
Effects of Ocean Currents in the Western Pacific Ocean on Net-Phytoplankton Community Compositions


 , and
, and
Abstract
:1. Introduction
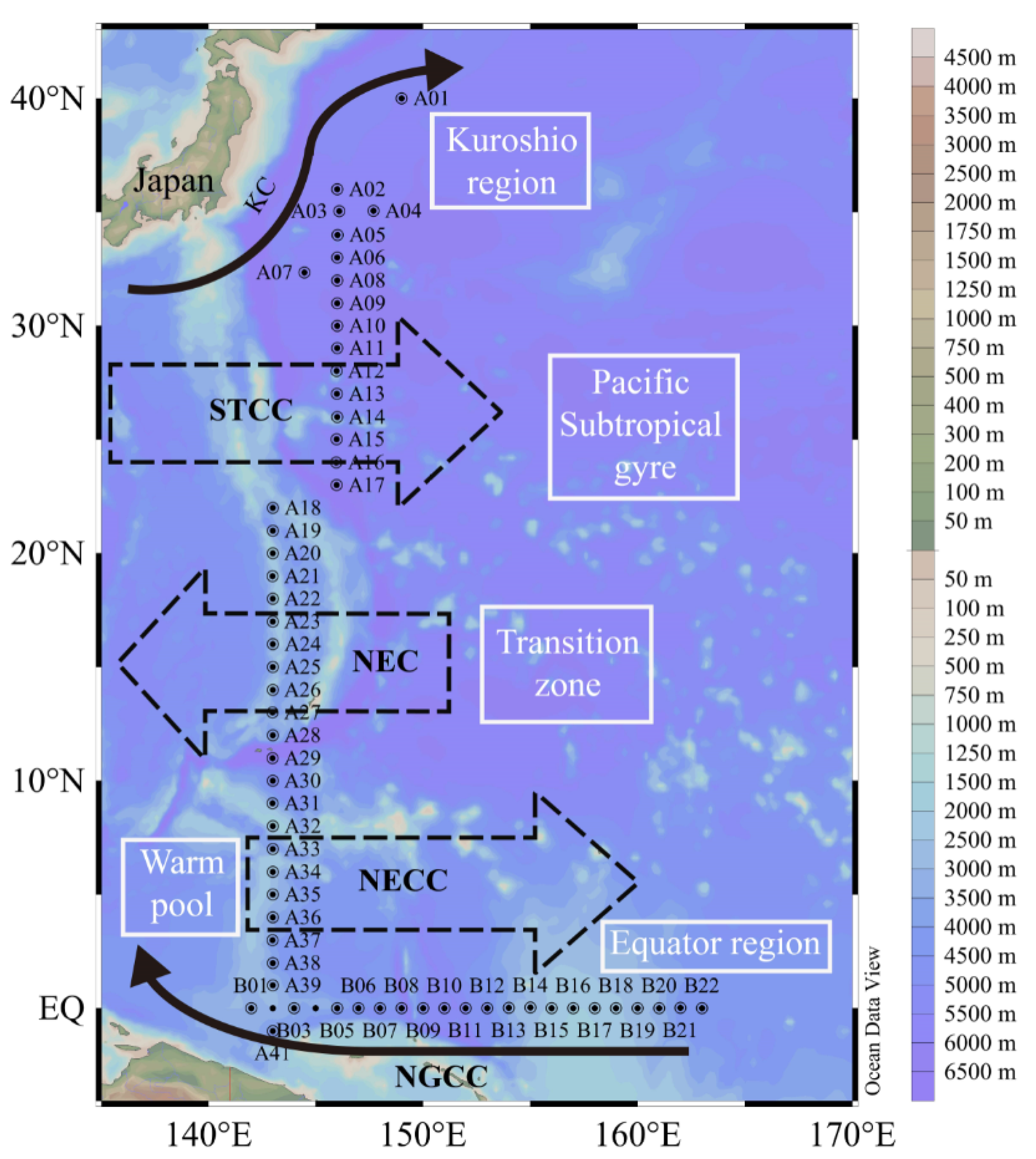
2. Material and Method
3. Result
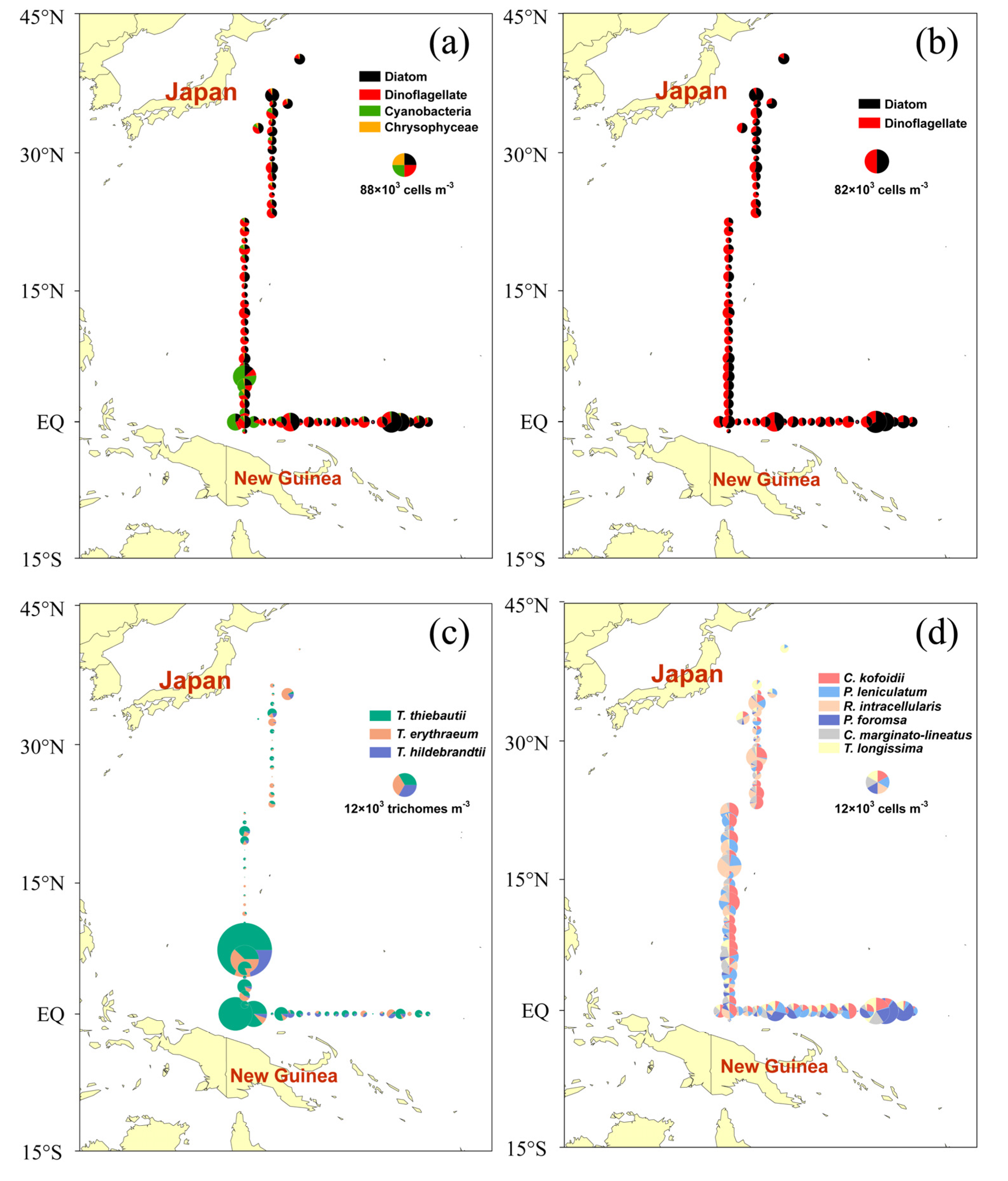
3.1. Phytoplankton Species Composition
3.2. Environmental Characteristics of the Survey Area
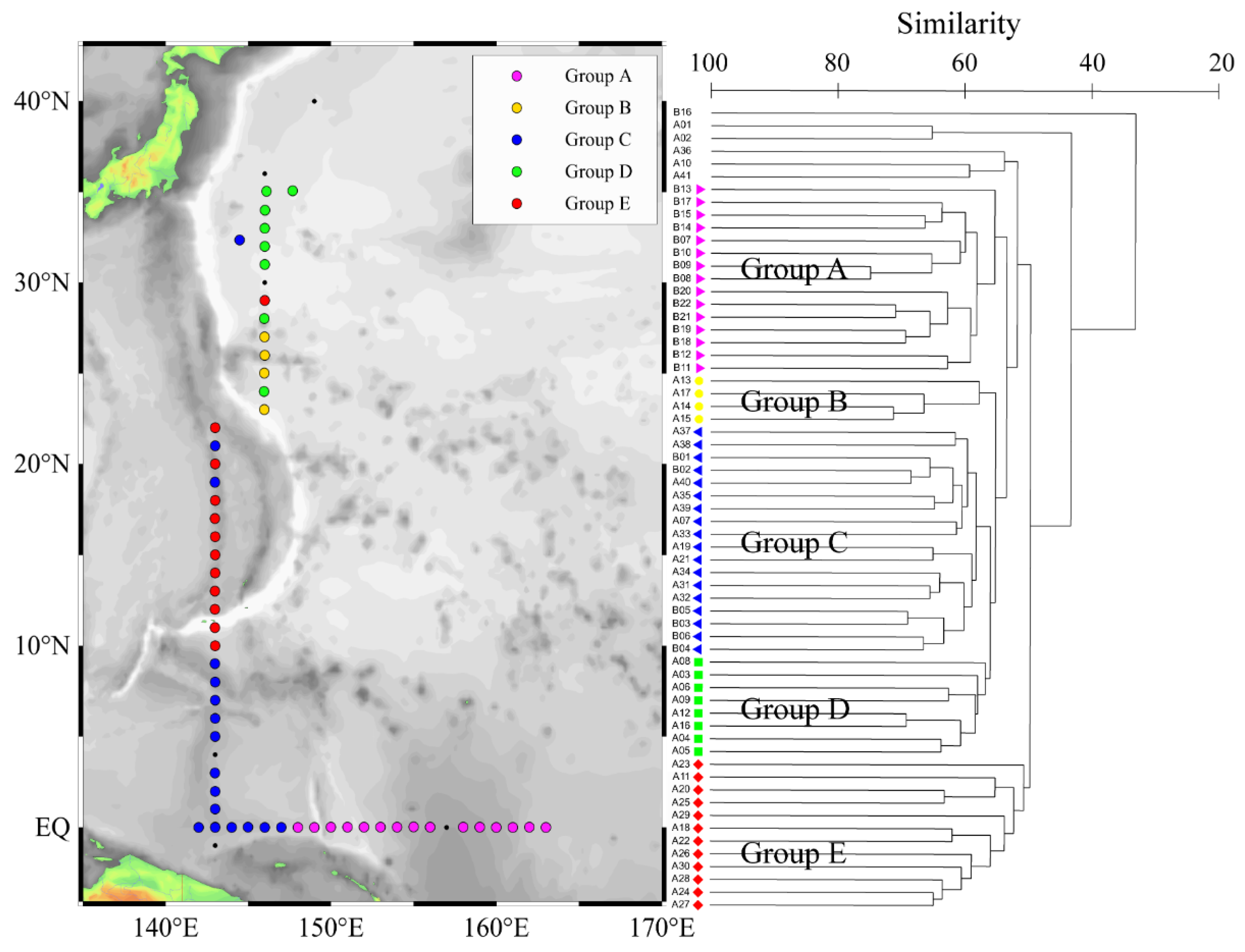
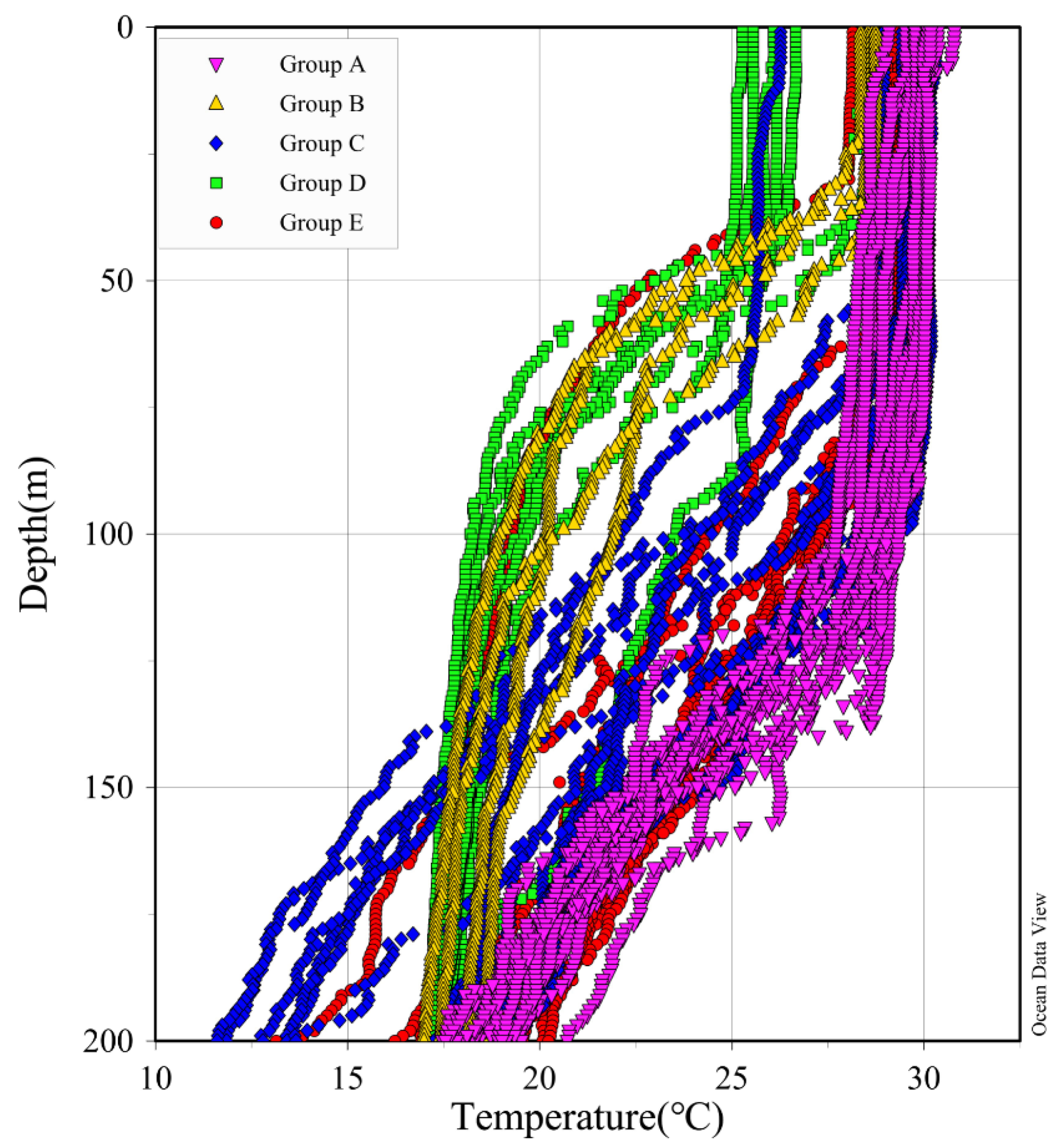
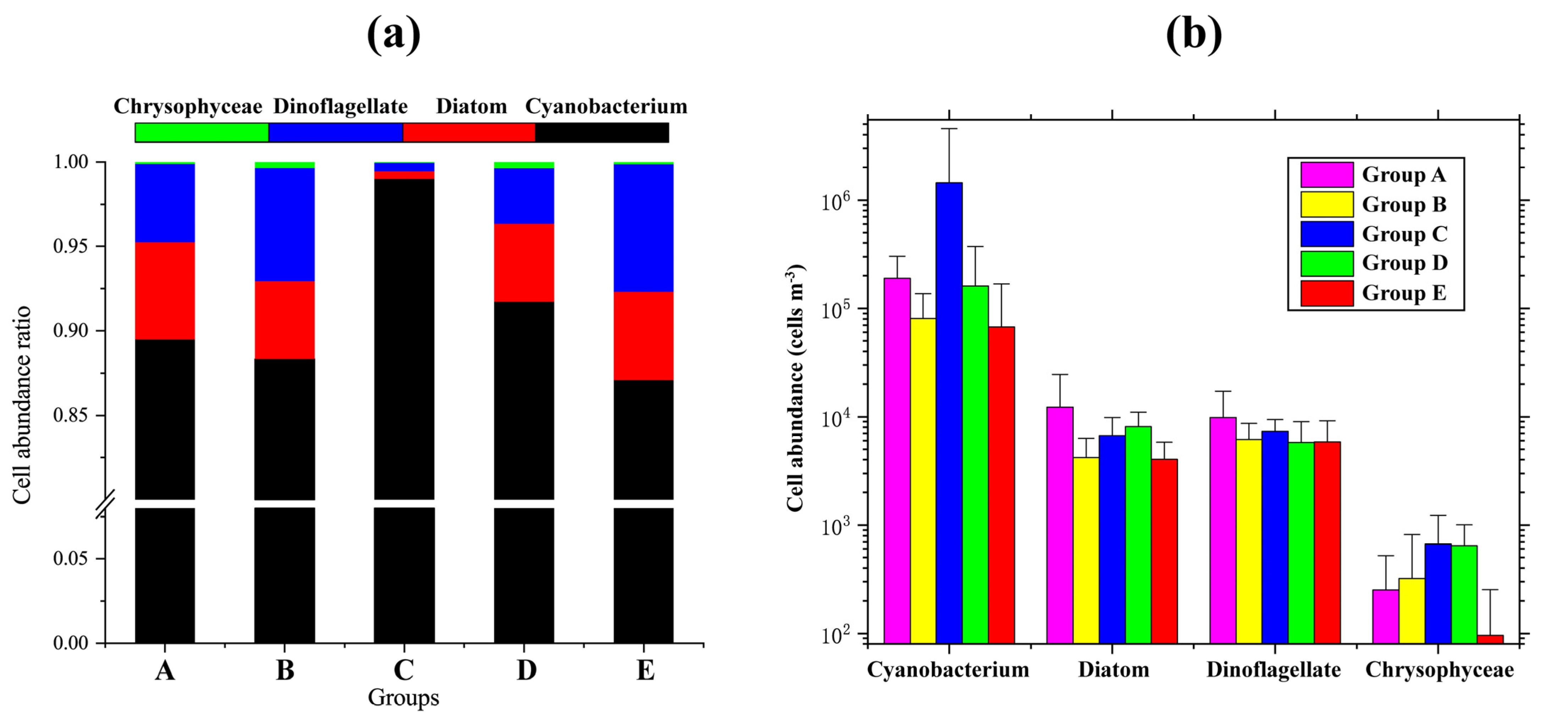
3.3. Phytoplankton Community Structure of Five Groups
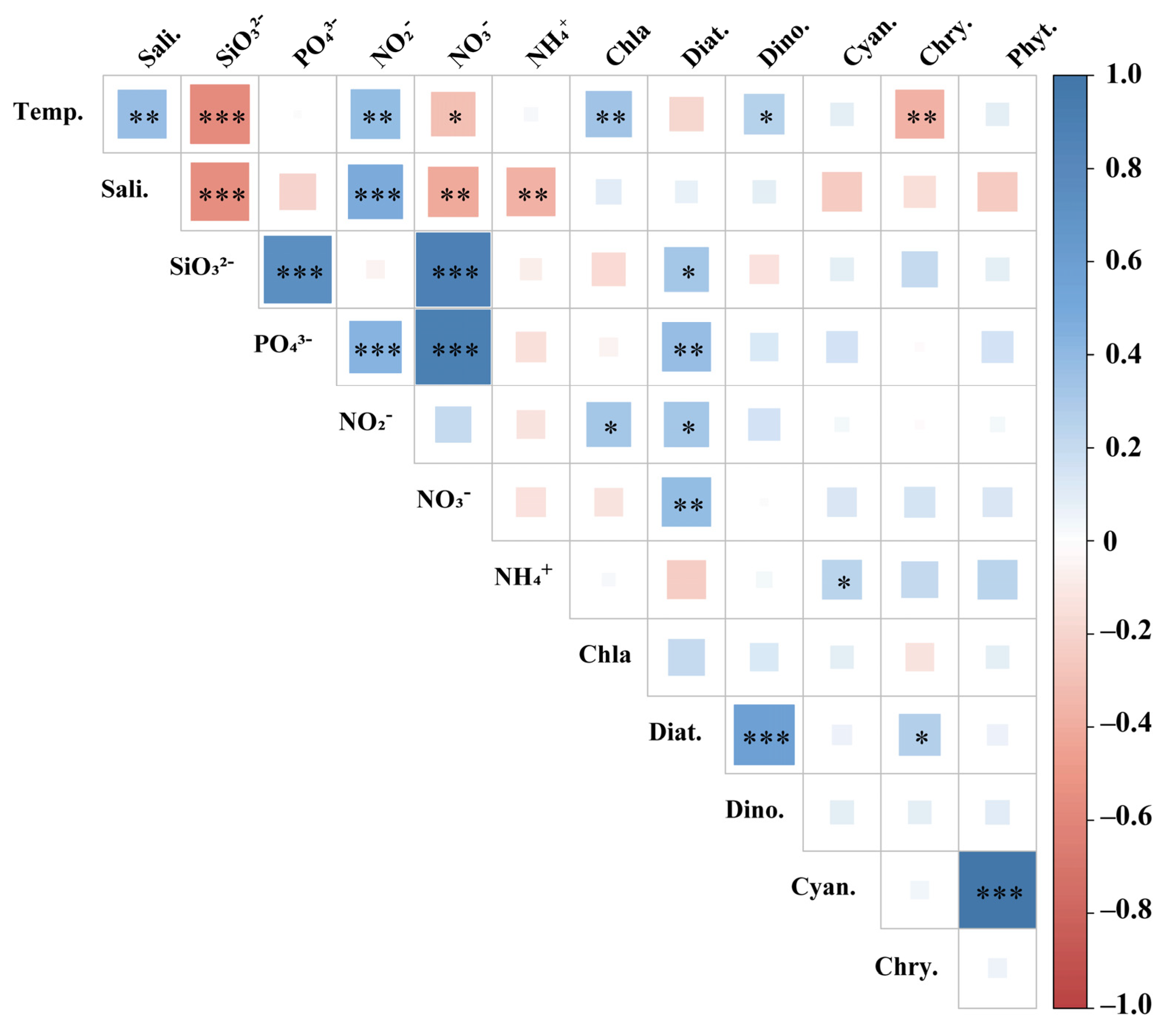
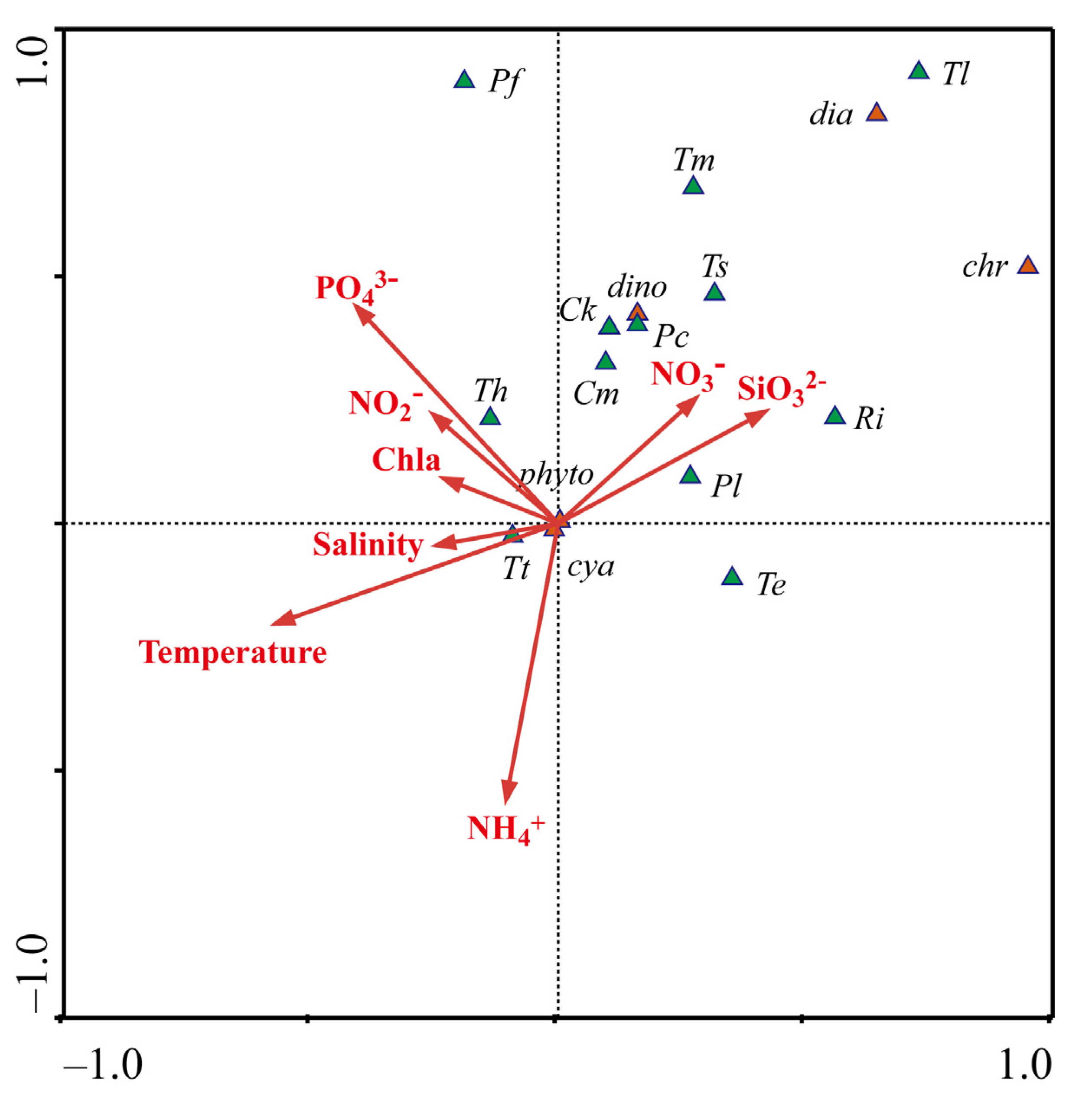
3.4. Phytoplankton Distribution in Relation to Environmental Factors
4. Discussion
4.1. Hydrological Conditions and Corresponding Phytoplankton Community Structure
4.2. Distribution of Trichodesmium
4.3. Dominant Species and Their Preferred Environmental Factors
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, J. Marine phytoplankton and biological carbon sink. Acta Ecol. Sin. 2011, 31, 5372–5378. [Google Scholar]
