records of Pelagic Polychaeta from the Northeast Pacific North Of Latitude 40° N. and East of Longitude 175° W., Together with Records of Siphonophora, Mollusca, and Tunicata from the Same Region. Can. J. Zool. 1960, 38, 787–799. [Google Scholar] [CrossRef]
- Hartman, O. Endemism in the North Pacific Ocean, with emphasis on the distribution of marine annelids, and descriptions of new or little known species. In Essays in the Natural Sciences in Honor of Captain Allan Hancock on the Occasion of His Birthday July 26; Allan Hancock Foundation, Ed.; University of Southern California Press: Los Angeles, CA, USA, 1955; pp. 39–60. [Google Scholar]
- Hartman, O. Polychaetous annelids collected by the USNS Eltanin and Staten Island cruises, chiefly from Antarctic Seas. Allan Hancock Monogr. Mar. Biol. 1967, 2, 1–387. [Google Scholar]
- Parapar, J.; Vijapure, T.; Moreira, J.; Sukumaran, S. A new species of Heterospio (Annelida, Longosomatidae) from the Indian Ocean. Eur. J. Taxon. 2016. [Google Scholar] [CrossRef] [Green Version]
- Blake, J.A. Family Paraonidae Cerruti 1909. In Taxonomic Atlas of the Benthic Fauna of the Santa Maria Basin and Western Santa Barbara Channel; 6—The Annelida Part 3. Polychaeta: Orbiniidae to Cossuridae; Blake, J.A., Hilbig, B., Scott, P.H., Eds.; Santa Barbara Museum of Natural History: Santa Barbara, CA, USA, 1996; pp. 27–70. [Google Scholar]
- Hartley, J.P. The family Paraonidae (Polychaeta) in british waters: A new species and new records with a key to species. J. Mar. Biol. Assoc. U. K. 1981, 61, 133–149. [Google Scholar] [CrossRef] [Green Version]
- Inajima, M. Paraonidae (Polychaeta) from Japan. Bull. Natl. Sci. Mus. Tokyo 1973, 16, 253–292. [Google Scholar]
- Ranauro, N.; Barroso, R.; Paiva, P.C.; Nogueira, J.M.d.M. On the genus Sabidius Strelzov, 1973 (Annelida: Paraonidae), with a redescription of the type species and the description of a new species. PLoS ONE 2020, 15, e0229717. [Google Scholar] [CrossRef] [Green Version]
- Arriaga-Hernández, S.; Hernández-Alcántara, P.; Solís-Weiss, V. Description and distribution of two new species of Paraonidae (Annelida: Polychaeta) from a lagoon-estuarine ecosystem in the Southern Gulf of Mexico. Zootaxa 2013, 3686, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Reuscher, M.G. Description of a new species of Aricidea (Polychaeta: Paraonidae) from Chinese Seas. Proc. Biol. Soc. Wash. 2013, 126, 276–283. [Google Scholar] [CrossRef]
- Erdoğan-Dereli, D.; Çinar, M.E.; Dağli, E. Cirrophorus species (Annelida: Polychaeta: Paraonidae) in the Sea of Marmara, with description of a new species. Zootaxa 2017, 4347, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Lovell, L. Paraonidae (Annelida: Polychaeta) Of The Andaman Sea, Thailand. Phuket Mar. Biol. Cent. Spec. Publ. 2002, 24, 33–56. [Google Scholar]
- Aguirrezabalaga, F.; Gil, J. Paraonidae (Polychaeta) from the Capbreton Canyon (Bay of Biscay, NE Atlantic) with the description of eight new species. Sci. Mar. 2009, 73, 631–666. [Google Scholar] [CrossRef]
- Langeneck, J.; Mazziotti, C.; Mikac, B.; Scirocco, T.; Castelli, A. Aricidea fragilis (Annelida: Paraonidae) in the Mediterranean Sea: Overlooked native or alien species? Eur. Zool. J. 2018, 85, 267–272. [Google Scholar] [CrossRef] [Green Version]
- Castelli, A. Paraonidae (Annelida, Polychaeta) des fonds meubles infralittoraux des cotes toscanes. Cah. de Biol. Mar. 1985, 26, 267–279. [Google Scholar]
- Laubier, L.; Ramos, J. Paraonidae (Polychètes Sédentaires) de Méditerranée. Bull. du Mus. Natl. d’Histoire Nat. Ser. 3 Zool. 1974, 168, 1097–1148. [Google Scholar]
- Hartman, O. Orbiniidae, Apistobranchidae, Paraonidae and Longosomidae. Allan Hancock Pac. Exped. 1957, 15, 211–393. [Google Scholar]
- Katzmann, W.; Laubier, L. Paraonidae (Polychètes Sédentaires) de l’Adriatique. Ann. der Nat. Mus. Wien 1975, 79, 567–588. [Google Scholar]
- Çinar, M.E.; Dagli, E.; Açik, S. Annelids (Polychaeta and Oligochaeta) from the Sea of Marmara, with descriptions of five new species. J. Nat. Hist. 2011, 45, 2105–2143. [Google Scholar] [CrossRef]
- Langeneck, J.; Barbieri, M.; Maltagliati, F.; Castelli, A. A new species of Cirrophorus (Annelida: Paraonidae) from Mediterranean organically enriched coastal environments, with taxonomic notes on the family. J. Mar. Biol. Assoc. U. K. 2017, 97, 871–880. [Google Scholar] [CrossRef]
- Mackie, A.S.Y. Paradoneis eliasoni sp. nov. (Polychaeta: Paraonidae) from northern European waters, with a redescription of Paradoneis lyra (Southern, 1914). Ophelia Suppl. 1991, 5, 147–155. [Google Scholar]
- de Leon-Gonzáles, J.A.; Díaz-Castañeda, V. A new species of Paraonidae (Polychaeta: Paraonidae) from the western coast of Baja California, Mexico. Proc. Biol. Soc. Wash. 2011, 124, 40–44. [Google Scholar] [CrossRef]
