
Coverage Increases of Porites astreoides in Grenada Determined by Shifts in Size-Frequency Distribution

 , ,
, ,
Abstract
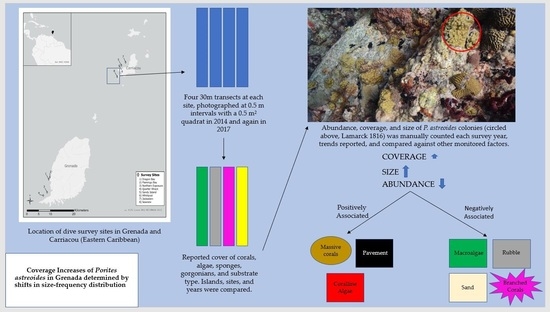
:
1. Introduction
2. Materials and Methods
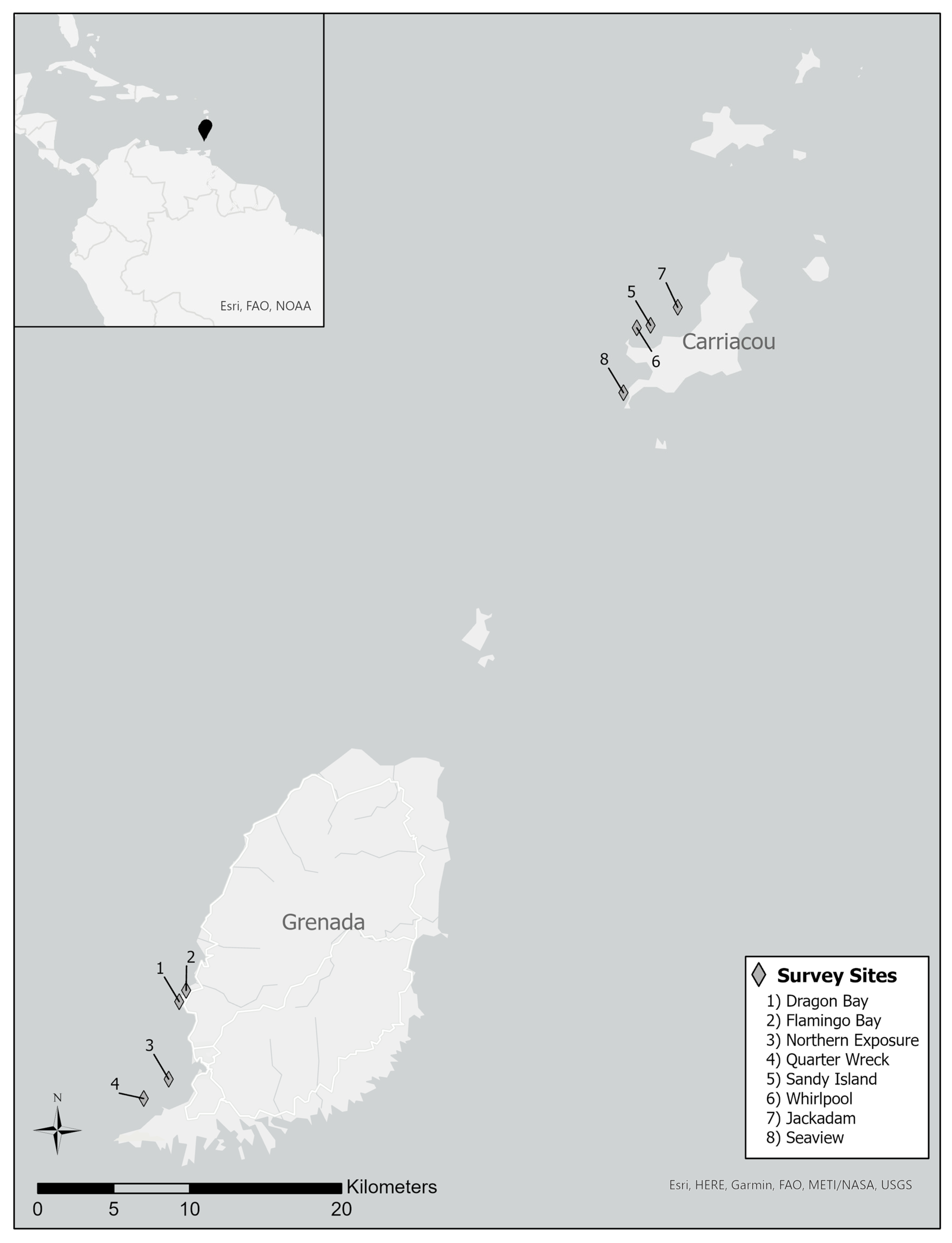
2.1. Study Sites
2.2. Benthic Community Assessment
2.3. P. astreoides Population Dynamics
3. Results
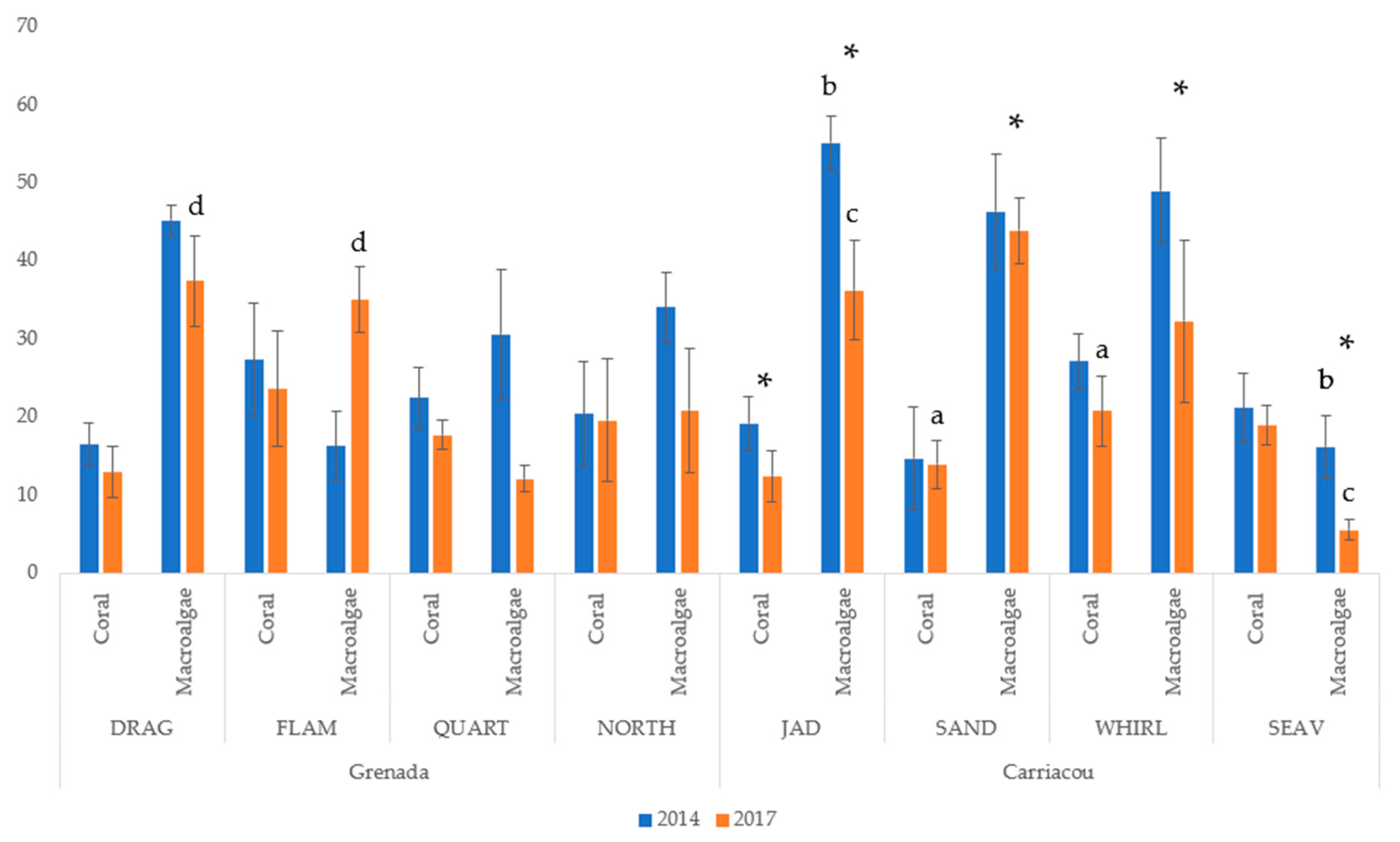
3.1. Benthic Community Assessment
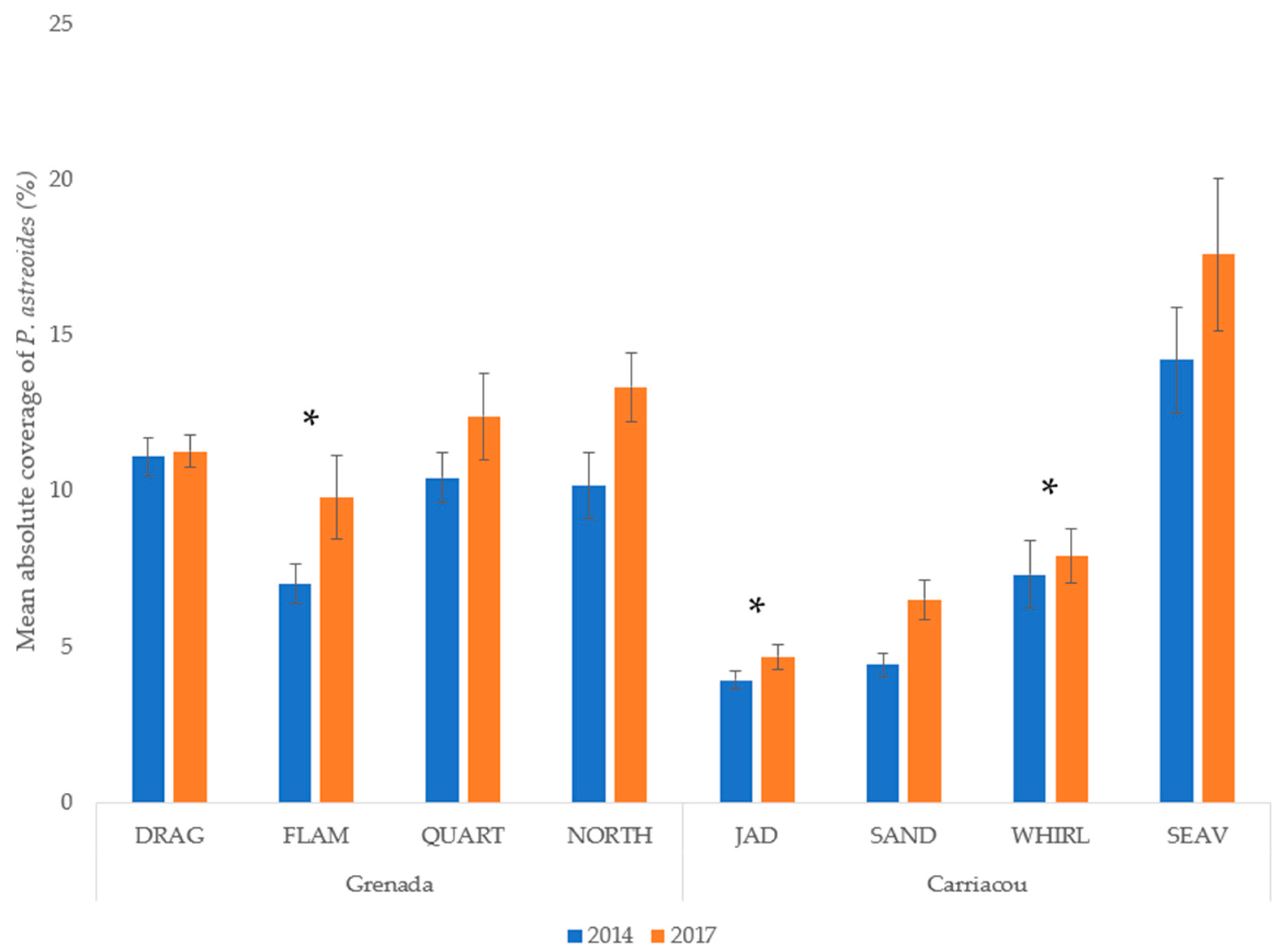
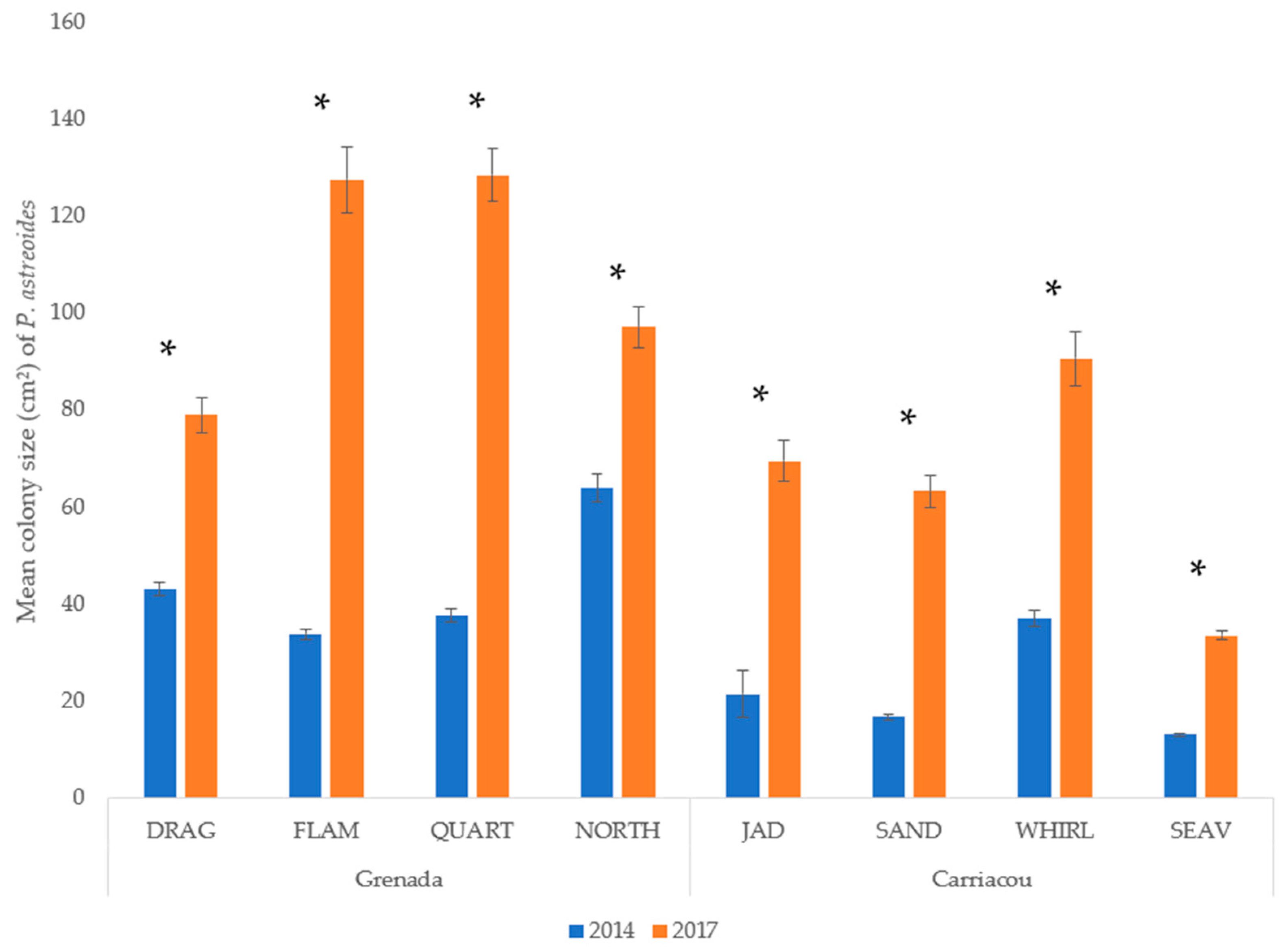
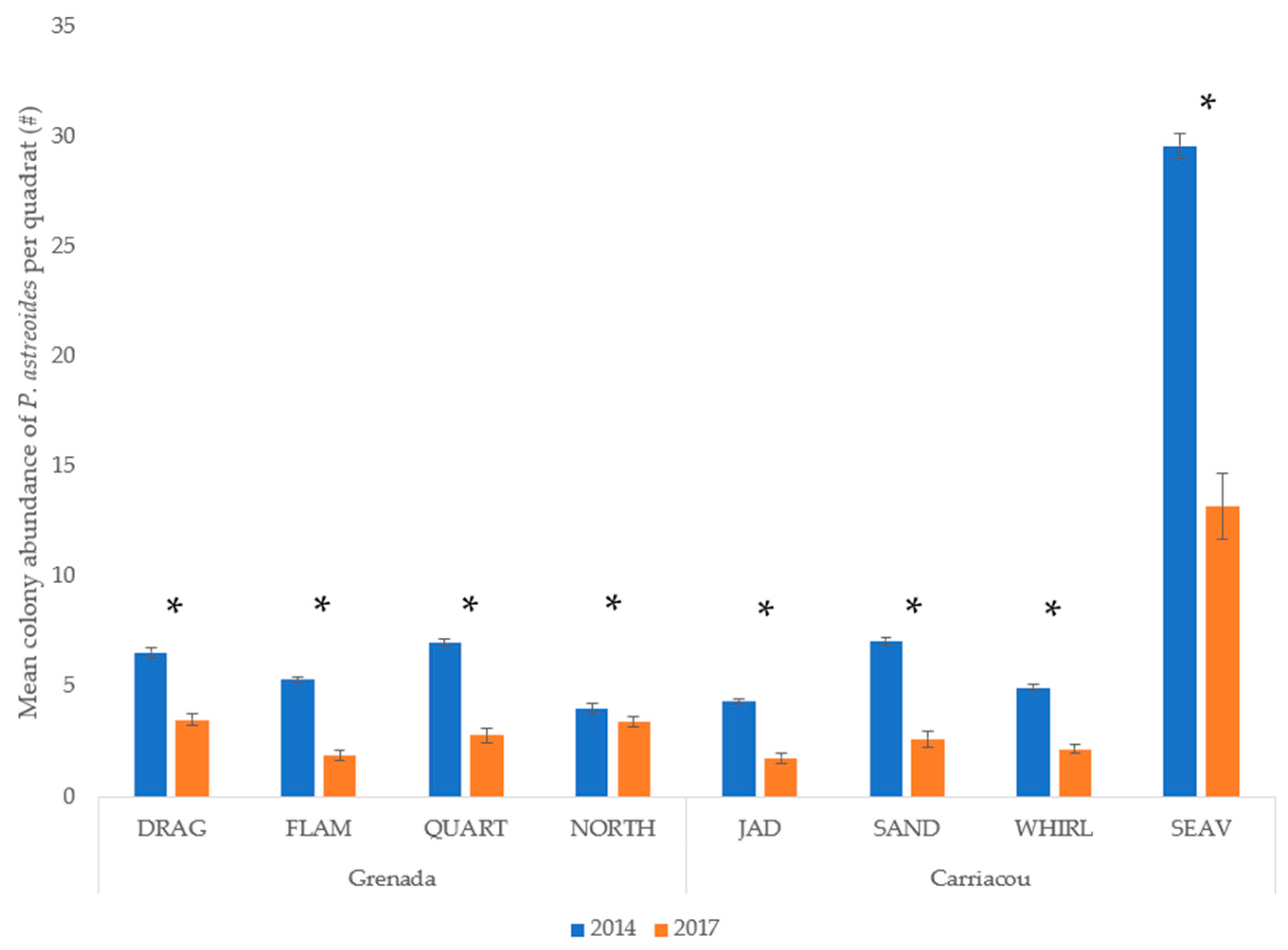
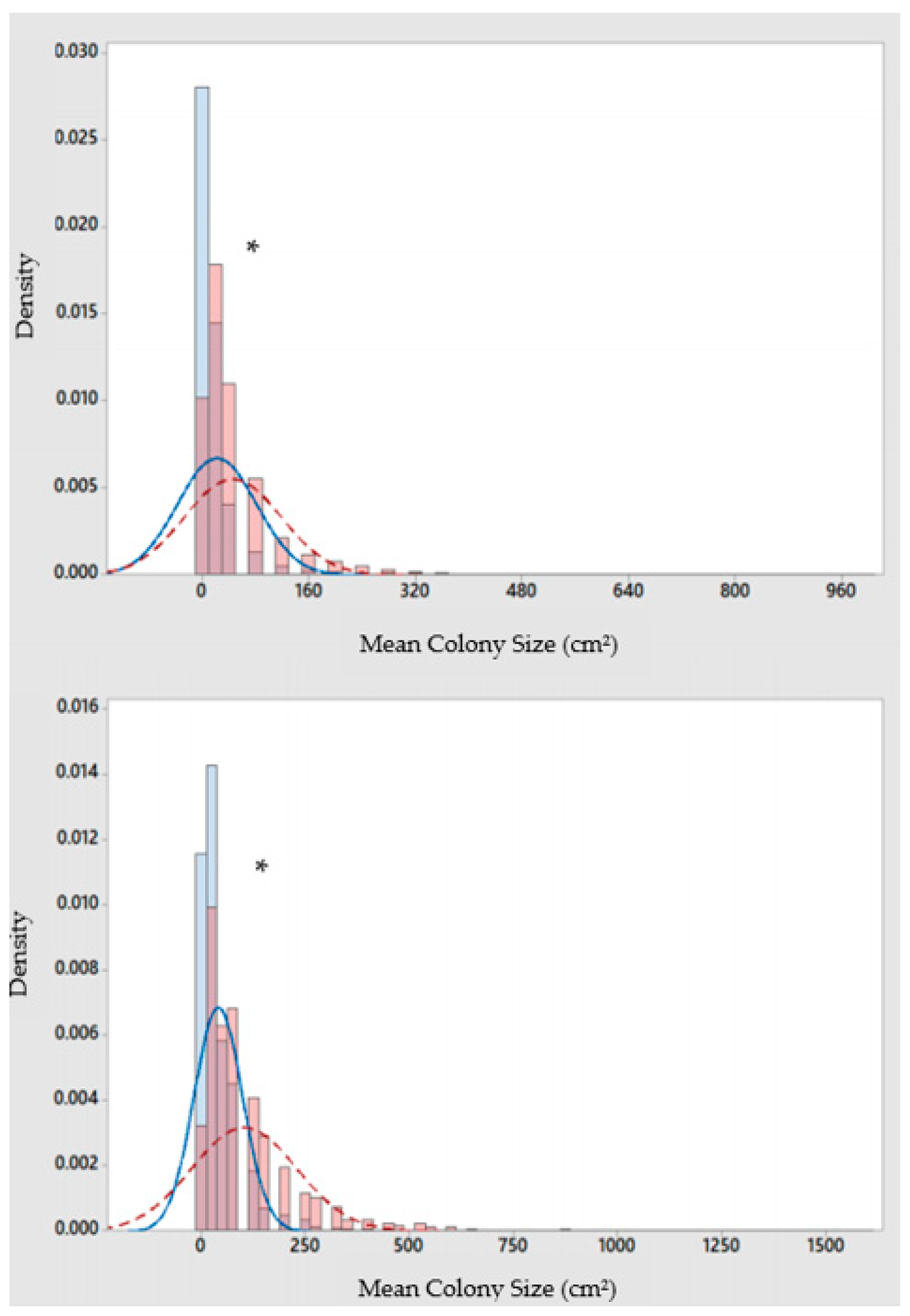
3.2. P. astreoides in Grenada
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gardner, T.A.; Côté, I.M.; Gill, J.A.; Grant, A.; Watkinson, A.R. Long-Term Region-Wide Declines in Caribbean Corals. Science 2003, 301, 958–960. [Google Scholar] [CrossRef] [Green Version]
- Jackson, J.B.C.; Donovan, M.K.; Cramer, K.L.; Lam, W. Status and Trends of Caribbean Coral Reefs: 1970–2012; International Union for the Conservation of Nature: Gland, Switzerland, 2014. [Google Scholar]
- Hughes, T.P.; Baird, A.H.; Bellwood, D.R.; Card, M.; Connolly, S.R.; Folke, C.; Grosberg, R.; Hoegh-Guldberg, O.; Jackson, J.B.C.; Kleypas, J.; et al. Climate Change, Human Impacts, and the Resilience of Coral Reefs. Science 2003, 301, 929–933. [Google Scholar] [CrossRef] [Green Version]
