
Do Anthropogenic Activities Affect Floristic Diversity and Vegetation Structure More Than Natural Soil Properties in Hyper-Arid Desert Environments?


Abstract
:1. Introduction
2. Materials and Methods
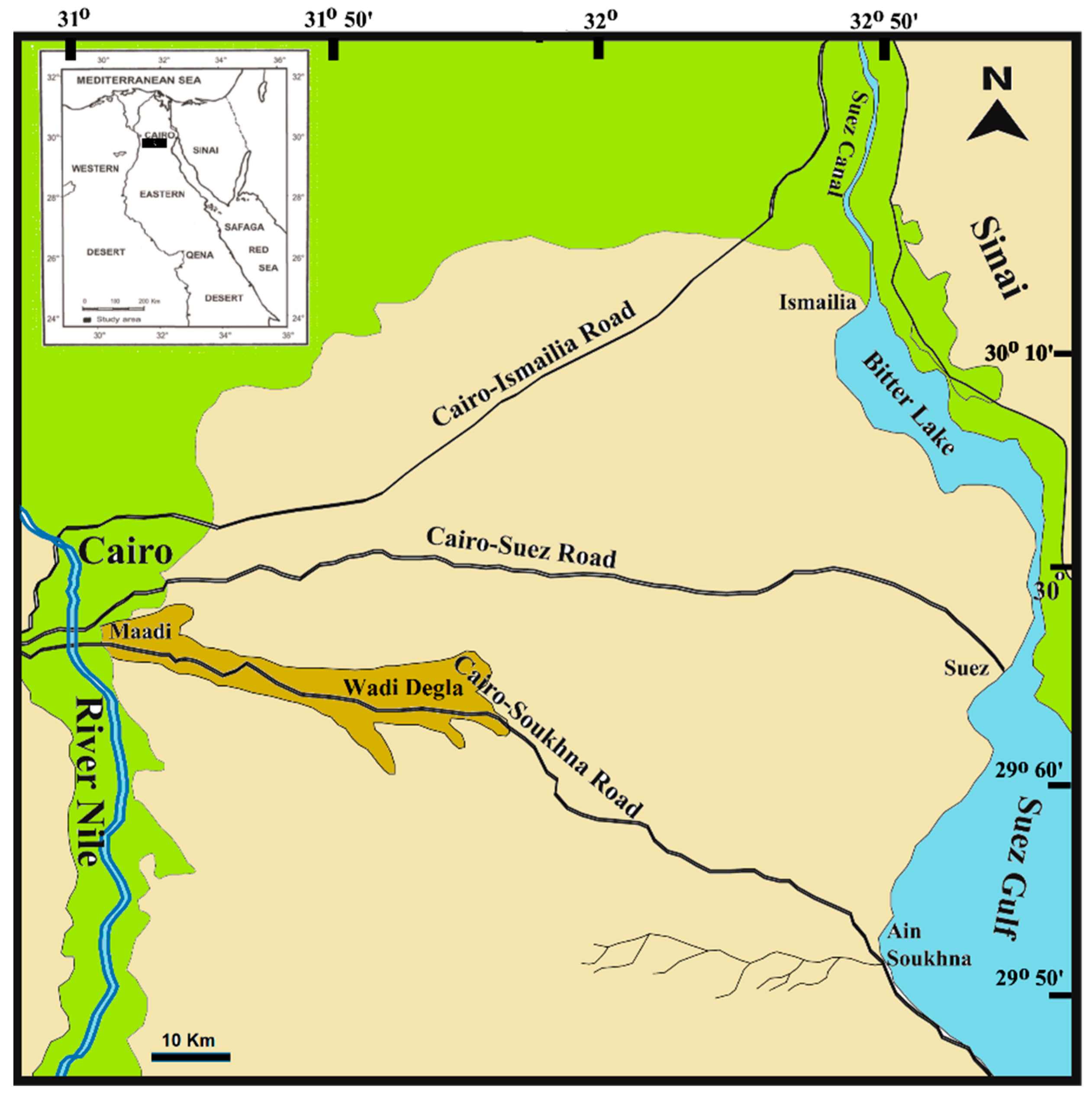
2.1. The Study Areas
2.2. Data Collection and Floristic Analysis
2.3. Soil Sampling and Analysis
2.4. Vegetation Data Analysis
2.5. Species Diversity
3. Results
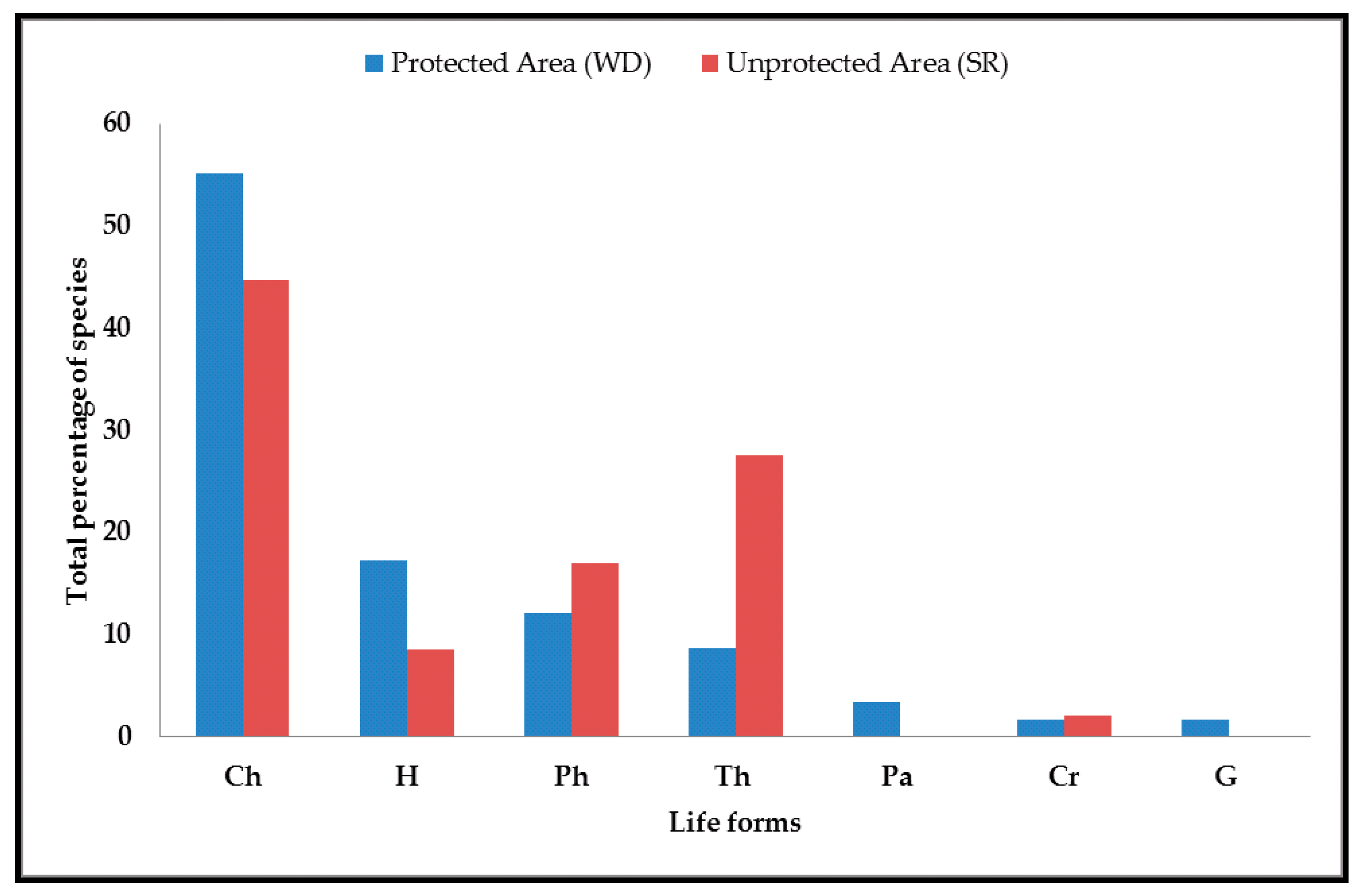
3.1. Characteristics of the Flora
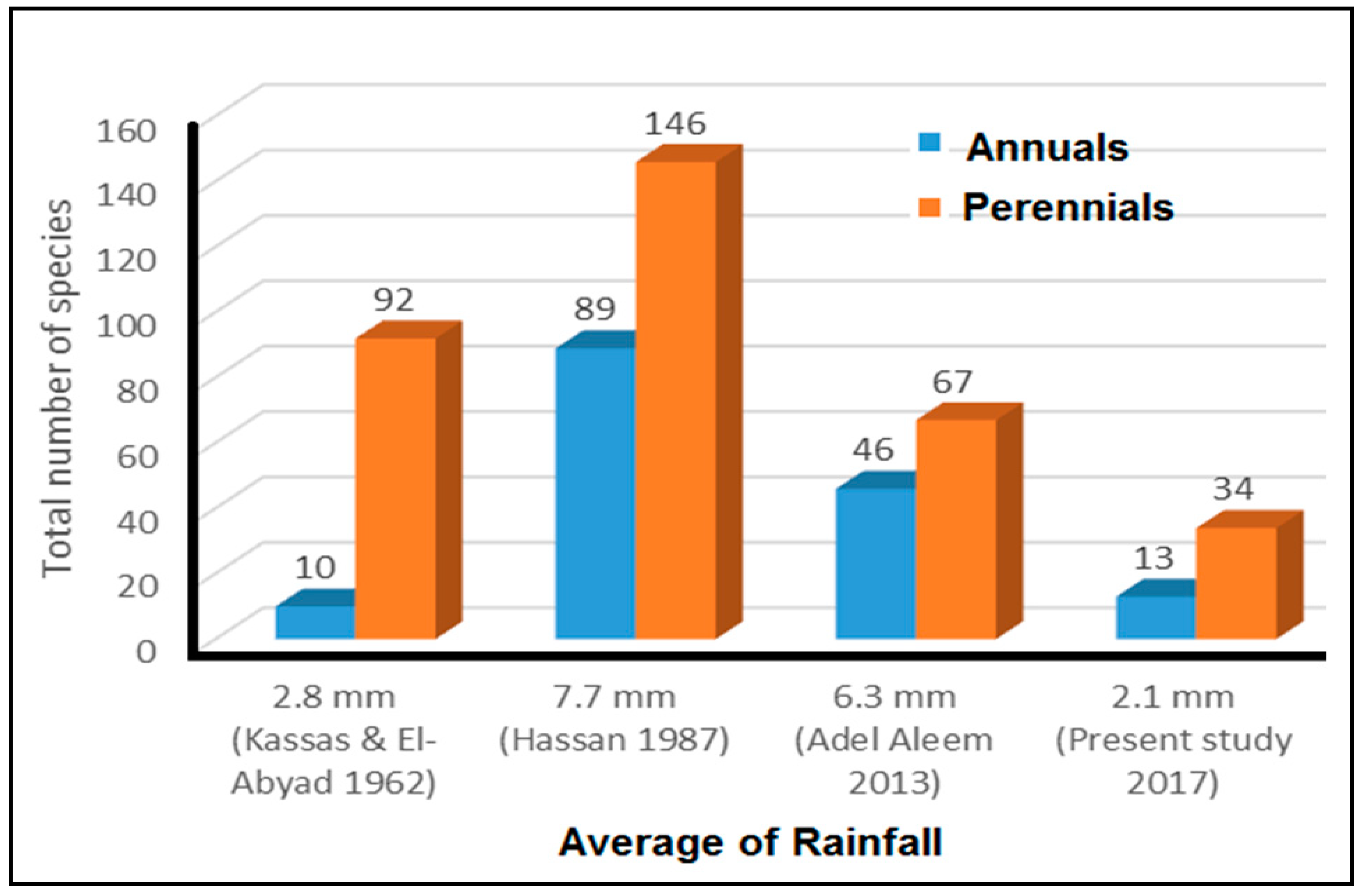
3.2. Floristic Changes over the Past Five Decades
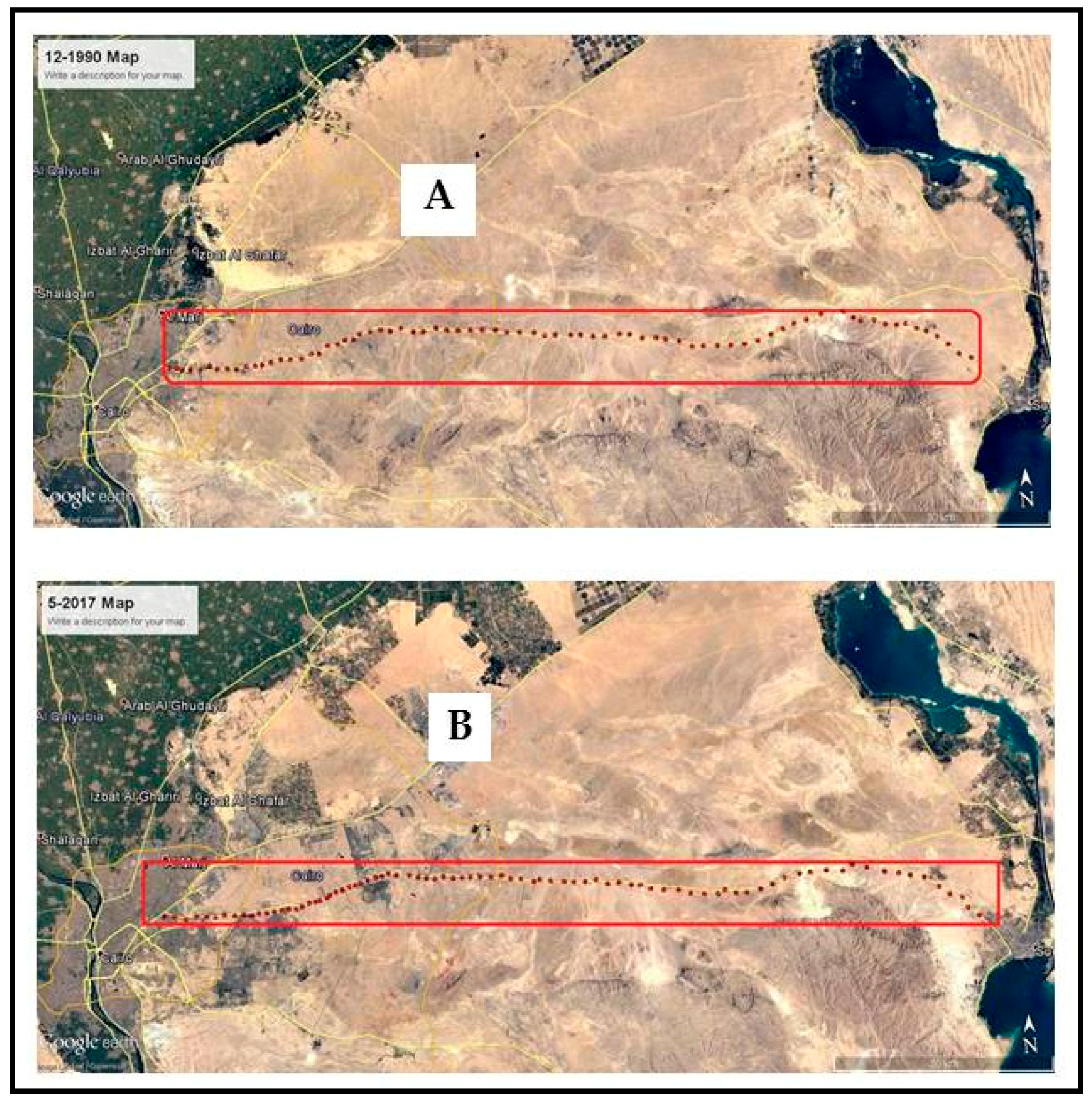
3.2.1. Cairo-Suez Road (Unprotected Area)
3.2.2. Wadi Degla Protected Area
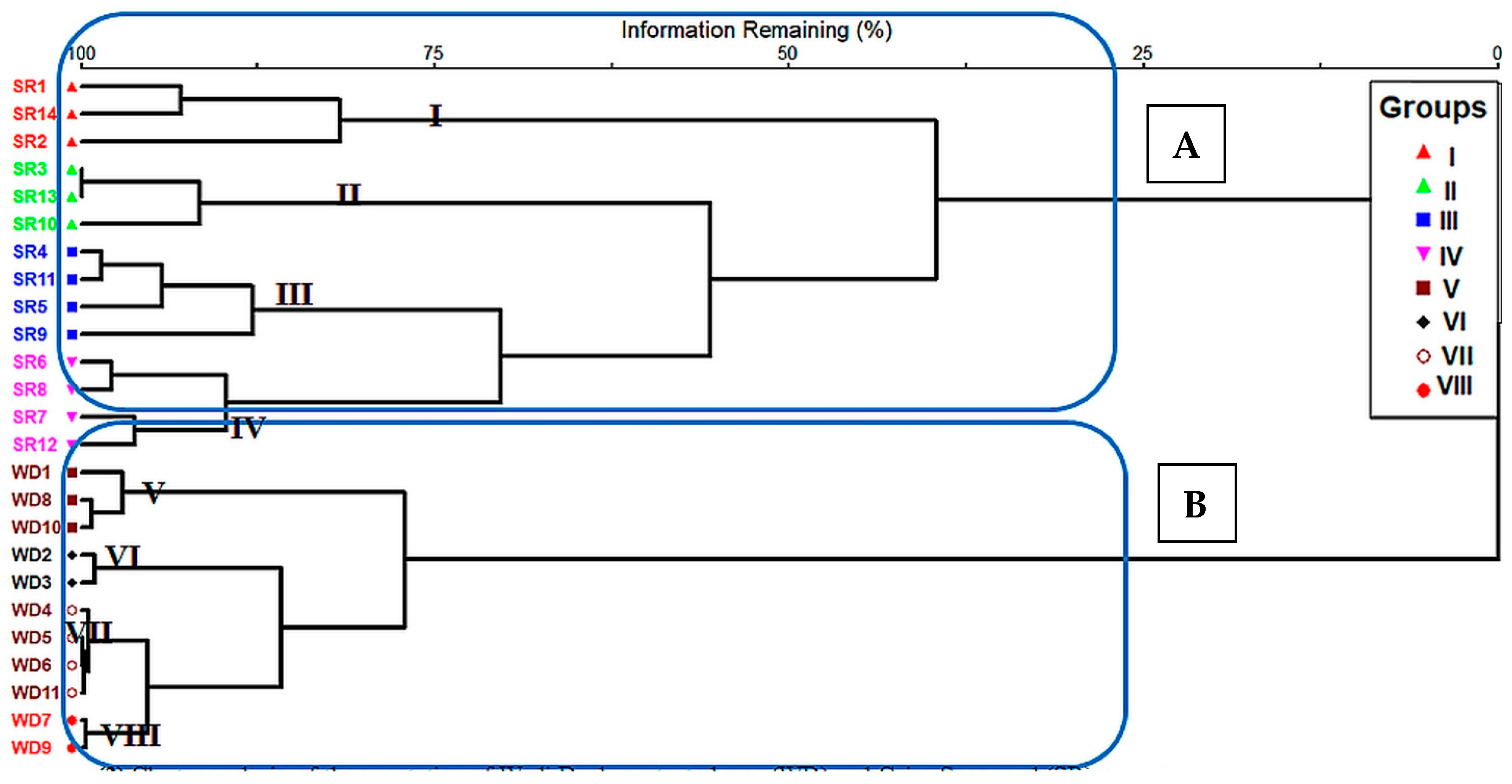
3.3. Classification, Indicator Species Analysis, and Description of Vegetation Groups
3.4. Description of Vegetation Groups
3.4.1. Cairo-Suez Road Unprotected Area
3.4.2. Wadi Degla Protected Area
3.5. Correlations between Soil Factors and Anthropogenic Variables
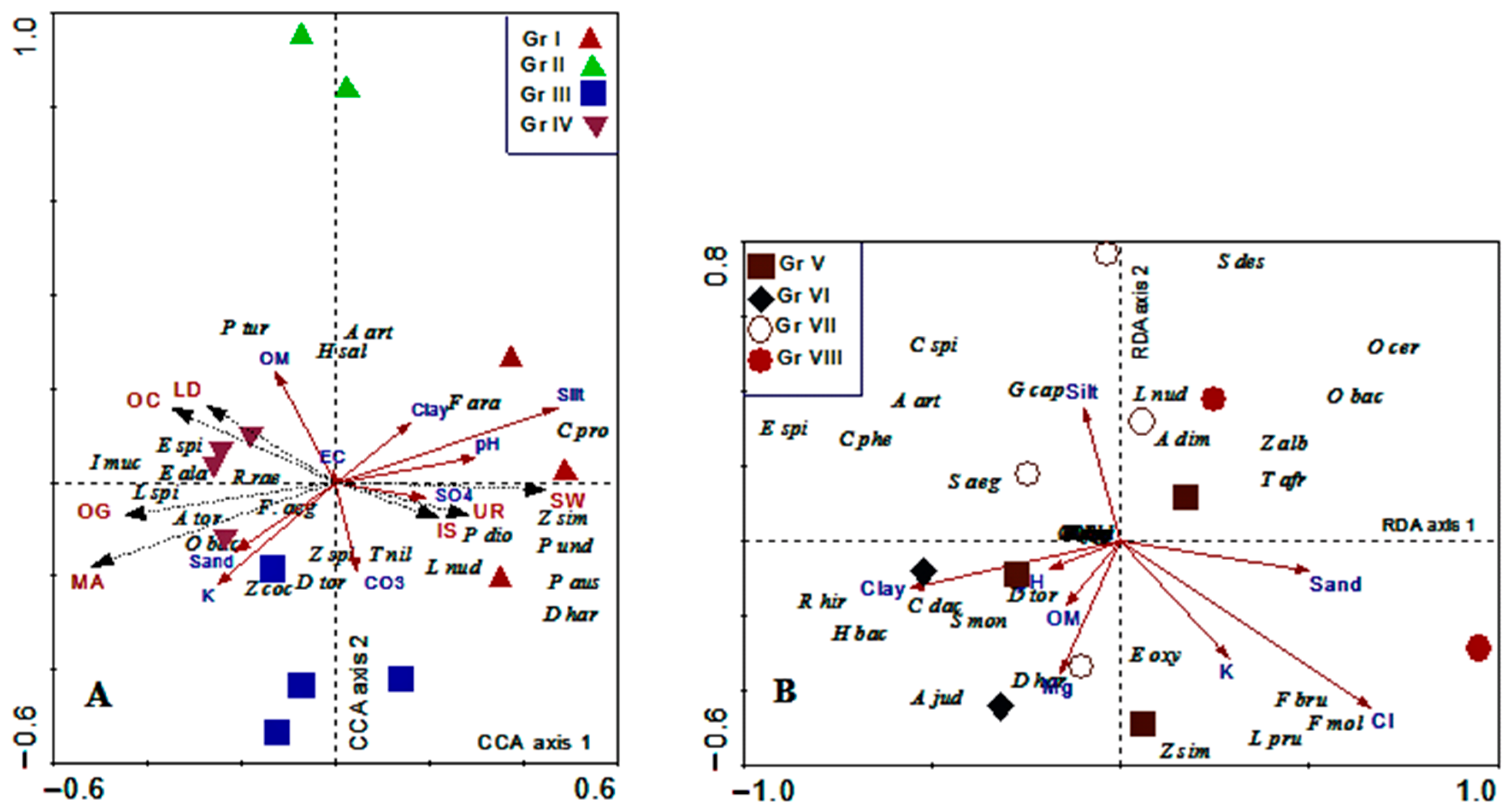
3.6. Vegetation–Soil–Anthropogenic Correlations
3.6.1. Cairo-Suez Road (Unprotected Area)
3.6.2. Wadi Degla Protected Area
3.6.3. Impact of Anthropogenic Variables (pCCA)
3.7. Correlations between Species Diversity, Soil Factors, and Anthropogenic Variables
4. Discussion
4.1. Floristic Composition, Biological Spectrum, and Chorological Affinities
4.2. Classification of the Vegetation
4.3. Soil–Vegetation Relationships
4.4. Anthropogenic Factors and Their Contribution
4.5. Plant Diversity and Its Significance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Millennium Ecosystem Assessment. Ecosystems and Human Well-Being: Desertification Synthesis; World Resources Institute: Washington DC, USA, 2005. [Google Scholar]
- Dregne, H.E. Land Degradation in the Drylands. Arid Land Res. Manag. 2002, 16, 99–132. [Google Scholar] [CrossRef]
- Dansereau, P. The origin and growth of plant communities. In Growth in Living System: Proceedings of International Symposium on Growth, Purdue University, Indiana; Zarrow, M.X., Ed.; Basic Books: New York, NY, USA, 1960; pp. 563–603. [Google Scholar]
- Moustafa, A.A.; Abd El-Wahab, R.H.; Zaghloul, M.S. Conservation and Sustainable Use of Medicinal Plants in Arid and Semi-Arid Ecosystems of Egypt; Final report; Egyptian Environmental Affairs Agency (EEAA), United Nations Development Programme (UNDP) and Global Environmental Facilities (GEF): Giza, Egypt, 1999. [Google Scholar]
- Nakahama, N.; Hirasawa, Y.; Minato, T.; Hasegawa, M.; Isagi, Y.; Shiga, T. Recovery of genetic diversity in threatened plants through use of germinated seeds from herbarium specimens. Plant Ecol. 2015, 216, 1635–1647. [Google Scholar] [CrossRef] [Green Version]
- Enright, N.J.; Miller, B.P.; Akhter, R. Desert vegetation and vegetation-environment relationships in Kirthar National Park, Sindh, Pakistan. J. Arid Environ. 2005, 61, 397–418. [Google Scholar] [CrossRef]
- Ward, D. The Biology of Deserts; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- San Martín, C.; Alvarez, M. Floristic composition of anthropogenic seasonal wetlands in the coastal mountain range of Cautin, Chile. Agrofor. Surv. 2009, 37, 9–25. [Google Scholar] [CrossRef]
- Kotańska, M.; Buziak-Chmielowiec, E.; Dąbrowska, A.; Gladysz, M.; Jakielaszek, A.; Wójcik, T. Human impact on the plant cover of four villages in SE Poland. Steciana 2015, 19, 115–121. [Google Scholar] [CrossRef]
- Abd El-Wahab, R.H. Plant assemblage and diversity variation with human disturbances in coastal habitats of the western Arabian Gulf. J. Arid Land 2016, 8, 787–798. [Google Scholar] [CrossRef] [Green Version]
- Neji, M.; Serbaji, M.M.; Hardy, O.; Chaieb, M. Floristic diversity and vegetation patterns along disturbance gradient in arid coasts in southern mediterranean: Case of the Gulf of Gabès, southern Tunisia. Arid Land Res. Manag. 2018, 32, 291–315. [Google Scholar] [CrossRef]
- Dhaou, S.O.; Abdallah, F.; Belgacem, A.O.; Chaieb, M. The protection effects on floristic diversity in North African pseudo-savanna. Pak. J. Bot. 2010, 42, 1501–1510. [Google Scholar]
- El-Sheikh, M.A.; Thomas, J.; Alatar, A.A.; Hegazy, A.K.; Abbady, G.A.; Alfarhan, A.H.; Okla, M.I. Vegetation of Thumamah Nature Park: A managed arid land site in Saudi Arabia. Rend. Fis. Acc. Lincei 2013, 24, 349–367. [Google Scholar] [CrossRef]