- López, E.; Sikorski, A. The Paraonidae (Annelida: Sedentaria) from Norway and adjacent seas, with two new species, four new records, and a redescription of Paraonides nordica Strelzov, 1968 based on type material. Zootaxa 2017, 4320, 41–67. [Google Scholar] [CrossRef]
- Hobson, K.D. Two new species and two new records of the Family Paraonidae (Annelida, Polychaeta) from the northeastern Pacific Ocean. Proc. Biol. Soc. Wash. 1972, 85, 549–556. [Google Scholar]
- Quintanar-Retama, O.; Hernández-Alcántara, P.; Solís-Weiss, V. Distribution patterns of three new species of Paradoneis (Annelida: Paraonidae) from the southern Gulf of Mexico, with a dichotomous key for the Grand Caribbean species. Mar. Biodivers. 2019, 49, 2851–2870. [Google Scholar] [CrossRef]
- Laubier, L. Sur quelques Aricidea (Polychètes, Paraonidae) de Banyuls-sur-Mer. Vie et Milieu Ser. A 1967, 18, 99–132. [Google Scholar]
- Jones, M.L. Paraonis pygoenigmatica new species, a new annelid from Massachusetts (Polychaeta: Paraonidae). Proc. Biol. Soc. Wash. 1968, 81, 323–334. [Google Scholar]
- Mclelland, J.; Gaston, G. 2 New Species Of Cirrophorus (Polychaeta, Paraonidae) From The Northern Gulf of-Mexico. Proc. Biol. Soc. Wash. 1994, 107, 524–531. [Google Scholar]
- Webster, H.E. The Annelida Chaetopoda of the Virginian coast. Trans. Albany Inst. 1879, 9, 202–269. [Google Scholar]
- Grube, A.E. Über ein paar neue Anneliden aus der Familie der Spiodeen. Jahres Bericht der Schlesischen Gesellschaft für Vaterländische Cultur 1872, 50, 57–58. [Google Scholar]
- McIntosh, W.C. On the Annelida obtained during the Cruise of H.M.S. ‘Valorous’ to Davis Strait in 1875. Trans. Linn. Soc. Lond. 1878, 1, 499–511. [Google Scholar] [CrossRef]
- Tauber, P. Annulata Danica. En Kritisk Revision af de in Danmark Fundne Annulata Chaetognatha, Gephyrea, Balanoglossi, Discophoreae, Oligochaeta, Gymnocopa og Polychaeta; Reitzer: Kobenhavn, Denmark, 1879; p. 143. [Google Scholar]
- Levinsen, G.M.R. Systematisk-geografisk Oversigt over de nordiske Annulata, Gephyrea, Chaetognathi og Balanoglossi. Videnskabelige Meddelelser fra den Naturhistoriske Forening i Kjöbenhavn 1884, 45, 92–350. [Google Scholar]
- Ehlers, E. Die bodensässigen Anneliden aus den Sammlungen der deutschen Tiefsee-Expedition. In Wissenschaftlichte Ergebnisse der Deutschen Tiefsee-Expedition auf dem Dampfer ‘Valdivia’ 1898–1899; Chun, C., Ed.; Gustav Fischer: Jena, Germany, 1908; Volume 16, pp. 1–168. [Google Scholar]
- Annenkova, N.P. Paraoniden der Meeren des Fernen Osten der USSR. Doklady Akademii Nauk SSSR 1934, 3, 656–658. [Google Scholar]
- Mesnil, F. Études de morphologie externe chez les Annélides. II. Remarques complémentaires sur les Spionidiens. La famille nouvelle des Disomidiens. La place des Aonides (sensu Tauber, Levinsen). Bull. Sci. Fr. Belg. 1897, 30, 83–100. [Google Scholar]
- Brasier, M.J.; Wiklund, H.; Neal, L.; Jeffreys, R.; Linse, K.; Ruhl, H.; Glover, A.G. DNA barcoding uncovers cryptic diversity in 50% of deep-sea Antarctic polychaetes. R. Soc. Open Sci. 2016, 3, 160432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilbig, B.; Blake, J.A. Deep-sea polychaete communities in the northeast Pacific Ocean off the Gulf of the Farallones, California. Bull. Mar. Sci. 2006, 78, 243–269. [Google Scholar]
- Reuscher, M.G. Biodiversity, Ecology, and Natural History of Polychaetous Annelids from the Gulf of Mexico. Ph.D. Thesis, Corpus Christi University, Corpus Christi, TX, USA, 2013. [Google Scholar]
- Ribeiro, R.P.; Alves, P.R.; de Almeida, Z.d.S.; Ruta, C. A new species of Paraonis and an annotated checklist of polychaetes from mangroves of the Brazilian Amazon Coast (Annelida, Paraonidae). ZooKeys 2018, 1–34. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Li, X.-Z. A report of the family Paraonidae (Annelida, Polychaeta) from China seas. Acta Zootaxon. Sin. 2007, 32, 275–282. [Google Scholar]
- Lovell, L.L.; Fitzhugh, K. Taking a closer look: An SEM review of Levinsenia species (Polychaeta: Paraonidae) reported from California. Zootaxa 2020, 4751, 256–275. [Google Scholar] [CrossRef] [PubMed]
- Langeneck, J.; Busoni, G.; Aliani, S.; Castelli, A. Deep-sea polychaetes (Annelida) from the Malta Escarpment (western Ionian Sea). Eur. Zool. J. 2017, 84, 142–152. [Google Scholar] [CrossRef]
- Langeneck, J.; Lezzi, M.; Pasqua, M.D.; Musco, L.; Gambi, M.C.; Castelli, A.; Giangrande, A. Non-indigenous polychaetes along the coasts of Italy: A critical review. Mediterr. Mar. Sci. 2020, 21, 238–275. [Google Scholar]
- Hartley, J.P. Cosmopolitan polychaete species: The status of Aricidea belgicae (Fauvel, 1936) and notes on the identity of A. suecica Eliason, 1920 (Polychaeta; Paraonidae). In Proceedings of the First International Polychaete Conference, Sydney, Australia, 4–9 July 1983; The Linnean Society of New South Wales: Sydney, Australia, 1984; pp. 7–20. [Google Scholar]
