
Genetic Investigation of Aral Wild Common Carp Populations (Cyprinus carpio) Using ddRAD Sequencing

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
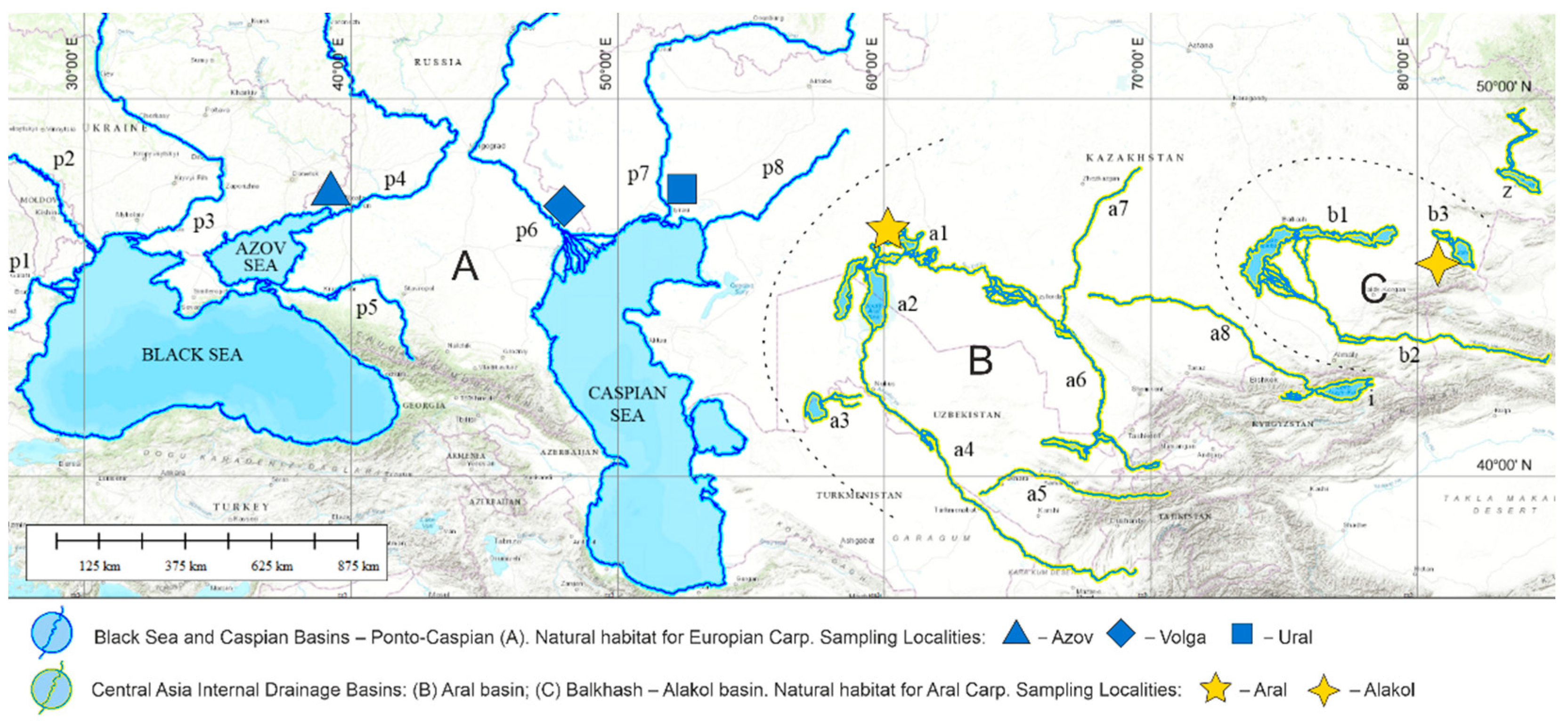
2.1. Fish Sampling, DNA Extraction and Sequencing
2.2. Data Analysis
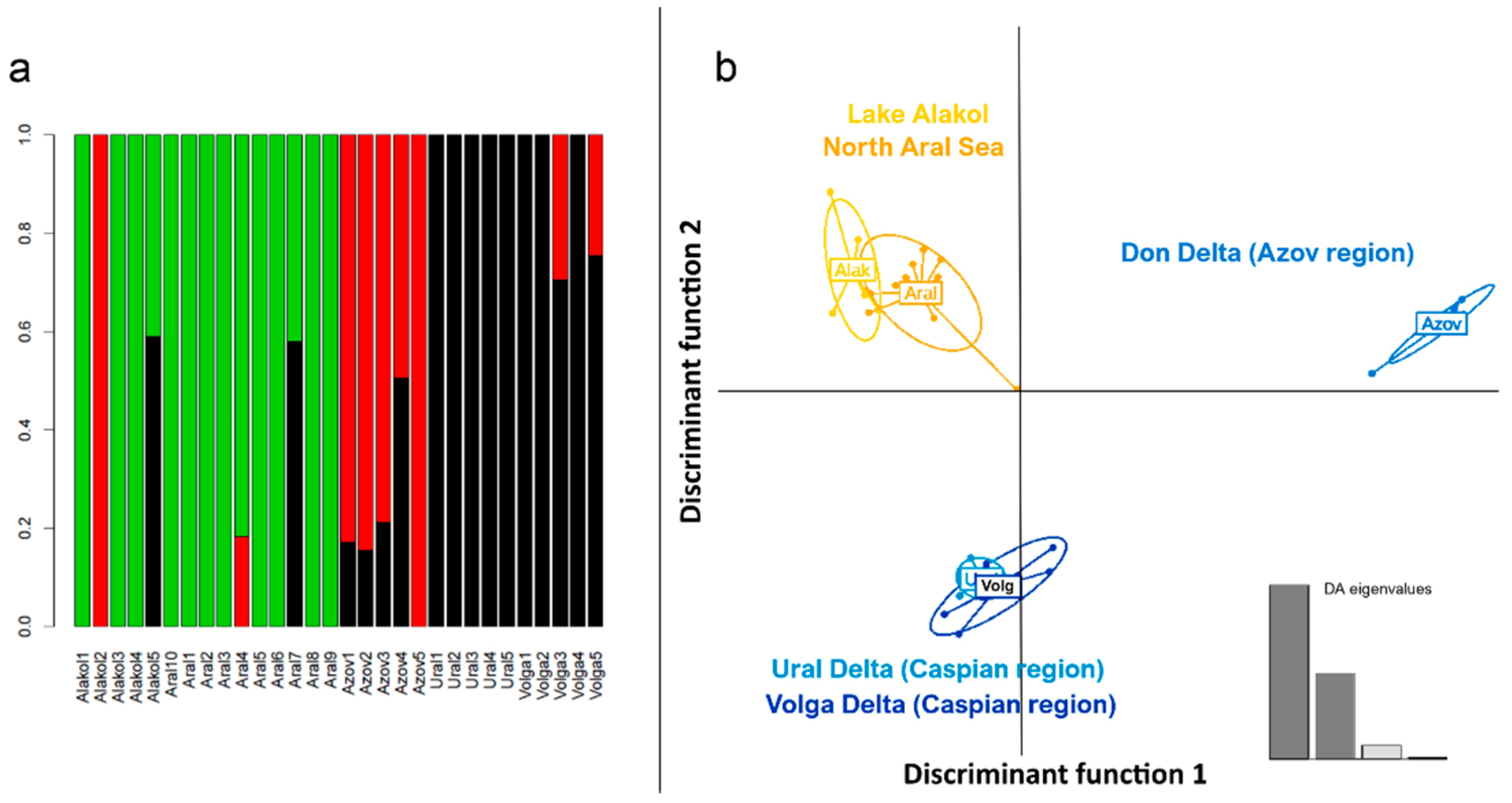
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nelson, J.S. Fishes of the World, 3rd ed.; John Wiley & Sons: New York, NY, USA, 1995. [Google Scholar]
- Gross, R.; Kohlmann, K.; Kersten, P. PCR–RFLP analysis of the mitochondrial ND-3/4 and ND-5/6 gene polymorphisms in the European and East Asian subspecies of common carp (Cyprinus carpio L.). Aquaculture 2002, 204, 507–516. [Google Scholar] [CrossRef]
- Balon, E.K. The oldest domesticated fishes, and the consequences of an epigenetic dichotomy. Int. J. Ichtyol. 2006, 2006, 44–86. [Google Scholar]
- Svetovidov, A.N. Ueber den europaeischen und ostasiatischen Karpfen (Cyprinus carpio). Zool. Anz. 1933, 104, 257–261. [Google Scholar]
- Xu, P.; Zhang, X.; Wang, X.; Li, J.; Liu, G.; Kuang, Y.; Xu, J.; Zheng, X.; Ren, L.; Wang, G.; et al. Genome sequence and genetic diversity of the common carp, Cyprinus carpio. Nat. Genet 2014, 46, 1212–1219. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Wu, Q.; Wang, Z.; Ye, Y. Molecular phylogeny of three subspecies of common carp Cyprinus carpio, based on sequence analysis of cytochrome b and control region of mtDNA. J. Zool. Syst. Evol. Res. 2004, 42, 266–269. [Google Scholar] [CrossRef]
- Chistiakov, D.A.; Voronova, N.V. Genetic evolution and diversity of common carp Cyprinus carpio L. Cent. Eur. J. Biol. 2009, 4, 304–312. [Google Scholar] [CrossRef]
- Gvozdev, E.V. Ryby Kazakhstana (Fishes of Kazakhstan); Nauka: Almaty, Kazakhstan, 1988; Volume 3, p. 304. [Google Scholar]
- Kirpitchnikov, V.S. Homologous hereditary variation and evolution of wild common carp Cyprinus carpio L. Genetika 1967, 8, 65–72. [Google Scholar]
