
Application of the Natural Products NOZEMAT HERB and NOZEMAT HERB PLUS Can Decrease Honey Bee Colonies Losses during the Winter




Abstract
:1. Introduction
1.1. Queen Bee Management
1.2. Beekeeping Management Practices
1.3. Forage Sources as a Risk Factor for Honey Bee Winter Losses
1.4. Role of Pathogens on Honey Bee Winter Losses
1.5. Effects from the Application of Herbal Supplements on Biochemical and Immunological Parameters in Honey Bee Haemolymph
2. Materials and Methods
2.1. Experimental Design
2.2. Evaluation of Food Consumption and Bee Mortality
- (1)
- The amount of food consumption from the bee colony during the winter—this parameter was evaluated based on the difference between the amount of honey during wintering and the amount of honey in the first clinical examination of honey bee colonies, following standard norms (Conditions and procedure for performance of selection and reproduction activities in beekeeping ORDINANCE No. 35, 2004, issued by The Bulgarian Ministry of Agriculture, Food and Forestry, https://www.mzh.government.bg/en/, accessed on 15 May 2021) (Table 1). The amount of the consumed food was added to the amount of the honey-sugar paste which bees used in the winter. Food consumption per unit of bees was evaluated according to the following formula:where M denoted the consumed food per unit of bees (frames, number, kg) in winter; C—the total amount of consumed food (kg); A—the strength of the bee colony in the autumn (frames, number, kg); and B—the strength of the bee colony in the spring (frames, number, kg).M = C/0.5 × (A + B)
- (2)
- Bee mortality (winter colony losses %)—the difference between the strength of the bee colony in the autumn and in the spring (Conditions and procedure for performance of selection and reproduction activities in beekeeping ORDINANCE No. 35, 2004, issued by The Bulgarian Ministry of Agriculture, Food and Forestry (https://www.mzh.government.bg/en/, accessed on 15 May 2021) (Table 2). This parameter was evaluated according to the following formula:where P denoted the amount of dead bees (winter colony losses, %); A—the strength of the bee colony in the autumn; and B—the strength of the bee colony in the spring.P = (A − B)/A × 100
2.3. Evaluation of Total Protein and Lysozyme Content in the Bee Haemolymph
2.4. Statistical Analysis
3. Results
3.1. Effect of the Herbal Supplements NH and NHP on Overwitering Food Consumption and Bee Mortality
3.1.1. Overwintering Food Consumption
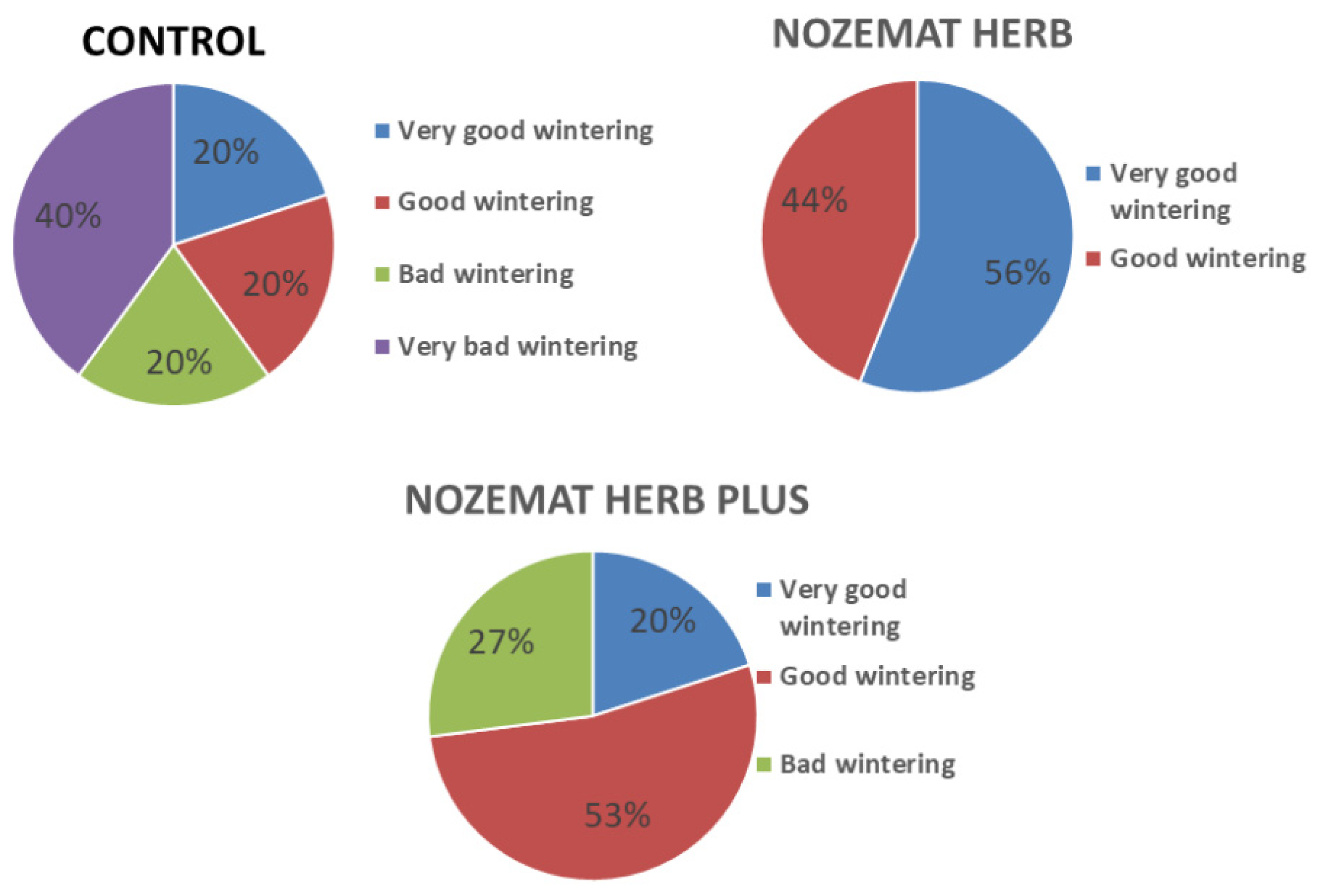
3.1.2. Overwintering Colony Losses
3.2. Effect of Herbal Supplements on Total Protein and Lysozyme Content in the Bee Haemolymph
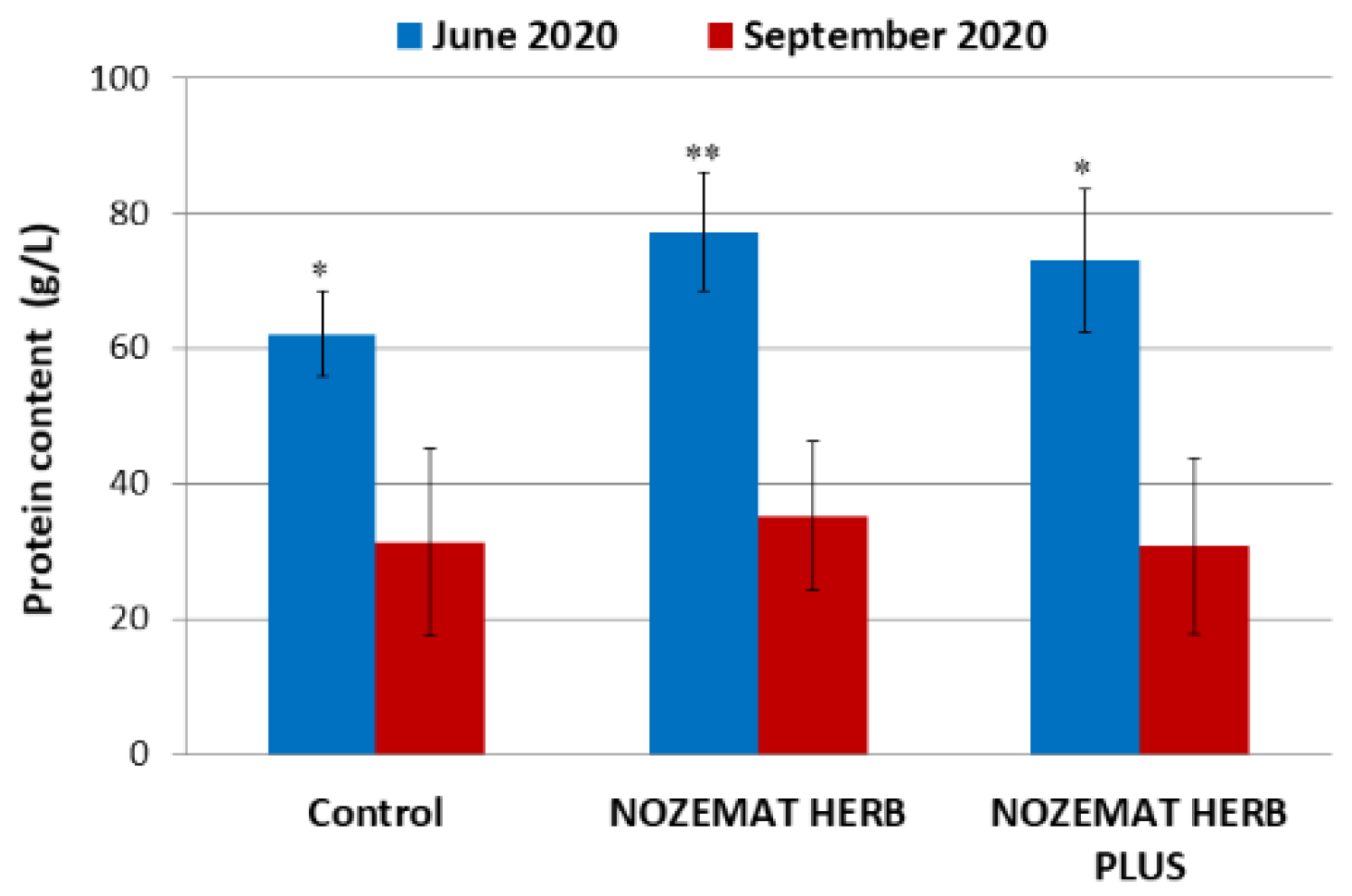
3.2.1. Influence of the Herbal Supplements NH and NHP on the Total Protein Content in the Bee Haemolymph
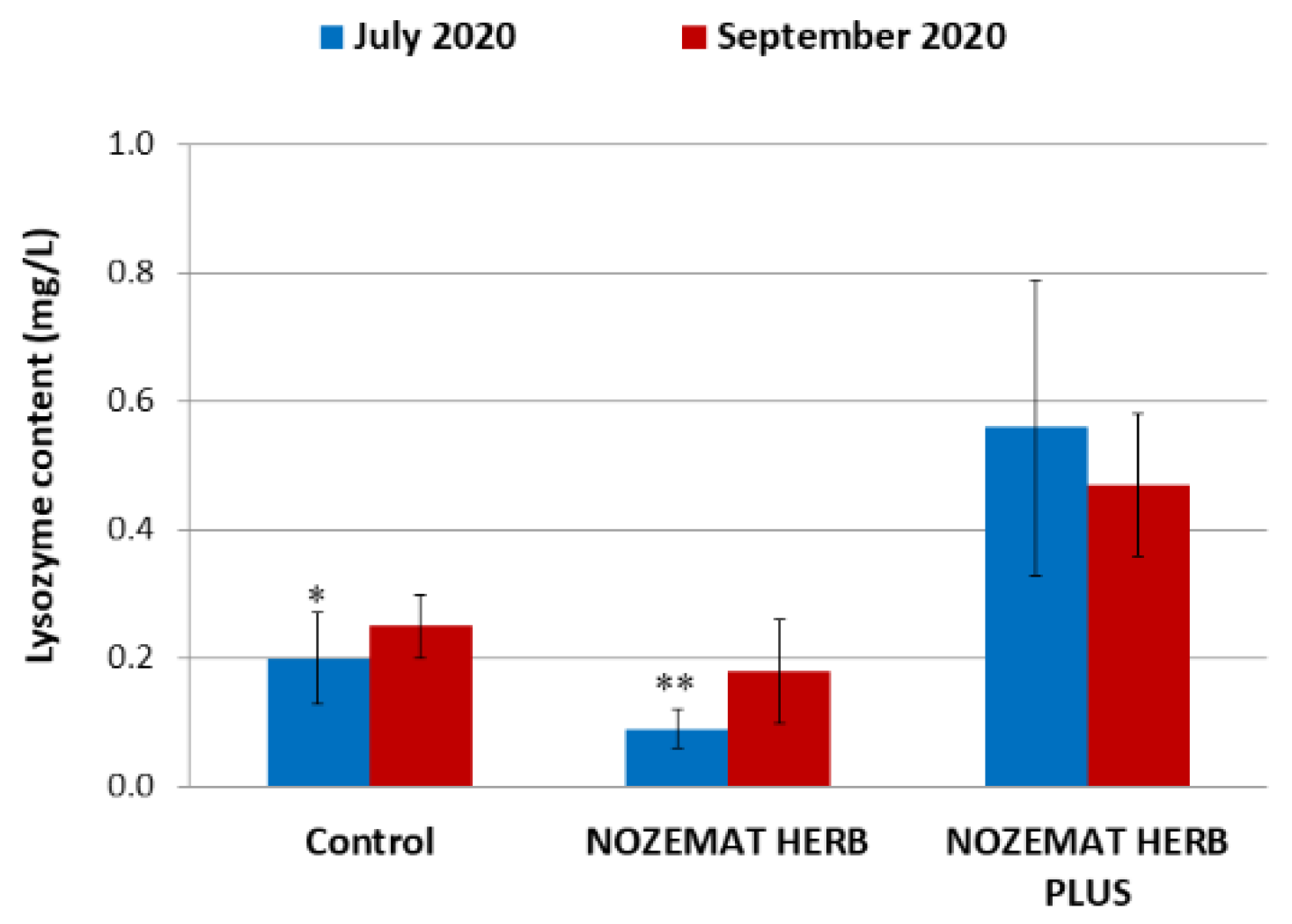
3.2.2. Influence of the Herbal Supplements NH and NHP on Lysozyme Content in the Bee Haemolymph
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ballantyne, G.; Baldock, K.C.R.; Rendell, L.; Willmer, P.G. Pollinator Importance Networks Illustrate the Crucial Value of Bees in a Highly Speciose Plant Community. Sci. Rep. 2017, 7, 8389. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.-Q.; Wang, K.; Marcucci, M.C.; Sawaya, A.C.H.F.; Hu, L.; Xue, X.-F.; Wu, L.-M.; Hu, F.-L. Nutrient-Rich Bee Pollen: A Treasure Trove of Active Natural Metabolites. J. Funct. Foods 2018, 49, 472–484. [Google Scholar] [CrossRef]
- El-Seedi, H.R.; Khalifa, S.A.M.; El-Wahed, A.A.; Gao, R.; Guo, Z.; Tahir, H.E.; Zhao, C.; Du, M.; Farag, M.A.; Musharraf, S.G.; et al. Honeybee Products: An Updated Review of Neurological Actions. Trends Food Sci. Technol. 2020, 101, 17–27. [Google Scholar] [CrossRef]
- Gray, A.; Adjlane, N.; Arab, A.; Ballis, A.; Brusbardis, V.; Charrière, J.D.; Chlebo, R.; Coffey, M.F.; Cornelissen, B.; da Costa, C.A.; et al. Honey bee colony winter loss rates for 35 countries participating in the COLOSS survey for winter 2018–2019, and the effects of a new queen on the risk of colony winter loss. J. Apic. Res. 2020, 59, 744–751. [Google Scholar] [CrossRef]