- Castillo, K.D.; Ries, J.B.; Weiss, J.M.; Lima, F.P. Decline of forereef corals in response to recent warming linked to history of thermal exposure. Nat. Clim. Chang. 2012, 2, 756–760. [Google Scholar] [CrossRef]
- Okazaki, R.R.; Towle, E.K.; Hooidonk, R.; Mor, C.; Winter, R.N.; Piggot, A.M.; Cunning, R.; Baker, A.C.; Klaus, J.S.; Swart, P.K.; et al. Species-specific responses to climate change and community composition determine future calcification rates of Florida Keys reefs. Glob. Chang. Biol. 2017, 23, 1023–1035. [Google Scholar] [CrossRef]
- Anthony, K.R.; Marshall, P.A.; Abdulla, A.; Beeden, R.; Bergh, C.; Black, R.; Eakin, C.M.; Game, E.; Gooch, M.; Graham, N.; et al. Operationalizing resilience for adaptive coral reef management under global environmental change. Glob. Chang. Biol. 2015, 21, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Perry, C.T.; Steneck, R.S.; Murphy, G.N.; Kench, P.S.; Edinger, E.N.; Smithers, S.G.; Mumby, P. Regional-scale dominance of non-framework building corals on Caribbean reefs affects carbonate production and future reef growth. Glob. Chang. Biol. 2015, 21, 1153–1164. [Google Scholar] [CrossRef]
- Van Woesik, R.; Sakai, K.; Ganase, A.; Loya, Y. Revisiting the winners and the losers a decade after coral bleaching. Mar. Ecol. Prog. Ser. 2011, 434, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Darling, E.S.; Alvarez-Filip, L.; Oliver, T.A.; McClanahan, T.R.; Côté, I.M. Evaluating life-history strategies of reef corals from species traits. Ecol. Lett. 2012, 15, 1378–1386. [Google Scholar] [CrossRef]