- Erdoğan-Dereli, D.; Çinar, M.E. The diversity of the genus Aricidea (Polychaeta: Paraonidae) from the Sea of Marmara, with descriptions of two new species and two new records for the Mediterranean fauna. Zootaxa 2020, 4844, 1–73. [Google Scholar] [CrossRef]
- Bakalem, A.; Gillet, P.; Pezy, J.-P.; Dauvin, J.-C. Inventory and the biogeographical affinities of Annelida Polychaeta in theAlgerian coastline (Western Mediterranean). Mediterr. Mar. Sci. 2020, 21, 157–182. [Google Scholar] [CrossRef]
- Gibbs, P.E. Recent additions to the marine fauna of Whitstable, with a description of Aricidea minuta southward (Annelida: Polychaeta). Ann. Mag. Nat. Hist. 1965, 8, 33–36. [Google Scholar] [CrossRef]
- Martín, D. La comunidad de Anélidos Poliquetos de las concreciones de algas calcáreas del litoral catalán. Caracterización de las especies. Publicaciones del Departamiento de Zoologia de Barcelona 1987, 13, 45–54. [Google Scholar]
- Box, A.; Martin, D.; Deudero, S. Changes in seagrass polychaete assemblages after invasion by Caulerpa racemosa var. cylindracea (Chlorophyta: Caulerpales): Community structure, trophic guilds and taxonomic distinctness. Sci. Mar. 2010, 74, 317–329. [Google Scholar] [CrossRef] [Green Version]
- Brito, M.C.; Martin, D.; Núñez, J. Polychaetes associated to a Cymodocea nodosa meadow in the Canary Islands: Assemblage structure, temporal variability and vertical distribution compared to other Mediterranean seagrass meadows. Mar. Biol. 2005, 146, 467–481. [Google Scholar] [CrossRef] [Green Version]
- Çinar, M.E.; Dagli, E. Polychaetes (Annelida: Polychaeta) from the Aegean and Levantine coasts of Turkey, with descriptions of two new species. J. Nat. Hist. 2013, 47, 911–947. [Google Scholar] [CrossRef]
- Langeneck, J.; Busoni, G.; Aliani, S.; Lardicci, C.; Castelli, A. Distribution and diversity of polychaetes along a bathyal escarpment in the western Mediterranean Sea. Deep Sea Res. Part I Oceanogr. Res. Pap. 2019, 144, 85–94. [Google Scholar] [CrossRef]
- Lardicci, C.; Como, S.; Corti, S.; Rossi, F. Changes and recovery of macrozoobenthic communities after restoration measures of the Orbetello Lagoon (Tyrrhenian coast, Italy). Aquat. Conserv. Mar. Freshw. Ecosyst. 2001, 11, 281–287. [Google Scholar] [CrossRef]
- Morri, C.; Castelli, A.; Diviacco, G.; Mori, M.; Bianchi, C.N. Zonazione di comunità bentiche lungo l’estuario della Magra (Mar Ligure). Atti Soc. Toscana Sci. Nat. Ser. B 1990, 97, 311–327. [Google Scholar]
- Magalhães, W.F.; Bailey-Brock, J.H.; Barroso, R. A new species of Paradoneis Hartman, 1965 (Annelida: Paraonidae) from Hawaii with notes on its reproductive strategy. Mar. Biodivers. 2019, 49, 1617–1625. [Google Scholar] [CrossRef]
- Gaston, G.R.; McLelland, J.A.; Heard, R.W. Feeding Biology, Distribution, and Ecology of Two Species of Benthic Polychaetes: Paraonis fulgens and Paraonis pygoenigmatica (Polychaeta: Paraonidae). Gulf Res. Rep. 1992, 8, 395–399. [Google Scholar] [CrossRef] [Green Version]
- Röder, H. Gangsysteme von Paraonis fulgens Levinsen 1883 (Polychaeta) in ökologischer, ethologischer und aktuopaläontologischer Sicht. Senckenbergiana Marit. 1971, 3, 3–51. [Google Scholar]
- Blake, J.A.; Baptiste, E.M. Life history studies on dominant polychaete species from Georges Bank. In Georges Bank Benthic Infauna Monitoring Program, Final Report for Third Year of Sampling, U.S.; Maciolek-Blake, N., Grassle, J.F., Neff, J.M., Eds.; Department of the Interior, Minerals Management Service: Herndon, VA, USA, 1985; Chapter 6; pp. 140–178. [Google Scholar]
- Levin, L.A.; Plaia, G.R.; Huggett, C.L. The influence of natural organic enhancement on life histories and community structure of bathyal polychaetes. In Reproduction, Larval biology and Recruitment of the Deep-Esa Benthos; Young, C.M., Eckelbarger, K.J., Eds.; Columbia University Press: New York, NY, USA, 1994; pp. 261–283. [Google Scholar]
- López-Jamar, E.; O’Connor, B.; González, G. Demography and gametogenic cycle of Paradoneis armata Glémarec (Polychaeta, Paraonidae) in Ría de la Coruña, North-West Spain. Ophelia 1987, 27, 127–136. [Google Scholar] [CrossRef]
- Giangrande, A. Polychaete reproductive patterns, life cycles and life histories: An overview. Oceanogr. Mar. Biol. Annu. Rev. 1997, 35, 323–386. [Google Scholar]
- Sendall, K.; Salazar-Vallejo, S. Revision of Sternaspis Otto, 1821 (Polychaeta, Sternaspidae). ZooKeys 2013, 286, 1–74. [Google Scholar] [CrossRef]
- Fiege, D. Sternaspidae Carus, 1863. In Handbook of Zoology Online. Annelida: Polychaetes; Westheide, W., Purschke, G., Eds.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2016; Available online: https://db.degruyter.com/view/Zoology/bp_029146-9_24 (accessed on 17 March 2021).