- Amiri, E.; Strand, M.K.; Rueppell, O.; Tarpy, D.R. Queen quality and the impact of honey bee diseases on queen health: Potential for interactions between two major threats to colony health. Insects 2017, 8, 48. [Google Scholar] [CrossRef]
- Steinhauer, N.; Kulhanek, K.; Antúnez, K.; Human, H.; Chantawannakul, P.; Chauzat, M.P.; vanEngelsdorp, D. Drivers of colony losses. Curr. Opin. Insect Sci. 2018, 26, 142–148. [Google Scholar] [CrossRef]
- vanEngelsdorp, D.; Tarpy, D.R.; Lengerich, E.J.; Pettis, J.S. Idiopathic brood disease syndrome and queen events as precursors of colony mortality in migratory beekeeping operations in the eastern United States. Prev. Vet. Med. 2013, 108, 225–233. [Google Scholar] [CrossRef] [Green Version]
- Giacobino, A.; Molineri, A.; Cagnolo, N.B.; Merke, J.; Orellano, E.; Bertozzi, E.; Masciangelo, G.; Pietronave, H.; Pacini, A.; Salto, C.; et al. Queen Replacement: The Key to Prevent Winter Colony Losses in Argentina. J. Apic. Res. 2016, 55, 335–341. [Google Scholar] [CrossRef]
- Zee, R.V.D.; Brodschneider, R.; Brusbardis, V.; Charriere, J.D.; Chlebo, R.; Coffey, M.F.; Dahle, B.; Drazic, M.M.; Kauko, L.; Kretavicius, J.; et al. Results of international standardised beekeeper surveys of colony losses for winter 2012–2013: Analysis of winter loss rates and mixed effects modelling of risk factors for winter loss. J. Apic. Res. 2014, 53, 19–34. [Google Scholar] [CrossRef] [Green Version]
- Oberreiter, H.; Brodschneider, R. Austrian COLOSS survey of honey bee colony winter losses 2018/19 and analysis of hive management practices. Diversity 2020, 12, 99. [Google Scholar] [CrossRef] [Green Version]
- Brown, P.; Newstrom-Lloyd, L.E.; Foster, B.J.; Badger, P.H.; McLean, J.A. Winter 2016 honey bee colony losses in New Zealand. J. Apic. Res. 2018, 57, 278–291. [Google Scholar] [CrossRef] [Green Version]
- Sperandio, G.; Simonetto, A.; Carnesecchi, E.; Costa, C.; Hatjina, F.; Tosi, S.; Gilioli, G. Beekeeping and Honey Bee Colony Health: A Review and Conceptualization of Beekeeping Management Practices Implemented in Europe. Sci. Total Environ. 2019, 696, 133795. [Google Scholar] [CrossRef]
- Steinhauer, N.; Saegerman, C. Prioritizing changes in management practices associated with reduced winter honey bee colony losses for US beekeepers. Sci. Total Environ. 2021, 753, 141629. [Google Scholar] [CrossRef] [PubMed]
- Meikle, W.G.; Weiss, M.; Maes, P.W.; Fitz, W.; Snyder, L.A.; Sheehan, T.; Mott, B.M.; Anderson, K.E. Internal Hive Temperature as a Means of Monitoring Honey Bee Colony Health in a Migratory Beekeeping Operation before and during winter. Apidologie 2017, 48, 666–680. [Google Scholar] [CrossRef] [Green Version]
- De Guzman, L.I.; Simone-Finstrom, M.; Frake, A.M.; Tokarz, P. Comb irradiation has limited, interactive effects on colony performance or pathogens in bees, Varroa destructor and wax based on two honey bee stocks. Insects 2019, 10, 15. [Google Scholar] [CrossRef] [Green Version]
- Shimshoni, J.A.; Sperling, R.; Massarwa, M.; Chen, Y.; Bommuraj, V.; Borisover, M.; Barel, S. Pesticide Distribution and Depletion Kinetic Determination in Honey and Beeswax: Model for Pesticide Occurrence and Distribution in Beehive Products. PLoS ONE 2019, 14, e0212631. [Google Scholar] [CrossRef]
- El Agrebi, N.; Traynor, K.; Wilmart, O.; Tosi, S.; Leinartz, L.; Danneels, E.; de Graaf, D.C.; Saegerman, C. Pesticide and Veterinary Drug Residues in Belgian Beeswax: Occurrence, Toxicity, and Risk to Honey Bees. Sci. Total Environ. 2020, 745, 141036. [Google Scholar] [CrossRef]
- Musinguzi, P.; Bosselmann, A.S.; Pouliot, M. Livelihoods-Conservation Initiatives: Evidence of Socio-Economic Impacts from Organic Honey Production in Mwingi, Eastern Kenya. For. Policy Econ. 2018, 97, 132–145. [Google Scholar] [CrossRef]
