
Interface of Human/Wildlife Interactions: An Example of a Bold Coyote (Canis latrans) in Atlanta, GA, USA


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
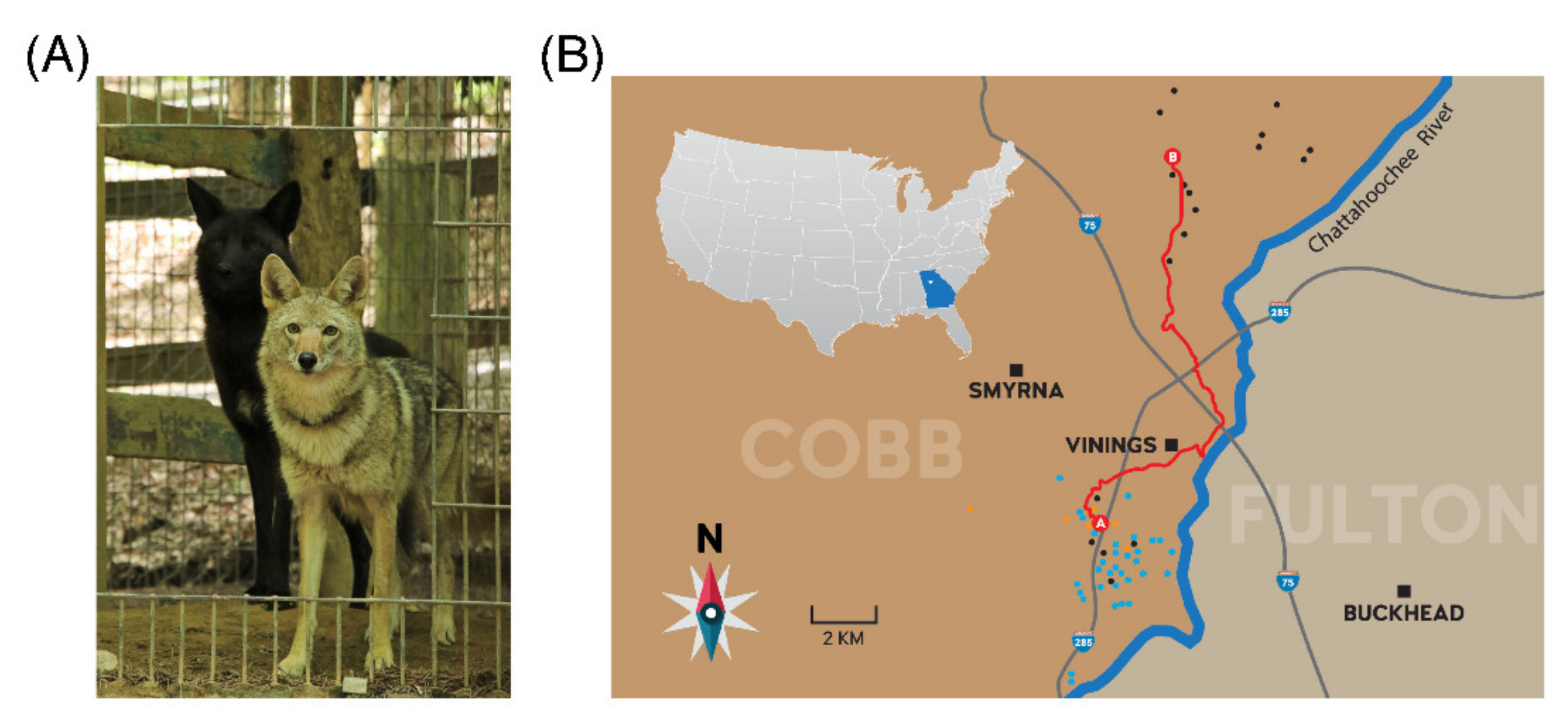
2.1. Tracking and Capture of Subject Animal
2.2. RAD Sequencing and Genomic Ancestry Analyses
2.3. Genotyping Variants Associated with Canine Human-Directed Hypersocial Behavior
2.4. Sequence Assay for the Deletion Associated with K Locus Melanism in Canids
3. Results
3.1. Behavior and Morphology
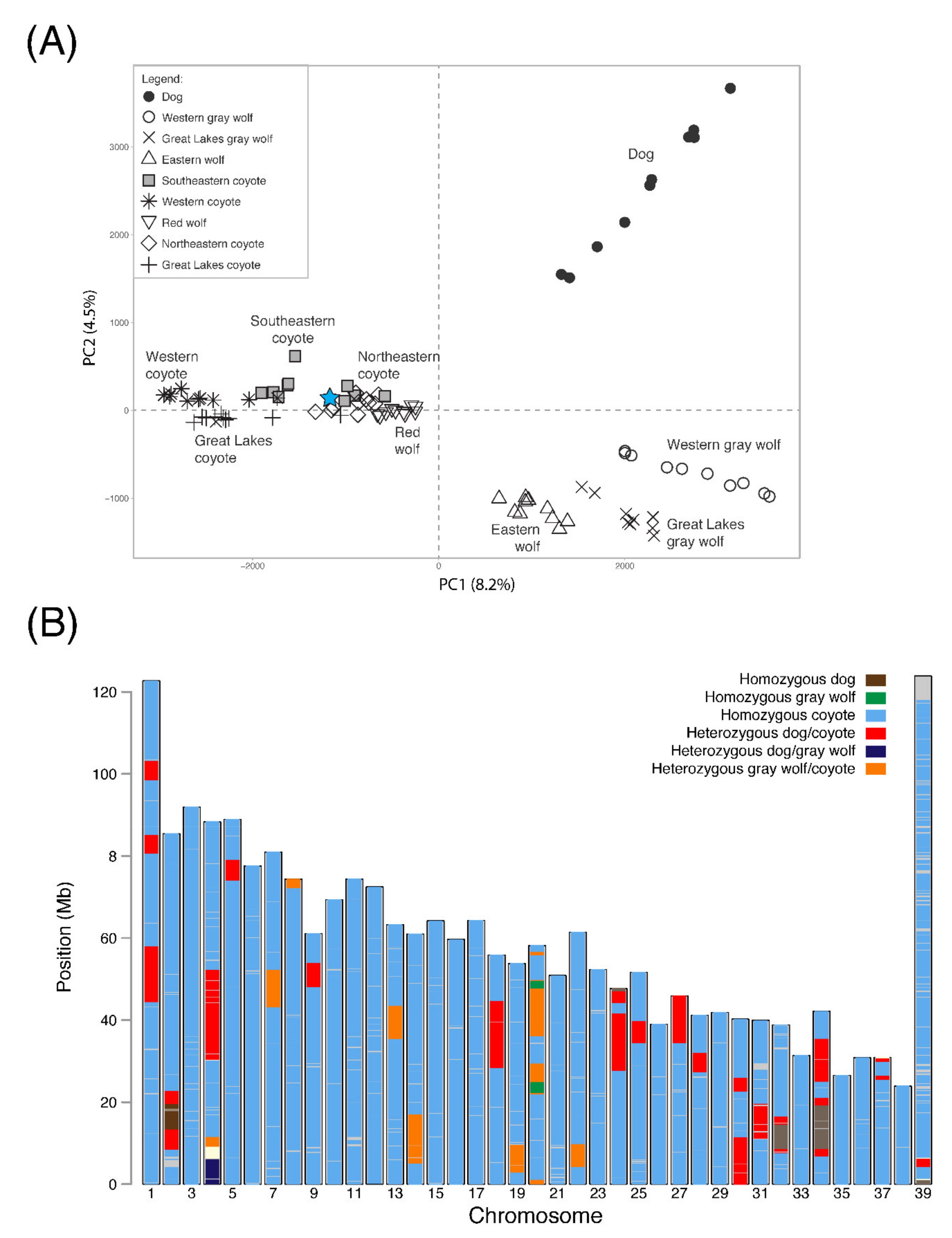
3.2. Ancestry
3.3. Canine Hypersociability and Melanism
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1. Detailed Materials and Methods
Appendix A.1.1. RAD Sequencing and Bioinformatic Processing
Appendix A.1.2. SNP Variant Discovery
Appendix A.1.3. Population Structure Analysis
Appendix A.1.4. Inference of Canid Ancestry
Appendix A.1.5. Genotyping Variants Associated with Canine Human-Directed Hypersocial Behavior
Appendix A.1.6. Sequence Assay for the Deletion Associated with K Locus Melanism in Canids
Appendix B
RADseq Results
References
- Frantz, L.A.F.; Bradley, D.G.; Larson, G.; Orlando, L. Animal Domestication in the Era of Ancient Genomics. Nat. Rev. Genet. 2020, 21, 449–460. [Google Scholar] [CrossRef]
- Li, Y.; vonHoldt, B.M.; Reynolds, A.; Boyko, A.R.; Wayne, R.K.; Wu, D.-D.; Zhang, Y.-P. Artificial Selection on Brain-Expressed Genes during the Domestication of Dog. Mol. Biol. Evol. 2013, 30, 1867–1876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akey, J.M.; Ruhe, A.L.; Akey, D.T.; Wong, A.K.; Connelly, C.F.; Madeoy, J.; Nicholas, T.J.; Neff, M.W. Tracking Footprints of Artificial Selection in the Dog Genome. Proc. Natl. Acad. Sci. USA 2010, 107, 1160–1165. [Google Scholar] [CrossRef] [Green Version]
- Axelsson, E.; Ratnakumar, A.; Arendt, M.-L.; Maqbool, K.; Webster, M.T.; Perloski, M.; Liberg, O.; Arnemo, J.M.; Hedhammar, Å.; Lindblad-Toh, K. The Genomic Signature of Dog Domestication Reveals Adaptation to a Starch-Rich Diet. Nature 2013, 495, 360–364. [Google Scholar] [CrossRef]
- vonHoldt, B.M.; Pollinger, J.P.; Lohmueller, K.E.; Han, E.; Parker, H.G.; Quignon, P.; Degenhardt, J.D.; Boyko, A.R.; Earl, D.A.; Auton, A.; et al. Genome-Wide SNP and Haplotype Analyses Reveal a Rich History Underlying Dog Domestication. Nature 2010, 464, 898–902. [Google Scholar] [CrossRef] [Green Version]