- Schutte, V.G.W.; Selig, E.R.; Bruno, J.F. Regional spatio-temporal trends in Caribbean coral reef benthic communities. Mar. Ecol. Prog. Ser. 2010, 402, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Martínez, R.; Banaszak, A.T.; McField, M.D.; Beltrán-Torres, A.U.; Álvarez-Filip, L. Assessment of Acropora palmata in the Mesoamerican Reef System. PLoS ONE 2014, 9, e96140. [Google Scholar] [CrossRef] [Green Version]
- Green, D.; Edmunds, P.; Carpenter, R. Increasing relative abundance of Porites astreoides on Caribbean reefs mediated by an overall decline in coral cover. Mar. Ecol. Prog. Ser. 2008, 359, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Medina-Valmaseda, A.E.; Rodríguez-Martínez, R.E.; Alvarez-Filip, L.; Jordan-Dahlgren, E.; Blanchon, P. The role of geomorphic zonation in long-term changes in coral-community structure on a Caribbean fringing reef. PeerJ 2020, 8, e10103. [Google Scholar] [CrossRef]
- Eagleson, R. Spatial Distribution of Benthic Habitats and Ecological Patterns of the Mustard Hill Coral (Porites Astreoides) in the Nearshore Waters of Grenada. Master’s Thesis, University of Guelph, Guelph, ON, Canada, 2019. [Google Scholar]
- Edmunds, P. Population biology of Porites astreoides and Diploria strigosa on a shallow Caribbean reef. Mar. Ecol. Prog. Ser. 2010, 418, 87–104. [Google Scholar] [CrossRef]