- Wintermantel, D.; Odoux, J.; Chadœuf, J.; Bretagnolle, V. Organic Farming Positively Affects Honeybee Colonies in a Flower-poor Period in Agricultural Landscapes. J. Appl. Ecol. 2019, 56, 1960–1969. [Google Scholar] [CrossRef] [Green Version]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 6229. [Google Scholar] [CrossRef]
- Durant, J.L. Where Have All the Flowers Gone? Honey Bee Declines and Exclusions from Floral Resources. J. Rural Stud. 2019, 65, 161–171. [Google Scholar] [CrossRef]
- Urbanowicz, C.; Baert, N.; Bluher, S.E.; Böröczky, K.; Ramos, M.; McArt, S.H. Low Maize Pollen Collection and Low Pesticide Risk to Honey Bees in Heterogeneous Agricultural Landscapes. Apidologie 2019, 50, 379–390. [Google Scholar] [CrossRef]
- Gray, A.; Brodschneider, R.; Adjlane, N.; Ballis, A.; Brusbardis, V.; Charrière, J.D.; Chlebo, R.; Coffey, M.F.; Cornelissen, B.; da Costa, C.A.; et al. Loss rates of honey bee colonies during winter 2017/18 in 36 countries participating in the COLOSS survey, including effects of forage sources. J. Apic. Res. 2019, 58, 479–485. [Google Scholar] [CrossRef] [Green Version]
- Schmolke, A.; Kearns, B.; O’Neill, B. Plant Guttation Water as a Potential Route for Pesticide Exposure in Honey Bees: A Review of Recent Literature. Apidologie 2018, 49, 637–646. [Google Scholar] [CrossRef] [Green Version]
- Pilling, E.; Campbell, P.; Coulson, M.; Ruddle, N.; Tornier, I. A Four-Year Field Program Investigating Long-Term Effects of Repeated Exposure of Honey Bee Colonies to Flowering Crops Treated with Thiamethoxam. PLoS ONE 2013, 8, e77193. [Google Scholar] [CrossRef] [PubMed]
- Charrière, J.-D.; Imdorf, A.; Koenig, C.; Gallmann, S.; Kuhn, R. Do Sunflowers Influence the Development of Honey Bee, Apis mellifera, Colonies in Areas with Diversified Crop Farming? J. Apic. Res. 2010, 49, 227–235. [Google Scholar] [CrossRef]
- Crailsheim, K.; Moosbeckhofer, R.; Brodschneider, R. Future of Honey Bees—Basic Research for Project for Honey Bee Health and Bee Protection. Final Report Poject ‘Zukunft Biene’ 2014–2018 Austria. 2018. Available online: https://www.ages.at/en/topics/environment/bees/research-projects-on-bees/future-of-honey-bees/ (accessed on 7 February 2020).
- Bernal, J.; Martin-Hernandez, R.; Diego, J.C.; Nozal, M.J.; Gozalez-Porto, A.V.; Bernal, J.L.; Higes, M. An exposure study to assess the potential impact of fipronil in treated sunflower seeds on honey bee colony losses in Spain. Pest. Manag. Sci. 2011, 67, 1320–1331. [Google Scholar] [CrossRef] [PubMed]
- Budge, G.E.; Garthwaite, D.; Crowe, A.; Boatman, N.D.; Delaplane, K.S.; Brown, M.A.; Thygesen, H.H.; Pietravalle, S. Evidence for Pollinator Cost and Farming Benefits of Neonicotinoid Seed Coatings on Oilseed Rape. Sci. Rep. 2015, 5, 12574. [Google Scholar] [CrossRef] [Green Version]
- Balfour, N.J.; Al Toufailia, H.; Scandian, L.; Blanchard, H.E.; Jesse, M.P.; Carreck, N.L.; Ratnieks, F.L.W. Landscape Scale Study of the Net Effect of Proximity to a Neonicotinoid-Treated Crop on Bee Colony Health. Environ. Sci. Technol. 2017, 51, 10825–10833. [Google Scholar] [CrossRef]
- Seeburger, V.C.; D’Alvise, P.; Shaaban, B.; Schweikert, K.; Lohaus, G.; Schroeder, A.; Hasselmann, M. The Trisaccharide Melezitose Impacts Honey Bees and Their Intestinal Microbiota. PLoS ONE 2020, 15, e0230871. [Google Scholar] [CrossRef] [Green Version]
- Gregorc, A. Monitoring of Honey Bee Colony Losses: A Special Issue. Diversity 2020, 12, 403. [Google Scholar] [CrossRef]
- Özcan, M.; Arslan, D.; Ali Ceylan, D. Effect of Inverted Saccharose on Some Properties of Honey. Food Chem. 2006, 99, 24–29. [Google Scholar] [CrossRef]
- Nguyen, B.K.; Ribière, M.; vanEngelsdorp, D.; Snoeck, C.; Saegerman, C.; Kalkstein, A.L.; Schurr, F.; Brostaux, Y.; Faucon, J.-P.; Haubruge, E. Effects of Honey Bee Virus Prevalence, Varroa destructor Load and Queen Condition on Honey Bee Colony Survival over the Winter in Belgium. J. Apic. Res. 2011, 50, 195–202. [Google Scholar] [CrossRef]