- Boyko, A.R.; Quignon, P.; Li, L.; Schoenebeck, J.J.; Degenhardt, J.D.; Lohmueller, K.E.; Zhao, K.; Brisbin, A.; Parker, H.G.; vonHoldt, B.M.; et al. A Simple Genetic Architecture Underlies Morphological Variation in Dogs. PLoS Biol. 2010, 8, e1000451. [Google Scholar] [CrossRef] [Green Version]
- Cadieu, E.; Neff, M.W.; Quignon, P.; Walsh, K.; Chase, K.; Parker, H.G.; vonHoldt, B.M.; Rhue, A.; Boyko, A.; Byers, A.; et al. Coat Variation in the Domestic Dog Is Governed by Variants in Three Genes. Science 2009, 326, 150–153. [Google Scholar] [CrossRef] [Green Version]
- vonHoldt, B.M.; Shuldiner, E.; Koch, I.J.; Kartzinel, R.Y.; Hogan, A.; Brubaker, L.; Wanser, S.; Stahler, D.; Wynne, C.D.L.; Ostrander, E.A.; et al. Structural Variants in Genes Associated with Human Williams-Beuren Syndrome Underlie Stereotypical Hypersociability in Domestic Dogs. Sci. Adv. 2017, 3, e1700398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Driscoll, C.A.; Macdonald, D.W.; O’Brien, S.J. From Wild Animals to Domestic Pets, an Evolutionary View of Domestication. Proc. Natl. Acad. Sci. USA 2009, 106, 9971–9978. [Google Scholar] [CrossRef] [Green Version]
- Price, E.O. Behavioral Aspects of Animal Domestication. Q. Rev. Biol. 1984, 59, 1–32. [Google Scholar] [CrossRef]
- Zeder, M.A. The Domestication of Animals. J. Anthropol. Res. 2012, 68, 161–190. [Google Scholar] [CrossRef] [Green Version]
- Coppinger, R.; Coppinger, L. Dogs: A New Understanding of Canine Origin, Behavior and Evolution; The University of Chicago: Chicago, IL, USA, 2002; ISBN 978-0-226-11563-4. [Google Scholar]
- Anderson, T.M.; vonHoldt, B.M.; Candille, S.I.; Musiani, M.; Greco, C.; Stahler, D.R.; Smith, D.W.; Padhukasahasram, B.; Randi, E.; Leonard, J.A.; et al. Molecular and Evolutionary History of Melanism in North American Gray Wolves. Science 2009, 323, 1339–1343. [Google Scholar] [CrossRef] [Green Version]
- Schweizer, R.M.; Durvasula, A.; Smith, J.; Vohr, S.H.; Stahler, D.R.; Galaverni, M.; Thalmann, O.; Smith, D.W.; Randi, E.; Ostrander, E.A.; et al. Natural Selection and Origin of a Melanistic Allele in North American Gray Wolves. Mol. Biol. Evol. 2018, 35, 1190–1209. [Google Scholar] [CrossRef] [Green Version]
- Hedrick, P.W. Wolf of a Different Colour. Heredity 2009, 103, 435–436. [Google Scholar] [CrossRef]
- Caudill, G.; Caudill, D. Melanism of Coyotes (Canis latrans) in Florida. AMID 2015, 174, 335–342. [Google Scholar] [CrossRef]