- Salazar-Vallejo, S.I. Six New Tropical Sternaspid Species (Annelida, Sternaspidae) with Keys to Identify Genera and Species. Zool. Stud. 2017, 56. [Google Scholar] [CrossRef]
- Fiege, D.; Barnich, R. A new genus and species of Sternaspidae (Annelida: Polychaeta) from the deep eastern Atlantic. Eur. J. Taxon. 2020, 699, 1–13. [Google Scholar] [CrossRef]
- Stimpson, W. Synopsis of the marine Invertebrata of Grand Manan: Or the region about the mouth of the Bay of Fundy, New Brunswick. Smithson. Contrib. Knowl. 1853, 6, 28–37. [Google Scholar]
- Petersen, M.E. Family Sternaspidae Carus, 1863. In Taxonomic Atlas of the Benthic Fauna of the Santa Maria Basin and Western Santa Barbara Channel; Volume 7–The Annelida Part 4. Polychaeta: Flabelligeridae to Sternaspidae; Blake, J.A., Hilbig, B., Scott, P.V., Eds.; Santa Barbara Museum of Natural History: Santa Barbara, CA, USA, 2000; pp. 331–336. [Google Scholar]
- Otto, A.G. Animalium maritimorum, nondum editorum, genera duo. I. Sternaspis thalassemoides. Verhandlungen der Kaiserlichen Leopoldinisch Carolinischen Academie der Naturforscher 1821, 2, 619–627. [Google Scholar]
- Sendall, K. Review and Revision of the Genus Sternaspis (Polychaeta: Sternaspidae) Using Cladistics on Morphological Characters. Master’s Thesis, University of Victoria, Greater Victoria, BC, Canada, 2006. [Google Scholar]
- Drennan, R.; Wiklund, H.; Rouse, G.W.; Georgieva, M.N.; Wu, X.; Kobayashi, G.; Yoshino, K.; Glover, A.G. Taxonomy and phylogeny of mud owls (Annelida: Sternaspidae), including a new synonymy and new records from the Southern Ocean, North East Atlantic Ocean and Pacific Ocean: Challenges in morphological delimitation. Mar. Biodivers. 2019, 49, 2659–2697. [Google Scholar] [CrossRef] [Green Version]
- Méndez, N.; Yáñez-Rivera, B. Distribution and morphometry of the deep-sea sternaspids, Sternaspis maior, Sternaspis uschakovi, and Caulleryaspis fauchaldi (Polychaeta), in Mexican Pacific waters. Bull. Mar. Sci. 2015, 91, 457–467. [Google Scholar] [CrossRef]
- Yoshino, K.; Nagayoshi, M.; Sato, M.; Katano, T.; Ito, Y.; Fuji, N.; Hamada, T.; Hayami, Y. Life history of Sternaspis costata (Sternaspidae: Polychaeta) in Ariake Bay, Japan. J. Mar. Biol. Assoc. U. K. 2016, 96, 647–655. [Google Scholar] [CrossRef]
- Fiege, D.; Bütfering, B. SEM-Investigations on the morphology and anatomy of Sternaspis scutata (Ranzani, 1817) (Annelida: Polychaeta). In Proceedings of the 6th International Polychete Conference, Held at Curitiba, Paraná, Brazil, 2–7 August 1998. Bull. Mar. Sci. 2000, 67, 662–663. [Google Scholar]
- Zhadan, A.E.; Tzetlin, A.B.; Salazar-Vallejo, S.I. Sternaspidae (Annelida, Sedentaria) from Vietnam with description of three new species and clarification of some morphological features. Zootaxa 2017, 4226, 75–92. [Google Scholar] [CrossRef] [PubMed]
- Stimpson, W. Descriptions of new species of marine Invertebrata from Puget Sound, collected by the Naturalists of the North-west Boundary Commission, A.H. Campbell, Esq., Commissioner. Proc. Acad. Nat. Sci. Phila. 1864, 16, 153–161. [Google Scholar]
- von Marenzeller, E. Südjapanische Anneliden I. Denkschriften der Kaiserlichen Akademie der Wissenschaften. Mathematisch Naturwissenschaftliche Classe 1879, 41, 109–154. [Google Scholar]
- Kobayashi, G.; Mukai, R.; Alalykina, I.; Miura, T.; Kojima, S. Phylogeography of benthic invertebrates in deep waters: A case study of Sternaspis cf. williamsae (Annelida: Sternaspidae) from the northwestern Pacific Ocean. Deep Sea Res. Part II Top. Stud. Oceanogr. 2018, 154, 159–166. [Google Scholar] [CrossRef]
- Salazar-Vallejo, S.; Buzhinskaja, G. Six new deep-water sternaspid species (Annelida, Sternaspidae) from the Pacific Ocean. ZooKeys 2013, 348, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Southern, R. Fauna of the Chilka Lake. Polychaeta of the Chilka Lake and also of fresh and brackish waters in other parts of India. Mem. Indian Mus. Calcutta 1921, 5, 563–693. [Google Scholar]
- Salazar-Vallejo, S.I. Three new polar species of Sternaspis Otto, 1821 (Polychaeta: Sternaspidae). Zootaxa 2014, 3861, 333. [Google Scholar] [CrossRef]
- Wu, X.; Salazar-Vallejo, S.; Xu, K. Two new species of Sternaspis Otto, 1821 (Polychaeta: Sternaspidae) from China seas. Zootaxa 2015, 4052, 373. [Google Scholar] [CrossRef]
