
Spatial Distribution Patterns of Appendicularians in the Drake Passage: Potential Indicators of Water Masses?


Abstract
:1. Introduction
2. Material and Methods
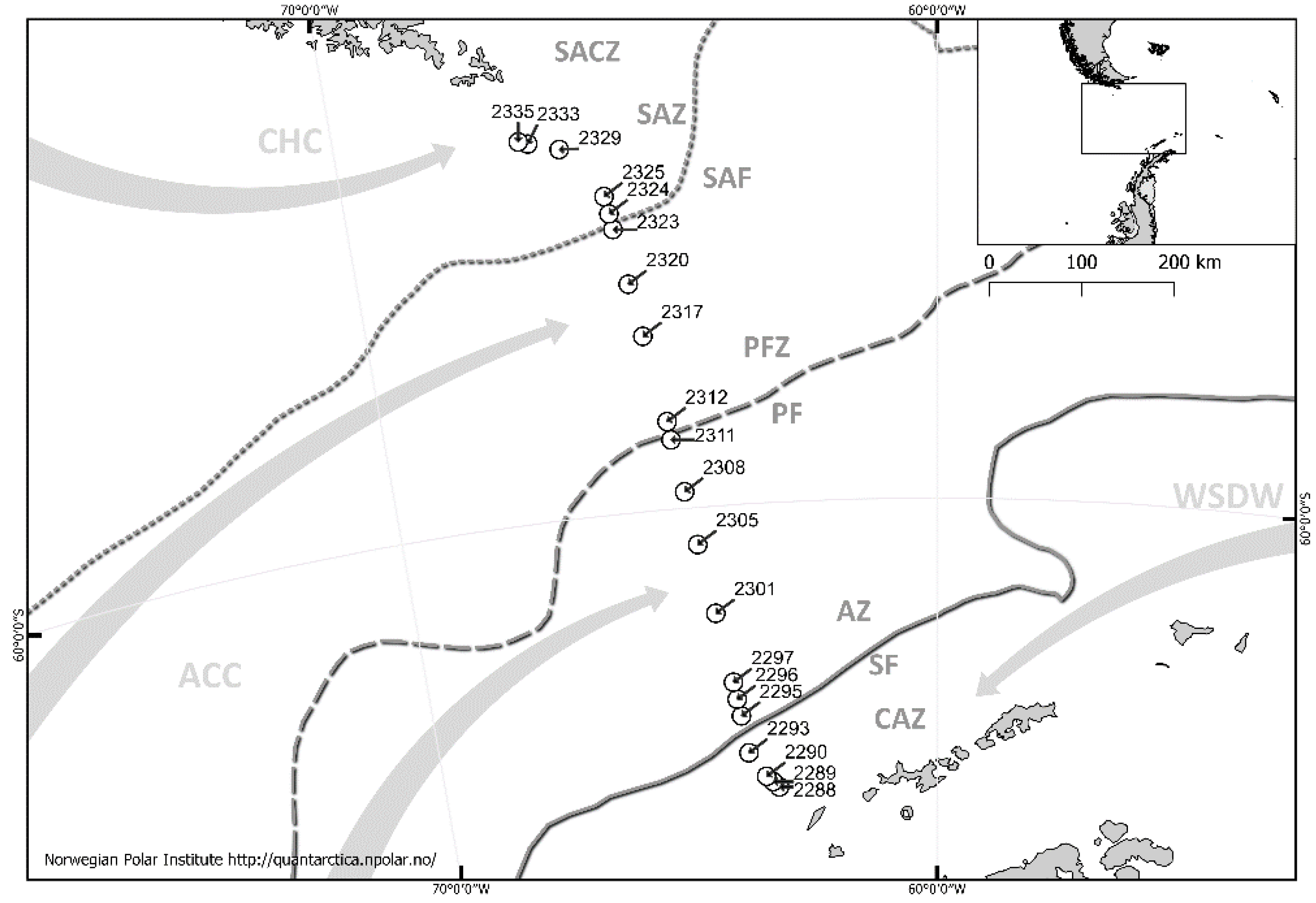
2.1. Study Area
2.2. Sampling Collection and Laboratory Studies
2.3. Statistical Analyses
3. Results
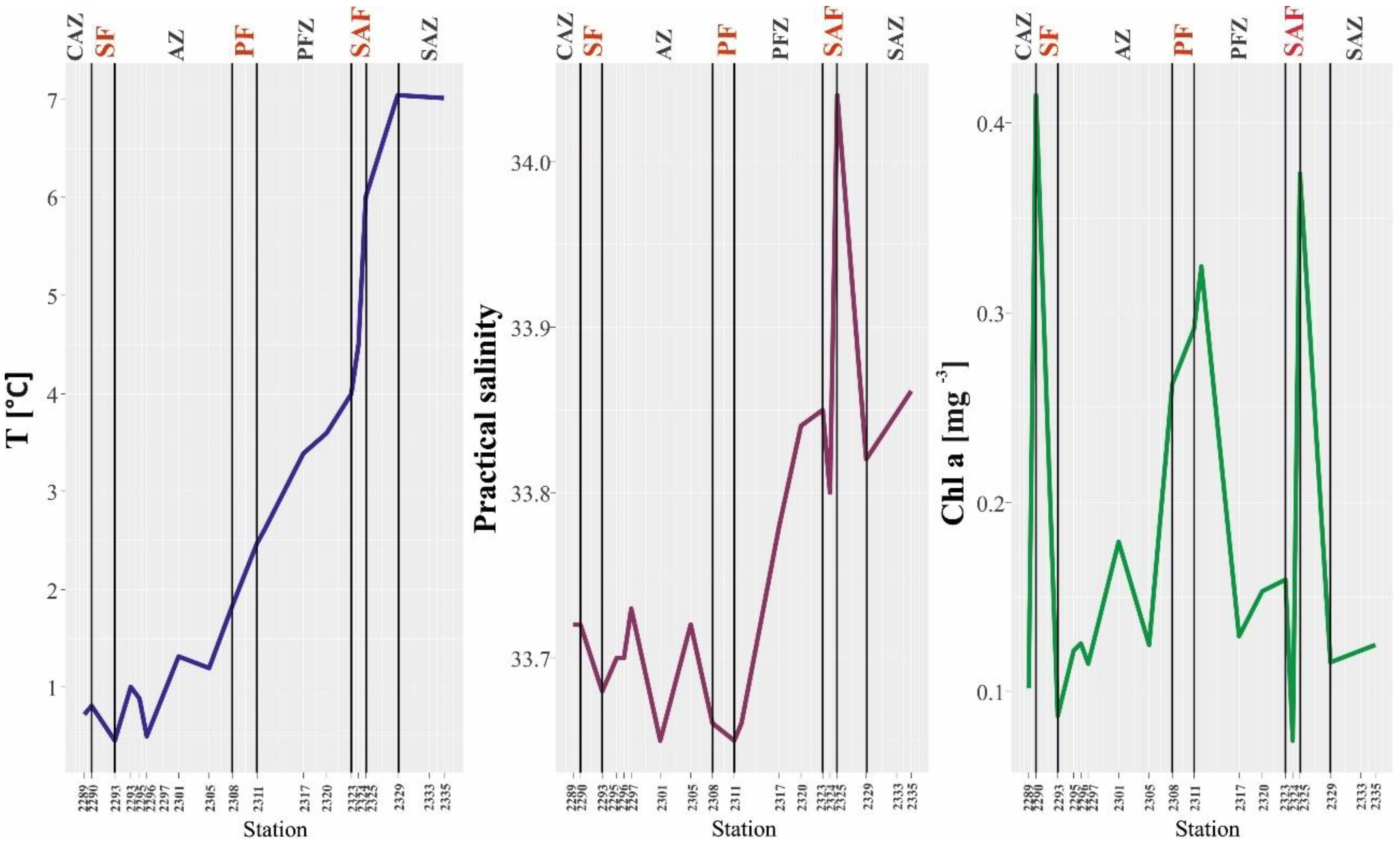
3.1. Hydrological Conditions
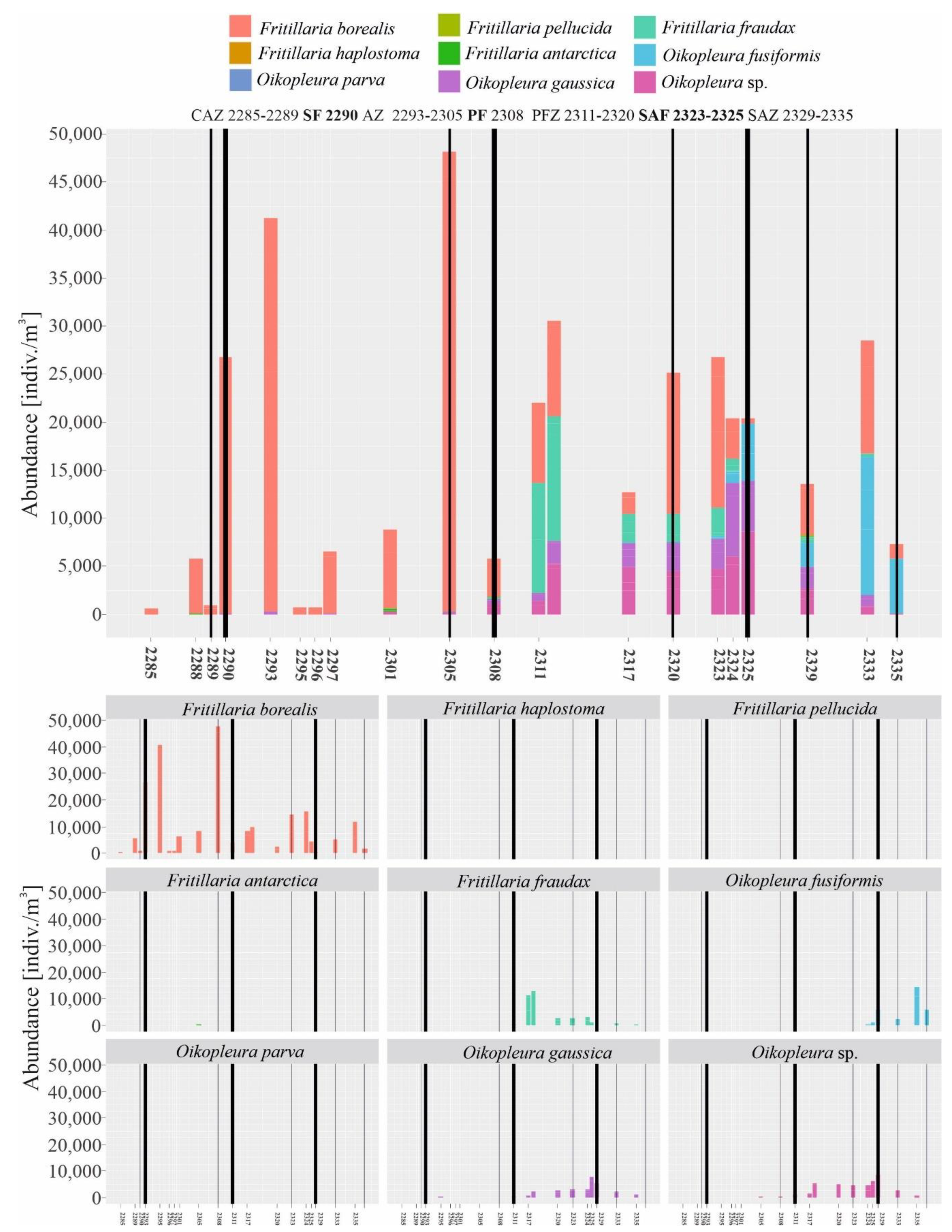
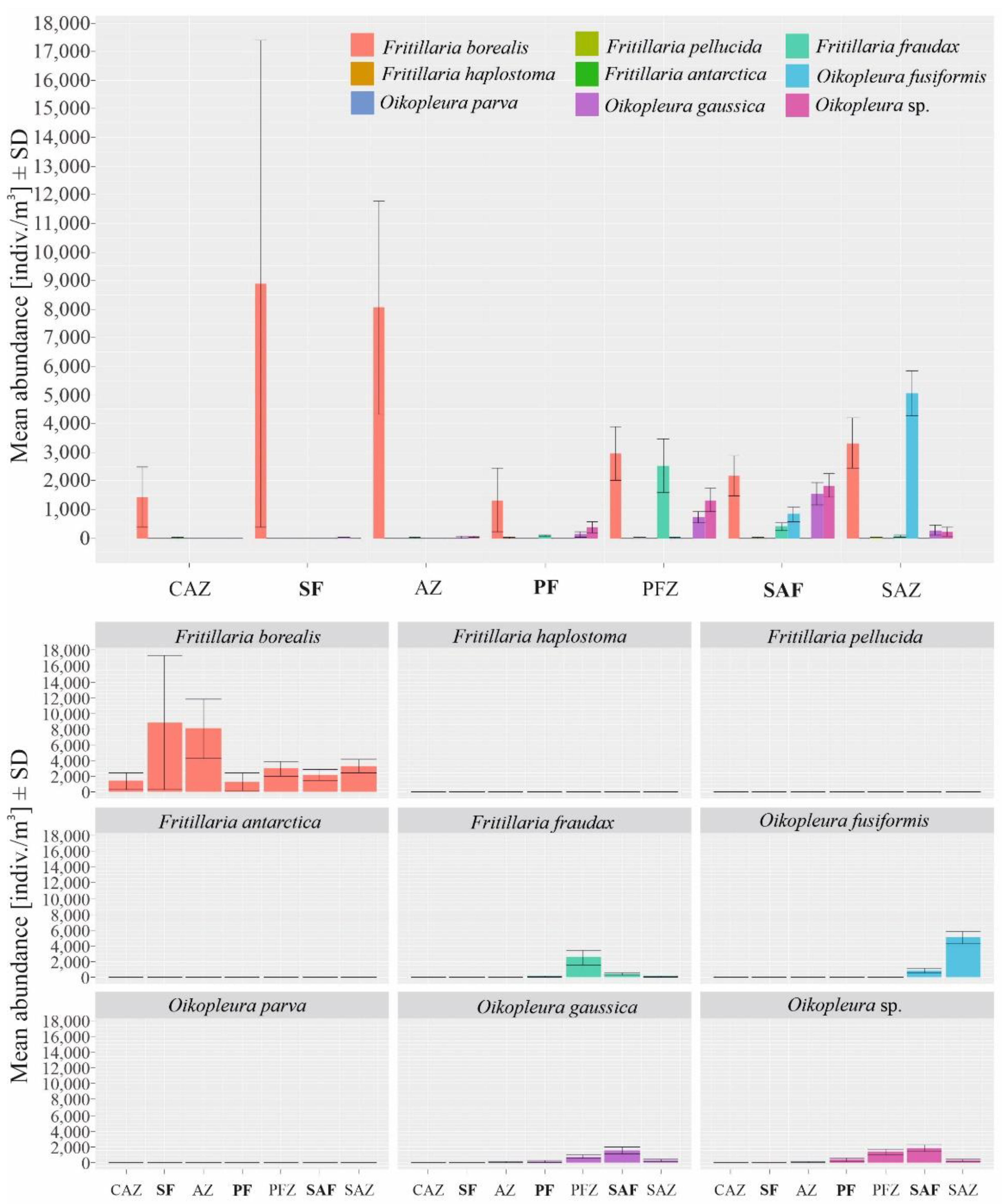
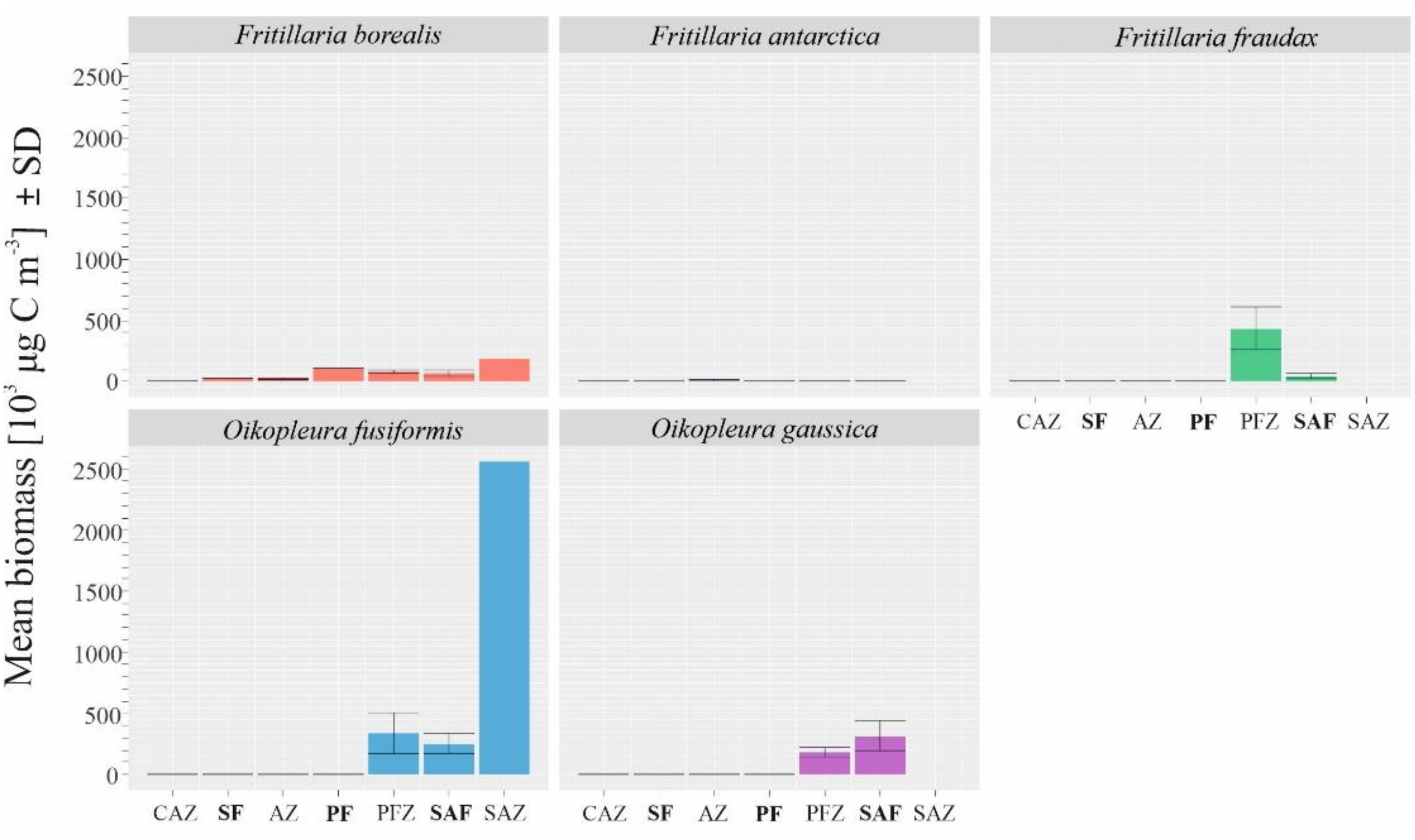
3.2. Biogeographical Distribution of Appendicularia in the Drake Passage
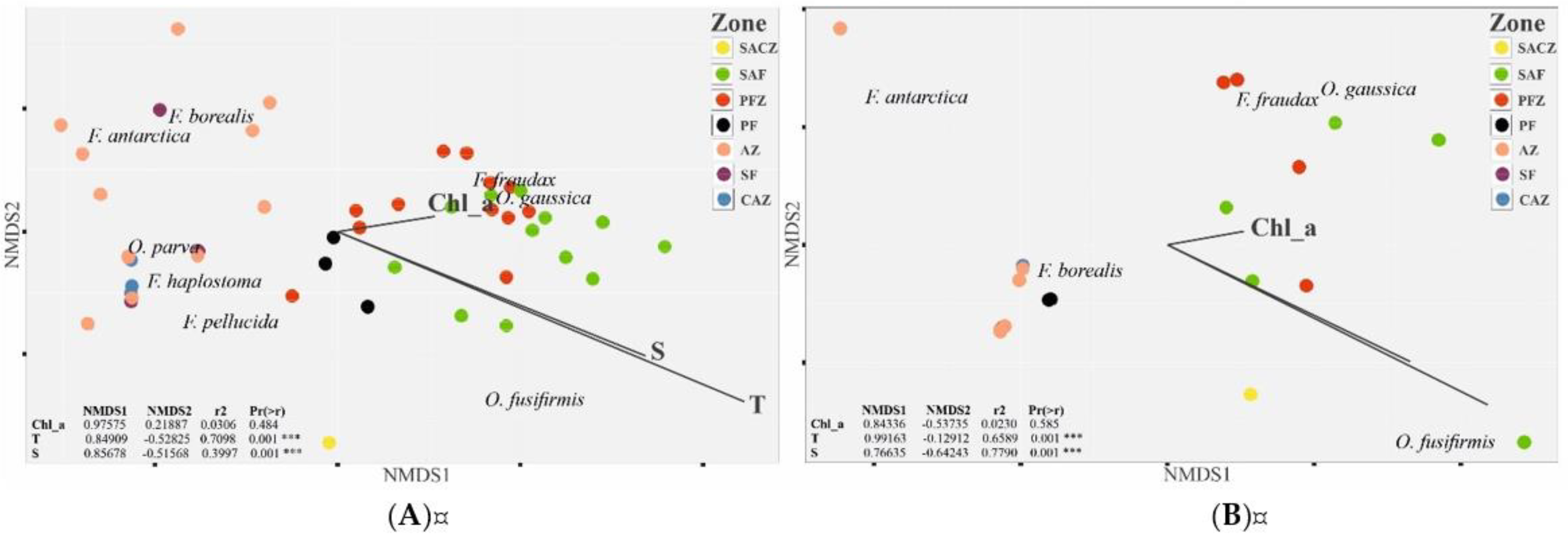
3.3. Environmental Factors Influencing the Zonal Distribution of Appendicularia
4. Discussion
4.1. Biogeographical Distribution of Appendicularia in the Drake Passage and the Main Zones of Occurrence
4.2. Environmental Factors Influencing the Zonal Distribution of Appendicullaria
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Esnal, G. Apendicularia. In Atlas del Zooplancton del Atlántico Sudoccidental y Métodosde Trabajo Con el Zooplancton Marino; Boltovskoy, D., Ed.; Publicación Especial; Instituto Nacional de Investigacióny Desarrollo Pesquero: Mar del Plata, Argentina, 1981; pp. 809–820. [Google Scholar]
- Aravena, G.; Palma, S. Taxonomic identification of appendicularians collected in the epipelagic waters off northern Chile (Tunicata, Appendicularia). Rev. Chil. Hist. Nat. 2002, 75, 307–325. [Google Scholar] [CrossRef]
- Tsujimoto, M.; Takahashi, K.T.; Hirawake, T.; Mitsuo, F. Unusual abundance of appendicularians in the seasonal ice zone (140°E) of the Southern Ocean. Polar Biosci. 2006, 19, 133–141. [Google Scholar]
- Esnal, G. Appendicularia. In South Atlantic Zooplankton; Boltovskoy, D., Ed.; Backhuys Publication: Leiden, The Netherlands, 1999; pp. 1375–1399. [Google Scholar]
- Sato, R.; Tanaka, Y.; Ishimaru, T. House Production by Oikopleura dioica (Tunicata, Appendicularia) Under Laboratory Conditions. J. Plankton Res. 2001, 23, 415–423. [Google Scholar] [CrossRef] [Green Version]