- Ayyad, M. Case studies in the conservation of biodiversity: Degradation and threats. J. Arid Environ. 2003, 54, 165–185. [Google Scholar] [CrossRef] [Green Version]
- EEAA Egyptian Environmental Affairs Agency: Egypt State of the Environment Report 2006. Available online: http://www.eeaa.gov.eg/en-us/mediacenter/reports/soereports/soe2006.aspx (accessed on 1 December 2007).
- Abd El-Ghani, M.M.; El-Sawaf, N. The coastal roadside vegetation and environmental gradients in the arid lands of Egypt. Community Ecol. 2005, 6, 143–154. [Google Scholar] [CrossRef] [Green Version]
- Gelbard, J.l.; Belnap, J. Roads as conduits for exotic plant invasions in a semiarid landscape. Conserv. Biol. 2003, 17, 420–432. [Google Scholar] [CrossRef]
- Sullivan, J.J.; Williams, P.A.; Timmins, S.M.; Mark, C.; Smale, M.C. Distribution and spread of environmental weeds along New Zealand roadsides. New Zeal. J. Ecol. 2009, 33, 190–204. [Google Scholar]
- Abd El-Ghani, M.M.; Salama, F.M.; El-Tayeh, N.A. Desert roadside vegetation in eastern Egypt and environmental determinants for its distribution. Phytol. Balcan. 2013, 19, 233–242. [Google Scholar]
- Solbrig, O.T. The origin and function of biodiversity. Environ. Sci. Policy Sustain. Dev. 1991, 33, 16–38. [Google Scholar] [CrossRef]
- Kassas, M.; Batanouny, K.H. Plant ecology in Sahara Desert. In Sahara Desert; Cloudsley-Thompson, J., Ed.; Pergamon Press: Oxford, UK, 1984; pp. 77–90. [Google Scholar]
- Abd El-Ghani, M.M.; Huerta-Martínez, F.M.; Hongyan, L.; Qureshi, R. Plant Responses to Hyperarid Desert Environments; Springer International Pubisher: Berlin/Heidelberg, Germany, 2017; ISSN 978-3-319-59134-6. [Google Scholar]
- Zahran, M.A.; Willis, A.J. The Vegetation of Egypt; Chapman and Hall: London, UK, 1992. [Google Scholar]
- Galal, T.M.; Fahmy, A.G. Plant diversity and community structure of Wadi Gimal protected area, Red Sea Coast of Egypt. Afr. J. Ecol. 2012, 50, 266–276. [Google Scholar] [CrossRef]
- Alatar, A.; El-Sheikh, M.A.; Thomas, J. Vegetation analysis of Wadi Al-Jufair, a hyper-arid region in Najd, Saudi Arabia. Saudi J. Biol. Sci. 2012, 19, 357–368. [Google Scholar] [CrossRef] [Green Version]
- Kadmon, R.; Danin, A. Distribution of plant species in Israel in relation to spatial variation in rainfall. J. Veg. Sci. 1999, 10, 421–432. [Google Scholar] [CrossRef]
- Li, B.; Wang, L.; Kaseke, K.F.; Vogt, R.; Li, L.; Seely, M.K. The impact of fog on soil moisture dynamics in the Namib Desert. Adv. Water Resour. 2018, 113, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Arshad, M.; Ul-Hussan, A.; Ashraf, M.Y.; Noureen, S.; Moazzam, M. Edaphic factors and distribution of vegetation in the cholistan desert, pakistan. Pak. J. Bot. 2008, 40, 1923–1931. [Google Scholar]
- Shupe, S.M. Multivariate characterization of Sonoran Desert vegetation in southwest Arizona using US Army field data. Plant Ecol. 2005, 176, 215–235. [Google Scholar] [CrossRef]
- Abdi, M.; Afsharzadeh, S. An analysis of vegetation and species diversity patterns in sand dune and gravel desert ecosystem. Bot. Sci. 2016, 94, 499–511. [Google Scholar] [CrossRef] [Green Version]
- Kassas, M.; Imam, M. Habitat and plant communities in the Egyptian desert, III. The Wadi Bed Ecosystem, J. Ecol. 1954, 42, 424–441. [Google Scholar]
- Hassib, M. Distribution of plant communities in Egypt. Bull. Fac. Sci. Univ. Fouad 1 Cairo Egypt 1951, 29, 59–261. [Google Scholar]
- Abdelaal, M. Current status of the floristic composition in Wadi Hagul, Northwest Suez Gulf, Egypt. Rend. Fis. Acc. Lincei 2017, 28, 81–92. [Google Scholar] [CrossRef]
- Hassan, M.O.; Hassan, Y.M. Effect of human activities onfloristic composition and diversity of desert and urban vegetation in a new urbanized desert ecosystem. Heliyon 2019, 5, e02283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, S.R.; Mallik, A.U. Disturbance effects on species diversity and functional diversity in riparian and upland plant communities. Ecology 2010, 91, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Kassas, M.; Imam, M. Habitat and plant communities in the Egyptian Desert. J. Ecol. 1959, 47, 289–310. [Google Scholar] [CrossRef]
