
Mediterranean Red Macro Algae Mats as Habitat for High Abundances of Serpulid Polychaetes





Abstract
:1. Introduction
- (1)
- What are the abundance and species richness of serpulid polychaetes in P. crispa mats compared to P. oceanica meadows?
- (2)
- Which species are found in both habitats and which species are unique to P. crispa mats?
- (3)
- What is the spatial variability of serpulid polychaetes associated with red algae mats?
2. Materials and Methods
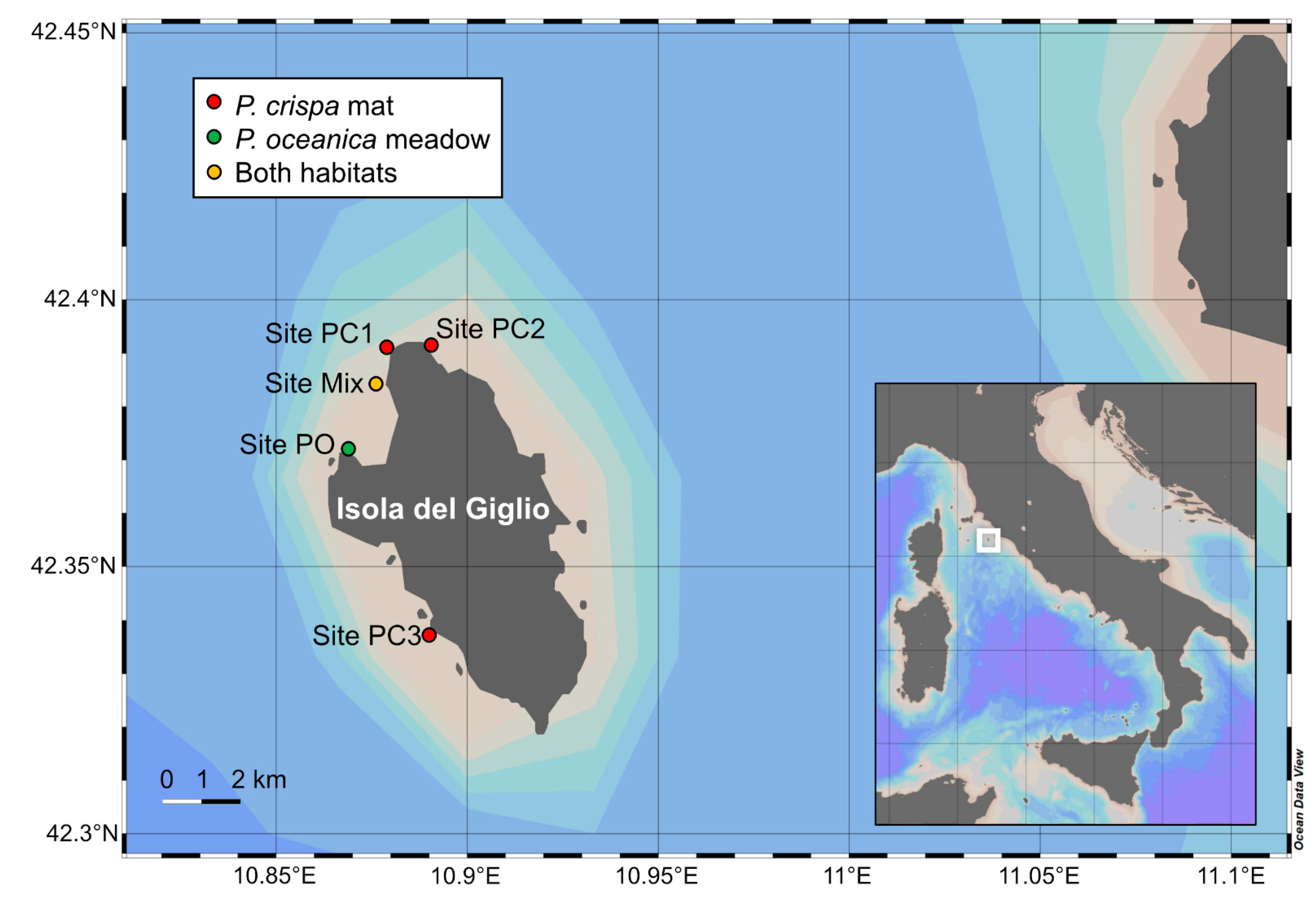
2.1. Study Area and Sampling Activities
2.2. Species Identification
2.3. Diversity Descriptors
2.4. Statistical Analysis
3. Results
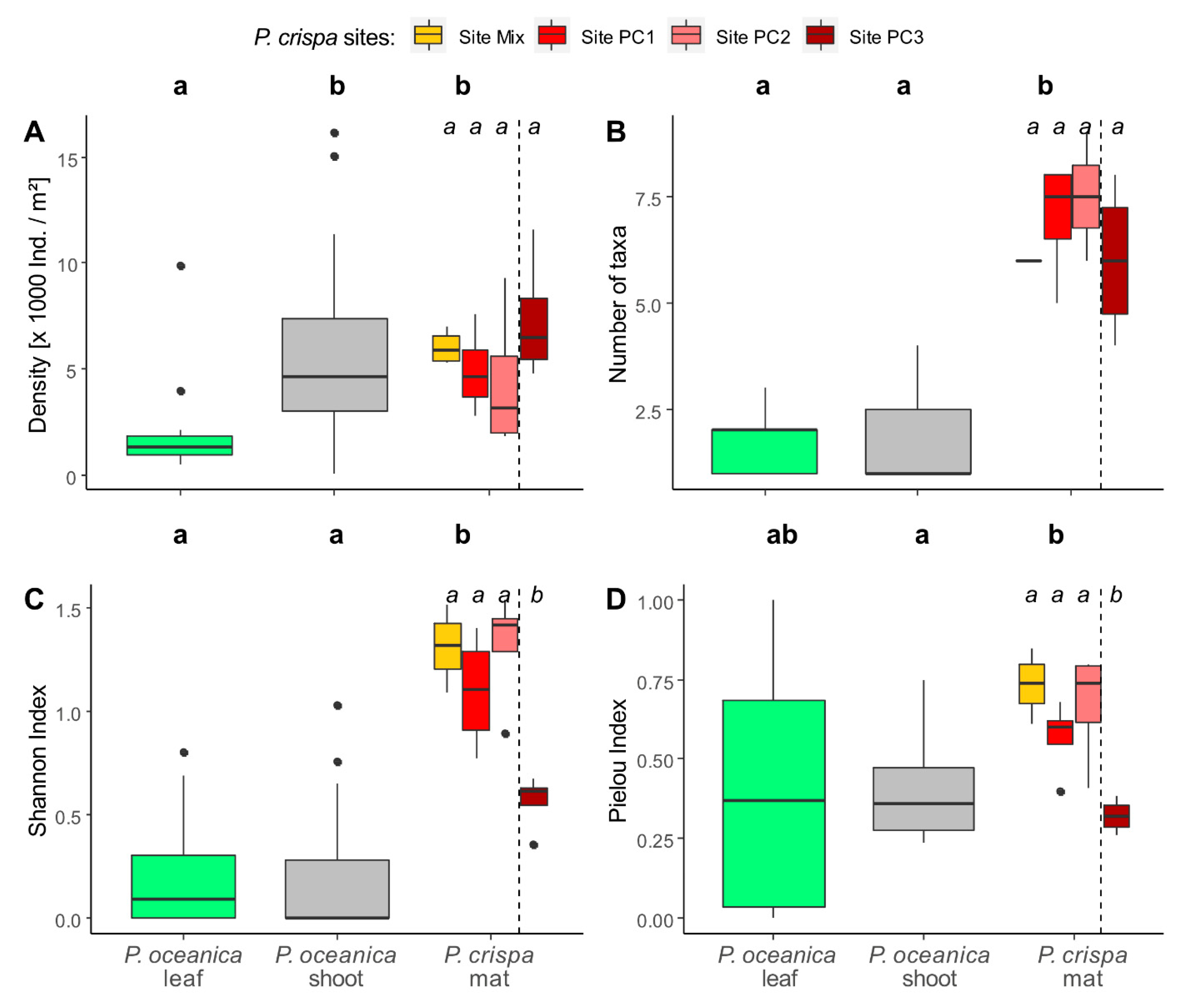
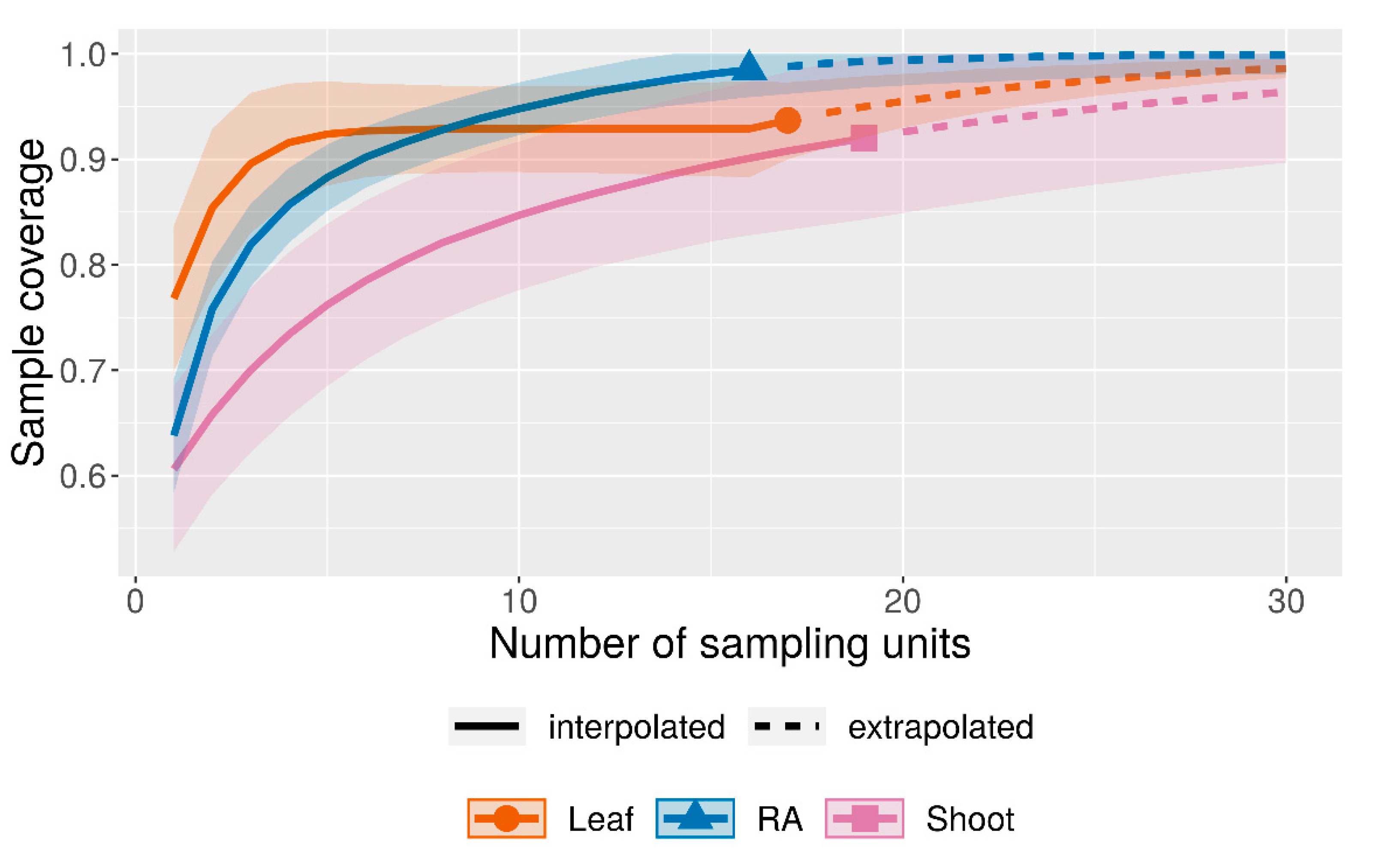
3.1. Diversity Descriptors
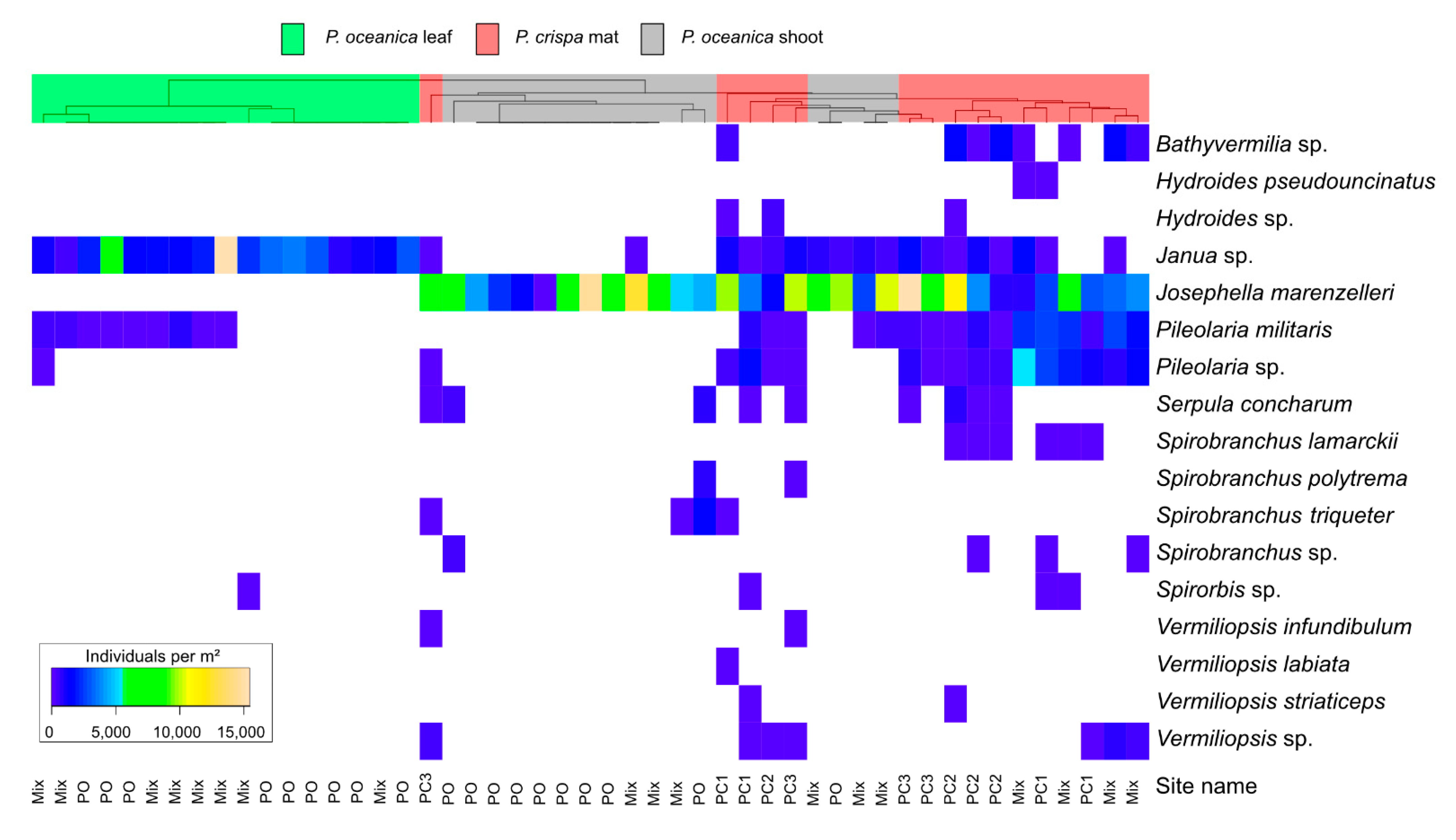
3.2. Analysis of Serpulid Assemblages
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author(s) | Year | Title |
|---|---|---|
| Riedl, R. | 2011 | Fauna und Flora des Mittelmeeres |
| Stresemann, E. | 1992 | Wirbellose |
| Hayward, P.J. and Ryland, J.S. | 1999 | Handbook of the Marine Fauna of North-West Europe |
| Bianchi et al. | 1981 | Guide per il riconoscimento delle specie animali delle acque lagunari e costiere italiane |
| Ten Hove et al. | 2009 | Taxonomy of Serpulidae (Annelida, Polychaeta): the state of affairs |
| Zibrowius H. | 1968 | Etude morphologique, systématique et écologique des Serpulidae (Annelida Polychaeta) de la région de Marseille |
| Zibrowius H. | 1972 | Mise au point sur les especes mediterraneennes de Serpulidae (Annelida Polychaeta) déecrites par Stefano delle Chiaje (1822–1829, 1841–1844) et Oronzio Gabriele Costa (1861) |