- Deibel, D.; Lee, S.H. Retention efficiency of sub-micrometer particles by the pharyngeal filter of the pelagic tunicate Oikopleura vanhoeffeni. Mar. Ecol. Prog. Ser. 1992, 81, 25–30. [Google Scholar] [CrossRef]
- Acuña, J.L.; Deibel, D.; Morris, C.C. Particle capture mechanism of the pelagic tunicate Oikopleura vanhoeffeni. Limnol. Oceanogr. 1996, 41, 1800–1814. [Google Scholar] [CrossRef]
- Vargas, C.A.; Tönnesson, K.; Sell, A.; Maar, M.; Friis Møller, E.; Zervoudaki, T.; Giannakourou, A.; Christou, E.; Satapoomin, S.; Kjerulf Petersen, J.; et al. Importance of copepods versus appendicularians in vertical carbon fluxes in a Swedish fjord. Mar. Ecol. Prog. Ser. 2002, 241, 125–138. [Google Scholar] [CrossRef] [Green Version]
- Hansen, J.L.S.; Kiørboe, T.; Alldredge, A.L. Marine snow derived from abandoned larvacean houses: Sinking rates, particle content and mechanisms of aggregate formation. Mar. Ecol. Prog. Ser. 1996, 141, 205–215. [Google Scholar] [CrossRef] [Green Version]
- Hopcroft, R.R.; Roff, J.C.; Bauman, H.A. Zooplankton growth rates: The larvaceans Appendicularia, Fritillaria and Oikopleura in tropical waters. J. Plankton Res. 1998, 20, 539–555. [Google Scholar] [CrossRef] [Green Version]
- Capitanio, F.; Pajaro, M.; Esnal, G. Appendicularians (Chordata, Tunicata) in the diet of anchovy (Engraulis anchoita) in the Argentine sea. Sci. Mar. 1997, 61, 9–15. [Google Scholar]
- Atkinson, A.; Ward, P.; Hunt, B.P.V.; Pakhomov, E.A.; Hosie, G.W. An Overview of Southern Ocean zooplankton data: Abundance, biomass, feeding and functional relationship. CCAMLR Sci. 2012, 19, 171–218. [Google Scholar]
- Spinelli, M.L.; Franzosi, C.; Olguin Salinas, H.; Capitanio, F.L.; Alder, V.A. Appendicularians and copepods from Scotia Bay (Laurie island, South Orkney, Antarctica): Fluctuations in community structure and diversity in two contrasting, consecutive summers. Pol. Biol. 2018, 41, 663–678. [Google Scholar] [CrossRef]