- Beyer, M.; Junk, J.; Eickermann, M.; Clermont, A.; Kraus, F.; Georges, C.; Reichart, A.; Hoffmann, L. Winter honey bee colony losses, Varroa destructor control strategies, and the role of weather conditions: Results from a survey among beekeepers. Res. Vet. Sci. 2018, 118, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Haber, A.I.; Steinhauer, N.A.; vanEngelsdorp, D. Use of chemical and nonchemical methods for the control of Varroa destructor (Acari: Varroidae) and associated winter colony losses in US beekeeping operations. J. Econ. Entomol. 2019, 112, 1509–1525. [Google Scholar] [CrossRef] [PubMed]
- Giusti, M.; Sabelli, C.; Di Donato, A.; Lamberti, D.; Paturzo, C.E.; Polignano, V.; Lazzari, R.; Felicioli, A. Efficacy and Safety of Varterminator, a New Formic Acid Medicine against the Varroa Mite. J. Apic. Res. 2017, 56, 162–167. [Google Scholar] [CrossRef]
- Maggi, M.D.; Amiani, N.D.; Uffinengo, S.R.; Rasesco, M.C.; Zawarski, N.S.; Itton, G.M.; Ariani, F.M.; Ammataro, D.S.; Uintana, S.Q.; Guaras, M.J. The susceptibility of Varroa destructor against oxalic acid: A study case. Bull. Insectol. 2017, 70, 39–44. [Google Scholar]
- Gregorc, A.; Alburaki, M.; Sampson, B.; Knight, P.R.; Adamczyk, J. Toxicity of selected acaricides to honey bees (Apis mellifera) and Varroa (Varroa destructor Anderson and Trueman) and their use in controlling varroa within honey bee colonies. Insects 2018, 9, 55. [Google Scholar] [CrossRef] [Green Version]
- Gregorc, A.; Adamczyk, J.; Kapun, S.; Planinc, I. Integrated Varroa Control in Honey Bee (Apis mellifera Carnica) Colonies with or without Brood. J. Apic. Res. 2017, 55, 253–258. [Google Scholar] [CrossRef]
- Gregorc, A.; Sampson, B. Diagnosis of Varroa Mite (Varroa destructor) and Sustainable Control in Honey Bee (Apis mellifera) Colonies—A Review. Diversity 2019, 11, 243. [Google Scholar] [CrossRef] [Green Version]
- Oddie, M.; Dahle, B.; Neumann, P. Reduced Postcapping Period in Honey Bees Surviving Varroa destructor by Means of Natural Selection. Insects 2018, 9, 149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamler, M.; Nesvorna, M.; Stara, J.; Erban, T.; Hubert, J. Comparison of Tau-Fluvalinate, Acrinathrin, and Amitraz Effects on Susceptible and Resistant Populations of Varroa destructor in a Vial Test. Exp. Appl. Acarol. 2016, 69, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gracia, M.J.; Moreno, C.; Ferrer, M.; Sanz, A.; Peribáñez, M.Á.; Estrada, R. Field Efficacy of Acaricides against Varroa destructor. PLoS ONE 2017, 12, e0171633. [Google Scholar] [CrossRef] [PubMed]
- Medici, S.K.; Maggi, M.D.; Sarlo, E.G.; Ruffinengo, S.; Marioli, J.M.; Eguaras, M.J. The Presence of Synthetic Acaricides in Beeswax and Its Influence on the Development of Resistance in Varroa destructor. J. Apic. Res. 2015, 54, 267–274. [Google Scholar] [CrossRef]
- Wilmart, O.; Legrève, A.; Scippo, M.-L.; Reybroeck, W.; Urbain, B.; de Graaf, D.C.; Steurbaut, W.; Delahaut, P.; Gustin, P.; Nguyen, B.K.; et al. Residues in Beeswax: A Health Risk for the Consumer of Honey and Beeswax? J. Agric. Food Chem. 2016, 64, 8425–8434. [Google Scholar] [CrossRef]
- Almecija, G.; Poirot, B.; Cochard, P.; Suppo, C. Inventory of Varroa destructor Susceptibility to Amitraz and Tau-Fluvalinate in France. Exp. Appl. Acarol. 2020, 82, 1–16. [Google Scholar] [CrossRef]
- Li, J.L.; Cornman, R.S.; Evans, J.D.; Pettis, J.S.; Zhao, Y.; Murphy, C.; Peng, W.J.; Wu, J.; Hamilton, M.; Boncristiani, H.F.; et al. Systemic Spread and Propagation of a Plant-Pathogenic Virus in European Honeybees, Apis mellifera. mBio 2014, 5, e00898-13. [Google Scholar] [CrossRef] [Green Version]
- Meixner, M.D.; Francis, R.M.; Gajda, A.; Kryger, P.; Andonov, S.; Uzunov, A.; Topolska, G.; Costa, C.; Amiri, E.; Berg, S.; et al. Occurrence of Parasites and Pathogens in Honey Bee Colonies Used in a European Genotype-Environment Interactions Experiment. J. Apic. Res. 2014, 53, 215–229. [Google Scholar] [CrossRef]