- Wang, Y.; Kang, J.; Ye, Y.; Lin, G.; Yang, Q.; Lin, M. Phytoplankton community and environmental correlates in a coastal upwelling zone along western Taiwan Strait. J. Mar. Syst. 2016, 154, 252–263. [Google Scholar] [CrossRef]
- Ahmed, A.; Kurian, S.; Gauns, M.; Chndrasekhararao, A.V.; Mulla, A.; Naik, B.; Naik, H.; Naqvi, S.W.A. Spatial variability in phytoplankton community structure along the eastern Arabian Sea during the onset of south-west monsoon. Cont. Shelf Res. 2016, 119, 30–39. [Google Scholar] [CrossRef]
- Okazaki, Y.; Nakata, H.; Kimura, S. Effects of frontal eddies on the distribution and food availability of anchovy larvae in the Kuroshio Extension. Mar. Freshw. Res. 2002, 53, 403. [Google Scholar] [CrossRef]
- Moita, M.T.; Pazos, Y.; Rocha, C.; Nolasco, R.; Oliveira, P.B. Toward predicting Dinophysis blooms off NW Iberia: A decade of events. Harmful Algae 2016, 53, 17–32. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Zhang, P.; Hu, D.X.; Seidel, H. Tropical Western Pacific circulation and seasonal changes. Sci. Bull. 2001, 46, 1998–2002. [Google Scholar] [CrossRef]