- Aladin, N.V.; Gontar, V.I.; Zhakova, L.V.; Plotnikov, I.S.; Smurov, A.O.; Rzymski, P.; Klimaszyk, P. The zoocenosis of the Aral Sea: Six decades of fast-paced change. Environ. Sci. Pollut. Res. Int. 2019, 26, 2228–2237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izhitskiy, A.S.; Zavialov, P.O.; Sapozhnikov, P.V.; Kirillin, G.B.; Grossart, H.P.; Kalinina, O.Y.; Zalota, A.K.; Goncharenko, I.V.; Kurbaniyazov, A.K. Present state of the Aral Sea: Diverging physical and biological characteristics of the residual basins. Sci. Rep. 2016, 6, 23906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohlmann, K.; Gross, R.; Murakaeva, A.; Kersten, P. Genetic variability and structure of common carp (Cyprinus carpio) populations throughout the distribution range inferred from allozyme, microsatellite and mitochondrial DNA markers, in: Aquatic Living Resources. Aquat. Living Resour. 2003, 16, 421–431. [Google Scholar] [CrossRef]
- Kohlmann, K.; Kersten, P.; Flajšhans, M. Microsatellite-based genetic variability and differentiation of domesticated, wild and feral common carp (Cyprinus carpio L.) populations. Aquaculture 2005, 247, 253–266. [Google Scholar] [CrossRef]
- Memiş, D.; Kohlmann, K. Genetic characterization of wild common carp (Cyprinus carpio L.) from Turkey. Aquaculture 2006, 258, 257–262. [Google Scholar] [CrossRef]
- Hohenlohe, P.A.; Bassham, S.; Etter, P.D.; Stiffler, N.; Johnson, E.A.; Cresko, W.A. Population genomics of parallel adaptation in threespine stickleback using sequenced RAD tags. PLoS Genet 2010, 6, e1000862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franchini, P.; Monne Parera, D.; Kautt, A.F.; Meyer, A. quaddRAD: A new high-multiplexing and PCR duplicate removal ddRAD protocol produces novel evolutionary insights in a nonradiating cichlid lineage. Mol. Ecol. 2017, 26, 2783–2795. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.F.; Russell, D.W.; Maniatis, T. (Eds.) Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor: New York, NY, USA, 1989; p. 676. [Google Scholar]
- Rochette, N.C.; Catchen, J.M. Deriving genotypes from RAD-seq short-read data using Stacks. Nat. Protoc. 2017, 12, 2640–2659. [Google Scholar] [CrossRef]