- Amiri, E.; Meixner, M.; Nielsen, S.L.; Kryger, P. Four Categories of Viral Infection Describe the Health Status of Honey Bee Colonies. PLoS ONE 2015, 10, e0140272. [Google Scholar] [CrossRef] [Green Version]
- McMenamin, A.J.; Genersch, E. Honey Bee Colony Losses and Associated Viruses. Curr. Opin. Insect Sci. 2015, 8, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Genersch, E.; von der Ohe, W.; Kaatz, H.; Schroeder, A.; Otten, C.; Büchler, R.; Berg, S.; Ritter, W.; Mühlen, W.; Gisder, S.; et al. The German Bee Monitoring Project: A Long Term Study to Understand Periodically High Winter Losses of Honey Bee Colonies. Apidologie 2010, 41, 332–352. [Google Scholar] [CrossRef] [Green Version]
- Cox-Foster, D.L.; Conlan, S.; Holmes, E.C.; Palacios, G.; Evans, J.D.; Moran, N.A.; Quan, P.-L.; Briese, T.; Hornig, M.; Geiser, D.M.; et al. A Metagenomic Survey of Microbes in Honey Bee Colony Collapse Disorder. Science 2007, 318, 283–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthoud, H.; Imdorf, A.; Haueter, M.; Radloff, S.; Neumann, P. Virus Infections and Winter Losses of Honey Bee Colonies (Apis mellifera). J. Apic. Res. 2010, 49, 60–65. [Google Scholar] [CrossRef]
- Highfield, A.C.; El Nagar, A.; Mackinder, L.C.M.; No�l, L.M.-L.J.; Hall, M.J.; Martin, S.J.; Schroeder, D.C. Deformed Wing Virus Implicated in Overwintering Honeybee Colony Losses. Appl. Environ. Microbiol. 2009, 75, 7212–7220. [Google Scholar] [CrossRef] [Green Version]
- Topolska, G.; Gajda, A.; Pohorecka, K.; Bober, A.; Kasprzak, S.; Skubida, M.; Semkiw, P. Winter colony losses in Poland. J. Apic. Res. 2010, 49, 126–128. [Google Scholar] [CrossRef]
- Dainat, B.; Neumann, P. Clinical Signs of Deformed Wing Virus Infection Are Predictive Markers for Honey Bee Colony Losses. J. Invertebr. Pathol. 2013, 112, 278–280. [Google Scholar] [CrossRef]
- Toplak, I.; Jamnikar Ciglenečki, U.; Aronstein, K.; Gregorc, A. Chronic Bee Paralysis Virus and Nosema Ceranae Experimental Co-Infection of Winter Honey Bee Workers (Apis mellifera L.). Viruses 2013, 5, 2282–2297. [Google Scholar] [CrossRef] [Green Version]
- Retschnig, G.; Neumann, P.; Williams, G.R. Thiacloprid–Nosema Ceranae Interactions in Honey Bees: Host Survivorship but Not Parasite Reproduction Is Dependent on Pesticide Dose. J. Invertebr. Pathol. 2014, 118, 18–19. [Google Scholar] [CrossRef]
- Doublet, V.; Labarussias, M.; de Miranda, J.R.; Moritz, R.F.; Paxton, R.J. Bees under stress: Sublethal doses of a neonicotinoid pesticide and pathogens interact to elevate honey bee mortality across the life cycle. Environ. Microbiol. 2015, 17, 969–983. [Google Scholar] [CrossRef]
- Tesovnik, T.; Cizelj, I.; Zorc, M.; Čitar, M.; Božič, J.; Glavan, G.; Narat, M. Immune related gene expression in worker honey bee (Apis mellifera carnica) pupae exposed to neonicotinoid thiamethoxam and Varroa mites (Varroa destructor). PLoS ONE 2017, 12, e0187079. [Google Scholar] [CrossRef]
- Straub, L.; Williams, G.R.; Vidondo, B.; Khongphinitbunjong, K.; Retschnig, G.; Schneeberger, A.; Chantawannakul, P.; Dietemann, V.; Neumann, P. Neonicotinoids and Ectoparasitic Mites Synergistically Impact Honeybees. Sci. Rep. 2019, 9, 8159. [Google Scholar] [CrossRef]
- Di Prisco, G.; Cavaliere, V.; Annoscia, D.; Varricchio, P.; Caprio, E.; Nazzi, F.; Gargiulo, G.; Pennacchio, F. Neonicotinoid Clothianidin Adversely Affects Insect Immunity and Promotes Replication of a Viral Pathogen in Honey Bees. Proc. Natl. Acad. Sci. USA 2013, 110, 18466–18471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coulon, M.; Schurr, F.; Martel, A.-C.; Cougoule, N.; Bégaud, A.; Mangoni, P.; Di Prisco, G.; Dalmon, A.; Alaux, C.; Ribière-Chabert, M.; et al. Influence of Chronic Exposure to Thiamethoxam and Chronic Bee Paralysis Virus on Winter Honey Bees. PLoS ONE 2019, 14, e0220703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pohorecka, K.; Szczęsna, T.; Witek, M.; Miszczak, A.; Sikorski, P. The Exposure of Honey Bees to Pesticide Residues in the Hive Environment with Regard to Winter Colony Losses. J. Apic. Sci. 2017, 61, 105–125. [Google Scholar] [CrossRef] [Green Version]