- Wu, X.; Xu, K. Diversity of Sternaspidae (Annelida: Terebellida) in the South China Sea, with descriptions of four new species. Zootaxa 2017, 4244, 403–415. [Google Scholar] [CrossRef]
- Díaz-Díaz, O.; Rozbaczylo, N. Article Sternaspis chilensis n. sp., a new species from austral Chilean channels and fjords (Annelida, Sternaspidae). Zootaxa 2017, 4254, 269–276. [Google Scholar] [CrossRef]
- Salazar-Vallejo, S.I. Sternaspis piotrowskiae sp. nov. (Polychaeta: Sternaspidae) from the Philippine Islands. In The Coral Triangle: The 2011 Hearst Philippine Biodiversity Expedition; Williams, G.C., Terrence, M., Eds.; California Academy of Sciences: San Francisco, CA, USA, 2014; pp. 165–169. [Google Scholar]
- Ranzani, C. Descrizione di una nova specie del genere Thalassema. Opusculi Sci. 1817, I, 112–116. [Google Scholar]
- Wang, Z.; Liu, C.; Liu, H.; Xu, Z.; Wang, B.; Zheng, F. Complete mitochondrial genome of the Sternaspis scutata (Sternaspidae, Annelida). Mitochondrial DNA Part B 2019, 4, 1700–1701. [Google Scholar] [CrossRef]
- Preetha Mini Jose, H.M.; Muthuvelu, S.; Sivaraj, S.; Murugesan, P. A note on the occurrence of Sternaspis scutata (Renier 1807)—A sternaspid polychaete from Sundarban Mangroves, West Bengal, India. J. Bombay Nat. Hist. Soc. 2014, 111, 62–64. [Google Scholar] [CrossRef]
- Pillai, N.G.K. On some benthic polychaetes from Cochin estuary. J. Mar. Biol. Assoc. of India 2001, 43, 120–135. [Google Scholar]
- Joydas, T.V.; Damodaran, R. Infaunal macrobenthos along the shelf waters of the west coast of India, Arabian Sea. Indian J. Mar. Sci. 2009, 38, 14. [Google Scholar]
- Abdelnaby, F.A. First record of Sternaspis scutata (Polychaeta, Sternaspidae) in Egyptian waters. Egypt. J. Aquat. Biol. Fish. 2020, 24, 317–326. [Google Scholar] [CrossRef] [Green Version]
- Townsend, M.; Worsfold, T.M.; Smith, P.R.J.; Martina, L.J.; McNeill, C.L.; Kendall, M.A. Occurrence of Sternaspis scutata (Polychaeta: Sternaspidae) in the English Channel. Cah. Biol. Mar. 2006, 47, 281–285. [Google Scholar]
- Wang, J.-B.; Li, X.-Z.; Wang, H.-F. Ecological characteristics of dominant polychaete species from the Jiaozhou Bay. Acta Zool. Sin. 2006, 52, 63–69. [Google Scholar]
- Por, F.D. Lessepsian Migration. The influx of Red Sea Biota into the Mediterranean by Way of the Suez Canal; Springer: Berlin, Germany, 1978; p. 228. [Google Scholar]
- Rietsch, M. Étude sur le Sternaspis scutata. Ann. des Sci. Nat. Sixieme Sér. Zool. 1882, 13, 1–84. [Google Scholar]
- Fauchald, K.; Jumars, P.A. The Diet of Worms: A Study of Polychaete Feeding Guilds. Oceanogr. Mar. Biol. Annu. Rev. 1979, 17, 193–284. [Google Scholar]
- Dales, R.P. The polychaete stomodeum and the inter-relationships of the families of Polychaeta. Proc. Zool. Soc. 1962, 39, 389–428. [Google Scholar] [CrossRef]
- Dorgan, K.M.; D’Amelio, C.; Lindsay, S.M. Strategies of burrowing in soft muddy sediments by diverse polychaetes. Invertebr. Biol. 2016, 135, 287–301. [Google Scholar] [CrossRef]
- Self, R.F.L.; Jumars, P.A. Cross-phyletic patterns of particle selection by deposit feeders. J. Mar. Res. 1988, 46, 119–143. [Google Scholar] [CrossRef]
- Lim, H.S.; Hong, J.S. Distribution and growth pattern of Sternaspis scutata (Polychaeta: Sternaspidae) in Chinhae Bay, Korea. J. Korean Fish. Soc. 1996, 29, 537–545. [Google Scholar]
- Ohwada, T. Mesh size for settlement and recruitment studies of polychaetes with reference to the effect of fixation on washing bottom samples. Asian Mar. Biol. 1988, 5, 35–40. [Google Scholar]
- Blake, J.A.; Göransson, P. Redescription of Tharyx killariensis (Southern) from Ireland and description of two new species of Tharyx from the Kattegat, Sweden (Polychaeta, Cirratulidae). Zootaxa 2015, 4039, 501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jägersten, G. Studies on the morpgology, larval development and biology of Protodrilus. Zool. Bidrag från Uppsala 1952, 29, 426–512. [Google Scholar]
- Helm, C.; Capa, M. Comparative analyses of morphological characters in Sphaerodoridae and allies (Annelida) revealed by an integrative microscopical approach. Front. Mar. Sci. 2015, 1, 82. [Google Scholar] [CrossRef] [Green Version]