| Group 1 | Group 2 | p-Value | p-Signif | |
|---|---|---|---|---|
| Density | Site Mix | Site PC1 | 0.34 | ns |
| Site Mix | Site PC2 | 0.34 | ns | |
| Site Mix | Site PC3 | 0.69 | ns | |
| Site PC1 | Site PC2 | 0.69 | ns | |
| Site PC1 | Site PC3 | 0.34 | ns | |
| Site PC2 | Site PC3 | 0.20 | ns | |
| No of taxa | Site Mix | Site PC1 | 0.278 | ns |
| Site Mix | Site PC2 | 0.069 | ns | |
| Site Mix | Site PC3 | 1.000 | ns | |
| Site PC1 | Site PC2 | 0.766 | ns | |
| Site PC1 | Site PC3 | 0.454 | ns | |
| Site PC2 | Site PC3 | 0.306 | ns | |
| Shannon index | Site Mix | Site PC1 | 0.486 | ns |
| Site Mix | Site PC2 | 0.686 | ns | |
| Site Mix | Site PC3 | 0.029 | * | |
| Site PC1 | Site PC2 | 0.200 | ns | |
| Site PC1 | Site PC3 | 0.029 | * | |
| Site PC2 | Site PC3 | 0.029 | * | |
| Pielou index | Site Mix | Site PC1 | 0.057 | ns |
| Site Mix | Site PC2 | 0.886 | ns | |
| Site Mix | Site PC3 | 0.029 | * | |
| Site PC1 | Site PC2 | 0.200 | ns | |
| Site PC1 | Site PC3 | 0.029 | * | |
| Site PC2 | Site PC3 | 0.029 | * |
References
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Lasram, F.B.R.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The Biodiversity of the Mediterranean Sea: Estimates, Patterns, and Threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [Green Version]
- Medail, F.; Quezel, P. Biodiversity Hotspots in the Mediterranean. Conserv. Biol. 1999, 13, 1510. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Bianchi, C.N.; Morri, C. Marine Biodiversity of the Mediterranean Sea: Situation, Problems and Prospects for Future. Mar. Pollut. Bull. 2016, 40, 367–376. [Google Scholar] [CrossRef]