- Gordon, A.L.; Fine, R.A. Pathways of water between the Pacific and Indian oceans in the Indonesian seas. Nature 1996, 379, 146–149. [Google Scholar] [CrossRef]
- Yan, X.H.; Ho, C.R.; Zheng, Q.; Klemas, V. Temperature and size variabilities of the Western Pacific Warm Pool. Science 1992, 258, 1643–1645. [Google Scholar] [CrossRef]
- Kawarada, Y. Distribution of plankton in the waters neighboring Japan in 1966 (CSK). Oceanogr. Mag. 1968, 20, 187–212. [Google Scholar]
- Shiro, F. Illustrations of the Kuroshio-Waters; Chang Qi Publishing Culture Association: Hunan, China, 1990; pp. 1–170. [Google Scholar]
- Sun, J.; Liu, D.Y.; Qian, S.B. Planktonic diatoms in the Ryukyu Islands and adjacent waters-species composition and abundance distribution in summer 1997. In Chinese Oceanographic Anthology (II); Ocean University of China: Qingdao, China, 2000; pp. 170–182. (In Chinese) [Google Scholar]
- Liu, D.Y.; Sun, J.; Qian, S.B. Planktonic dinoflagellates in the Ryukyu Islands and adjacent waters-species composition and abundance distribution in summer 1997. In Chinese Oceanographic Anthology (II); Ocean University of China: Qingdao, China, 2000; pp. 158–169. (In Chinese) [Google Scholar]
- Chen, Y.; Sun, X.; Zhu, M.; Zheng, S.; Yuan, Y.; Denis, M. Spatial variability of phytoplankton in the Pacific western boundary currents during summer 2014. Mar. Freshw. Res. 2017, 68, 1887–1900. [Google Scholar] [CrossRef]
- Chen, Z.; Sun, J.; Zhang, G.C. Netz-phytoplankton community structure of the tropical Western Pacific Ocean in summer 2016. Mar. Sci. 2018, 42, 114–130. (In Chinese) [Google Scholar] [CrossRef]
- Sun, J.; Liu, D.Y. A quantative research and analysis method for marine phytoplankton: An introduction to Utermöhl method and Its modification. Adv. Mar. Sci. 2002, 20, 105–112. [Google Scholar] [CrossRef]
- Jin, D.X.; Chen, J.H. Chinese Marine Planktonic Diatoms; Shanghai Scientific & Technical Press: Shanghai, China, 1965; pp. 1–230. [Google Scholar]