- Surugiu, V.; Capa, M. The Occurrence of Amphiglena mediterranea (Leydig, 1851) (Annelida: Sabellidae) at the Romanian Coast of the Black Sea: A Case on an Unsuccessful Invasion? Russ. J. Biol. Invasions 2020, 11, 293–299. [Google Scholar] [CrossRef]
- Nygren, A. Cryptic polychaete diversity: A review. Zool. Scr. 2014, 43, 172–183. [Google Scholar] [CrossRef]
- Choi, H.K.; Kim, H.J.; An, Y.R.; Kim, H. The complete mitochondrial genome of Cirriformia cf. tentaculata (Polychaeta, Cirratulidae) from Dok-Do, Korea. Mitochondrial DNA B 2017, 2, 847–848. [Google Scholar]
- Kim, H.; Min, G.-S.; Choi, H.K. The complete mitochondrial genome of Korean endemic species, Timarete posteria (Polychaeta, Cirratulidae). Mitochondrial DNA Part B 2019, 4, 501–502. [Google Scholar] [CrossRef]
- Di Giuseppe, G.; Barbieri, M.; Vallesi, A.; Luporini, P.; Dini, F. Phylogeographical pattern of Euplotes nobilii, a protist ciliate with a bipolar biogeographical distribution. Mol. Ecol. 2013, 22, 4029–4037. [Google Scholar] [CrossRef] [PubMed]
- Flot, J.-F.; Couloux, A.; Tillier, S. Haplowebs as a graphical tool for delimiting species: A revival of Doyle’s “field for recombination” approach and its application to the coral genus Pocillopora in Clipperton. BMC Evol. Biol. 2010, 10, 372. [Google Scholar] [CrossRef] [PubMed]
- Christie, G. A Comparative Study of the Reproductive Cycles of Three Northumberland Populations of Chaetozone Setosa (Polychaeta:Cirratulidae). J. Mar. Biol. Assoc. U. K. 1985, 65, 239–254. [Google Scholar] [CrossRef]
- Day, J.H. A Monograph on the Polychaeta of Southern Africa. Part 2. Sedentaria; Trustees of the British Museum (Natural History): London, UK, 1967; p. 419. [Google Scholar]
- Olive, P.J.W. Ovary structure and oogenesis in Cirratulus cirratus (Polychaeta: Cirratulidae). Mar. Biol. 1971, 8, 243–259. [Google Scholar] [CrossRef]
- Parapar, J.; Zhadan, A.; Tzetlin, A.B.; Vortsepneva, E.V.; Moreira, J. Exploring the anatomy of Cossura pygodactylata Jones, 1956 (Annelida, Cossuridae) using micro-computed tomography, with special emphasis on gut architecture. Mar. Biodivers. 2018, 48, 751–761. [Google Scholar] [CrossRef]
- Zhadan, A.; Vortsepneva, E.; Tzetlin, A. Three-dimensional reconstruction of the musculature of Cossura pygodactylata Jones, 1956 (Annelida: Cossuridae). Zool. Anz. 2014, 253, 181–191. [Google Scholar] [CrossRef]
- Pamungkas, J.; Glasby, C.J.; Read, G.B.; Wilson, S.P.; Costello, M.J. Progress and perspectives in the discovery of polychaete worms (Annelida) of the world. Helgol. Mar. Res. 2019, 73, 10. [Google Scholar] [CrossRef]







| Geographic Distribution | No. of Species | Species |
|---|---|---|
| Pacific Ocean | 18 | |
| NE Pacific, North American coast | 7 | C. bansei, C. brunnea, C. candida, C. modica, C. pygodactylata, C. rostrata, C. sima |
| SE Pacific, South American coast | 4 | C. abyssalis, C. alba, C. chilensis, C. laeviseta |
| NW Pacific, Japan, China | 3 | C. coasta, C. duplex, C. aciculata |
| SW Pacific, Australia, New Zeeland, Fiji | 4 | C. consimilis, C. hutchingsae, C. keablei, C. queenslandensis |
| Atlantic Ocean | 10 | |
| NW Atlantic, North American coast | 3 | C. delta, C. longocirrata, C. pseudokaina |
| NW Atlantic, Caribbean Sea, South American coast | 2 | C. pettibonea, C. ginesi |
| NE Atlantic, Mediterranean Sea | 1 | C. soyeri |
| SW Atlantic, South American coast | 2 | C. heterochaeta, C. yacy |
| SE Atlantic, Congo deep-sea fan | 2 | C. flabelligera, C. platypus |
| Indian Ocean | 2 | C. dayi, C. dimorpha |
| Species | Distribution | Depth (m) | Type Locality |
|---|---|---|---|
| Caulleryaspis fauchaldi Salazar-Vallejo and Buzhinskaja, 2013 | NE Pacific, Oregon to Southern California | 1387–2800 | Oregon, Cascadia Abyssal Plain, W of Yaquina Bay, 44°30.0′ N, 125°34.0′ W to 44°39.0′ N, 125°33.2′ W, 2800 m |
| Caulleryaspis gudmundssoni Sendall and Salazar-Vallejo, 2013 | N Atlantic, Iceland | 452–1207 | Iceland, southeast of Vestmannaey jar, 63°02.30′ N, 21°50.80′ W, 1072 m |
| Caulleryaspis laevis (Caullery, 1944) | Andaman Sea to South China Sea, Southeastern Australia | 39–147 | Sumbawa Island, Bay of Bima, 08°27.5′ S, 118°43.5′ E, 55 m |
| Caulleryaspis nuda Salazar-Vallejo and Buzhinskaja, 2013 | NE Pacific, off Oregon NW Pacific, Sea of Okhotsk, off Kuril Islands as C. nuda | 2519 | Off Oregon, 48°9.0′ N, 127°4.2′ W, 2519 m |