- Defant, A. Physical Oceanography; Pergamon: New York, NY, USA, 1961; Volume 1. [Google Scholar]
- Boudouresque, C.F.; Bernard, G.; Bonhomme, P.; Charbonnel, E.; Diviacco, G.; Meinesz, A.; Pergent, G.; Pergent-Martini, C.; Ruitton, S.; Tunesi, L. Préservation et Conservation des Herbiers à Posidonia Oceanica; Ramoge and RAC/SPA: Tunis, Tunesia, 2006; ISBN 2905540303. [Google Scholar]
- Mazzella, L.; Buia, M.C.; Gambi, M.C.; Lorenti, M.; Russo, G.F.; Scipione, M.B.; Zupo, V. Plant-animal trophic relationships in the Posidonia oceanica ecosystem of the Mediterranean Sea: A review. Plant Anim. Interact. Mar. Benthos 1992, 46, 165–187. [Google Scholar]
- Ballesteros, E. Mediterranean coralligenous assemblages: A synthesis of present knowledge. In Oceanography and Marine Biology: An Annual Review; Gibson, R.N., Atkinson, R.J.A., Gordon, J.D.M., Eds.; Taylor & Francis: London, UK, 2006; pp. 123–195. [Google Scholar]
- Cocito, S. Bioconstruction and biodiversity: Their mutual influence. Sci. Mar. 2004, 68, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Ingrosso, G.; Abbiati, M.; Badalamenti, F.; Bavestrello, G.; Belmonte, G.; Cannas, R.; Benedetti-Cecchi, L.; Bertolino, M.; Bevilacqua, S.; Bianchi, C.N.; et al. Mediterranean Bioconstructions Along the Italian Coast, 1st ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2018; Volume 79, ISBN 9780128151013. [Google Scholar]