- Baumann, J.H.; Townsend, J.E.; Courtney, T.A.; Aichelman, H.E.; Davies, S.W.; Lima, F.P.; Castillo, K.D. Temperature Regimes Impact Coral Assemblages along Environmental Gradients on Lagoonal Reefs in Belize. PLoS ONE 2016, 11, e0162098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodbody-Gringley, G.; de Putron, S.J. Brooding Corals: Planulation Patterns, Larval Behavior, and Recruitment Dynamics in the Face of Environmental Change. In The Cnidaria, Past, Present and Future; Springer International Publishing: Cham, Switzerland, 2016; pp. 279–289. [Google Scholar]
- Edmunds, P.J.; Didden, C.; Frank, K. Over three decades, a classic winner starts to lose in a Caribbean coral community. Ecosphere 2021, 12, e03517. [Google Scholar] [CrossRef]
- Horricks, R.A.; Herbinger, C.M.; Lillie, B.N.; Taylor, P.; Lumsden, J.S. Differential protein abundance during the first month of regeneration of the Caribbean star coral Montastraea cavernosa. Coral Reefs 2018, 38, 45–61. [Google Scholar] [CrossRef]
- Anderson, R.; Morrall, C.; Nimrod, S.; Balza, R.; Berg, C.; Jossart, J. Benthic and fish population monitoring associated with a marine protected area in the nearshore waters of Grenada, Eastern Caribbean. Rev. Biol. Trop. 2012, 60, 71–87. [Google Scholar] [CrossRef] [Green Version]