- Brockerville, R.M.; McGrath, M.J.; Pilgrim, B.L.; Marshall, H.D. Sequence Analysis of Three Pigmentation Genes in the Newfoundland Population of Canis latrans Links the Golden Retriever Mc1r Variant to White Coat Color in Coyotes. Mamm. Genome 2013, 24, 134–141. [Google Scholar] [CrossRef]
- Mowry, C.B.; Edge, J.L. Melanistic Coyotes in Northwest Georgia. SENA 2014, 13, 280–287. [Google Scholar] [CrossRef]
- Gipson, P.S. Melanistic Canis in Arkansas. Southwest. Nat. 1976, 21, 124–126. [Google Scholar] [CrossRef]
- vonHoldt, B.M.; Pollinger, J.P.; Earl, D.A.; Knowles, J.C.; Boyko, A.R.; Parker, H.; Geffen, E.; Pilot, M.; Jedrzejewski, W.; Jedrzejewska, B.; et al. A Genome-Wide Perspective on the Evolutionary History of Enigmatic Wolf-like Canids. Genome Res. 2011, 21, 1294–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heppenheimer, E.; Brzeski, K.; Wooten, R.; Waddell, W.; Rutledge, L.; Chamberlain, M.; Stahler, D.; Hinton, J.; vonHoldt, B. Rediscovery of Red Wolf Ghost Alleles in a Canid Population Along the American Gulf Coast. Genes 2018, 9, 618. [Google Scholar] [CrossRef] [Green Version]
- Heppenheimer, E.; Brzeski, K.E.; Hinton, J.W.; Chamberlain, M.J.; Robinson, J.; Wayne, R.K.; vonHoldt, B.M. A Genome-Wide Perspective on the Persistence of Red Wolf Ancestry in Southeastern Canids. J. Hered. 2020, 111, 277–286. [Google Scholar] [CrossRef]
- Bozarth, C.A.; Hailer, F.; Rockwood, L.L.; Edwards, C.W.; Maldonado, J.E. Coyote Colonization of Northern Virginia and Admixture with Great Lakes Wolves. J. Mammal. 2011, 92, 1070–1080. [Google Scholar] [CrossRef] [Green Version]
- Monzón, J.; Kays, R.; Dykhuizen, D.E. Assessment of Coyote-Wolf-Dog Admixture Using Ancestry-Informative Diagnostic SNPs. Mol. Ecol. 2014, 23, 182–197. [Google Scholar] [CrossRef] [Green Version]
- vonHoldt, B.M.; Cahill, J.A.; Fan, Z.; Gronau, I.; Robinson, J.; Pollinger, J.P.; Shapiro, B.; Wall, J.; Wayne, R.K. Whole-Genome Sequence Analysis Shows That Two Endemic Species of North American Wolf Are Admixtures of the Coyote and Gray Wolf. Sci. Adv. 2016, 2, e1501714. [Google Scholar] [CrossRef] [Green Version]
- Sinding, M.-H.S.; Gopalakrishan, S.; Vieira, F.G.; Samaniego Castruita, J.A.; Raundrup, K.; Heide Jørgensen, M.P.; Meldgaard, M.; Petersen, B.; Sicheritz-Ponten, T.; Mikkelsen, J.B.; et al. Population Genomics of Grey Wolves and Wolf-like Canids in North America. PLoS Genet. 2018, 14, e1007745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- vonHoldt, B.M.; Kays, R.; Pollinger, J.P.; Wayne, R.K. Admixture Mapping Identifies Introgressed Genomic Regions in North American Canids. Mol. Ecol. 2016, 25, 2443–2453. [Google Scholar] [CrossRef]