- Lepoint, G.; Balancier, B.; Gobert, S. Seasonal and depth-related biodiversity of leaf epiphytic Cheilostome Bryozoa in a Mediterranean Posidonia oceanica meadow. Cah. Biol. Mar. 2014, 55, 57–67. [Google Scholar]
- Stachowicz, J.J. The Structure of Ecological Communities. Bioscience 2001, 51, 235–246. [Google Scholar] [CrossRef]
- Stachowicz, J.J.; Hay, M.E. Facultative mutualism between an herbivorous crab and a coralline alga: Advantages of eating noxious seaweeds. Oecologia 1996, 105, 377–387. [Google Scholar] [CrossRef]
- Duffy, J.E. Amphipods on seaweeds: Partners or pests? Oecologia 1990, 83, 267–276. [Google Scholar] [CrossRef]
- Tomas, F.; Turon, X. Seasonal and small-scale spatial variability of herbivory pressure on the temperate seagrass Posidonia oceanica. Mar. Ecol. Prog. Ser. 2005, 301, 95–107. [Google Scholar] [CrossRef]
- Piazzi, L.; Balata, D.; Ceccherelli, G. Epiphyte assemblages of the Mediterranean seagrass Posidonia oceanica: An overview. Mar. Ecol. 2016, 37, 3–41. [Google Scholar] [CrossRef]
- Casoli, E.; Bonifazi, A.; Gravina, M.F.; Russo, G.F.; Sandulli, R.; Donnarumma, L. Comparative Analysis of Mollusc Assemblages from Different Hard Bottom Habitats in the Central Tyrrhenian Sea. Diversity 2019, 11, 74. [Google Scholar] [CrossRef] [Green Version]
- Bonifazi, A.; Ventura, D.; Gravina, M.F.; Lasinio, G.J.; Belluscio, A.; Ardizzone, G.D. Unusual algal turfs associated with the rhodophyta Phyllophora crispa: Benthic assemblages along a depth gradient in the Central Mediterranean Sea. Estuar. Coast. Shelf Sci. 2017, 185, 77–93. [Google Scholar] [CrossRef]