- Anderson, R.A.; Morrall, C.B.; Nimrod, S.B.; Balza, R.A.; Berg, C.; Jossart, J.A. Marine Protected Area Monitoring in the Nearshore Waters of Grenada, Eastern Caribbean: Benthic Cover and Fish Populations. Rev. Biol. Trop. 2014, 62, 273–286. [Google Scholar] [CrossRef] [Green Version]
- Horricks, R.A. Tissue Regeneration of Artificially Induced Lesions in the Caribbean Great Star Coral (Montastraea Cavernosa) in the Nearshore Waters of Grenada. Ph.D. Thesis, University of Guelph, Guelph, ON, Canada, 2017. [Google Scholar]
- Kohler, K.E.; Gill, S.M. Coral Point Count with Excel extensions (CPCe): A Visual Basic program for the determination of coral and substrate coverage using random point count methodology. Comput. Geosci. 2006, 32, 1259–1269. [Google Scholar] [CrossRef]
- Dumas, P.; Bertaud, A.; Peignon, C.; Léopold, M.; Pelletier, D. A “quick and clean” photographic method for the description of coral reef habitats. J. Exp. Mar. Biol. Ecol. 2009, 368, 161–168. [Google Scholar] [CrossRef]
- Alquezar, R.; Boyd, W. Development of rapid, cost effective coral survey techniques: Tools for management and conservation planning. J. Coast. Conserv. 2007, 11, 105–119. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; ISSN 17513243. ISBN 9780080554501. [Google Scholar]
- Oksanen, J. Vegan: Ecological Diversity. 2016, p. 12. Available online: https://www.semanticscholar.org/paper/Vegan%3A-ecological-diversity-Oksanen/6d1f21af6a90af203ab680657c215008e292cea6 (accessed on 27 November 2020).
