
Reciprocal Garden Study Reveals Acute Spatial-Edaphic Adaptation for Cycas micronesica


Abstract
:
1. Introduction
2. Materials and Methods
3. Results
3.1. Soils
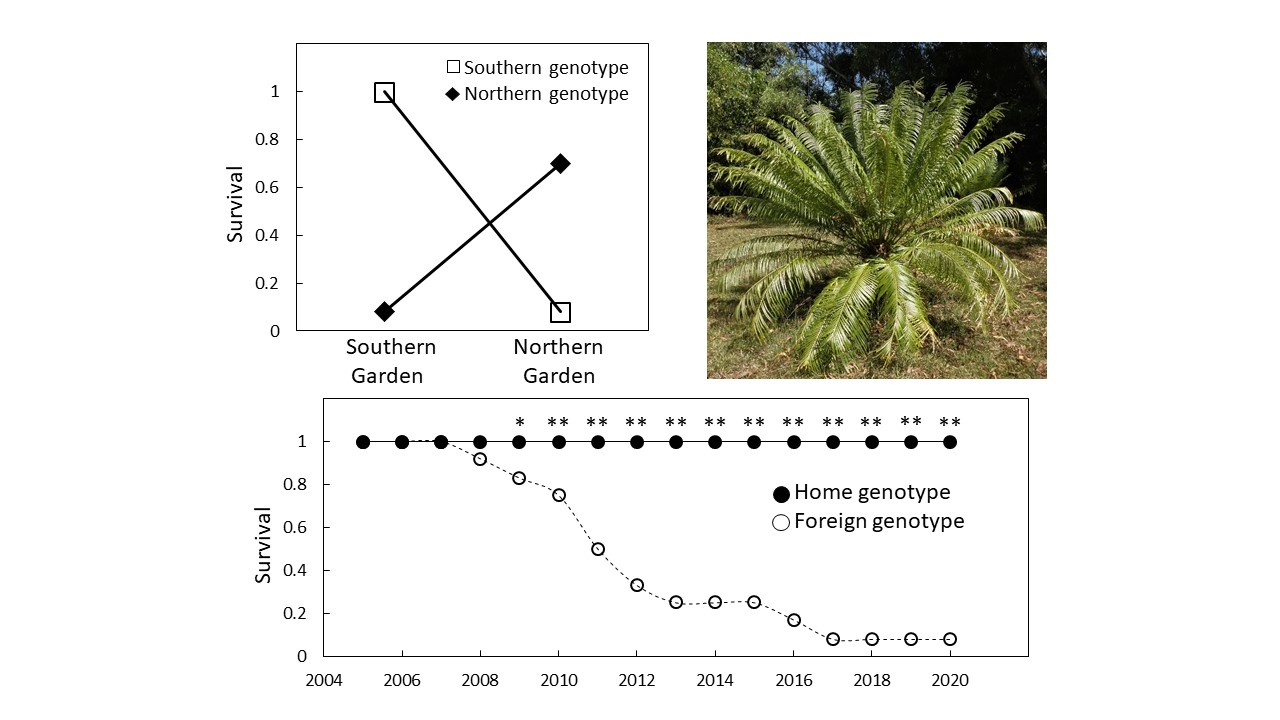
3.2. Survival
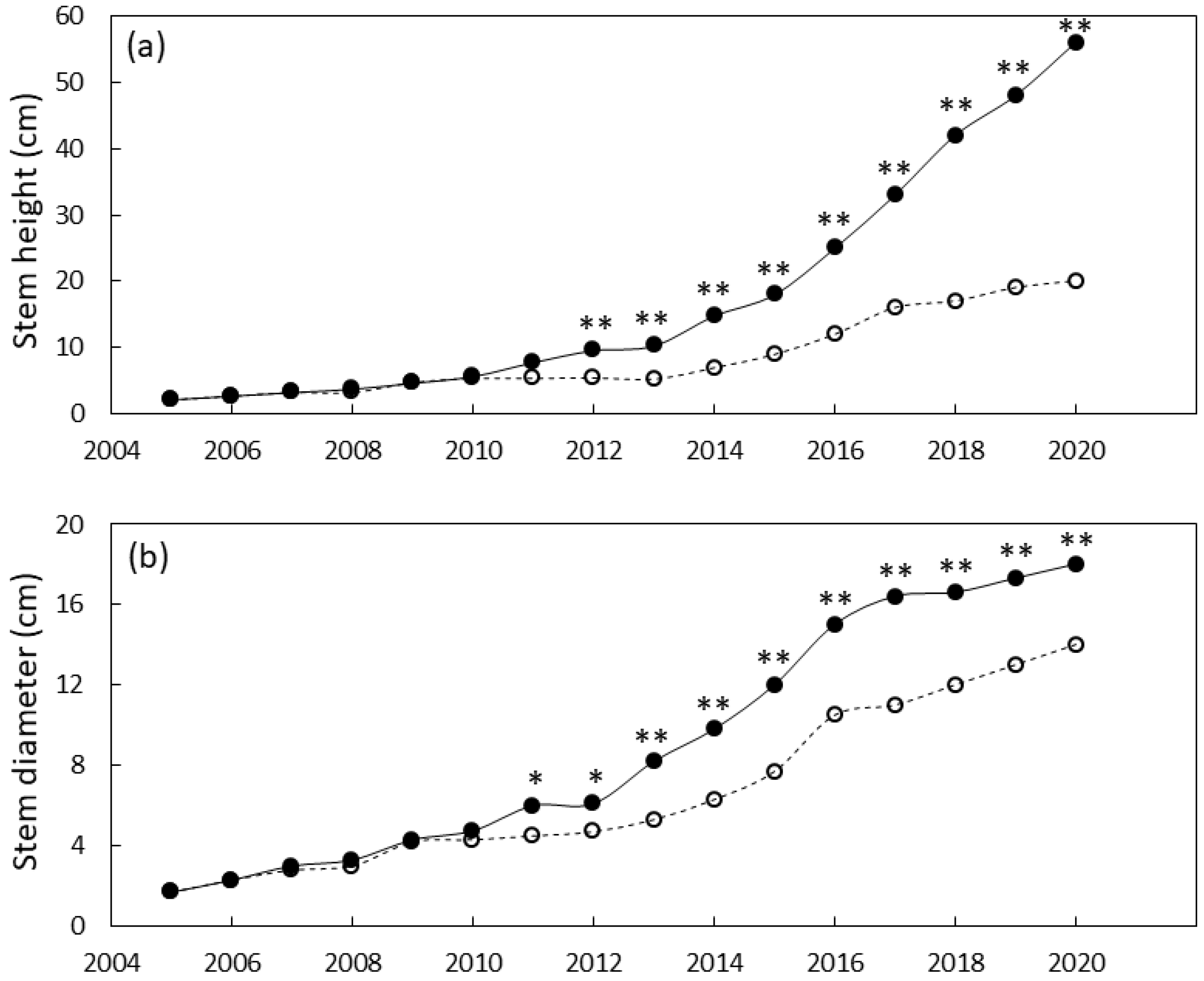
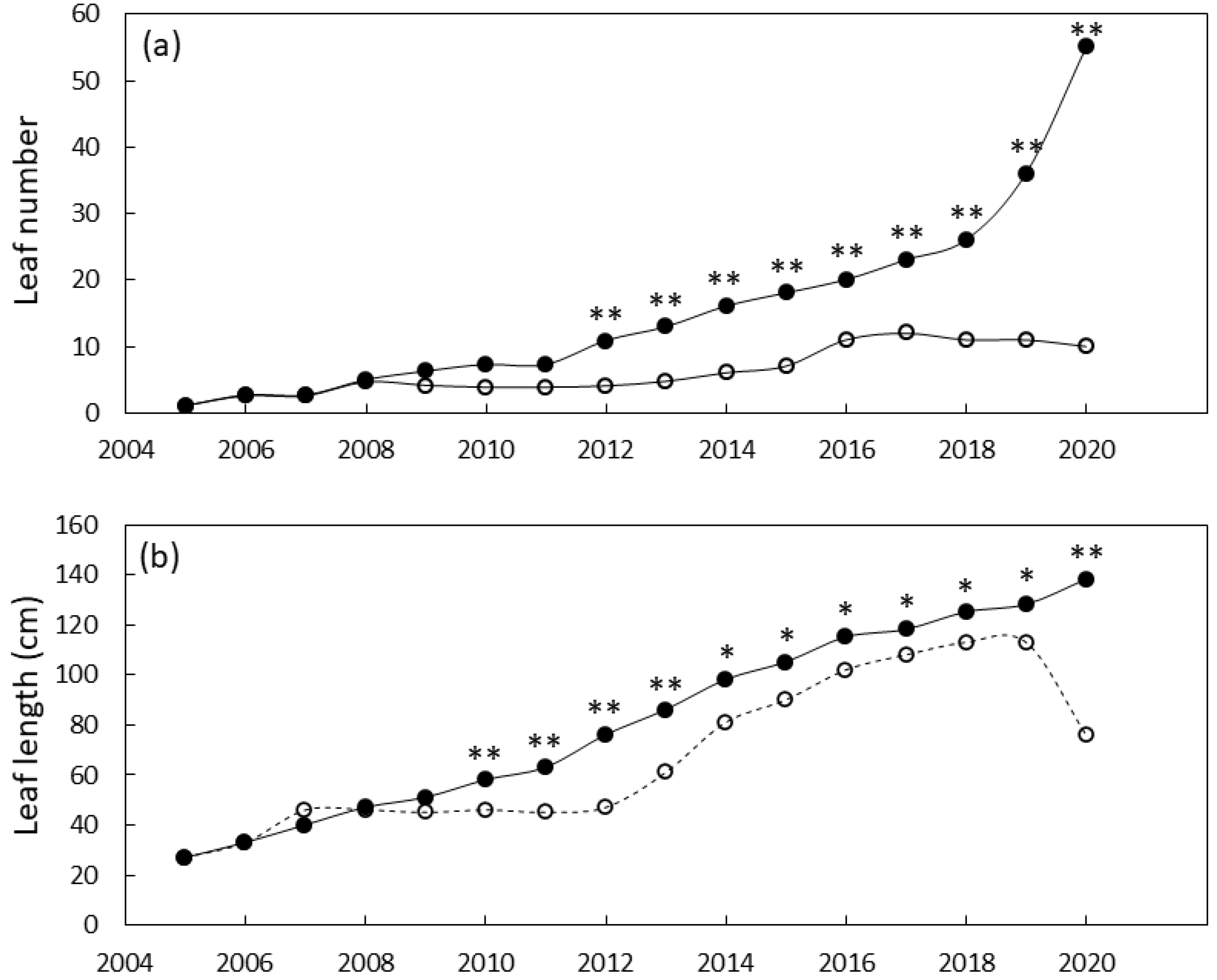
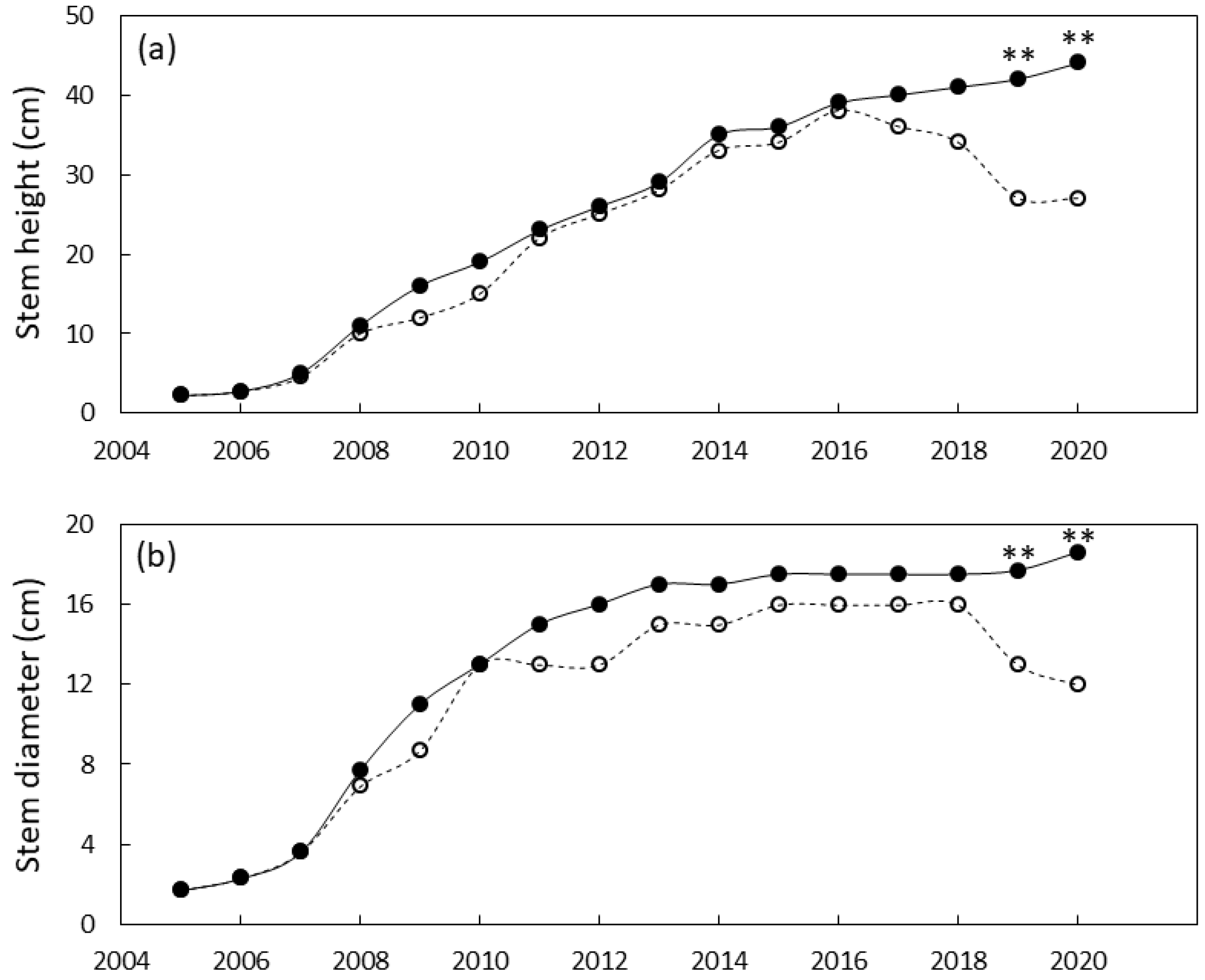
3.3. Plant Growth
4. Discussion
4.1. Lessons
4.2. Future Research Needs
4.3. Conservation Applications
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mounce, R.; Smith, P.; Brockington, S. Ex situ conservation of plant diversity in the world’s botanic gardens. Nat. Plants 2017, 3, 795–802. [Google Scholar] [CrossRef]
- Werden, L.K.; Sugii, N.C.; Weisenberger, L.; Keir, M.J.; Koob, G.; Zahawi, R.A. Ex situ conservation of threatened plant species in island biodiversity hotspots: A case study from Hawai‘i. Biol. Conserv. 2020, 243, 108435. [Google Scholar] [CrossRef]