- Kostylev, E.F.; Tkachenko, F.P.; Tretiak, I.P. Establishment of “Zernov’s Phyllophora field” marine reserve: Protection and restoration of a unique ecosystem. Ocean Coast. Manag. 2010, 53, 203–208. [Google Scholar] [CrossRef]
- Casoli, E.; Bonifazi, A.; Ardizzone, G.; Gravina, M.F. How algae influence sessile marine organisms: The tube worms case of study. Estuar. Coast. Shelf Sci. 2016, 178, 12–20. [Google Scholar] [CrossRef]
- Navone, A.; Bianchi, C.N.; Orru, P.; Ulzega, A. Saggio di cartografia geomorfologica e bionomica nel parco marino di Tavolara-Capo Coda di Cavallo (Sardegna nord-orientale). Oebalia 1992, XVII, 469–478. [Google Scholar]
- Bianchi, C.N.; Morri, C.; Navone, A. I popolamenti delle scogliere rocciose sommerse dell’Area Marina Protetta di Tavolara Punta Coda Cavallo (Sardegna nord-orientale). Sci. Rep. Port Cros Natl. Park 2010, 24, 39–85. [Google Scholar]
- Donnarumma, L.; Sandulli, R.; Appolloni, L.; Russo, G.F. Assessing molluscs functional diversity within different coastal habitats of Mediterranean marine protected areas. Ecol. Quest. 2018, 29, 35–51. [Google Scholar] [CrossRef] [Green Version]
- Buonocore, E.; Donnarumma, L.; Appolloni, L.; Miccio, A.; Russo, G.F.; Franzese, P.P. Marine natural capital and ecosystem services: An environmental accounting model. Ecol. Modell. 2020, 424, 109029. [Google Scholar] [CrossRef]
- Horton, T.; Kroh, A.; Ahyong, S.; Bailly, N.; Boyko, C.B.; Brandão, S.N.; Gofas, S.; Hooper, J.N.A.; Hernandez, F.; Holovachov, O.; et al. World Register of Marine Species (WoRMS). Available online: https://www.marinespecies.org (accessed on 27 January 2021).