- Bonett, D.G.; Wright, T.A. Sample size requirements for estimating pearson, kendall and spearman correlations. Psychometrika 2000, 65, 23–28. [Google Scholar] [CrossRef]
- Greenstein, B.J.; Curran, H.A.; Pandolfi, J.M. Shifting ecological baselines and the demise of Acropora cervicornis in the western North Atlantic and Caribbean Province: A Pleistocene perspective. Coral Reefs 1998, 17, 249–261. [Google Scholar] [CrossRef]
- Toth, L.T.; Stathakopoulos, A.; Kuffner, I.B.; Ruzicka, R.R.; Colella, M.A.; Shinn, E.A. The unprecedented loss of Florida’s reef-building corals and the emergence of a novel coral-reef assemblage. Ecology 2019, 100, e02781. [Google Scholar] [CrossRef] [Green Version]
- Burke, L.; Maidens, J. Reefs at Risk in the Caribbean; World Resources Institute: Washington, DC, USA, 2004; ISBN 1559632574. [Google Scholar]
- Neal, B.P.; Khen, A.; Treibitz, T.; Beijbom, O.; O’Connor, G.; Coffroth, M.A.; Knowlton, N.; Kriegman, D.; Mitchell, B.G.; Kline, D. Caribbean massive corals not recovering from repeated thermal stress events during 2005–2013. Ecol. Evol. 2017, 7, 1339–1353. [Google Scholar] [CrossRef]
- Aronson, R.B.; MacIntyre, I.G.; Wapnick, C.M.; O’Neill, M.W. Phase Shifts, Alternative States, and the Unprecedented Convergence of Two Reef Systems. Ecology 2004, 85, 1876–1891. [Google Scholar] [CrossRef]
- Goodwin, M.; Cole, M.; Stewart, W.; Zimmerman, B. Species density and associations in Caribbean reef corals. J. Exp. Mar. Biol. Ecol. 1976, 24, 19–31. [Google Scholar] [CrossRef]
- Deschamps, A.; DesRochers, A.; Klomp, K.D. St. Vincent. A Rapid Assessment of the Horseshoe Reef, Tobago Cays Marine Park, St. Vincent, West Indies (Stony Corals, Algae and Fishes). Atoll Res. Bull. 2003, 496, 438–459. [Google Scholar] [CrossRef] [Green Version]
- Lewis, J.B. A preliminary description of the coral reefs of the Tobago Cays, Grenadines, West Indies. Atoll Res. Bull. 1975, 178, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.R. Patterns of Coral Recruitment and Post-Settlement Mortality on Bermuda’s Reefs: Comparisons to Caribbean and Pacific Reefs. Am. Zool. 1992, 33, 663–673. [Google Scholar] [CrossRef]
- Votaw, G.S.; Rosa, L.; Snell, W.; Anselmi, C.; History, S. Tropical Storm Chantal; National Weather Service: Washington, DC, USA, 2013. [Google Scholar]
- Mallela, J.; Crabbe, M. Hurricanes and coral bleaching linked to changes in coral recruitment in Tobago. Mar. Environ. Res. 2009, 68, 158–162. [Google Scholar] [CrossRef] [Green Version]
- Madin, J.S.; Anderson, K.D.; Andreasen, M.H.; Bridge, T.C.; Cairns, S.D.; Connolly, S.R.; Darling, E.S.; Diaz, M.; Falster, D.S.; Franklin, E.C.; et al. The Coral Trait Database, a curated database of trait information for coral species from the global oceans. Sci. Data 2016, 3, 160017. [Google Scholar] [CrossRef] [Green Version]
- Chornesky, E.; Peters, E. Sexual Reproduction and Colony Growth in the Scleractinian Coral Porites Astreoides. Biol. Bull. 1987, 172, 161–177. [Google Scholar] [CrossRef]
- Bythell, J.C. Assessment of the Impacts of Hurricanes Marilyn and Luis and Post-Hurricane Community Dynamics at Buck Island Reef National Monument; Technical Report; US Department of Interior, National Park Service: Washington, DC, USA, 1997. [Google Scholar] [CrossRef]
- Brazeau, D.A.; Gleason, D.F.; Morgan, M.E. Self-Fertilization in Brooding Hermaphroditic Caribbean Corals: Evidence from Molecular Markers. J. Exp. Mar. Biol. Ecol. 1998, 231, 225–238. [Google Scholar] [CrossRef]
- Vega Thurber, R.; Burkepile, D.E.; Correa, A.M.S.; Thurber, A.R.; Shantz, A.A.; Welsh, R.; Pritchard, C.; Rosales, S. Macroalgae Decrease Growth and Alter Microbial Community Structure of the Reef-Building Coral, Porites Astreoides. PLoS ONE 2012, 7, e44246. [Google Scholar] [CrossRef] [Green Version]
- Cameron, C.M.; Pausch, R.E.; Miller, M.W. Coral Recruitment Dynamics and Substrate Mobility in a Rubble-Dominated Back Reef Habitat. Bull. Mar. Sci. 2016, 92, 123–136. [Google Scholar] [CrossRef]
- Ritson-Williams, R.; Arnold, S.; Paul, V. Patterns of larval settlement preferences and post-settlement survival for seven Caribbean corals. Mar. Ecol. Prog. Ser. 2016, 548, 127–138. [Google Scholar] [CrossRef]
- Ross, C.; Ritson-Williams, R.; Olsen, K.; Paul, V.J. Short-Term and Latent Post-Settlement Effects Associated with Elevated Temperature and Oxidative Stress on Larvae from the Coral Porites Astreoides. Coral Reefs 2013, 32, 71–79. [Google Scholar] [CrossRef]
- Olsen, K.; Sneed, J.; Paul, V.J. Differential larval settlement responses of Porites astreoides and Acropora palmata in the presence of the green alga Halimeda opuntia. Coral Reefs 2016, 35, 521–525. [Google Scholar] [CrossRef]
- Fava, F.; Ponti, M.; Scinto, A.; Calcinai, B.; Cerrano, C. Possible effects of human impacts on epibenthic communities and coral rubble features in the marine Park of Bunaken (Indonesia). Estuarine Coast. Shelf Sci. 2009, 85, 151–156. [Google Scholar] [CrossRef]
- Gilby, B.L.; Maxwell, P.S.; Tibbetts, I.; Stevens, T. Bottom-Up Factors for Algal Productivity Outweigh No-Fishing Marine Protected Area Effects in a Marginal Coral Reef System. Ecosystems 2015, 18, 1056–1069. [Google Scholar] [CrossRef]
- Olsen, K.; Paul, V.J.; Ross, C. Direct effects of elevated temperature, reduced pH, and the presence of macroalgae (Dictyota spp.) on larvae of the Caribbean coral Porites astreoides. Bull. Mar. Sci. 2015, 91, 255–270. [Google Scholar] [CrossRef]
- Burkepile, D.E.; Hay, M. Impact of Herbivore Identity on Algal Succession and Coral Growth on a Caribbean Reef. PLoS ONE 2010, 5, e8963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atlantic and Gulf Rapid Reef Assessment. Grenada’s Coral Reef Report Card; Ocean Research and Education Foundation: Key Largo, FL, USA, 2016; Available online: http://caribnode.org/documents/85 (accessed on 15 November 2017).