- Wade, A.; Lin, C.-H.; Kurkul, C.; Regan, E.R.; Johnson, R.M. Combined Toxicity of Insecticides and Fungicides Applied to California Almond Orchards to Honey Bee Larvae and Adults. Insects 2019, 10, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gajger, I.T.; Vlainić, J.; Šoštarić, P.; Prešern, J.; Bubnič, J.; Smodiš Škerl, M.I. Effects on Some Therapeutical, Biochemical, and Immunological Parameters of Honey Bee (Apis mellifera) Exposed to Probiotic Treatments, in Field and Laboratory Conditions. Insects 2020, 11, 638. [Google Scholar] [CrossRef] [PubMed]
- Schulz, M.; Łoś, A.; Grzybek, M.; Ścibior, R.; Strachecka, A. Piperine as a new natural supplement with beneficial effects on the life-span and defence system of honeybees. J. Agric. Sci. 2019, 157, 140–149. [Google Scholar] [CrossRef]
- Tawfik, A.I.; Ahmed, Z.H.; Abdel-Rahman, M.F.; Moustafa, A.M. Influence of winter feeding on colony development and the antioxidant system of the honey bee, Apis mellifera. J. Apic. Res. 2020, 59, 752–763. [Google Scholar] [CrossRef]
- Delaplane, K.S.; van der Steen, J.; Guzman-Novoa, E. Standard Methods for Estimating Strength Parameters of Apis mellifera Colonies. J. Apic. Res. 2013, 52, 1–12. [Google Scholar] [CrossRef]
- Borsuk, G.; Ptaszyńska, A.A.; Olszewski, K.; Domaciuk, M.; Krutmuang, P.; Paleolog, J. A New Method for Quick and Easy Hemolymph Collection from Apidae Adults. PLoS ONE 2017, 12, e0170487. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 93, 265–275. [Google Scholar] [CrossRef]
- Wang, T.; Riegger, A.; Lamla, M.; Wiese, S.; Oeckl, P.; Otto, M.; Wu, Y.; Fischer, S.; Barth, H.; Kuan, S.L.; et al. Water-Soluble Allyl Sulfones for Dual Site-Specific Labelling of Proteins and Cyclic Peptides. Chem. Sci. 2016, 7, 3234–3239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shumkova, R.; Balkanska, R.; Hristov, P. The Herbal Supplements NOZEMAT HERB® and NOZEMAT HERB PLUS®: An Alternative Therapy for N. ceranae Infection and Its Effects on Honey Bee Strength and Production Traits. Pathogens 2021, 10, 234. [Google Scholar] [CrossRef]
- Charistos, L.; Parashos, N.; Hatjina, F. Long Term Effects of a Food Supplement HiveAliveTM on Honey Bee Colony Strength and Nosema Ceranae Spore Counts. J. Apic. Res. 2015, 54, 420–426. [Google Scholar] [CrossRef]
- Al-Ghamdi, A.A.; Abou-Shaara, H.F.; Ansari, M.J. Effects of Sugar Feeding Supplemented with Three Plant Extracts on Some Parameters of Honey Bee Colonies. Saudi J. Biol. Sci. 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- Ptaszyńska, A.A.; Załuski, D. Extracts from Eleutherococcus Senticosus (Rupr. et Maxim.) Maxim. Roots: A New Hope Against Honeybee Death Caused by Nosemosis. Molecules 2020, 25, 4452. [Google Scholar] [CrossRef] [PubMed]
- Archer, C.R.; Köhler, A.; Pirk, C.W.W.; Oosthuizen, V.; Apostolides, Z.; Nicolson, S.W. Antioxidant Supplementation Can Reduce the Survival Costs of Excess Amino Acid Intake in Honeybees. J. Insect Physiol. 2014, 71, 78–86. [Google Scholar] [CrossRef] [Green Version]
- Stevanovic, J.; Stanimirovic, Z.; Simeunovic, P.; Lakic, N.; Radovic, I.; Sokovic, M.; Griensven, L.J.L.D.V. The Effect of Agaricus Brasiliensis Extract Supplementation on Honey Bee Colonies. An. Acad. Bras. Ciênc. 2018, 90, 219–229. [Google Scholar] [CrossRef]
- Singh, M.; Pandey, N.; Agnihotri, V.; Singh, K.K.; Pandey, A. Antioxidant, Antimicrobial Activity and Bioactive Compounds of Bergenia Ciliata Sternb.: A Valuable Medicinal Herb of Sikkim Himalaya. J. Tradit. Complement. Med. 2017, 7, 152–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, D.-P.; Li, Y.; Meng, X.; Zhou, T.; Zhou, Y.; Zheng, J.; Zhang, J.-J.; Li, H.-B. Natural Antioxidants in Foods and Medicinal Plants: Extraction, Assessment and Resources. Int. J. Mol. Sci. 2017, 18, 96. [Google Scholar] [CrossRef]