- Isamu, Y. Illustrations of the Marine Plankton of Japan; Hoikusha Press: Tokyo, Japan, 1991; pp. 1–158. [Google Scholar]
- Sun, J.; Liu, D.Y. The Preliminary Notion on Nomenclature of Common Phytoplankton in China Sea Waters. Oceanol. Limnol. Sin. 2002, 33, 271–286. [Google Scholar] [CrossRef]
- Ehrhardt, M.A.; Kremling, K.; Almgren, T.; Grasshoff, K.M. Methods of seawater analysis. Chemie 1976. [Google Scholar] [CrossRef]
- Dai, M.; Wang, L.; Guo, X.; Zhai, W.; Li, Q.; He, B.; Kao, S.J. Nitrification and inorganic nitrogen distribution in a large perturbed river/estuarine system: The Pearl River estuary, China. Biogeosciences 2008, 5, 1227–1244. [Google Scholar] [CrossRef] [Green Version]
- Lorenzen, C.J. A method for the continuous measurement of in vivo chlorophyll concentration. Deep-Sea Res. Oceanogr. Abstr. 1966, 13, 223–227. [Google Scholar] [CrossRef]
- Hu, D.X.; Wu, L.X.; Cai, W.J.; Gupta, A.S.; Ganachaud, A.; Qiu, B.; Gordon, A.L.; Lin, X.P.; Chen, Z.H.; Shijian Hu, S.J.; et al. Pacific western boundary currents and their roles in climate. Nature 2015, 522, 299–308. [Google Scholar] [CrossRef]
- Lukas, R.; Firing, E.; Hacker, P.; Richardson, P.L.; Collins, C.A.; Fine, R.; Gammon, R. Observations of the Mindanao Current during the western equatorial Pacific Ocean circulation study. J. Geophys. Res. Oceans 1991, 96, 7089–7104. [Google Scholar] [CrossRef]
- Zhu, J.; Zheng, Q.N.; Hu, J.Y.; Lin, H.Y.; Chen, D.W.; Chen, Z.Z.; Sun, Z.Y.; Li, L.Y.; Kong, H. Classification and 3-D distribution of upper layer water masses in the northern South China Sea. Acta Oceanol. Sin. 2019, 38, 126–135. [Google Scholar] [CrossRef]
- Oksanen, J.F.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’ Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 20 August 2021).
- Cravatte, S.; Delcroix, T.; Zhang, D.; Mcphaden, M.; Leloup, J. Observed freshening and warming of the western Pacific Warm Pool. Clim. Dyn. 2009, 33, 565–589. [Google Scholar] [CrossRef]