- Westwood, M.; Cavender, N.; Meyer, A.; Smith, P. Botanic garden solutions to the plant extinction crisis. Plants People Planet 2020, 3, 22–32. [Google Scholar] [CrossRef]
- Viani, R.A.G.; Barreto, T.E.; Farah, F.T.; Rodrigues, R.R.; Brancalion, P.H.S. Monitoring young tropical forest restoration sites: How much to measure? Trop. Conserv. Sci. 2018, 11, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Volis, S. Conservation-oriented restoration—A two for one method to restore both threatened species and their habitats. Plant Divers. 2019, 41, 50–58. [Google Scholar] [CrossRef]
- Abeli, T.; Dixon, K. Translocation ecology: The role of ecological sciences in plant translocation. Plant Ecol. 2016, 217, 123–125. [Google Scholar] [CrossRef] [Green Version]
- Marler, T.E. Conservation of local genotypes when planting native plants on Guam. Micrones. Suppl. 2002, 6, 55–60. [Google Scholar]
- Broadhurst, L.M.; Lowe, A.; Coates, D.J.; Cunningham, S.A.; McDonald, M.; Vesk, P.A.; Yates, C. Seed supply for broadscale restoration: Maximizing evolutionary potential. Evol. Appl. 2008, 1, 587–597. [Google Scholar] [CrossRef]
- Johnson, L.C.; Olsen, J.T.; Tetreault, H.; DeLaCruz, A.; Bryant, J.; Morgan, T.J.; Knapp, M.; Bello, N.M.; Baer, S.G.; Maricle, B.R. Intraspecific variation of a dominant grass and local adaptation in reciprocal garden communities along a US Great Plains’ precipitation gradient: Implications for grassland restoration with climate change. Evol. Appl. 2015, 8, 705–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuismer, S.L.; Gandon, S. Moving beyond common-garden and transplant designs: Insight into the causes of local adaptation in species interactions. Amer. Nat. 2008, 171, 658–668. [Google Scholar] [CrossRef] [PubMed]
- Ågren, J.; Schemske, D.W. Reciprocal transplants demonstrate strong adaptive differentiation of the model organism Arabidopsis thaliana in its native range. New Phytol. 2012, 194, 1112–1122. [Google Scholar] [CrossRef]
- Fragniere, Y.; Bétrisey, S.; Cardinaux, L.; Stoffel, M.; Kozlowski, G. Fighting their last stand? A global analysis of the distribution and conservation status of gymnosperms. J. Biogeogr. 2015, 42, 809–820. [Google Scholar] [CrossRef]
- Marler, P.N.; Marler, T.E. An assessment of Red List data for the Cycadales. Trop. Conserv. Sci. 2015, 8, 1114–1125. [Google Scholar] [CrossRef] [Green Version]
- Calonje, M.; Stevenson, D.W.; Osborne, R. The World List of Cycads, Online Edition ed. 2013–2021. Available online: http://www.cycadlist.org (accessed on 29 May 2021).