- Langmead, B.; Wilks, C.; Antonescu, V.; Charles, R. Scaling read aligners to hundreds of threads on general-purpose processors. Bioinformatics 2019, 35, 421–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larhammar, D.; Risinger, C. Molecular genetic aspects of tetraploidy in the common carp Cyprinus carpio. Mol. Phylogenet Evol. 1994, 3, 59–68. [Google Scholar] [CrossRef]
- Knaus, B.J.; Grunwald, N.J. vcfr: A package to manipulate and visualize variant call format data in R. Mol. Ecol. Resour. 2017, 17, 44–53. [Google Scholar] [CrossRef]
- Jombart, T.; Ahmed, I. adegenet 1.3-1: New tools for the analysis of genome-wide SNP data. Bioinformatics 2011, 27, 3070–3071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.; Levine, D.; Shen, J.; Gogarten, S.M.; Laurie, C.; Weir, B.S. A high-performance computing toolset for relatedness and principal component analysis of SNP data. Bioinformatics 2012, 28, 3326–3328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef] [Green Version]
- Quinlan, A.R.; Hall, I.M. BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef] [Green Version]
- Mount, D.W. Using the Basic Local Alignment Search Tool (BLAST). CSH Protoc. 2007, 2007. [Google Scholar] [CrossRef]
- Gruber, B.; Unmack, P.J.; Berry, O.F.; Georges, A. DARTR: An R package to facilitate analysis of SNP data generated from reduced representation genome sequencing. Mol. Ecol. Resour. 2018, 18, 691–699. [Google Scholar] [CrossRef]
- Sherwin, W.B.; Chao, A.; Jost, L.; Smouse, P.E. Information Theory Broadens the Spectrum of Molecular Ecology and Evolution. Trends Ecol. Evol. 2017, 32, 948–963. [Google Scholar] [CrossRef] [PubMed]
- Hartl, D.L.; Clark, A.G. Principles of Population Genetics, 4th ed.; Sinauer Associates: Sunderland, MA, USA, 2007; 652p. [Google Scholar]
- Nikolsky, G.V. Ichtyology; Vysshaya Shkola: Moscow, Russia, 1971; p. 471. [Google Scholar]