| Caulleryaspis villamari Salazar-Vallejo, 2017 | E Pacific, off northern Peru | 160 | Peru, off Isla Lobos de Tierra, 06°27′ S, 80°56′ W to 06°23′ S, 80°55′ W, 160 m |
| Mauretanaspis longichaeta Fiege and Barnich, 2020 | CE and SE Atlantic, off Mauritania and Angola | 2700–4400 | Off Mauretania; 18°21.27′ N, 17°57.44′ W; 2712 m |
| Petersenaspis capillata (Nonato, 1966) | SW Atlantic, Brazil | (intertidal) 52–138 | Brazil, Vitoria Island, 23°45′18” S, 44°00′54” W, 52 m |
| Petersenaspis deani Salazar-Vallejo, 2017 | E Pacific, Costa Rica | 22 | Costa Rica, Golfo de Nicoya, 09°44′00″ N, 84°59′25″ W, 22 m |
| Petersenaspis harrisae Salazar-Vallejo, 2017 | SW Indian Ocean, between South Africa and Madagascar | 15–128 | South Africa, off Durban, 29°11′ S, 31°37′ E, 15 m |
| Petersenaspis palpallatoci Sendall and Salazar-Vallejo, 2013 | Philippine Islands to Malaysia | 5.5–93 | Philippine Islands, Sibuyan Sea, E off Kalibo, 11°42.6′ N, 122°31.5′ E, 93 m |
| Petersenaspis salazari Wu and Xu, 2017 | Northern South China Sea | 174–182 | South China Sea, 19°30′ N, 112°30′ E, 174 m |
| Sternaspis affinis Stimpson, 1864 | NE Pacific Ocean, Alaska to Gulf of California | 95–154 | Canada, British Columbia, Strait of Georgia |
| Sternaspis Africana Augener, 1918 | E Atlantic, W African coast from Ghana to Angola | 20–70 | Angola, St. Paul Loanda |
| Sternaspis andamanensis Sendall and Salazar-Vallejo, 2013 | Indian Ocean, Andaman Sea and Malaysia, South China Sea | 5–45 | Andaman Sea, Thailand, 45 m |
| Sternaspis annenkovae Salazar-Vallejo and Buzhinskaja, 2013 | NW Pacific, east of northern Kuril I lands SW Pacific, Australia, off E Gippsland | 3980–4070 | East of northern Kuril Island, 49°46′01″ N, 157°48′06″ E, 3980–4070 m |
| Sternaspis britayevi Zhadan, Tzetlin and Salazar-Vallejo, 2017 | South China Sea, Gulf of Tonkin | 65 | South China Sea, Gulf of Tonkin, 19°16′ N, 107°19′ E, 65 m |
| Sternaspis buzhinskajae Salazar-Vallejo, 2014 | Arctic Ocean, Okhotsk, Chukchi, Bering and Beaufort Seas | 3.5–90 | Alaska, Beaufort Sea, off Point Barrow, 3.5–5.0 m |
| Sternaspis chilensis Díaz-Díaz and Rozbaczylo, 2017 | SE Pacific, Austral Chilean channel and fjord region | 50–515 | Chile, Magellan Province, Moraleda Channel, 45°06′ S, 73°38′ W, 250 m |
| Sternaspis chinensis Wu, Salazar-Vallejo and Xu, 2015 | Yellow and East China Sea, Bohai Sea | 8–63.7; 500 | Bohai Sea, 38°38′ N, 120°23′ E, 39 m |
| Sternaspis costata von Marenzeller, 1879 | NW Pacific, Southern Sakhalin Island to Philippines, South China Sea | 15–70 | Japan, Honshu Island, Chiba, Boso Peninsula |
| Sternaspis fossor Stimpson, 1853 | NW Atlantic, Canada to northeastern United States | 20–153 | Canada, Nova Scotia, 88 km E Cape Sable, 153 m |
| Sternaspis islandica Malmgren, 1867 | NE Atlantic, Norwegian Sea, Iceland and Faroe Islands | 7–226 | Iceland, Berufjord, 64°48′ N, 14°30′ W, 27–55 m |
| Sternaspis lindae Salazar-Vallejo, 2017 | E Pacific, Gulf of Panama to NW Colombia | 119–275 | Panama, 08°39.3′ N, 79°31.7′ W, 22–27 m |
| Sternaspis liui Wu, Salazar-Vallejo and Xu, 2015 | Yellow Sea | 21–43.3 | Yellow Sea, 35°59′ N, 122°00′ E; 43.3 m |
| Sternaspis londognoi Salazar-Vallejo, 2017 | Gulf of Mexico and Caribbean Sea | intertidal-344 | SW Caribbean, Panama, Caledonia Bay, intertidal |
| Sternaspis maior Chamberlin, 1919 | E Pacific, Central Gulf of California | 180–1158 | Gulf of California, Sinaloa, Off Isla Altamura, 24°52′ N, 108°58′ W, 830 m |
| Sternaspis maureri Salazar-Vallejo and Buzhinskaja, 2013 | E Pacific and SW Pacific | 795–6489 | Peru, W of Trujillo, 08°16′ S, 81°05′ W, 6156–6489 m |
| Sternaspis nana Zhadan, Tzetlin and Salazar-Vallejo, 2017 | Vietnam, Nha Trang Bay, South China Sea | 15–40 | South China Sea, Nha Trang Bay, 12°11′50″ N, 109°15′06.6″ E, 23 m |
| Sternaspis papillosa Zhadan, Tzetlin and Salazar-Vallejo, 2017 | Vietnam, Nha Trang Bay, South China Sea | 11–40 | South China Sea, Nha Trang Bay, 12°10′24″ N, 109°15′42″ E, 21.1 m |
| Sternaspis piotrowskiae Salazar-Vallejo, 2014 | Philippine Islands | 272–636 | Philippines, Luzon Island, Balayan Bay, 13.82°N, 120.75°E, 272–291 m |
| Sternaspis princeps Selenka, 1885 | SW Pacific, New Zealand, off North Island | 1274 | New Zealand, North Island, NE off Gisborne, 37°34′ S, 179°22′ E, 1274 m |