- Jain, C.; Khatana, S.; Vijayvergia, R. Bioactivity of secondary metabolites of various plants: A review. Int. J. Pharm. Sci. Res. 2019, 10, 494–498. [Google Scholar]
- Rosado-Pérez, J.; Aguiñiga-Sánchez, I.; Arista-Ugalde, T.L.; Santiago-Osorio, E.; Mendoza-Núñez, V.M. The Biological Significance of Oxidative Stress, Effects of Fruits as Natural Edible Antioxidants. Curr. Pharm. Des. 2019, 24, 4807–4824. [Google Scholar] [CrossRef]
- Moghrovyan, A.; Sahakyan, N.; Babayan, A.; Chichoyan, N.; Petrosyan, M.; Trchounian, A. Essential oil and ethanol extract of oregano (Origanum vulgare L.) from Armenian flora as a natural source of terpenes, flavonoids and other phytochemicals with antiradical, antioxidant, metal chelating, tyrosinase inhibitory and antibacterial activity. Curr. Pharm. Des. 2019, 25, 1809–1816. [Google Scholar] [CrossRef]
- Geneva, M.; Kostadinov, K.; Filipov, S.; Kirova, E.; Stancheva, I. Analysis of the antioxidant capacity of lettuce growth at different fertilizer regimes. CR Acad. Bulg. Sci. 2021, 74, 1–10. [Google Scholar]
- Alonso-Salces, R.M.; Cugnata, N.M.; Guaspari, E.; Pellegrini, M.C.; Aubone, I.; De Piano, F.G.; Antunez, K.; Fuselli, S.R. Natural Strategies for the Control of Paenibacillus larvae, the Causative Agent of American Foulbrood in Honey Bees: A Review. Apidologie 2017, 48, 387–400. [Google Scholar] [CrossRef]
- Isidorov, V.A.; Buczek, K.; Segiet, A.; Zambrowski, G.; Swiecicka, I. Activity of Selected Plant Extracts against Honey Bee Pathogen Paenibacillus larvae. Apidologie 2018, 49, 687–704. [Google Scholar] [CrossRef] [Green Version]
- Kirova, E.; Pecheva, D.; Simova-Stoilova, L. Drought response in winter wheat: Protection from oxidative stress and mutagenesis effect. Acta Physiol. Plant. 2021, 43, 8. [Google Scholar] [CrossRef]
- Mao, W.; Schuler, M.A.; Berenbaum, M.R. A Dietary Phytochemical Alters Caste-Associated Gene Expression in Honey Bees. Sci. Adv. 2015, 1, e1500795. [Google Scholar] [CrossRef] [Green Version]
- Gong, Y.; Diao, Q. Current Knowledge of Detoxification Mechanisms of Xenobiotic in Honey Bees. Ecotoxicology 2017, 26, 1–12. [Google Scholar] [CrossRef]
- Liao, L.-H.; Wu, W.-Y.; Berenbaum, M. Impacts of Dietary Phytochemicals in the Presence and Absence of Pesticides on Longevity of Honey Bees (Apis mellifera). Insects 2017, 8, 22. [Google Scholar] [CrossRef] [Green Version]
- Manjon, C.; Troczka, B.J.; Zaworra, M.; Beadle, K.; Randall, E.; Hertlein, G.; Singh, K.S.; Zimmer, C.T.; Homem, R.A.; Lueke, B.; et al. Unravelling the Molecular Determinants of Bee Sensitivity to Neonicotinoid Insecticides. Curr. Biol. 2018, 28, 1137–1143.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazarov, S.B.; Veleva, P.; Zhelyazkova, I. Statistical Models for Assessing the Influence of Hygienic Behaviour of Worker Bees on the Level of Lysozyme and Total Protein Content in Their Hаemolymph. Iran. J. Appl. Anim. Sci. 2020, 10, 365–373. [Google Scholar]
- Leclercq, G.; Pannebakker, B.; Gengler, N.; Nguyen, B.K.; Francis, F. Drawbacks and Benefits of Hygienic Behavior in Honey Bees (Apis mellifera L.): A Review. J. Apic. Res. 2017, 56, 366–375. [Google Scholar] [CrossRef] [Green Version]
- Reeves, A.M.; O’Neal, S.T.; Fell, R.D.; Brewster, C.C.; Anderson, T.D. In-Hive Acaricides Alter Biochemical and Morphological Indicators of Honey Bee Nutrition, Immunity, and Development. J. Insect Sci. 2018, 18, 8. [Google Scholar] [CrossRef]
- Kunc, M.; Dobeš, P.; Hurychová, J.; Vojtek, L.; Poiani, S.B.; Danihlík, J.; Havlík, J.; Titěra, D.; Hyršl, P. The Year of the Honey Bee (Apis mellifera L.) with Respect to Its Physiology and Immunity: A Search for Biochemical Markers of Longevity. Insects 2019, 10, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knoll, S.; Pinna, W.; Varcasia, A.; Scala, A.; Cappai, M.G. The honey bee (Apis mellifera L., 1758) and the seasonal adaptation of productions. Highlights on summer to winter transition and back to summer metabolic activity. A review. Livest. Sci. 2020, 235, 104011. [Google Scholar] [CrossRef]