- Giangrande, A.; Gravina, M.F. Brackish-water polychaetes, good descriptors of environmental changes in space and time. Transit. Waters Bull. 2015, 9, 42–55. [Google Scholar]
- Casoli, E.; Ricci, S.; Antonelli, F.; Sacco Perasso, C.; Ardizzone, G.; Gravina, M.F. Colonization dynamic on experimental limestone substrata: The role of encrusting epilithics favouring boring polychaetes. Hydrobiologia 2019, 842, 101–112. [Google Scholar] [CrossRef]
- Musco, L. Ecology and diversity of Mediterranean hard-bottom Syllidae (Annelida): A community-level approach. Mar. Ecol. Prog. Ser. 2012, 461, 107–119. [Google Scholar] [CrossRef] [Green Version]
- Giangrande, A.; Licciano, M.; Musco, L. Polychaetes as environmental indicators revisited. Mar. Pollut. Bull. 2005, 50, 1153–1162. [Google Scholar] [CrossRef]
- Watson, D.I.; Barnes, D.K.A. Quantifying assemblage distinctness with time: An example using temperate epibenthos. J. Exp. Mar. Bio. Ecol. 2004, 312, 367–383. [Google Scholar] [CrossRef]
- Ben-Eliahu, M.N.; Fiege, D. Serpulid tube-worms (Annelida: Polychaeta) of the Central and Eastern Mediterranean with particular attention to the Levant Basin. Senckenberg. Marit. 1996, 28, 1–51. [Google Scholar] [CrossRef]
- Piazzi, L.; Balata, D.; Cinelli, F. Epiphytic macroalgal assemblages of Posidonia oceanica rhizomes in the western Mediterranean. Eur. J. Phycol. 2002, 37, 69–76. [Google Scholar] [CrossRef]
- Fagerstrom, J.A. Reef-building guilds and a checklist for determining guild membership—A new approach for study of communities. Coral Reefs 1991, 10, 47–52. [Google Scholar] [CrossRef]
- Sanfilippo, R.; Vertino, A.; Rosso, A.; Beuck, L.; Freiwald, A.; Taviani, M. Serpula aggregates and their role in deep-sea coral communities in the southern Adriatic Sea. Facies 2013, 59, 663–677. [Google Scholar] [CrossRef]
- Vinn, O.; Ten Hove, H.A.; Mutvei, H.; Kirsimäe, K. Ultrastructure and mineral composition of serpulid tubes (Polychaeta, Annelida). Zool. J. Linn. Soc. 2008, 154, 633–650. [Google Scholar] [CrossRef]
- Ten Hove, H.A.; Kupriyanova, E.K. Taxonomy of Serpulidae (Annelida, Polychaeta): The State of Affairs. Zootaxa 2009, 2036, 1–126. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, C.N. Ecologia dei Serpuloidea (Annelida, Polychaeta) del piano infralitorale presso Portofino (Genova). Boll. Musei Inst. Biol. Univ. Genova 1979, 47, 101–115. [Google Scholar]
- Çinar, M.E. Serpulid species (Polychaeta: Serpulidae) from the Levantine coast of Turkey (eastern Mediterranean), with special emphasis on alien species. Aquat. Invasions 2006, 1, 223–240. [Google Scholar] [CrossRef]
- Balduzzi, A.; Bianchi, C.N.; Burlando, B.; Cattaneo-Vietti, R.; Manconi, R.; Morri, C.; Pansini, M.; Pronzato, R.; Sara, M. Zoobenthos di substrato duro delle isole di Capraia e del Giglio (Arcipelago Toscano). Atti Soc. Toscana Sci. Nat. Resid. 1995, 52, 12. [Google Scholar]
- Zenetos, A.; Çinar, M.E.; Panucci-Papadopoulou, M.A.; Harmelin, J.G.; Furnari, G.; Andaloro, F.; Bellou, N.; Streftaris, N.; Zibrowius, H. Annotated list of marine alien species in the Mediterranean with records of the worst invasive species. Mediterr. Mar. Sci. 2005, 6, 63–118. [Google Scholar] [CrossRef]
- Zibrowius, H. Ongoing modification of the Mediterranean marine fauna and flora by the establishment of exotic species. Mésogée 1991, 51, 83–107. [Google Scholar]
- Hopkins, G.A.; Forrest, B.M. Management options for vessel hull fouling: An overview of risks posed by in-water cleaning. ICES J. Mar. Sci. 2008, 65, 811–815. [Google Scholar] [CrossRef] [Green Version]
- Abbiati, M.; Bianchi, C.N.; Castelli, A. Polychaete Vertical Zonation along a Littoral Cliff in the Western Méditerranean. Mar. Ecol. 1987, 8, 33–48. [Google Scholar] [CrossRef]
- Giangrande, A. Polychaete zonation and its relation to algal distribution down a vertical cliff in the western Mediterranean (Italy): A structural analysis. J. Exp. Mar. Bio. Ecol. 1988, 120, 263–276. [Google Scholar] [CrossRef]
- Kikuchi, T. Handbook of Seagrass Biology: An Ecosystem Perspective; Phillips, R.C., McRoy, C.P., Eds.; Garland STPM Press: New York, NY, USA, 1980. [Google Scholar]