- Hody, J.W.; Kays, R. Mapping the Expansion of Coyotes (Canis latrans) across North and Central America. ZK 2018, 759, 81–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poessel, S.A.; Gese, E.M.; Young, J.K. Environmental Factors Influencing the Occurrence of Coyotes and Conflicts in Urban Areas. Landsc. Urban Plan. 2017, 259–269. [Google Scholar] [CrossRef] [Green Version]
- Gompper, M.E. Top Carnivores in the Suburbs? Ecological and Conservation Issues Raised by Colonization of North-Eastern North America by Coyotes. BioScience 2002, 52, 185–190. [Google Scholar] [CrossRef] [Green Version]
- Parker, G.R. Eastern Coyote: The Story of Its Success; Nimbus: Halifax, NS, Canada, 1995. [Google Scholar]
- Thurber, J.M.; Peterson, R.O. Changes in Body Size Associated with Range Expansion in the Coyote (Canis latrans). J. Mammal. 1991, 72, 750–755. [Google Scholar] [CrossRef]
- Hill, E.P.; Sumner, P.W.; Wooding, J.B. Human Influences on Range Expansion of Coyotes in the Southeast. Wildl. Soc. Bull. 1987, 15, 521–524. [Google Scholar]
- Koolhaas, J.M.; Korte, S.M.; De Boer, S.F.; Van Der Vegt, B.J.; Van Reenen, C.G.; Hopster, H.; De Jong, I.C.; Ruis, M.A.; Blokhuis, H.J. Coping Styles in Animals: Current Status in Behavior and Stress-Physiology. Neurosci. Biobehav. Rev. 1999, 23, 925–935. [Google Scholar] [CrossRef]
- Sloan Wilson, D.; Clark, A.B.; Coleman, K.; Dearstyne, T. Shyness and Boldness in Humans and Other Animals. Trends Ecol. Evol. 1994, 9, 442–446. [Google Scholar] [CrossRef]
- Ali, O.A.; O’Rourke, S.M.; Amish, S.J.; Meek, M.H.; Luikart, G.; Jeffres, C.; Miller, M.R. RAD Capture (Rapture): Flexible and Efficient Sequence-Based Genotyping. Genetics 2016, 202, 389–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catchen, J.; Hohenlohe, P.A.; Bassham, S.; Amores, A.; Cresko, W.A. Stacks: An Analysis Tool Set for Population Genomics. Mol. Ecol. 2013, 22, 3124–3140. [Google Scholar] [CrossRef] [Green Version]
- Rochette, N.C.; Rivera-Colón, A.G.; Catchen, J.M. Stacks 2: Analytical Methods for Paired—End Sequencing Improve RADseq—based Population Genomics. Mol. Ecol. 2019, 28, 4737–4754. [Google Scholar] [CrossRef] [PubMed]
- Lindblad-Toh, K.; Wade, C.M.; Mikkelsen, T.S.; Karlsson, E.K.; Jaffe, D.B.; Kamal, M.; Clamp, M.; Chang, J.L.; Kulbokas, E.J.; Zody, M.C.; et al. Genome Sequence, Comparative Analysis and Haplotype Structure of the Domestic Dog. Nature 2005, 438, 803–819. [Google Scholar] [CrossRef]