- Danihlík, J.; Aronstein, K.; Petřivalský, M. Antimicrobial Peptides: A Key Component of Honey Bee Innate Immunity: Physiology, Biochemistry, and Chemical Ecology. J. Apic. Res. 2015, 54, 123–136. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, C.; Qi, S.; He, J.; Bai, Y. The Sublethal Effects of Ethiprole on the Development, Defense Mechanisms, and Immune Pathways of Honeybees (Apis mellifera L.). Environ. Geochem. Health 2021, 43, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Alaux, C.; Ducloz, F.; Crauser, D.; Le Conte, Y. Diet effects on honeybee immunocompetence. Biol. Lett. 2010, 6, 562–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orsavová, J.; Hlaváčová, I.; Mlček, J.; Snopek, L.; Mišurcová, L. Contribution of phenolic compounds, ascorbic acid and vitamin E to antioxidant activity of currant (Ribes L.) and gooseberry (Ribes uva-crispa L.) fruits. Food Chem. 2019, 284, 323–333. [Google Scholar] [CrossRef]
- Perna, A.; Intaglietta, I.; Simonetti, A.; Gambacorta, E. A comparative study on phenolic profile, vitamin C content and antioxidant activity of Italian honeys of different botanical origin. Int. J. Food Sci. Technol. 2013, 48, 1899–1908. [Google Scholar] [CrossRef]
- Farjan, M.; Dmitryjuk, M.; Lipiński, Z.; Biernat-Łopieńska, E.; Żółtowska, K. Supplementation of the honey bee diet with vitamin C: The effect on the antioxidative system of Apis mellifera carnica brood at different stages. J. Apic. Res. 2012, 51, 263–270. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Rank | Food Consumption from One Frame with Bees (Kg) | Assessment of the Parameter |
|---|---|---|
| 5 | 0.6–1.0 | Very good wintering |
| 4 | 1.1–1.4 | Good wintering |
| 3 | 1.5–1.8 | Satisfactory wintering |
| 2 | 1.9–2.2 | Bad wintering |
| 1 | Over 2.3 | Very bad wintering |
| Rank | Quantity of Fead Bees (%) | Assessment of the Parameter |
|---|---|---|
| 5 | Under 15 | Very good wintering |
| 4 | 15–19.99 | Good wintering |
| 3 | 20–29.99 | Satisfactory wintering |
| 2 | 30–49.99 | Bad wintering |
| 1 | Over 50 | Very bad wintering |
| Groups | Quantity of Honey in Autumn Review (kg) | Amount of Honey-Sugar Paste Given in the Winter (kg) | Quantity of Honey in Spring Review (kg) | Total Amount of Consumed Food (kg) | Consumed Food per One Frame of Bees (kg) |
|---|---|---|---|---|---|
| Control (C) | 7.14 ± 0.13 a | 3.0 | 0.48 ± 0.27 a | 9.67 ± 0.19 a | 2.08 ± 0.19 a |
| NOZEMAT HERB® | 7.18 ± 0.05 a | 3.0 | 3.42 ± 0.27 b | 6.76 ± 0.30 b | 1.29 ± 0.01 b |
| NOZEMAT HERB PLUS® | 7.31 ± 0.08 a | 3.0 | 3.24 ± 0.14 b | 7.07 ± 0.14 b | 1.38 ± 0.09 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shumkova, R.; Balkanska, R.; Koynarski, T.; Hristov, P. Application of the Natural Products NOZEMAT HERB and NOZEMAT HERB PLUS Can Decrease Honey Bee Colonies Losses during the Winter. Diversity 2021, 13, 228. https://doi.org/10.3390/d13060228
Shumkova R, Balkanska R, Koynarski T, Hristov P. Application of the Natural Products NOZEMAT HERB and NOZEMAT HERB PLUS Can Decrease Honey Bee Colonies Losses during the Winter. Diversity. 2021; 13(6):228. https://doi.org/10.3390/d13060228
Chicago/Turabian StyleShumkova, Rositsa, Ralitsa Balkanska, Tsvetoslav Koynarski, and Peter Hristov. 2021. "Application of the Natural Products NOZEMAT HERB and NOZEMAT HERB PLUS Can Decrease Honey Bee Colonies Losses during the Winter" Diversity 13, no. 6: 228. https://doi.org/10.3390/d13060228
APA StyleShumkova, R., Balkanska, R., Koynarski, T., & Hristov, P. (2021). Application of the Natural Products NOZEMAT HERB and NOZEMAT HERB PLUS Can Decrease Honey Bee Colonies Losses during the Winter. Diversity, 13(6), 228. https://doi.org/10.3390/d13060228