- Kassas, M.; El-Abyad, M.S. On the phytosociology of the desert vegetation of Egypt. Ann. Arid Zone 1962, 1, 54–83. [Google Scholar]
- Zahran, M.A.; Willis, A.J. The Vegetation of Egypt, 2nd ed.; Springer Science+Business Media: London, UK, 2009. [Google Scholar]
- Hassan, L.M. Plant life in the Digla conserved area, hyperarid desert, Egypt. Online J. Biol. Sci. 2002, 2, 533–537. [Google Scholar]
- Said, R. The Geology of Egypt; Pergamon Press: Oxford, UK; New York, NY, USA; Seoul, Korea; Tokyo, Japan, 1990; p. 593. [Google Scholar]
- Eid, R.A.H. Approaching Industrial and Environmental Reform for Shaq Al-Thu’ban Marble and Granite Industrial Cluster; The American University in Cairo: Cairo, Egypt, 2011; 158p. [Google Scholar]
- Emberger, L. Rapport sur les régions arides et semi- arides de l’Afrique du Nord; Union Int. Soc. Biologiques, Série B, Colloques: Paris, France, 1951; Volume 9, pp. 50–61. [Google Scholar]
- El-Sheikh, M.A. Weed vegetation ecology of arable land in Salalah, Southern Oman. Saudi J. Biol. Sci. 2013, 20, 291–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulos, L. Flora of Egypt; Checklist; Al Hadara Publishing: Cairo, Egypt, 1995. [Google Scholar]
- Boulos, L. Flora of Egypt; Vol. 1 (Azollaceae—Oxalidaceae); Al-Hadara Publishing: Cairo, Egypt, 1999. [Google Scholar]
- Boulos, L. Flora of Egypt; Vol. 2 (Geraniaceae—Boraginaceae); Al-Hadara Publishing: Cairo, Egypt, 2000. [Google Scholar]
- Boulos, L. Flora of Egypt; Vol. 3 (Verbinaceae—Compositae); Al-Hadara Publishing: Cairo, Egypt, 2002. [Google Scholar]
- Boulos, L. Flora of Egypt; Vol. 4 (Monocotyledons: Alismataceae—Orchidaceae); AL Hadara Publishing: Cairo, Egypt, 2005. [Google Scholar]
- Boulos, L. Flora of Egypt; Checklist Revised. Annotated Edition; Al Hadara Publishing: Cairo, Egypt, 2009; pp. 29–30. [Google Scholar]
- Raunkiaer, C. The Plant Life Forms and Statistical Plant Geography; Clarendon Press: Oxford, UK, 1934. [Google Scholar]
- Pansu, M.; Gautheyrou, J. Handbook of Soil Analysis: Mineralogical, Organic and Inorganic Methods; Springer: Berlin/Heidelberg, Germany, 2007; ISBN1 10 3540312102. ISBN2 13 9783540312109. [Google Scholar]
- Davey, B.G.; Bembrick, M.J. The potentiometric estimation of chloride in water extracts of soils. Soil Sci. Soc. Amer. Proc. 1969, 33, 385–387. [Google Scholar] [CrossRef]
- Allen, S.E.; Grimshaw, H.M.; Parkinson, J.A.; Quarmby, C. Chemical Analysis of Ecological Materials; Blackwell Scientific Publication: Oxoford, UK, 1974; 565p. [Google Scholar]
- Upadhyay, R.M.; Sharma, N.L. Manual of Soil, Plant, Water and Fertilizer Analysis; Kalyani Publishers: New Delhi, India, 2005. [Google Scholar]
- McCune, B.; Mefford, M.J. PC-ORD for Windows. Multivariate Analysis of Ecological Data; Version 4.14. User’s Guide; MjM Software: Oregon, USA, 1999. [Google Scholar]
- Orlóci, L. Multivariate Analysis in Vegetation Research, 2nd ed.; W. Junk, B.V. Publishers: The Hague, The Netherlands; Boston, MA, USA, 1978. [Google Scholar]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F. CANOCO, version 4.52; Wageningen University and Research Centre: Wageningen, The Netherlands, 2003. [Google Scholar]
- SPSS. SPSS Interactive Graphics 10.0, A Comprehensive System for Analyzing Data; SPSS Incorporation: Chicago, IL, USA, 1999. [Google Scholar]
- Hill, M.O.; Gauch, H.G. Detrended Correspondence Analysis: An improved ordination technique. Vegetatio 1980, 42, 47–58. [Google Scholar] [CrossRef]
- Ter Braak, C.F.J.; Verdonschot, P.F.M. Canonical correspondence analysis and the related multivariate methods in aquatic ecology. Aquat. Sci. 1995, 57, 254–289. [Google Scholar] [CrossRef]
- Rao, C.R. The use and interpretation of principal component analysis in applied research. Sankhya A 1964, 26, 329–350. [Google Scholar]
- Jongman, R.H.G.; ter Braak, C.J.F.; van Tregen, O.F.R. Data Analysis in Community and Landscape Ecology; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Zar, J.H. Biostatistical Analysis, 4th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 1999. [Google Scholar]
- Ter Braak, C.J.F.; Šmilauer, P. CANOCO Reference Manual and Cano Draw for Windows User’s Guide: Software for Canonical Community Ordination, version 4.5; Microcomputer power: Ithaca, NY, USA, 2002; 500p. [Google Scholar]