- Kitajima, S.; Furuya, K.; Hashihama, F.; Takeda, S.; Kanda, J. Latitudinal distribution of diazotrophs and their nitrogen fixation in the tropical and subtropical western North Pacific. Limnol. Oceanogr. 2009, 54, 537–547. [Google Scholar] [CrossRef]
- Sekine, Y.; Miyamoto, S. Influence of Kuroshio Flow on the Horizontal Distribution of North Pacific Intermediate Water in the Shikoku Basin. J. Oceanogr. 2002, 58, 611–616. [Google Scholar] [CrossRef]
- Xie, L.; Tian, J.; Hu, D.X.; Wang, F. A quasi-synoptic interpretation of water mass distribution and circulation in the western North Pacific II: Circulation. Chin. J. Oceanol. Limnol. 2009, 27, 630–639. [Google Scholar] [CrossRef]
- Lukas, R.; Lindstrom, E. The mixed layer of the western equatorial Pacific Ocean. J. Geophys. Res. Oceans 1991, 96, 3343. [Google Scholar] [CrossRef]
- Mathias, G.; Hisayuki, A.; Fuminori, H. Phosphorus stress of microphytoplankton community in the western subtropical North Pacific. J. Plankton Res. 2013, 35, 146–157. [Google Scholar] [CrossRef] [Green Version]