- Kikuchi, T.; Pérès, J.M. Animal communities in seagrass beds: A review. In Seagrass Ecosystems: A Scientific Perspective; McRoy, C.P., Helfferich, C., Eds.; Marcel Dekker: New York, NY, USA, 1967; pp. 147–193. [Google Scholar]
- Mounir, B.B.; Asma, H.; Sana, B.I.; Lotfi, M.; Abderrahmen, B.; Lotfi, A. What factors drive seasonal variation of phytoplankton, protozoans and metazoans on leaves of Posidonia oceanica and in the water column along the coast of the Kerkennah Islands, Tunisia? Mar. Pollut. Bull. 2013, 71, 286–298. [Google Scholar] [CrossRef]
- Donnarumma, L.; Lombardi, C.; Cocito, S.; Gambi, M.C. Settlement pattern of Posidonia oceanica epibionts along a gradient of ocean acidification: An approach with mimics. Mediterr. Mar. Sci. 2014, 15, 498–509. [Google Scholar] [CrossRef] [Green Version]
- Albano, P.G.; Sabelli, B. The molluscan assemblages inhabiting the leaves and rhizomes of a deep water Posidonia oceanica settlement in the central Tyrrhenian Sea. Sci. Mar. 2012, 76, 721–732. [Google Scholar] [CrossRef] [Green Version]
- Piazzi, L.; Balata, D.; Cinelli, F.; Benedetti-Cecchi, L. Patterns of spatial variability in epiphytes of Posidonia oceanica: Differences between a disturbed and two reference locations. Aquat. Bot. 2004, 79, 345–356. [Google Scholar] [CrossRef]
- Mecca, S.; Casoli, E.; Ardizzone, G.; Gambi, M.C. Effects of ocean acidification on phenology and epiphytes of the seagrass Posidonia oceanica at two CO2 vent systems of Ischia (Italy). Mediterr. Mar. Sci. 2020, 21, 70–83. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- McArdle, B.H.; Anderson, M.J. Fitting multivariate models to community data: A comment on distance-based redundancy analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef]
- Free Software Foundation The R Project for Statistical Computing. Available online: http://www.r-project.org (accessed on 10 June 2020).
- Marzialetti, S.; Nicoletti, L.; Ardizzone, G.D. The polychaete community of the Fregene artificial reef (Tyrrhenian Sea, Italy): A 20-year study (1981–2001). Zoosymposia 2009, 2, 551–566. [Google Scholar] [CrossRef]
- Giangrande, A. Trophic structure changes of a polychaete community along a vertical cliff. In Rapport Commission Internationale pour L’exploration Scientifique de la Mer Méditerranée; CIESM: Villa Girasole, Monaco, 1986; Volume 30, p. 252. [Google Scholar]
- Mikac, B.; Licciano, M.; Jaklin, A.; Iveša, L.; Giangrande, A.; Musco, L. Diversity and distribution patterns of hard bottom polychaete assemblages in the north adriatic sea (Mediterranean). Diversity 2020, 12, 408. [Google Scholar] [CrossRef]
- Sanfilippo, R.; Rosso, A.; Sciuto, F.; Serio, D.; Catra, M.; Alongi, G.; Negri, M.P.; Leonardi, R.; Viola, A. Serpulid polychaetes from Cystoseira communities in the Ionian Sea, Mediterranean. Vie Milieu 2017, 67, 217–226. [Google Scholar]
- Mabrouk, L.; Ben Brahim, M.; Hamza, A.; Bradai, M.N. Temporal and spatial zonation of macroepiphytes on Posidonia oceanica (L.) Delile leaves in a meadow off Tunisia. Mar. Ecol. 2015, 36, 77–92. [Google Scholar] [CrossRef]
- Ippolitov, A.P.; Rzhavsky, A.V. Tube morphology, ultrastructures and mineralogy in recent Spirorbinae (Annelida: Polychaeta: Serpulidae). III. Tribe Circeini. Invertebr. Zool. 2015, 12, 151–173. [Google Scholar] [CrossRef]
- Sanfilippo, R. Micromorphology, microstructure and functional morphology of the Josephella marenzelleri (Polychaeta Serpulidae) tube. In Autoecology of Selected Organisms: Achievements and Problems; Bollettino della Società Paleontologica Italiana: Milan, Italy, 1996; Volume 3, pp. 205–211. [Google Scholar]
- Boero, F.; De Leo, F.; Fraschetti, S.; Ingrosso, G. The Cells of Ecosystem Functioning: Towards a Holistic Vision of Marine Space, 1st ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2019; Volume 82. [Google Scholar]
- Chao, A.; Ma, K.H.; Hsieh, T.C. iNEXT (iNterpolation and EXTrapolation) Online: Software for Interpolation and Extrapolation of Species Diversity. Program and User’s Guide. 2016. Available online: http://chao.stat.nthu.edu.tw/wordpress/software_download/ (accessed on 28 May 2021).