- Ter Braak, C.J.F. Canonical community ordination. Part 1. Basic theory and linear methods. Ecoscience 1994, 1, 127–140. [Google Scholar] [CrossRef]
- Borcard, D.; Legendre, P.; Drapeau, P. Partialling out the spatial component of ecological variation. Ecology 1992, 73, 1045–1055. [Google Scholar] [CrossRef] [Green Version]
- Pielou, E.C. Ecological Diversity; Wiley: New York, NY, USA, 1975; p. 165. [Google Scholar]
- Magurran, A.E. Ecological Diversity and Its Measurement; Chapman and Hall: London, UK, 1988; 179p. [Google Scholar]
- Mahdavi, P.; Akhani, H.; Van der Maarel, E. Species diversity and life-form patterns in steppe vegetation along a 3000 m altitudinal gradient in the Alborz Mountains, Iran. Folia Geobot. 2013, 48, 7–22. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.Y.H.; Taylor, A.R.; Ostertag, R. Positive species diversity and above-ground biomass relationships are ubiquitous across forest strata despite interference from overstorey trees. Funct. Ecol. 2017, 31, 419–426. [Google Scholar] [CrossRef]
- Whittaker, R.H. Evolution of species diversity in land communities. Evol. Biol. 1977, 10, 1–67. [Google Scholar]
- Abd El-Ghani, M.M.; Salama, F.M.; Salem, B.; El-Hadidy, A.; Abdel-Aleem, M. Biogeographical relations of a hyperarid desert flora in eastern Egypt. Afr. J. Ecol. 2014, 52, 173–191. [Google Scholar] [CrossRef]
- Hassan, L.M. Studies on the Flora of Eastern Desert, Egypt. PhD Thesis, Faculty of Science, Cairo University, Giza, Egypt, 1987. [Google Scholar]
- Tadros, T.M. Osmotic pressure of Egyptian desert plants in relation to their water supply. Bull. Fac. Sci. Cairo Univ. 1936, 7, 1–35. [Google Scholar]
- El-Adawy, H.A. Variation of Population Dynamics and Phenology of Desert Plant Communities. PhD Thesis, Cairo University, Giza, Egypt, 2011. [Google Scholar]
- Ter Braak, C.J.F. Canonical correspondence analysis: A new eigenvector technique for multivariate direct gradient analysis. Ecology 1986, 67, 1167–1179. [Google Scholar] [CrossRef] [Green Version]
- El-Ghanem, W.A.; Hassan, L.M.; Galal, T.M.; Badr, A. Floristic composition and vegetation analysis in Hail region north of central Saudi Arabia. Saudi J. Biol. Sci. 2010, 17, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Shaheen, H.; Qureshi, R.; Akram, A.; Gulfraz, M.; Potter, D. A preliminary floristic checklist of Thal desert Punjab, Pakistan. Pak. J. Bot. 2014, 46, 13–18. [Google Scholar]
- Salama, F.M.; Abd El-Ghani, M.M.; Amro, A.A.E.R.; Gaafar, A.E.S.; Abd El Galil, A.A.E.M. Vegetation dynamics and species diversity in a Saharan Oasis. Egypt. Not. Scient. Biol. 2018, 10, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Croce, A.; Stinca, A.; Santangelo, A.; Esposito, A. Exploring vascular flora diversity of two protected sandy coastal areas in southern Italy. Rend. Fis. Acc. Lincei 2019, 30, 323–336. [Google Scholar] [CrossRef]
- McIntyre, S.; Lavorel, S.; Trémont, R.M. Plant life-history attributes: Their relationship to disturbance response in herbaceous vegetation. J. Ecol. 1995, 83, 31–44. [Google Scholar] [CrossRef]
- Sternberg, M.; Shoshany, M. Aboveground biomass allocation and water content relationships in Mediterranean trees and shrubs at two climatological regions in Israel. Plant Ecol. 2001, 157, 171–179. [Google Scholar] [CrossRef]
- Hosni, H.A.; Hegazy, A.K. Contribution to the flora of Asir, Saudi Arabia. Candollea 1996, 51, 169–202. [Google Scholar]
- Shaltout, K.H.; Sheded, M.G.; Salem, A.M. Vegetation spatial heterogeneity in a hyper arid biosphere reserve area in North Africa. Acta Bot. Croat. 2010, 69, 31–46. [Google Scholar]
- Abd El-Ghani, M.M.; Bornkamm, R.; El-Sawaf, N.; Turky, H. Heterogeneity of Soil and Vegetation in the Urban Habitats of New Industrial Cities in the Desert Landscape of Egypt. Not. Scient. Biol. 2015, 7, 26–36. [Google Scholar] [CrossRef]
- Sans, F.X.; Masalles, R.M. Phenological patterns in an arable land weed community related to disturbance. Weed Res. 1995, 35, 321–332. [Google Scholar] [CrossRef]
- Abd El-Ghani, M.M. Floristics and environmental relations in two extreme desert zones of western Egypt. Global Ecol. Biogeogr. 2000, 9, 499–516. [Google Scholar] [CrossRef]