- Nedoluzhko, A.V.; Slobodova, N.V.; Sharko, F.; Shalgimbayeva, G.M.; Tsygankova, S.V.; Boulygina, E.S.; Jeney, Z.; Nguyen, V.Q.; Pham, T.T.; Nguyen, D.T.; et al. A new strain group of common carp: The genetic differences and admixture events between Cyprinus carpio breeds. Ecol. Evol. 2020, 10, 5431–5439. [Google Scholar] [CrossRef] [PubMed]
- Nedoluzhko, A.V.; Gladysheva-Azgari, M.V.; Shalgimbayeva, G.M.; Volkov, A.A.; Slobodova, N.V.; Tsygankova, S.V.; Boulygina, E.S.; Nguyen, V.Q.; Pham, T.T.; Nguyen, D.T.; et al. Genetic contribution of domestic European common carp (Cyprinus carpio carpio) and Amur carp (Cyprinus carpio haematopterus) to the wild Vietnamese carp population as revealed by ddRAD sequencing. Aquaculture 2021, 544, 737049. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Sample Name/DAPC Abbreviation | Sampling Locality | Collection Repository | Number of Specimens | NCBI Accessions |
|---|---|---|---|---|
| Alak | Lake Alakol, Kazakhstan | FRPC * | 5 | SAMN12827353- SAMN12827357 |
| Aral | North Aral Sea, Kazakhstan | FRPC | 10 | SAMN12827368- SAMN12827377 |
| Azov | Don Delta, Azov Sea, Russia | VNIRO ** | 5 | SAMN12827378- SAMN12827382 |
| Volga | Volga Delta, Caspian Sea, Russia | VNIRO | 5 | SAMN12827446- SAMN12827450 |
| Ural | Ural Delta, Caspian Sea, Kazakhstan | FRPC | 5 | SAMN12827431- SAMN12827435 |
| SNP Position According to GCA_004011595.1 | DP | Ar/Ur Allele | Gene | Gene Description/NCBI Access# |
|---|---|---|---|---|
| SAUJ01045606.1_275599 | 0.005815405 | A/T | – | – |
| SAUJ01041730.1_1504105 | 0.005552164 | T/C | – | – |
| SAUJ01042215.1_278967 | 0.005461386 | A/G | ano10b | anoctamin 10b |
| SAUJ01041730.1_1504004 | 0.005149558 | G/T | ccl36.1 | chemokine (C-C motif) ligand 36 |
| SAUJ01045606.1_275498 | 0.005103472 | G/T | – | – |
| SAUJ01046983.1_1471585 | 0.003859412 | A/T | brd9 | bromodomain containing 9 |
| SAUJ01045448.1_218919 | 0.003709669 | C/T | – | CR855320.1 |
| SAUJ01041569.1_35062 | 0.003702634 | C/A | si:ch73-233k15.2 | Orthologous to human APOL3 |
| SAUJ01047296.1_2308559 | 0.003442002 | T/C | npffr2b | neuropeptide FF receptor 2b |