- Done, T.J. Coral Community Adaptability to Environmental Change at the Scales of Regions, Reefs and Reef Zones. Am. Zool. 1999, 39, 66–79. [Google Scholar] [CrossRef] [Green Version]
- Williams, S.; Mumby, P.; Chollett, I.; Cortés, J. Importance of differentiating Orbicella reefs from gorgonian plains for ecological assessments of Caribbean reefs. Mar. Ecol. Prog. Ser. 2015, 530, 93–101. [Google Scholar] [CrossRef]
- Riegl, B.; Purkis, S.J. Coral population dynamics across consecutive mass mortality events. Glob. Chang. Biol. 2015, 21, 3995–4005. [Google Scholar] [CrossRef]
- De Barros, M.M.L.; Pires, D.D.O. Colony size-frequency distributions among different populations of the scleractinan coral Siderastrea stellata in Southwestern Atlantic: Implications for life history patterns. Braz. J. Oceanogr. 2006, 54, 213–223. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Island | Site Name | Site Code | GPS Coordinates | R |
|---|---|---|---|---|
| Grenada | Dragon Bay | DRAG | 12°5′6.00″ N 61°45′45.36″ W | 0.958 * |
| Grenada | Flamingo Bay | FLAM | 12°5′30.36″ N 61°45′30.60″ W | 1.00 * |
| Grenada | Northern Exposure | NORTH | 12°2′22.14″ N 61°46′4.74″ W | 0.990 * |
| Grenada | Quarter Wreck | QUART | 12°1′40.98″ N 61°47′0.84″ W | 0.990 * |
| Carriacou | Sandy Island | SAND | 12°29′7.77″ N 61°29′2.00″ W | 0.656 * |
| Carriacou | Whirlpool | WHIRL | 12°29′2.50″ N 61°29′31.35″ W | 0.927 * |
| Carriacou | Jackadam | JAD | 12°29′46.25″ N 61°28′4.11″ W | 0.937 * |
| Carriacou | Seaview | SEAV | 12°26′44.13″ N 61°29′59.70″ W | 1.000 * |
| Factor | |
|---|---|
| Acropora cervicornis | Mycetophyllia aliciae |
| Acropora palmata | Mycetophyllia danaana |
| Acropora prolifera | Mycetophyllia ferox |
| Agaricia agaricites | Mycetophyllia lamarckiana |
| Agaricia fragilis | Oculina diffusa |
| Agaricia grahamae | Orbicella annularis |
| Agaricia lamarcki | Orbicella faveolata |
| Agaricia tenuifolia | Orbicella franksi |
| Agaricia undata | Porites branneri |
| Colpophyllia breviserialis | Porites divaricate |
| Colpophyllia natans | Porites furcata |
| Dendrogyra cylindrus | Porites porites |
| Dichocoenia stellaris | Scolymia cubensis |
| Dichocoenia stokes | Scolymia lacera |
| Diploria clivosa | Siderastrea radians |
| Diploria labyrinthiformis | Siderastrea siderea |
| Diploria strigosa | Solenastrea bournoni |
| Eusmilia fastigiata | Solenastrea hyades |
| Favia fragum | Stephanocoenia michelinii |
| Isophyllia sinuosa | Tubastraea aurea |
| Leptoseris cucullata | Coralline algae |
| Madracis decactis | Gorgonians |
| Madracis mirabilis | Macroalgae |
| Manicina areolata | Sponges |
| Meandrina meandrites | Zoathids |
| Millipora alcicornis | Sand |
| Millipora complanata | Rubble |
| Millipora squarrosa | Pavement |
| Montastraea cavernosa | |
| Mussa angulosa | |
| Taxon | CAR 2014 | CAR 2017 | GND 2014 | GND 2017 |
|---|---|---|---|---|
| Agaricia agaricites | 0.037 ± 0.003 | 0.046 ± 0.004 | 0.014 ± 0.002 | 0.016 ± 0.002 |
| Agaricia lamarcki | 0.000 ± 0.000 | 0.003 ± 0.001 | 0.000 ± 0.000 | 0.001 ± 0.000 |
| Colpophyllia natans | 0.005 ± 0.001 | 0.092 ± 0.005 | 0.000 ± 0.000 | 0.039 ± 0.004 |
| Dendrogyra cylindrus | 0.003 ± 0.001 | 0.001 ± 0.001 | 0.003 ± 0.001 | 0.001 ± 0.001 |
| Diploria labyrinthiformis | 0.013 ± 0.002 | 0.021 ± 0.003 | 0.004 ± 0.001 | 0.005 ± 0.001 |
| Diploria strigosa | 0.081 ± 0.005 | 0.006 ± 0.001 | 0.035 ± 0.003 | 0.001 ± 0.001 |
| Eusmilia fastigiata | 0.000 ± 0.000 | 0.001 ± 0.001 | 0.002 ± 0.001 | 0.001 ± 0.001 |
| Madracis decactis | 0.000 ± 0.000 | 0.019 ± 0.003 | 0.000 ± 0.000 | 0.015 ± 0.002 |
| Madracis mirabilis | 0.100 ± 0.005 | 0.102 ± 0.006 | 0.250 ± 0.007 | 0.253 ± 0.008 |
| Meandrina meandrites | 0.015 ± 0.002 | 0.017 ± 0.002 | 0.007 ± 0.001 | 0.005 ± 0.001 |
| Millipora alcicornis | 0.033 ± 0.003 | 0.040 ± 0.004 | 0.014 ± 0.002 | 0.013 ± 0.002 |