- Hunt, B.P.V.; Hosie, G.W. Zonal structure of zooplankton communities in the Southern Ocean South of Australia: Results from a 2150 km continuous plankton recorder transect. Deep Sea Res. I 2005, 52, 1241–1271. [Google Scholar] [CrossRef]
- Capitanio, F.L.; Daponte, M.C.; Esnal, G.B. The classification of Antarctic appendicularians: The Oikopleura gaussica group. Ant. Sci. 2003, 15, 476–482. [Google Scholar] [CrossRef]
- Chiba, S.; Ishimaru, T.; Hosie, G.W.; Fukuchi, M. Spatio-temporal variability of zooplankton community structure off east Antarctica (90 to 160°E). Mar. Ecol. Prog. Ser. 2001, 216, 95–108. [Google Scholar] [CrossRef] [Green Version]
- Ward, P.; Shreeve, R.; Whitehouse, M.; Korb, B.; Atkinson, A.; Meredith, M.; Pond, D.; Watkins, J.; Goss, C.; Cunningham, N. Phyto- and zooplankton community structure and production around South Georgia (Southern Ocean) during summer 2001/02. Deep Sea Res. I 2005, 52, 421–441. [Google Scholar] [CrossRef]
- McLeod, D.J.; Hosie, G.W.; Kitchener, J.A.; Takahashi, K.T.; Hunt, B.P.V. Zooplankton atlas of the Southern Ocean: The SCAR SO-CPR survey (1991–2008). Polar Sci. 2010, 4, 353–385. [Google Scholar] [CrossRef] [Green Version]
- Aguirre, G.E.; Capitanio, F.L.; Lovrich, G.A.; Esnal, G.B. Seasonal variability of metazooplankton in coastal sub-Antarctic waters (Beagle Channel). Mar. Biol. Res. 2012, 8, 341–353. [Google Scholar] [CrossRef]
- Turner, J.; Barrand, N.E.; Bracegirdle, T.J.; Convey, P.; Hodgson, D.; Jarvis, M.; Jenkins, A.; Marshall, G.; Meredith, M.P.; Roscoe, H.; et al. Antarctic climate change and the environment: An update. Polar Rec. 2014, 50, 237–259. [Google Scholar] [CrossRef] [Green Version]
- Convey, P.; Peck, L.S. Antarctic environmental change and biological responses. Sci. Adv. 2019, 5, eaaz0888. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.R.; Raymond, B.; Bracegirdle, T.J.; Chadès, I.; Fuller, R.A.; Shaw, J.D.; Terauds, A. Climate change drives expansion of Antarctic ice-free habitat. Nature 2017, 547, 49–54. [Google Scholar] [CrossRef]