- Lopez-Gallego, C. Genotype-by-environment interactions for seedling establishment across native and degraded-forest habitats in a long-lived cycad. Bot. Rev. 2013, 79, 542–558. [Google Scholar] [CrossRef]
- Deloso, B.E.; Ferreras, U.F.; Marler, T.E. Does phytogeography change with shifts in geopolitics? The curious case of cycads in the United States. Diversity 2020, 12, 445. [Google Scholar] [CrossRef]
- Hill, K.D. The Cycas rumphii complex (Cycadaceae) in New Guinea and the western Pacific. Aust. Syst. Bot. 1994, 7, 543–567. [Google Scholar] [CrossRef]
- Donnegan, J.A.; Butler, S.L.; Grabowiecki, W.; Hiserote, B.A.; Limtiaco, D. Guam’s Forest Resources, 2002. Resource Bulletin PNW-RB-243; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 2004.
- Deloso, B.E.; Terry, L.I.; Yudin, L.S.; Marler, T.E. Biotic threats to Cycas micronesica continue to expand to complicate conservation decisions. Insects 2020, 11, 888. [Google Scholar] [CrossRef] [PubMed]
- Marler, T.E.; Krishnapillai, M.V. Longitude, forest fragmentation, and plant size influence Cycas micronesica mortality following island insect invasions. Diversity 2020, 12, 194. [Google Scholar] [CrossRef]
- Marler, T.; Haynes, J.; Lindstrom, A. Cycas micronesica. The IUCN Red List of Threatened Species; E.T61316A12462113. IUCN Global Species Programme; Cambridge, UK, 2010; Available online: https://dx.doi.org/10.2305/IUCN.UK.2010-3.RLTS.T61316A12462113.en (accessed on 29 May 2021).
- United States Fish & Wildlife Service. Endangered and threatened wildlife and plants; endangered status for 16 species and threatened status for 7 species in Micronesia. Fed. Regist. 2015, 80, 59424–59497. [Google Scholar]
- Cibrian-Jaramillo, A.; Daly, A.C.; Brenner, E.; DeSalle, R.; Marler, T.E. When North and South don’t mix: Genetic connectivity of a recently endangered oceanic cycad, Cycas micronesica, in Guam using EST-microsatellites. Mol. Ecol. 2010, 19, 2364–2379. [Google Scholar] [CrossRef]
- Norstog, K.J.; Nicholls, T.J. The Biology of the Cycads; Cornell Univ. Press: Ithaca, NY, USA, 1997; ISBN 978-0-8014-3033-6. [Google Scholar]
- Santini, G.; Biondi, N.; Rodolfi, L.; Tredici, M.R. Plant biostimulants from cyanobacteria: An emerging strategy to improve yields and sustainability in agriculture. Plants 2021, 10, 643. [Google Scholar] [CrossRef]
- Young, F.J. Soil Survey of Territory of Guam; U. S. Dept. of Agric. Soil Conservation Service: Washington, DC, USA, 1988.
- Olsen, S.R.; Cole, C.V.; Watanabe, F.S.; Dean, L.A. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; U.S. Dept. of Agric. Circ.: Washington, DC, USA, 1954; p. 939.