- Abd El-Ghani, M.M. Environmental correlates of species distribution in arid desert ecosystems of eastern Egypt. J. Arid Environ. 1998, 38, 297–313. [Google Scholar] [CrossRef]
- Salama, F.M.; Abd El-Ghani, M.M.; Gadallah, M.; El-Naggar, S.M. Amro, Diversity and responses of plant functional groups to soil variables in the arid desert landscape of southern Egypt. J. Biodivers. Ecol. Sci. 2016, 5, 24–38. [Google Scholar]
- Abdel Khalik, K.; Al-Gohary, I.; Al-Sodany, Y. Floristic composition and vegetation: Environmental relationships of Wadi Fatimah, Mecca, Saudi Arabia. Arid Land Res. Manag. 2017, 31, 316–334. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, J.; Wang, X.; Pann, H.; Gu, Z.; Ban, B. The distribution pattern of biological soil crust in the Gurbantunggut Desert. Acta Geogr. Sin. 2005, 60, 53–60. [Google Scholar]
- Medeiros, A.S.; Drezner, T.D. Vegetation, climate, and soil relationships across Sonoran Desert. Ecoscience 2012, 19, 148–160. [Google Scholar] [CrossRef]
- Feng, Y.; Jia-Qiang, L.; Xing-Wen, X.; Bo-Rong, P. Composition and characteristics of Libyan flora. Arch. Biol. Sci. 2013, 65, 651–657. [Google Scholar]
- Baameur, M.; Abdelguerfi, A.; Bouhoun, D.M.; Saadi, H.; El Hadj, O.M. Distribution study of some species of spontaneous Flora in two Saharan Regions of the North-East of Algeria (Ouargla and Ghardaïa). Int. J. Biodivers. Conserv. 2015, 7, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Salama, F.M.; Abd El-Ghani, M.M.; El-Naggar, S.M.; Baayo, K.A. Floristic composition and chorological analysis of the Sallum area, west Mediterranean, Egypt. J. Union Arab Biol. Cairo 2003, 13, 27–47. [Google Scholar]
- Abd El-Ghani, M.M.; Abdel-Khalik, K.N. Floristic diversity and phytogeography of Gebel Elba National Park, south-east Egypt. Turk. J. Bot. 2006, 30, 121–136. [Google Scholar]
- Zohary, M. Geobotanical Foundation of the Middle East; Gus. Fischer-Verlag: Stuttgart, Germany, 1983. [Google Scholar]
- White, F.; Leonard, J. Phytogeographical links between Africa and southwest Asia. Flora Veg. Mundi 1991, 9, 229–246. [Google Scholar]
- Kürschner, H.; Neef, R. A first synthesis of the flora and vegetation of the Tayma oasis and surroundings (Saudi Arabia). Plant Divers. Evol. 2011, 129, 27–58. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology, 2nd ed.; Elsevier B.V.: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Pillar, V.D. How sharp are classifications? Ecology 1999, 80, 2508–2516. [Google Scholar] [CrossRef]
- McCune, B.; Grace, J.B.; Urban, D.L. Analysis of Ecological Communities; MjM Software Design: Gleneden Beach, OR, USA, 2002; 300p. [Google Scholar]
- Fossati, J.; Pautou, G.; Peltier, J.P. Wadi vegetation of the North-Eastern desert of Egypt. Feddes Repert. 1998, 109, 313–327. [Google Scholar] [CrossRef]
- Salama, F.; Abd El-Ghani, M.M.; El-Tayeh, N.; Amro, A.; Gaafar, A.; Abd El Galil, A. Assessing the role of environmental gradients on the phytodiversity in Kharga Oasis of Western Desert, Egypt. Jordan J. Biol. Sci. 2019, 12, 421–434. [Google Scholar]
- Salama, F.M.; Abd El-Ghani, M.M.; El Naggar, S.M.; Baayo, K.A. Vegetation structure and environmental gradients in the Sallum area, Egypt. Ecol. Medit. 2005, 31, 15–32. [Google Scholar] [CrossRef]
- Abd El-Ghani, M.M.; Amer, W.M. Soil-vegetation relationships in a coastal desert plain of southern Sinai, Egypt. J. Arid Environ. 2003, 55, 607–628. [Google Scholar] [CrossRef]
- Salama, F.M.; Abd El-Ghani, M.M.; El-Tayeh, N. Vegetation and soil relationships in the inland wadi ecosystem of central Eastern Desert Egypt. Turk. J. Bot. 2013, 37, 489–498. [Google Scholar]
- Tielbörger, K. The vegetation of linear desert dunes in the north-western Negev, Israel. Flora 1997, 192, 261–278. [Google Scholar] [CrossRef]
- Noy-Meir, I. Desert ecosystems: Environment and producers. Annu. Rev. Ecol. Syst. 1973, 4, 25–51. [Google Scholar] [CrossRef]
- Epstein, H.E.; Lauenroth, W.K.; Burke, I.C.; Coffin, D.P. Ecological responses of dominant grasses along two climatic gradients in the great plains of the United States. J. Veg. Sci. 1996, 7, 777–788. [Google Scholar] [CrossRef]
- Duckworth, J.C.; Bunce, R.G.H.; Malloch, A.J.C. Vegetation environment relationships in Atlantic European calcareous grasslands. J. Veg. Sci. 2000, 11, 15–22. [Google Scholar] [CrossRef]