- Kobusińska, M.E.; Lewandowski, K.K.; Panasiuk, A.; Łęczyński, L.; Urbaniak, M.; Ossowski, T.; Niemirycz, E. Precursors of polychlorinated dibenzo-p-dioxins and dibenzofurans in Arctic and Antarctic marine sediments: Environmental concern in the face of climate change. Chemosphere 2020, 260, 127605. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, A.; Siegel, V.; Pakhomov, E.; Rothery, P. Long-term decline in krill stock and increase in salps within the Southern Ocean. Nature 2004, 432, 100–103. [Google Scholar] [CrossRef]
- Peck, L.S. Antarctic marine biodiversity: Adaptations environments and responses to change. Oceanogr. Mar. Biol. 2018, 56, 105–236. [Google Scholar]
- Henley, S.; Schofield, O.; Hendry, K.; Schloss, I.; Steinberg, D.; Moffat, C.; Peck, L.; Costa, D.; Bakker, D.; Hughes, C.; et al. Variability and change in the west Antarctic Peninsula marine system: Research priorities and opportunities. Prog. Oceanogr. 2019, 173, 208–237. [Google Scholar] [CrossRef] [Green Version]
- Potocka, M.; Kidawa, A.; Panasiuk, A.; Bielecka, L.; Wawrzynek-Borejko, J.; Patuła, W.; Wójcik, K.A.; Plenzler, J.; Janecki, T.; Bialik, R.J. The effect of glacier recession on benthic and pelagic communities: Case study in Herve cove, Antarctica. J. Mar. Sci. Eng. 2019, 7, jmse7090285. [Google Scholar] [CrossRef] [Green Version]
- Panasiuk, A.; Wawrzynek-Borejko, J.; Musiał, A.; Korczak-Abshire, M. Pygoscelis penguin diets on King George Island, South Shetland Islands, with a special focus on the krill Euphausia Superba. Ant. Sci. 2020, 32, 21–28. [Google Scholar] [CrossRef]
- Wirtz, K.W. Who is eating whom? Morphology and feeding type determine the size relation between planktonic predators and their ideal prey. Mar. Ecol. Prog. Ser. 2012, 445, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Garcia, M.D.; Dutto, M.S.; Chazarreta, C.J.; Berasategui, A.A.; Schloss, I.R.; Hoffmeyer, M.S. Micro- and mesozooplankton successions in an Antarctic coastal environment during a warm year. PLoS ONE 2020, 15, e0232614. [Google Scholar] [CrossRef] [PubMed]