| SAUJ01232093.1_1293 | 0.003257919 | G/A | yjefn3 | YjeF N-terminal domain containing 3 |
| SAUJ01045205.1_157187 | 0.003208534 | C/T | ywhae2 | tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein, epsilon polypeptide 2 |
| SAUJ01046983.1_1471607 | 0.003102829 | G/T | brd9 | bromodomain containing 9 |
| SAUJ01045179.1_41814 | 0.003032678 | G/A | apol1 | apolipoprotein L, 1 |
| SAUJ01019207.1_771112 | 0.002956798 | C/T | inpp4aa | inositol polyphosphate-4-phosphatase type I Aa |
| SAUJ01035516.1_/ | 0.002949396 | A/G | Trap | transformation/transcription domain-associated protein |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shalgimbayeva, G.; Volkov, A.; Slobodova, N.; Sharko, F.; Tsygankova, S.; Boulygina, E.; Nguyen, V.Q.; Pham, T.T.; Nguyen, D.T.; Assylbekova, S.Z.; et al. Genetic Investigation of Aral Wild Common Carp Populations (Cyprinus carpio) Using ddRAD Sequencing. Diversity 2021, 13, 295. https://doi.org/10.3390/d13070295
Shalgimbayeva G, Volkov A, Slobodova N, Sharko F, Tsygankova S, Boulygina E, Nguyen VQ, Pham TT, Nguyen DT, Assylbekova SZ, et al. Genetic Investigation of Aral Wild Common Carp Populations (Cyprinus carpio) Using ddRAD Sequencing. Diversity. 2021; 13(7):295. https://doi.org/10.3390/d13070295
Chicago/Turabian StyleShalgimbayeva, Gulmira, Alexander Volkov, Natalia Slobodova, Fedor Sharko, Svetlana Tsygankova, Eugenia Boulygina, Van Q. Nguyen, The T. Pham, Duc T. Nguyen, Saule Zh. Assylbekova, and et al. 2021. "Genetic Investigation of Aral Wild Common Carp Populations (Cyprinus carpio) Using ddRAD Sequencing" Diversity 13, no. 7: 295. https://doi.org/10.3390/d13070295
APA StyleShalgimbayeva, G., Volkov, A., Slobodova, N., Sharko, F., Tsygankova, S., Boulygina, E., Nguyen, V. Q., Pham, T. T., Nguyen, D. T., Assylbekova, S. Z., Alekseev, Y., Nedoluzhko, A., Fernandes, J. M. O., & Rastorguev, S. (2021). Genetic Investigation of Aral Wild Common Carp Populations (Cyprinus carpio) Using ddRAD Sequencing. Diversity, 13(7), 295. https://doi.org/10.3390/d13070295