- Amezaga, I.; Mendarte, S.; Albizu, I.; Besga, G.; Garbisu, C.; Onaindia, M. Grazing intensity, aspect, and slope effects on lime-stone grassland structure. Rangeland Ecol. Manag. 2004, 57, 606. [Google Scholar] [CrossRef]
- Zheng, Y.; Xie, Z.; Jiang, L.; Shimizu, H.; Rimmington, G.; Zhou, G. Vegetation responses along environmental gradients on the Ordos plateau, China. Ecol. Res. 2006, 21, 396–404. [Google Scholar] [CrossRef]
- Arekhi, S.; Heydari, M.; Pourbabaei, H. Vegetation-environmental relationships and ecological species groups of the Ilam Oak forest landscape, Iran. Caspian J. Env. Sci. 2010, 8, 115–125. [Google Scholar]
- Kargar-Chigani, H.; Javadi, S.A.; Zahedi-Amiri, G.; Khajeddin, J.; Jafari, M. Vegetation composition differentiation and species-environment relationships in the northern part of Isfahan Province, Iran. J. Arid Land 2017, 9, 161–175. [Google Scholar] [CrossRef] [Green Version]
- Abrams, P.A. Monotonic or unimodal diversity-productivity gradients: What does competition theory predict? Ecology 1995, 76, 2019–2027. [Google Scholar] [CrossRef]
- Yair, A.; Sharon, D.; Lavee, H. Trends in runoff and erosion processes over an arid limestone hillside, North Negev, Israel. Hydrolog. Sci. Bull. 1980, 25, 243–255. [Google Scholar] [CrossRef] [Green Version]
- Mashaly, I.A. Vegetation-soil relationships in lake borollus protected area, Egypt. Am. Eurasian J. Agric. Environ. Sci. 2006, 1, 229–238. [Google Scholar]
- He, M.Z.; Zheng, J.G.; Li, X.R.; Qian, Y.L. Environmental factors affecting vegetation composition in the Alxa Plateau, China. J. Arid Environ. 2007, 69, 473–489. [Google Scholar] [CrossRef]
- Zuo, X.A.; Wang, S.K.; Zhao, X.Y.; Lian, J. Scale dependence of plant species richness and vegetation-environment relationship along a gradient of dune stabilization in Horqin Sandy Land, Northern China. J. Arid Land 2014, 6, 334–342. [Google Scholar] [CrossRef] [Green Version]
- Huerta-Martínez, F.M.; García-Moya, E.; Flores-Flores, J.L.; Pi-mienta-Barrios, E. Ordenación de las poblaciones silves-tres de pitayo y cardón en la Cuenca de Sayula, Jalisco. Boletínde la Soc. Botánica de México 1999, 64, 11–24. [Google Scholar]
- El-Khatib, A.A.; Abd El-Ghani, M.M.; El-Sayed, K.N. Soil-vegetation relationship and distribution of Convolvulus in Egypt. Feddes Repert. 1998, 109, 175–185. [Google Scholar] [CrossRef]
- Abd El-Ghani, M.M.; Bornkamm, R.; El-Sawaf, N.; Turky, H. Plant species distribution and spatial habitat heterogeneity in the landscape of urbanizing desert ecosystem of Egypt. Urban Ecosyst. 2011, 14, 585–616. [Google Scholar] [CrossRef]
- Hassan, M.O. Leaf litter of Bombax ceiba L. threatens plant cover and floristic diversity in a new urban ecosystem. Flora 2018, 242, 22–30. [Google Scholar] [CrossRef]
- Forman, R.T.; Alexander, L.E. Roads and their major ecological effects. Annu. Rev. Ecol. Syst. 1998, 29, 207–231. [Google Scholar] [CrossRef] [Green Version]
- Davis, M.A.; Grime, J.P.; Thompson, K. Fluctuating resources in plant communities: A general theory of invasibility. Ecology 2000, 88, 528–534. [Google Scholar] [CrossRef] [Green Version]
- Huerta-Martínez, F.M.; Vázquez-García, J.A.; García-Moya, E.; López-Mata, L.; Vaquera-Huerta, H. Vegetation ordination at the southern Chihuahuan Desert (San Luis Potosi, Mexico). Plant Ecol. 2004, 174, 79–87. [Google Scholar] [CrossRef]
- Zuo, X.A.; Zhao, X.Y.; Zhao, H.L.; Zhang, T.; Li, Y.; Wang, S.; Li, W.; Powers, R. Scale dependent effects of environmental factors on vegetation pattern and composition in Horqin Sandy Land, Northern China. Geoderma 2012, 173–174, 1–9. [Google Scholar] [CrossRef]
- Wang, J.H.; Zhou, S.W.; Ren, P.Z. Studies on the species diversity of plant communities and their ecotopes in the northeastern Ulan Buh Desert. J. Desert Res. 1996, 16, 258–265. [Google Scholar]
- Ali, E.A.; Kafkafi, U.; Yamaguchi, I.; Sugimoto, Y.; Inanga, S. Growth, transpiration, cytokinins and gibberellins, nutrient compositional changes in sesame exposed to low root-zone temperature under different ratios of nitrate: Ammonium supply. J. Plant Nutr. 2000, 23, 123–140. [Google Scholar] [CrossRef]
- Wilson, M.V.; Shmida, A. Measuring beta diversity with presence–absence data. J. Ecol. 1984, 72, 1055–1064. [Google Scholar] [CrossRef]