- Lebrato, M.; de Jesus Mendes, P.; Steinberg, D.K.; Cartes, J.E.; Jones, B.M.; Birsa, L.M.; Benavides, R.; Oschlies, A. Jelly biomass sinking speed reveals a fast carbon export mechanism. Limnol. Oceanogr. 2013, 58, 1113–1122. [Google Scholar] [CrossRef] [Green Version]
- Winder, M.; Bouquet, J.-M.; Bermúdez, J.R.; Berger, S.A.; Hansen, T.; Brandes, J.; Sazhin, A.F.; Nejstgaard, J.C.; Båmstedt, U.; Jakobsen, H.H.; et al. Increased appendicularian zooplankton alter carbon cycling under warmer more acidified ocean conditions. Limnol. Oceanogr. 2017, 62, 1541–1551. [Google Scholar] [CrossRef]
- Richardson, A.J.; Bakun, A.; Hays, G.C.; Gibbons, M.J. The jellyfish joyride: Causes, consequences and management responses to a more gelatinous future. Trends Ecol. Evol. 2009, 24, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Troedsson, C.; Bouquet, J.-M.; Lobon, C.M.; Novac, A.; Nejstgaard, J.C.; Dupont, S.; Bosak, S.; Jakobsen, H.H.; Romanova, N.; Pankoke, L.M.; et al. Effects of ocean acidification, temperature and nutrient regimes on the appendicularian Oikopleura dioica: A mesocosm study. Mar. Biol. 2012, 160, 2175–2187. [Google Scholar] [CrossRef]
- Deacon, G.E.R. A general account of the hydrology of the South Atlantic Ocean. Discov. Rep. 1933, VII, 177–238. [Google Scholar]
- Toggweiler, J.R.; Samuels, B. Effect of Drake Passage on the global thermohaline circulation. Deep Sea Res. I 1995, 42, 477–500. [Google Scholar] [CrossRef]
- Cox, M.D. An idealized model of the world ocean. Part I: The global-scale water masses. J. Phys. Oceanogr. 1989, 19, 1730–1752. [Google Scholar] [CrossRef] [Green Version]
- Whitworth, T., III.; Peterson, R.G. Volume transport of the Antarctic Circumpolar Current from bottom pressure measurements. J. Phys. Oceanogr. 1985, 15, 810–816. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, S.A.; Alderson, S.G.; King, B.A. Transport and variability of the Antarctic Current in Drake Passage. J. Geophys. Res. 2003, 108, 80–84. [Google Scholar] [CrossRef] [Green Version]