- Hue, N.V.; Ikawa, H.; Huang, X. Predicting soil phosphorus requirements. In Plant Nutrient Management in Hawaii’s Soils, Approaches for Tropical and Subtropical Agriculture; Silva, J.A., Uchida, R., Eds.; College of Tropical Agriculture and Human Resources, University of Hawaii at Manoa: Honolulu, HI, USA, 2000; pp. 95–99. [Google Scholar]
- Berghage, R.D.; Krauskopf, D.M.; Warncke, D.D.; Widders, I. Micronutrient Testing of Plant Growth Media Extractant, Identification and Evaluation. Commun. Soil Sci. Plant Anal. 1987, 18, 1089–1109. [Google Scholar] [CrossRef]
- Mann, H.B.; Whitney, D.R. On a test of whether one of two random variables is stochastically larger than the other. Ann. Math. Stat. 1947, 18, 50–60. [Google Scholar] [CrossRef]
- Joshi, J.; Schmid, B.; Caldeira, M.C.; Dimitrakopoulos, P.G.; Good, J.; Harris, R.; Hector, A.; Huss-Danell, K.; Jumpponen, A.; Minns, A.; et al. Local adaptation enhances performance of common plant species. Ecol. Lett. 2001, 4, 536–544. [Google Scholar] [CrossRef] [Green Version]
- Baltzer, J.L.; Thomas, S.C.; Nilus, R.; Burslem, D.F.R.P. Edaphic specialization in tropical trees: Physiological correlates and responses to reciprocal transplantation. Ecology 2005, 86, 3063–3077. [Google Scholar] [CrossRef] [Green Version]
- Ellis, A.G.; Weis, A.E. Coexistence and differentiation of ‘flowering stones’: The role of local adaptation to soil microenvironment. J. Ecol. 2006, 94, 322–335. [Google Scholar] [CrossRef]
- Nagy, E.S.; Rice, K.J. Local adaptation in two subspecies of an annual plant: Implications for migration and gene flow. Evolution 1997, 51, 1079–1089. [Google Scholar] [CrossRef]
- Kawecki, T.J.; Ebert, D. Conceptual issues in local adaptation. Ecol. Lett. 2004, 7, 1225–1241. [Google Scholar] [CrossRef] [Green Version]
- Marler, T.E.; Lindström, A.J. The value of research to selling the conservation of threatened species: The case of Cycas micronesica. J. Threat. Taxa 2014, 6, 6523–6528. [Google Scholar] [CrossRef]
- Bennington, C.; Fetcher, N.; Vavrek, M.; Shaver, G.; Cummings, K.; McGraw, J. Home site advantage in two long-lived arctic plant species: Results from two 30-year reciprocal transplant studies. J. Ecol. 2012, 100, 841–851. [Google Scholar] [CrossRef] [Green Version]
- Schoen, D.J.; Stewart, S.C.; Lechowicz, M.J.; Bell, G. Partitioning the transplant site effect in reciprocal transplant experiments with Impatiens capensis and Impatiens pallida. Oecologia 1986, 70, 149–154. [Google Scholar] [CrossRef]
- Deloso, B.E.; Krishnapillai, M.V.; Ferreras, U.F.; Lindström, A.J.; Calonje, M.; Marler, T.E. Chemical element concentrations of cycad leaves: Do we know enough? Horticulturae 2020, 6, 85. [Google Scholar] [CrossRef]
- Miglia, K.; McArthur, E.; Redman, R.; Rodriguez, R.; Zak, J.; Freeman, D. Genotype, soil type, and locale effects on reciprocal transplant vigor, endophyte growth, and microbial functional diversity of a narrow sagebrush hybrid zone in Salt Creek Canyon, Utah. Amer. J. Bot. 2007, 94, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Sork, V.L.; Stowe, K.A.; Hochwender, C. Evidence for local adaptation in closely adjacent subpopulations of Northern Red Oak (Quercus rubra L.) expressed as resistance to leaf herbivores. Amer. Nat. 1993, 142, 928–936. [Google Scholar] [CrossRef]