- Guan, Y. Detecting Structure of Haplotypes and Local Ancestry. Genetics 2014, 196, 625–642. [Google Scholar] [CrossRef] [Green Version]
- Johnson, N.A.; Coram, M.A.; Shriver, M.D.; Romieu, I.; Barsh, G.S.; London, S.J.; Tang, H. Ancestral Components of Admixed Genomes in a Mexican Cohort. PLoS Genet. 2011, 7, e1002410. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.R.; Adams, J.R.; Waits, L.P. Pedigree—Based Assignment Tests for Reversing Coyote (Canis latrans) Introgression into the Wild Red Wolf (Canis rufus) Population. Mol. Ecol. 2003, 12, 3287–3301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vonholdt, B.M.; Stahler, D.R.; Smith, D.W.; Earl, D.A.; Pollinger, J.P.; Wayne, R.K. The Genealogy and Genetic Viability of Reintroduced Yellowstone Grey Wolves. Mol. Ecol. 2008, 17, 252–274. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, P.W.; Peterson, R.O.; Vucetich, L.M.; Adams, J.R.; Vucetich, J.A. Genetic Rescue in Isle Royale Wolves: Genetic Analysis and the Collapse of the Population. Conserv. Genet. 2014, 15, 1111–1121. [Google Scholar] [CrossRef]
- Mech, L.D.; Barber-Meyer, S.M.; Erb, J. Wolf (Canis lupus) Generation Time and Proportion of Current Breeding Females by Age. PLoS ONE 2016, 11, e0156682. [Google Scholar] [CrossRef] [PubMed]
- Albers, G.; Edwards, J.W.; Rogers, R.E.; Mastro, L.L. Natality of Yearling Coyotes in West Virginia. J. Fish Wildl. Manag. 2016, 7, 192–197. [Google Scholar] [CrossRef] [Green Version]
- Kilgo, J.C.; Shaw, C.E.; Vukovich, M.; Conroy, M.J.; Ruth, C. Reproductive Characteristics of a Coyote Population before and during Exploitation: Reproduction of Southeastern Coyotes. Jour. Wild. Mgmt. 2017, 81, 1386–1393. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup The Sequence Alignment/Map Format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce Framework for Analyzing next-Generation DNA Sequencing Data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Poplin, R.; Ruano-Rubio, V.; DePristo, M.A.; Fennell, T.J.; Carneiro, M.O.; Van der Auwera, G.A.; Kling, D.E.; Gauthier, L.D.; Levy-Moonshine, A.; Roazen, D.; et al. Scaling Accurate Genetic Variant Discovery to Tens of Thousands of Samples. BioRxiv 2018, 201178. [Google Scholar] [CrossRef] [Green Version]