- Holm-Hansen, O.; Mitchell, B.G. Spatial and temporal distribution of phytoplankton and primary production in the western Bransficld Strait region. Deep Sea Res. 1991, 38, 961–9811. [Google Scholar] [CrossRef]
- Demidov, A.B.; Gagarin, V.I.; Grigoriev, A.V. Seasonal Variability of the Surface Chlorophyll a in the Drake Passage. Oceanology 2010, 50, 327–341. [Google Scholar] [CrossRef]
- Demidov, A.B.; Mosharov, S.A.; Gagarin, I. Phytoplankton Production Characteristics in the Southern Atlantic and the Atlantic Sector of the Southern Ocean in the Astral Summer of 2009–2010. Oceanology 2012, 52, 206–218. [Google Scholar] [CrossRef]
- Fenaux, R. The Classification of Appendicularia (Tunicata): History and Current State. Mem. L’Institut Oceanogr. 1993, 17, 1–123. [Google Scholar]
- O’Sullivan, D. A Guide to the Pelagic Tunicates of the Southern Ocean and Adjacent Waters; ANARE Research Notes 8; Antarctic Division: Kingston, Australia, 1983. [Google Scholar]
- Capitanio, F.L.; Curelovich, J.; Tresguerres, M.; Negri, R.M.; Viñas, M.D.; Esnal, G.B. Seasonal cycle of Appendicularians at a coastal station (38°28′ S, 57°41′ W) of the SW Atlantic Ocean. Bull. Mar. Sci. 2008, 82, 171–184. [Google Scholar]
- Sato, R.; Tanaka, Y.; Ishimaru, T. Species-specific house productivity of appendicularians. Mar. Ecol. Prog. Ser. 2003, 259, 163–172. [Google Scholar] [CrossRef]
- Uye, S. Length-weight relationship of important zooplankton from the Inland Sea of Japan. J. Oceanogr. 1982, 38, 149–158. [Google Scholar] [CrossRef]
- Deibel, D. Feeding mechanism and house of the appendicularian. Oikopleura Vanhoeffeni. Mar. Biol. 1986, 93, 429–436. [Google Scholar] [CrossRef]
- Fenaux, R. Anatomy and functional morphology of the Appendicularia. In The Biology of Pelagic Tunicates; Bone, Q., Ed.; Oxford University Press: New York, NY, USA, 1998; pp. 25–34. [Google Scholar]
- Venables, W.N.; Smith, D.M.; R Core Team. An Introduction to R, Notes on R: A Programming Environment for Data Analysis and GraphicsVersion 4.1.0. Available online: https://cran.r-project.org/doc/manuals/r-release/R-intro.pdf (accessed on 27 May 2021).