| Sternaspis radiate Wu and Xu, 2017 | Northern South China Sea | 16–105.5 | South China Sea 22°15′ N, 115°00′ E, 41 m |
| Sternaspis rietschi Caullery, 1944 | Arafura Sea, Indonesia, off Wokam Island | 1788 | Indonesia, W of Wokam Island, 5°46′ S, 134°00′ E, 1788 m |
| Sternaspis scutata (Renier in Ranzani, 1817) | Mediterranean Sea | 9–36 | Aegean Sea, Izmir Bay, 38°30′00″ N, 26°50′00”E, 33 m |
| Sternaspis sendalli Salazar-Vallejo, 2014 incl. S. monroi Salazar-Vallejo, 2014 | Southern Ocean, South Georgia to Antarctic Peninsula and Weddell Sea | 160–1500 | South Orkneys, Scotia Sea |
| Sternaspis sherlockae Salazar-Vallejo, 2017 | Red Sea | Presumed shallow water | Red Sea |
| Sternaspis spinosa Sluiter, 1882 | W Pacific from South China Sea to Andaman Sea and Coral Sea off Australia | 7–110 | Indonesia, Java, Bay of Batavia, “Batavia Roads”, outside Jakarta, 30 m |
| Sternaspis sunae Wu and Xu, 2017 | South China Sea | 224 | South China Sea, Nansha Islands, 224 m |
| Sternaspis thalassemoides Otto, 1821 | NE Atlantic and Mediterranean Sea | n.d. | Tyrrhenian Sea, Naples |
| Sternaspis thorsoni Sendall and Salazar-Vallejo, 2013 | Indian Ocean, Arabian/Iranian Gulf to Andaman Sea | 3–110 | Gulf of Oman, off Jask, 25°45′ N, 57°12′ E, 110 m |
| Sternaspis uschakovi Salazar-Vallejo and Buzhinskaja, 2013 | NW and E Pacific, Okhotsk Sea, California, Gulf of California, W Mexico | 592–2548 | Okhotsk Sea, 55°13′ N, 146°52′ E, 592 m |
| Sternaspis williamsae Salazar-Vallejo and Buzhinskaja, 2013 | NE Pacific, Oregon to California NW Pacific, off Kuril Islands as S. cf. williamsae | 1000–2800 | Oregon, W of Yaquina Bay, 44°30.0′ N, 125°34.0′ W to 44°39.0′ N, 125°33.2′ W, 2800 m |
| Sternaspis wui Wu and Xu, 2017 | Northern South China Sea | 173–230 | South China Sea, 17°45′ N, 110°30′ E, 230 m |
| Species | Distribution | Depth (m) |
|---|---|---|
| Caulleryaspis fauchaldi Salazar-Vallejo and Buzhinskaja, 2013 | NE Pacific, Oregon to Southern California | 1387–2800 |
| Caulleryaspis gudmundssoni Sendall and Salazar-Vallejo, 2013 | North Atlantic, Iceland | 452–1207 |
| Caulleryaspis nuda Salazar-Vallejo and Buzhinskaja, 2013 | NE Pacific, off Oregon | 2519 |
| Mauretanaspis longichaeta Fiege and Barnich, 2020 | CE and SE Atlantic, off Mauritania and Angola | 2700–4400 |
| Sternaspis annenkovae Salazar-Vallejo and Buzhinskaja, 2013 | NW Pacific, east of northern Kurile Island | 3980–4070 |
| Sternaspis chilensis Díaz-Díaz and Rozbaczylo, 2017 | SE Pacific, Austral Chilean channel and fjord region | 50–515 |
| Sternaspis maior Chamberlin, 1919 | E Pacific, Central Gulf of California | 180–1158 |
| Sternaspis maureri Salazar-Vallejo and Buzhinskaja, 2013 | E and SW Pacific | 795–6489 |
| Sternaspis piotrowskiae Salazar-Vallejo, 2014 | Philippine Islands | 272–636 |
| Sternaspis princeps Selenka, 1885 | SW Pacific, New Zealand | 1274 |
| Sternaspis rietschi Caullery, 1944 | Arafura Sea, off Wokam Island | 1788 |
| Sternaspis sendalli Salazar-Vallejo, 2014 | Southern Ocean and Antarctic Peninsula | 160–1500 |
| Sternaspis uschakovi Salazar-Vallejo and Buzhinskaja, 2013 | N Pacific, Okhotsk Sea, California, Gulf of California | 592–2548 |
| Sternaspis williamsae Salazar-Vallejo and Buzhinskaja, 2013 | NE Pacific, Oregon to California | 1000–2800 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grosse, M.; Zhadan, A.; Langeneck, J.; Fiege, D.; Martínez, A. Still Digging: Advances and Perspectives in the Study of the Diversity of Several Sedentarian Annelid Families. Diversity 2021, 13, 132. https://doi.org/10.3390/d13030132
Grosse M, Zhadan A, Langeneck J, Fiege D, Martínez A. Still Digging: Advances and Perspectives in the Study of the Diversity of Several Sedentarian Annelid Families. Diversity. 2021; 13(3):132. https://doi.org/10.3390/d13030132
Chicago/Turabian StyleGrosse, Maël, Anna Zhadan, Joachim Langeneck, Dieter Fiege, and Alejandro Martínez. 2021. "Still Digging: Advances and Perspectives in the Study of the Diversity of Several Sedentarian Annelid Families" Diversity 13, no. 3: 132. https://doi.org/10.3390/d13030132
APA StyleGrosse, M., Zhadan, A., Langeneck, J., Fiege, D., & Martínez, A. (2021). Still Digging: Advances and Perspectives in the Study of the Diversity of Several Sedentarian Annelid Families. Diversity, 13(3), 132. https://doi.org/10.3390/d13030132