- Candille, S.I.; Kaelin, C.B.; Cattanach, B.M.; Yu, B.; Thompson, D.A.; Nix, M.A.; Kerns, J.A.; Schmutz, S.M.; Millhauser, G.L.; Barsh, G.S. A β-Defensin Mutation Causes Black Coat Color in Domestic Dogs. Science 2007, 318, 1418. [Google Scholar] [CrossRef] [Green Version]
- Gehrt, S.D.; Brown, J.L.; Anchor, C. Is the Urban Coyote a Misanthropic Synanthrope? The Case from Chicago. Cities Environ. (CATE) 2011, 4, 3. [Google Scholar]
- Ellington, E.H.; Gehrt, S.D. Behavioral Responses by an Apex Predator to Urbanization. Behav. Ecol. 2019, 30, 821–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larson, R.N.; Brown, J.L.; Karels, T.; Riley, S.P.D. Effects of Urbanization on Resource Use and Individual Specialization in Coyotes (Canis Latrans) in Southern California. PLoS ONE 2020, 15, e0228881. [Google Scholar] [CrossRef]
- Adducci, A., II; Jasperse, J.; Riley, S.; Brown, J.; Honeycutt, R.; Monzón, J. Urban Coyotes Are Genetically Distinct from Coyotes in Natural Habitats. J. Urban Ecol. 2020, 6, juaa010. [Google Scholar] [CrossRef]
- Jones, P.; Chase, K.; Martin, A.; Davern, P.; Ostrander, E.A.; Lark, K.G. Single-Nucleotide-Polymorphism-Based Association Mapping of Dog Stereotypes. Genetics 2008, 179, 1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Våge, J.; Wade, C.; Biagi, T.; Fatjó, J.; Amat, M.; Lindblad-Toh, K.; Lingaas, F. Association of Dopamine- and Serotonin-Related Genes with Canine Aggression. Genes Brain Behav. 2010, 9, 372–378. [Google Scholar] [CrossRef] [PubMed]
- MacLean, E.L.; Snyder-Mackler, N.; vonHoldt, B.M.; Serpell, J.A. Highly Heritable and Functionally Relevant Breed Differences in Dog Behaviour. Proc. R. Soc. B Biol. Sci. 2019, 286, 20190716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saetre, P.; Strandberg, E.; Sundgren, P.-E.; Pettersson, U.; Jazin, E.; Bergström, T.F. The Genetic Contribution to Canine Personality. Genes Brain Behav. 2006, 5, 240–248. [Google Scholar] [CrossRef]
- Svartberg, K. A Comparison of Behaviour in Test and in Everyday Life: Evidence of Three Consistent Boldness-Related Personality Traits in Dogs. Appl. Anim. Behav. Sci. 2005, 91, 103–128. [Google Scholar] [CrossRef]
- Fox, M.W. Socio-Ecological Implications of Individual Differences in Wolf Litters: A Developmental and Evolutionary Perspective. Behaviour 1972, 41, 298–313. [Google Scholar] [CrossRef]
- Breck, S.W.; Poessel, S.A.; Mahoney, P.; Young, J.K. The Intrepid Urban Coyote: A Comparison of Bold and Exploratory Behavior in Coyotes from Urban and Rural Environments. Sci. Rep. 2019, 9, 2104. [Google Scholar] [CrossRef]