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Henry, M.; Stevens, H. The vegan package. Community Ecol. Package 2013, 10, 631–637. [Google Scholar]
- Jażdżewski, K.; Kittel, W.; Łotocki, K. Zooplankton studies in the southern Drake Passage and in the Bransfield Strait during the austral summer (BIOMAS-FIBEX, February-March 1981). Pol. Polar Res. 1982, 3, 203–242. [Google Scholar]
- Witek, Z.; Kittel, W.; Czykieta, H.; Żmijewska, M.I.; Presler, E. Macrozooplankton in the southern Drake Passage and in Bransfield Strait during BIOMASS-SIBEX (December 1983–January 1984). Pol. Polar Res. 1985, 6, 95–115. [Google Scholar]
- Tokioka, T. Taxonomic studies of Appendicularians collected by the Japanese Antarctic Research Expedition, 1957. Sci. Rep. Jap. Antarct. Res. Exped. 1964, 21, 1–16. [Google Scholar]
- Lindsay, M.; Williams, G. Distribution and abundance of Larvaceans in the Southern Ocean between 30 and 80°E. Deep Sea Res. II 2010, 57, 905–915. [Google Scholar] [CrossRef]
- Troedsson, C.; Bouquet, J.-M.; Aksnes, D.L.; Thompson, E.M. Resource allocation between somatic growth and reproductive output in the pelagic chordate Oikopleura dioica allows opportunistic response to nutritional variation. Mar. Ecol. Prog. Ser. 2002, 243, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Gorsky, G.; Palazzoli, I.; Fenaux, R. Influence of temperature changes on oxygen uptake and ammonia and phosphate excretion. In relation to body size and weight, in Oikopleura dioica (Appendicularia). Mar. Biol. 1987, 94, 191–201. [Google Scholar] [CrossRef]
- Pakhomov, E.A.; McQuaid, C.D. Distribution of surface zooplankton and seabirds across the Southern Ocean. Polar Biol. 1996, 16, 271–289. [Google Scholar] [CrossRef]
- Pakhomov, E.A.; Perissinotto, R.; McQuaid, C.D.; Froneman, P.W. Zooplankton structure and grazing in the Atlantic sector of the Southern Ocean in late summer 1993. Part 1. Ecological zonation. Deep Sea Res. I 2000, 47, 1663–1686. [Google Scholar] [CrossRef]
- Hosie, G.W.; Fukuchi, M.; Kawaguchi, S. Development of the Southern Ocean Continuous Plankton Recorder survey. Prog. Oceanogr. 2003, 58, 263–283. [Google Scholar] [CrossRef]
- Takahashi, K.T.; Kawaguchi, S.; Hosie, G.W.; Toda, T.; Naganobu, M.; Fukuchi, M. Surface zooplankton distribution in the Drake Passage recorded by Continuous Plankton Recorder (CPR) in late austral summer of 2000. Polar Sci. 2010, 3, 235–245. [Google Scholar] [CrossRef] [Green Version]
- López Urrutia, A.; Harris, R.P.; Smith, T. Predation by calanoid copepods on the appendicularian Oikopleura dioica. Limnol. Oceanogr. 2004, 49, 303–307. [Google Scholar] [CrossRef]
- Swart, N.C.; Gille, S.T.; Fyfe, J.C.; Gillett, N.P. Recent Southern Ocean warming and freshening driven by greenhouse gas emissions and ozone depletion. Nat. Geosci. 2018, 11, 836–841. [Google Scholar] [CrossRef]
- Trull, T.W.; Passmore, A.; Davies, D.M.; Smith, T.; Berry, K.; Tilbrook, B. Distribution of planktonic biogenic carbonate organisms in the Southern Ocean south of Australia: A baseline for ocean acidification impact assessment. Biogeosciences 2018, 15, 31–49. [Google Scholar] [CrossRef] [Green Version]
- Mańko, M.K.; Panasiuk-Chodnicka, A.A.; Żmijewska, M.I. Pelagic coelenterates in the Atlantic sector of the Arctic Ocean: Species diversity and distribution as water mass indicators. Oceanol. Hydrobiol. Stud. 2015, 44, 466–479. [Google Scholar] [CrossRef]
- Słomska, A.W.; Panasiuk-Chodnicka, A.A.; Żmijewska, M.I.; Mańko, M.K. Variability of Salpa thompsoni population structure in the Drake Passage in summer season 2010. Pol. Polar Res. 2015, 36, 391–404. [Google Scholar] [CrossRef]
- Groeneveld, J.; Berger, U.; Henschke, N.; Pakhomov, E.A.; Reiss, C.S.; Meyer, B. Blooms of a key grazer in the Southern Ocean –An individual-based model of Salpa Thompsoni. Prog. Oceanogr. 2020, 185, 102339. [Google Scholar] [CrossRef]
- Plum, C.; Hillebrand, H.; Moorthi, S. Krill vs salps: Dominance shift from krill to salps is associated with higher dissolved N:P ratios. Sci. Rep. 2020, 10, 5911. [Google Scholar] [CrossRef] [Green Version]