| Millipora complanata | 0.018 ± 0.002 | 0.000 ± 0.000 | 0.003 ± 0.001 | 0.000 ± 0.000 |
| Millipora squarrosa | 0.000 ± 0.000 | 0.003 ± 0.001 | 0.000 ± 0.000 | 0.001 ± 0.001 |
| Montastraea cavernosa | 0.039 ± 0.004 | 0.032 ± 0.003 | 0.036 ± 0.003 | 0.026 ± 0.003 |
| Oculina diffusa | 0.000 ± 0.000 | 0.001 ± 0.001 | 0.000 ± 0.000 | 0.000 ± 0.000 |
| * Orbicella annularis | 0.147 ± 0.006 | 0.122 ± 0.006 | 0.169 ± 0.006 | 0.094 ± 0.005 |
| Orbicella faveolata | 0.003 ± 0.001 | 0.000 ± 0.000 | 0.005 ± 0.001 | 0.000 ± 0.000 |
| Orbicella franksi | 0.000 ± 0.000 | 0.020 ± 0.003 | 0.000 ± 0.000 | 0.074 ± 0.005 |
| Porites astreoides | 0.221 ± 0.008 | 0.199 ± 0.008 | 0.212 ± 0.007 | 0.211 ± 0.007 |
| * Porites divaricata | 0.029 ± 0.003 | 0.178 ± 0.007 | 0.056 ± 0.004 | 0.163 ± 0.007 |
| * Porites porites | 0.174 ± 0.007 | 0.000 ± 0.000 | 0.118 ± 0.005 | 0.005 ± 0.001 |
| Siderastrea siderea | 0.077 ± 0.005 | 0.089 ± 0.005 | 0.072 ± 0.004 | 0.070 ± 0.005 |
| Solenastrea bournoni | 0.000 ± 0.000 | 0.007 ± 0.002 | 0.000 ± 0.000 | 0.004 ± 0.001 |
| Stephanocoenia michelinii | 0.006 ± 0.001 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 |
| Carriacou | Grenada | ||||
|---|---|---|---|---|---|
| 2014 | 2014 | ||||
| Factor | r | p-value | Factor | r | p-value |
| Rubble | −0.168 | 0.0001 * | Rubble | −0.101 | 0.002 * |
| Pavement | 0.193 | 0.0001 * | Sand | −0.122 | 0.0001 * |
| Sand | −0.168 | 0.0001 * | Madracis auretenra | −0.127 | 0.0001 * |
| Diploria strigosa | 0.159 | 0.0001 * | Orbicella annularis | 0.047 | 0.148 |
| Orbicella annularis | −0.031 | 0.345 | Porites divaricata | −0.006 | 0.844 |
| Porites porites | −0.118 | 0.0001 * | Porites porites | −0.048 | 0.143 |
| Siderastrea siderea | 0.013 | 0.685 | Siderastrea siderea | −0.063 | 0.054 |
| Gorgonians | −0.095 | 0.003 * | Gorgonians | −0.156 | 0.0001 * |
| Sponges | 0.259 | 0.0001 * | Sponges | −0.128 | 0.0001 * |
| Macroalgae | −0.288 | 0.0001 * | Macroalgae | 0.071 | 0.03 * |
| Coralline Algae | 0.199 | 0.0001 * | Coralline Algae | 0.099 | 0.002 * |
| 2017 | 2017 | ||||
| Factor | r | p-value | Factor | r | p-value |
| Rubble | −0.196 | 0.0001 * | Rubble | −0.073 | 0.027 * |
| Sand | −0.145 | 0.0001 * | Sand | −0.114 | 0.001 * |
| Colpophyllia natans | 0.126 | 0.0001 * | Madracis auretenra | −0.101 | 0.002 * |
| Orbicella annularis | −0.044 | 0.179 | Orbicella annularis | −0.03 | 0.364 |
| Porites divaricata | −0.078 | 0.017 * | Orbicella franksi | 0.088 | 0.008 * |
| Siderastrea siderea | −0.026 | 0.417 | Porites divaricata | −0.049 | 0.141 |
| Sponges | 0.199 | 0.0001 * | Siderastrea siderea | −0.017 | 0.61 |
| Macroalgae | −0.238 | 0.0001 * | Gorgonians | −0.14 | 0.0001 * |
| Sponges | −0.057 | 0.083 | |||
| Coralline Algae | 0.116 | 0.0001 * | Macroalgae | −0.083 | 0.012 * |
| Coralline Algae | 0.119 | 0.0001 * | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eagleson, R.G.; Lumsden, J.S.; Álvarez-Filip, L.; Herbinger, C.M.; Horricks, R.A. Coverage Increases of Porites astreoides in Grenada Determined by Shifts in Size-Frequency Distribution. Diversity 2021, 13, 288. https://doi.org/10.3390/d13070288
Eagleson RG, Lumsden JS, Álvarez-Filip L, Herbinger CM, Horricks RA. Coverage Increases of Porites astreoides in Grenada Determined by Shifts in Size-Frequency Distribution. Diversity. 2021; 13(7):288. https://doi.org/10.3390/d13070288
Chicago/Turabian StyleEagleson, Ryan G., John S. Lumsden, Lorenzo Álvarez-Filip, Christophe M. Herbinger, and Ryan A. Horricks. 2021. "Coverage Increases of Porites astreoides in Grenada Determined by Shifts in Size-Frequency Distribution" Diversity 13, no. 7: 288. https://doi.org/10.3390/d13070288
APA StyleEagleson, R. G., Lumsden, J. S., Álvarez-Filip, L., Herbinger, C. M., & Horricks, R. A. (2021). Coverage Increases of Porites astreoides in Grenada Determined by Shifts in Size-Frequency Distribution. Diversity, 13(7), 288. https://doi.org/10.3390/d13070288