- Vázquez García, J.A.; Givnish, T.J. Altitudinal gradients in tropical forest composition, structure, and diversity in the Sierra de Manantlán. J. Ecol. 1998, 86, 999–1020. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (A) Cairo-Suez road (SR) | |||||
| Authors of Earlier Studies | Kassas and El-Abyad (1962) | Hassan (1987) | Abd El-Ghani et al. (2014) | Number of Species | |
| Kassas and El-Abyad (1962) | 102 | ||||
| Hassan (1987) | 53 | 235 | |||
| Abd El-Ghani et al. (2014) | 52 | 53 | 113 | ||
| Current study (2017) | 39 | 27 | 42 | 47 | |
| (B) Wadi Degla protected area (WD) | |||||
| Authors of Earlier Studies | Tadros (1936) | Imam and Kassas (1954) | Hassan (2002) | El-Adawy (2011) | Number of Species |
| Tadros (1936) | 17 | ||||
| Imam and Kassas (1954) | 37 | 37 | |||
| Hassan (2002) | 30 | 5 | 75 | ||
| El-Adawy (2011) | 27 | 44 | 77 | 88 | |
| Current study (2017) | 38 | 53 | 73 | 63 | 58 |
| Vegetation Group | I | II | III | IV | V | VI | VII | VIII | p-Value | Species Abbreviations |
|---|---|---|---|---|---|---|---|---|---|---|
| Number of Sample Plots | 3 | 3 | 4 | 4 | 3 | 2 | 4 | 2 | ||
| Group I (3 significant indicator species) | ||||||||||
| Calotropis procera (Aiton) W.T. Aiton | 67 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.040 | C pro |
| Phragmites australis (Cav.) Trin.ex Steud. | 67 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.040 | P aus |
| Pulicariaundulata (Lam.) DC. | 67 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.037 | P und |
| Group II (1 significant indicator species) | ||||||||||
| Haloxylon salicornicum (Moq.) Bunge | 27 | 67 | 2 | 27 | 0 | 0 | 0 | 7 | 0.022 | H sal |
| Group III (2 significant indicator species) | ||||||||||
| Ochradenus baccatus Delile | 0 | 0 | 55 | 17 | 17 | 7 | 17 | 27 | 0.014 | O bac |
| Zygophyllum coccineum L. | 0 | 0 | 71 | 22 | 12 | 0 | 17 | 22 | 0.041 | Z coc |
| Group IV (3 significant indicator species) | ||||||||||
| Acacia tortilis (Forssk.) Hayne subsp. raddiana (Savi) Brenan | 0 | 0 | 6 | 56 | 0 | 0 | 0 | 0 | 0.046 | A tor |
| Panicum turgidum Forssk. | 0 | 10 | 0 | 52 | 0 | 0 | 0 | 0 | 0.050 | P tur |
| Launaea spinosa (Forssk.) Sch. Bip. ex Kuntze | 0 | 0 | 0 | 75 | 0 | 0 | 0 | 0 | 0.025 | L spi |
| Group V (2 significant indicator species) | ||||||||||
| Artemisia judaica L. | 0 | 0 | 0 | 0 | 38 | 21 | 0 | 0 | 0.016 | A jud |
| Reaumuria hirtella Jaub. and Spach | 0 | 0 | 0 | 0 | 36 | 17 | 20 | 0 | 0.022 | R hir |
| Group VI (1 significant indicator species) | ||||||||||
| Heliotropium bacciferum Forssk. | 0 | 0 | 0 | 0 | 0 | 80 | 5 | 0 | 0.013 | H bac |
| Group VII (2 significant indicator species) | ||||||||||
| Lasiurus scindicus Henrard | 0 | 0 | 0 | 0 | 4 | 9 | 71 | 35 | 0.038 | L sci |
| Nitraria retusa (Forssk.) Asch. | 0 | 0 | 0 | 0 | 0 | 0 | 50 | 0 | 0.044 | N ret |
| Group VIII (2 significant indicator species) | ||||||||||
| Orobanche cernua Loefl. | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 80 | 0.021 | O cer |
| Fagonia bruguieri DC. | 0 | 0 | 0 | 0 | 6 | 14 | 0 | 55 | 0.049 | F bru |
| Total Mean | Vegetation Groups | F-Ratio | p-Value | ||||
|---|---|---|---|---|---|---|---|
| I | II | III | IV | ||||
| Total Number of Sample Plots | 3 | 3 | 4 | 4 | |||
| Soil Variables | |||||||
| Sand (%) | 93.1 ± 1.5 | 92.3 ± 2.1 | 94.0 ± 0.5 | 93.7 ± 0.9 | 92.4 ± 2.0 | 3.06 | 0.08 |
| Silt (%) | 1.5 ± 0.2 | 1.8 ± 0.3 | 1.3 ± 0.1 | 1.4 ± 0.1 | 1.6 ± 0.1 | 4.06 * | 0.04 |
| Clay (%) | 5.3 ± 1.5 | 5.8 ± 2.0 | 4.6 ± 0.4 | 4.9 ± 0.8 | 6.0 ± 2.1 | 2.65 | 0.10 |
| OM (%) | 0.4 ± 0.2 | 0.23 ± 0.1 | 0.7 ± 0.2 | 0.4 ± 0.1 | 0.3 ± 0.2 | 3.33 | 0.06 |
| pH | 7.6 ± 0.3 | 7.7 ± 0.1 | 8.0 ± 0.03 | 7.6 ± 0.1 | 7.4 ± 0.4 | 18.17 ** | 0.001 |
| EC (dS m−1) | 2.0 ± 0.9 | 1.7 ± 1.1 | 3.0 ± 0.2 | 2.03 ± 0.3 | 1.5 ± 0.9 | 2.12 | 0.16 |
| Na+ (meq L−1) | 8.7 ± 3.8 | 7.1 ± 4.12 | 13.6 ± 2.3 | 8.5 ± 1.0 | 6.31 ± 3.5 | 2.73 | 0.09 |
| K+ (meq L−1) | 1.4 ± 0.7 | 0.9 ± 1.0 | 1.8 ± 0.4 | 1.6 ± 0.4 | 1.2 ± 0.8 | 0.76 | 0.57 |
| Ca2+ (meq L−1) | 5.4 ± 2.2 | 4.8 ± 3.0 | 7.8 ± 0.3 | 5.4 ± 0.5 | 3.9 ± 2.3 | 1.84 | 0.20 |
| Mg2+ (meq L−1) | 4.3 ± 2.0 | 3.7 ± 2.7 | 6.4 ± 0.3 | 4.9 ± 0.9 | 3.1 ± 2.2 | 1.55 | 0.27 |
| Cl− (meq L−1) | 11.5 ± 5.2 | 8.4 ± 5.1 | 18.5 ± 2.6 | 11.7 ± 1.6 | 8.4 ± 4.7 | 3.46 * | 0.05 |