- Panasiuk, A.; Grzonka, L.; Prątnicka, P.; Wawrzynek-Borejko, J.; Szymelfenig, M. Zonal variability of pelagic Siphonophora (Cnidaria) in the atlantic sector of the southern ocean. J. Sea. Res. 2020, 165, 101951. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Source | |
|---|---|---|
| Fritillaria borealis | log DW(µg) = 3.86 log TL(µm)–11.72 | [45] |
| Fritillaria fraudax | log DW(µg) = 3.86 log TL(µm)–11.72 | [45] |
| Oikopleura gaussica | C(µg) = 4.59 TL(mm) | [48] |
| Oikopleura fusiformis | Log DW(µg) = 2.1 log TL(µm)–6.82 | [46] |
| Species | Drake Passage | Zone | |||||||
|---|---|---|---|---|---|---|---|---|---|
| CAZ | SF | AZ | PF | PFZ | SAF | SAZ | SACZ | ||
| Fritillaria borealis | 3177 ± 4514 | 5684 | 8887 | 3418 ± 5242 | 1989 | 1392 ± 1153 | 866 ± 1108 | 433 | 2125 ± 861 |
| Fritillaria haplostoma | 1 ± 5 | 0 | 0 | 0 | 20 | 0 | 0 | 0 | 0 |
| Fritillaria pellucida | 2 ± 7 | 0 | 0 | 0 ± 2 | 0 | 0 | 0 | 0 | 20 ± 10 |
| Fritillaria antarctica | 18 ± 62 | 81 | 0 | 45 ± 111 | 0 | 0 | 0 | 0 | 0 |
| Fritillaria fraudax | 405 ± 880 | 0 | 0 | 0 | 81 | 1490 ± 1561 | 623 ± 551 | 68 | 61 ± 86 |
| Oikopleura fusiformis | 602 ± 1510 | 0 | 0 | 0 | 0 | 7 ± 14 | 821 ± 1180 | 284 | 4635 ± 1656 |
| Oikopleura parva | 1 ± 4 | 0 | 0 | 2 ± 0 | 0 | 3 ± 7 | 0 | 0 | 0 |
| Oikopleura sp | 212 ± 334 | 0 | 24 | 9 ± 42 | 183 | 330 ± 239 | 785 ± 501 | 122 | 41 ± 57 |
| Oikopleura gaussica | 321 ± 484 | 0 | 11 | 0 | 508 | 641 ± 652 | 1011 ± 353 | 135 | 61 ± 48 |
| Appendicularia | 4779 ± 4310 | 5765 | 8911 | 3474 ± 5278 | 2274 | 3223 ± 2523 | 3095 ± 536 | 907 | 6882 ± 641 |
| Species | Biomass (103 μg C m−3) |
|---|---|
| Fritillaria borealis | 73.79 ± 90.35 |
| Fritillaria fraudax | 96.85 ± 308.26 |
| Oikopleura gaussica | 101.69 ± 232.87 |
| Oikopleura fusiformis | 321.28 ± 677.54 |
| Abundance (ind. 10−3 m3) | Biomass (103 μg C m−3) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ef | Df R | App | Fb | Fh | Fp | Fa | Ff | Of | Op | Og | O | Fb | Fa | Of | Og |
| Chl a | 1 45 | 0.607 | 1.2827 | 0.4724 | 0.5911 | 0.0166 | 1.1251 | 1.3576 | 0.4371 | 3.5536 * | 0.2951 | 2.3743 | 0.0193 | 2.4803 | 1.7626 |
| T | 17 29 | 2.0373 *** | 1.1479 | 0.7985 | 2.0905 | 10.319 *** | 8.5419 *** | 6.8241 *** | 0.7509 | 2.5872 *** | 3.9824 *** | −1.7976 × 1017 | 9.156 × 1017 *** | 9.6813 × 1016 | 36.071 *** |
| S | 1 45 | 5.6876 *** | 0.6531 | 0.9379 | 0.0561 | 2.0514 | 2.7163 | 30.529 *** | 0.1458 | 6.4235 *** | 8.2766 *** | 6.1528 ** | 2.4086 | 39.397 *** | 5.5897 ** |
| T * Chl_a | 17 29 | 2.0373 *** | 1.1479 | 0.7985 | 2.0905 | 10.319 *** | 8.5419 *** | 6.8241 *** | 0.7509 | 2.5872 *** | 3.9824 *** | −1.7976 × 1017 | 9.156 × 1017 *** | 9.6813 × 1016 | 36.071 *** |
| T * S | 17 29 | 2.0373 *** | 1.1479 | 0.7985 | 2.0905 | 10.319 *** | 8.5419 *** | 6.8241 *** | 0.7509 | 2.5872 *** | 3.9824 *** | −1.7976 × 1017 | 9.156 × 1017 *** | 9.6813 × 1016 | 36.071 *** |
| Z | 1 45 | 9.9469 *** | 1.8119 | 0.0213 | 2.6486 | 1.0618 | 26.043 *** | 27.878 *** | 0.0906 | 16.678 *** | 30.08 *** | 24.818 *** | 1.2421 | 32.616 *** | 25.034 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalarus, M.; Panasiuk, A. Spatial Distribution Patterns of Appendicularians in the Drake Passage: Potential Indicators of Water Masses? Diversity 2021, 13, 286. https://doi.org/10.3390/d13070286
Kalarus M, Panasiuk A. Spatial Distribution Patterns of Appendicularians in the Drake Passage: Potential Indicators of Water Masses? Diversity. 2021; 13(7):286. https://doi.org/10.3390/d13070286
Chicago/Turabian StyleKalarus, Marcin, and Anna Panasiuk. 2021. "Spatial Distribution Patterns of Appendicularians in the Drake Passage: Potential Indicators of Water Masses?" Diversity 13, no. 7: 286. https://doi.org/10.3390/d13070286
APA StyleKalarus, M., & Panasiuk, A. (2021). Spatial Distribution Patterns of Appendicularians in the Drake Passage: Potential Indicators of Water Masses? Diversity, 13(7), 286. https://doi.org/10.3390/d13070286