- Young, J.K.; Mahe, M.; Breck, S. Evaluating Behavioral Syndromes in Coyotes (Canis latrans). J. Ethol. 2015, 33, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Lunter, G.; Goodson, M. Stampy: A Statistical Algorithm for Sensitive and Fast Mapping of Illumina Sequence Reads. Genome Res. 2011, 21, 936–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-Generation PLINK: Rising to the Challenge of Larger and Richer Datasets. GigaScience 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Abraham, G.; Inouye, M. Fast Principal Component Analysis of Large-Scale Genome-Wide Data. PLoS ONE 2014, 9, e93766. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.K.; Ruhe, A.L.; Dumont, B.L.; Robertson, K.R.; Guerrero, G.; Shull, S.M.; Ziegle, J.S.; Millon, L.V.; Broman, K.W.; Payseur, B.A.; et al. A Comprehensive Linkage Map of the Dog Genome. Genetics 2010, 184, 595–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.; Kim, J.; Kim, S.; Cook, D.E.; Evans, K.S.; Andersen, E.C.; Lee, J. Long-Read Sequencing Reveals Intra-Species Tolerance of Substantial Structural Variations and New Subtelomere Formation in C. elegans. Genome Res. 2019, 29, 1023–1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heppenheimer, E.; Harrigan, R.J.; Rutledge, L.Y.; Koepfli, K.-P.; DeCandia, A.L.; Brzeski, K.E.; Benson, J.F.; Wheeldon, T.; Patterson, B.R.; Kays, R.; et al. Population Genomic Analysis of North American Eastern Wolves (Canis lycaon) Supports Their Conservation Priority Status. Genes 2018, 9, 606. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mowry, C.B.; Wilson, L.A.; vonHoldt, B.M. Interface of Human/Wildlife Interactions: An Example of a Bold Coyote (Canis latrans) in Atlanta, GA, USA. Diversity 2021, 13, 372. https://doi.org/10.3390/d13080372
Mowry CB, Wilson LA, vonHoldt BM. Interface of Human/Wildlife Interactions: An Example of a Bold Coyote (Canis latrans) in Atlanta, GA, USA. Diversity. 2021; 13(8):372. https://doi.org/10.3390/d13080372
Chicago/Turabian StyleMowry, Christopher B., Lawrence A. Wilson, and Bridgett M. vonHoldt. 2021. "Interface of Human/Wildlife Interactions: An Example of a Bold Coyote (Canis latrans) in Atlanta, GA, USA" Diversity 13, no. 8: 372. https://doi.org/10.3390/d13080372
APA StyleMowry, C. B., Wilson, L. A., & vonHoldt, B. M. (2021). Interface of Human/Wildlife Interactions: An Example of a Bold Coyote (Canis latrans) in Atlanta, GA, USA. Diversity, 13(8), 372. https://doi.org/10.3390/d13080372