| HCO3−(meq L−1) | 5.6 ± 2.2 | 5.0 ± 3.0 | 7.7 ± 0.3 | 5.8 ± 0.7 | 4.1 ± 2.6 | 1.33 | 0.33 |
| CO32− (meq L−1) | 0.3 ± 0.4 | 0.2 ± 0.3 | 0.5 ± 0.5 | 0.3 ± 0.4 | 0.2 ± 0.3 | 0.54 | 0.71 |
| SO42− (meq L−1) | 2.6 ± 1.4 | 3.02 ± 2.6 | 3.5 ± 0.7 | 2.1 ± 0.3 | 2.0 ± 1.5 | 0.73 | 0.59 |
| Anthropogenic Factors | |||||||
| Over-collection (OC) | 1.93 ± 0.6 | 1.00 ± 0.0 (L) | 1.0 ± 0.5 (L) | 1.3 ± 0.6 (L) | 2.5 ± 0.6 (H) | 3.07 * | 0.03 |
| Over-grazing (OG) | 1.36 ± 0.7 | 1.00 ± 0.0 (L) | 1.0 ±0.0 (L) | 1.2 ± 0.5 (L) | 2.0 ± 1.2 (H) | 4.50 * | 0.01 |
| Introduced species (IS) | 1.21 ± 1.0 | 1.67 ± 1.2 (M) | 0.7 ±1.2 (L) | 1.0 ± 0.8 (L) | 1.7 ± 1.3 (M) | 2.21 * | 0.04 |
| Land degradation (LD) | 1.50 ± 0.9 | 1.00 ± 0.0 (L) | 1.7 ± 1.2 (M) | 1.2 ± 0.5 (L) | 0.7 ± 1.0 (L) | 1.26 | 0.17 |
| Urbanization (UR) | 1.43 ± 1.1 | 1.50 ± 1.2 (M) | 1.3 ± 1.2 (L) | 1.2 ± 1.5 (L) | 1.7 ± 1.0 (M) | 6.10 ** | 0.009 |
| Solid wastes (SW) | 1.36 ± 1.1 | 1.00 ± 1.0 (L) | 2.3 ± 1.2 (H) | 0.7 ± 1.0 (L) | 0.7 ± 1.0 (L) | 1.59 | 0.40 |
| Military activities (MA) | 1.29 ± 1.1 | 0.33 ± 0.6 (L) | 1.2 ± 1.5 (L) | 2.5 ± 0.6 (H) | 1.0 ± 1.3 (L) | 1.76 | 0.22 |
| Diversity Indices | |||||||
| Species richness (α-diversity) | 6.9 ± 4.5 | 9.7 ± 2.5 | 1.7 ± 1.2 | 4.0 ± 1.4 | 11.7 ± 1.3 | 29.76 ** | 0.001 |
| Shannon–Wiener index (H’) | 1.6 ± 0.9 | 2.2 ± 0.3 | 0.4 ± 0.6 | 1.3 ± 0.3 | 2.4 ± 0.1 | 22.81 ** | 0.002 |
| Total Mean | Vegetation Groups | F-Ratio | p-Value | ||||
|---|---|---|---|---|---|---|---|
| V | VI | VII | VIII | ||||
| Total Number of Sample Plots | 3 | 2 | 4 | 2 | |||
| Soil Factors | |||||||
| Sand (%) | 94.8 ± 1.5 | 94.6 ± 2.3 | 94.7 ± 0.7 | 94.36 ± 1.5 | 96.35 ± 0.4 | 0.750 | 0.56 |
| Silt (%) | 2.03 ± 0.8 | 1.8 ± 0.4 | 1.5 ± 0.4 | 2.56 ± 1.2 | 1.84 ± 0.1 | 0.975 | 0.46 |
| Clay (%) | 3.1 ± 1.2 | 3.6 ± 2.1 | 3.8 ± 0.3 | 3.09 ± 0.6 | 1.81 ± 0.5 | 1.164 | 0.39 |
| OM (%) | 0.4 ± 0.2 | 0.5 ± 0.3 | 0.5 ± 0.2 | 0.27 ± 0.2 | 0.19 ± 0.03 | 1.111 | 0.41 |
| pH | 7.7 ± 0.1 | 7.8 ± 0.1 | 8.0 ± 0.1 | 7.89 ± 0.03 | 7.84 ± 0.02 | 2.002 | 0.20 |
| EC (dS m−1) | 2.9 ± 1.2 | 3.0 ± 0.7 | 3.0 ± 1.9 | 2.36 ± 0.7 | 3.94 ± 2.1 | 0.745 | 0.56 |
| Na+ (meq L−1) | 6.3 ± 6.4 | 4.9 ± 2.1 | 9.3 ± 1.1 | 4.31 ± 4.4 | 9.68 ± 1.2 | 0.422 | 0.74 |
| K+ (meq L−1) | 1.2 ± 0.4 | 1.2 ± 0.5 | 1.3 ± 0.8 | 1.12 ± 0.2 | 1.52 ± 0.5 | 0.331 | 0.80 |
| Ca2+ (meq L−1) | 16.7 ± 6.1 | 19.3 ± 6.6 | 13.2 ± 4.0 | 13.75 ± 2.1 | 22.50 ± 10.6 | 1.497 | 0.048 * |
| Mg2+ (meq L−1) | 5.0 ± 2.0 | 4.8 ± 2.4 | 6.1 ± 2.6 | 4.40 ± 1.9 | 5.65 ± 2.3 | 0.336 | 0.80 |
| Cl− (meq L−1) | 11.6 ± 7.5 | 12.7 ± 5.5 | 10.5 ± 9.2 | 8.25 ± 3.6 | 18.00 ± 15.6 | 0.727 | 0.036 * |
| HCO3− (meq L−1) | 0.5 ± 0.1 | 0.5 ± 0.1 | 0.4 ± 0.0 | 0.50 ± 0.1 | 0.50 ± 0.1 | 0.604 | 0.63 |
| SO42− (meq L−1) | 17.3 ± 5.1 | 16.9 ± 3.3 | 19.1 ± 9.5 | 14.83 ± 4.5 | 20.85 ± 5.4 | 0.654 | 0.61 |
| Diversity Indices | |||||||
| Species richness α-diversity) | 28.8 ± 2.4 | 26.3 ± 1.5 | 31.0 ± 1.4 | 28.7 ± 2.1 | 30.5 ± 2.1 | 3.29 | 0.088 |
| Shannon–Wiener index (H’) | 3.4 ± 0.1 | 3.3 ± 0.1 | 3.4 ± 0.04 | 3.4 ± 0.1 | 3.4 ± 0.1 | 3.37 | 0.084 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussein, E.A.; Abd El-Ghani, M.M.; Hamdy, R.S.; Shalabi, L.F. Do Anthropogenic Activities Affect Floristic Diversity and Vegetation Structure More Than Natural Soil Properties in Hyper-Arid Desert Environments? Diversity 2021, 13, 157. https://doi.org/10.3390/d13040157
Hussein EA, Abd El-Ghani MM, Hamdy RS, Shalabi LF. Do Anthropogenic Activities Affect Floristic Diversity and Vegetation Structure More Than Natural Soil Properties in Hyper-Arid Desert Environments? Diversity. 2021; 13(4):157. https://doi.org/10.3390/d13040157
Chicago/Turabian StyleHussein, Ethar A., Monier M. Abd El-Ghani, Rim S. Hamdy, and Lamiaa F. Shalabi. 2021. "Do Anthropogenic Activities Affect Floristic Diversity and Vegetation Structure More Than Natural Soil Properties in Hyper-Arid Desert Environments?" Diversity 13, no. 4: 157. https://doi.org/10.3390/d13040157
APA StyleHussein, E. A., Abd El-Ghani, M. M., Hamdy, R. S., & Shalabi, L. F. (2021). Do Anthropogenic Activities Affect Floristic Diversity and Vegetation Structure More Than Natural Soil Properties in Hyper-Arid Desert Environments? Diversity, 13(4), 157. https://doi.org/10.3390/d13040157