| Serpulid Assemblages on P. oceanica Subhabitats | |||||
|---|---|---|---|---|---|
| Source | Df | SS | R2 | F | p |
| Habitat | 1 | 6.1885 | 0.5350 | 50.946 | 0.001 |
| Site | 1 | 0.2829 | 0.0267 | 2.329 | 0.094 |
| Habitat:Site | 1 | 0.2474 | 0.2333 | 2.037 | 0.117 |
| Residuals | 32 | 3.8871 | 0.3665 | ||
| Total | 35 | 10.6059 | 1.0000 | ||
| Pairwise Comparison (All Habitats, N Sites) | |||||
| Pairs | p | p adj | |||
| P. crispa mat | P. oceanica leaf | 0.001 | 0.003 | ||
| P. crispa mat | P. oceanica shoot | 0.001 | 0.003 | ||
| P. oceanica leaf | P. oceanica shoot | 0.001 | 0.003 | ||
| Serpulid Assemblages on P. crispa Mats | |||||
|---|---|---|---|---|---|
| Source | Df | SS | R2 | F | p |
| Site | 3 | 0.9182 | 0.4066 | 2.7407 | 0.012 |
| Residual | 12 | 1.3401 | 0.5934 | ||
| Total | 15 | 2.2583 | 1.0000 | ||
| Pairwise Comparison (P. crispa Sites) | |||||
| Pairs | p | p adj. | |||
| Mix | PC1 | 0.444 | 1.000 | ||
| Mix | PC2 | 0.032 | 0.192 | ||
| Mix | PC3 | 0.033 | 0.198 | ||
| PC1 | PC2 | 0.351 | 1.000 | ||
| PC1 | PC3 | 0.104 | 0.624 | ||
| PC2 | PC3 | 0.062 | 0.372 | ||
| Species | Shallow Infralittoral (1) | P. crispa Mat (1),(2) | Coralligènous Habitat (2) | P. oceanica Meadow (2) | |
|---|---|---|---|---|---|
| Bathyvermilia sp. | x | ||||
| Filograna implexa (Berkeley) | x | x | |||
| Hydroides spp. (Gunnerus) | x | x | x | x | x |
| Janita fimbriata (Delle Chiaje) | x | ||||
| Janua sp. | x | x | x | x | |
| Josephella marenzelleri (Caullery and Mesnil) | x | x | x | x | x |
| Metavermilia multicristata (Philippi) | x | ||||
| Pileolaria spp. | x | x | x | x | x |
| Protula sp. (Risso) | x | ||||
| Semivermilia crenata (O. G. Costa) | x | x | |||
| Semivermilia cribrata (O. G. Costa) | x | ||||
| Serpula spp. | x | x | x | x | x |
| Serpula vermicularis (Linnaeus) | x | x | |||
| Spiraserpula massiliensis (Zibrowius) | x | ||||
| Spirobranchus spp. | x | x | x | x | x |
| Spirorbis spp. | x | x | x | ||
| Vermiliopsis spp. | x | x | x | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossbach, F.I.; Casoli, E.; Beck, M.; Wild, C. Mediterranean Red Macro Algae Mats as Habitat for High Abundances of Serpulid Polychaetes. Diversity 2021, 13, 265. https://doi.org/10.3390/d13060265
Rossbach FI, Casoli E, Beck M, Wild C. Mediterranean Red Macro Algae Mats as Habitat for High Abundances of Serpulid Polychaetes. Diversity. 2021; 13(6):265. https://doi.org/10.3390/d13060265
Chicago/Turabian StyleRossbach, Felix I., Edoardo Casoli, Milan Beck, and Christian Wild. 2021. "Mediterranean Red Macro Algae Mats as Habitat for High Abundances of Serpulid Polychaetes" Diversity 13, no. 6: 265. https://doi.org/10.3390/d13060265
APA StyleRossbach, F. I., Casoli, E., Beck, M., & Wild, C. (2021). Mediterranean Red Macro Algae Mats as Habitat for High Abundances of Serpulid Polychaetes. Diversity, 13(6), 265. https://doi.org/10.3390/d13060265