- Marler, T.E.; Muniappan, R. Pests of Cycas micronesica leaf, stem, and male reproductive tissues with notes on current threat status. Micronesica 2006, 39, 1–9. [Google Scholar]
- Marler, T.E. Temporal variations in leaf miner, butterfly, and stem borer infestations of Cycas micronesica in relation to Aulacaspis yasumatsui incidence. HortScience 2013, 48, 1334–1338. [Google Scholar] [CrossRef]
- Griffith, M.P.; Barber, G.; Tucker Lima, J.; Barros, M.; Calonje, C.; Noblick, L.R.; Calonje, M.; Magellan, T.; Dosmann, M.; Thibault, T.; et al. Plant collection “half-life:” can botanic gardens weather the climate? Curator Mus. J. 2017, 60, 395–410. [Google Scholar] [CrossRef] [Green Version]
- Hoban, S. New guidance for ex situ gene conservation: Sampling realistic population systems and accounting for collection attrition. Biol. Conserv. 2019, 235, 199–208. [Google Scholar] [CrossRef]
- Marler, T.E.; Calonje, M. Stem branching of cycad plants informs horticulture and conservation decisions. Horticulturae 2020, 6, 65. [Google Scholar] [CrossRef]
- Marler, T.E.; Griffith, M.P.; Krishnapillai, M.V. Height increment of Cycas micronesica informs conservation decisions. Plant Signal. Behav. 2020, 15, e1830237. [Google Scholar] [CrossRef] [PubMed]
- Marler, T.E.; Matanane, F.C.; Terry, L.I. Burrowing activity of coconut rhinoceros beetle on Guam cycads. Commun. Integr. Biol. 2020, 13, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Marler, T.E. Three invasive tree species change soil chemistry in Guam forests. Forests 2020, 11, 279. [Google Scholar] [CrossRef] [Green Version]
- Joint Region Marianas. Final Integrated Natural Resources Management Plan Joint Region Marianas—Guam, Tinian, and Farallon de Medinilla; Prepared under Contract #SF1449-N40192-10-R-9915; NAVFAC Marianas: Piti, Guam, 2012. [Google Scholar]
- Marler, T.E.; Lindström, A.J. First, do no harm. Commun. Integr. Biol. 2017, 10, e1393593. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Levels |
|---|---|
| Genotype | Home versus Foreign |
| Year | 2005–2020 |
| Nutrient | North Garden | South Garden |
|---|---|---|
| Calcium | 6.56 ± 0.06 mg·g−1 | 0.72 ± 0.05 mg·g−1 |
| Carbon | 17.22 ± 0.65 mg·g−1 | 2.19 ± 0.63 mg·g−1 |
| Copper | 1.09 ± 0.04 µg·g−1 | 1.77 ± 0.03 µg·g−1 |
| Iron | 12.73 ± 1.13 µg·g−1 | 28.25 ± 2.66 µg·g−1 |
| Magnesium | 102.25 ± 1.38 µg·g−1 | 267.25 ± 8.98 µg·g−1 |
| Manganese | 5.11 ± 0.15 µg·g−1 | 7.63 ± 0.21 µg·g−1 |
| Nitrogen | 0.93 ± 0.06 mg·g−1 | 0.68 ± 0.11 mg·g−1 |
| Phosphorus | 42.25 ± 1.38 µg·g−1 | 6.03 ± 0.11 µg·g−1 |
| Potassium | 16.63 ± 0.55 µg·g−1 | 38.75 ± 4.84 µg·g−1 |
| Zinc | 1.37 ± 0.22 µg·g−1 | 1.91 ± 0.16 µg·g−1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marler, T.E. Reciprocal Garden Study Reveals Acute Spatial-Edaphic Adaptation for Cycas micronesica. Diversity 2021, 13, 237. https://doi.org/10.3390/d13060237
Marler TE. Reciprocal Garden Study Reveals Acute Spatial-Edaphic Adaptation for Cycas micronesica. Diversity. 2021; 13(6):237. https://doi.org/10.3390/d13060237
Chicago/Turabian StyleMarler, Thomas E. 2021. "Reciprocal Garden Study Reveals Acute Spatial-Edaphic Adaptation for Cycas micronesica" Diversity 13, no. 6: 237. https://doi.org/10.3390/d13060237
APA StyleMarler, T. E. (2021). Reciprocal Garden Study Reveals Acute Spatial-Edaphic Adaptation for Cycas micronesica. Diversity, 13(6), 237. https://doi.org/10.3390/d13060237