- Jeong, H.J.; Yoo, T.D.; Kim, J.S.; Seong, K.A.; Kang, N.S.; Kim, T.H. Growth, feeding and ecological roles of the mixotrophic and heterotrophic dinoflagellates in marine planktonic food webs. Ocean Sci. J. 2010, 45, 65–91. [Google Scholar] [CrossRef] [Green Version]
- Greta, B.; Fabio, R.J.V.; Alberto, A.; Eric, P.; Pierella, K.; Ferrante, M.I.; Patrick, W.; Alessandra, R.; Chris, B.; Remo, S.; et al. Meta-omics reveals genetic flexibility of diatom nitrogen transporters in response to environmental changes. Mol. Biol. Evol. 2019, 11, 2522–2535. [Google Scholar] [CrossRef] [Green Version]
- Luigi, C.; Quentin, C.; Damien, E.; Amos, K.; Eric, P.; Juan, J.P.K.; Fabio, R.J.V.; Emilie, V.; Samuel, C.; Shruti, M.; et al. Community-Level Responses to Iron Availability in Open Ocean Plankton Ecosystems. Glob. Biogeochem. Cycles 2019, 33, 391–419. [Google Scholar] [CrossRef]
- Girault, M.; Arakawa, H.; Barani, A.; Ceccaldi, H.J.; Gregori, G. Distribution of ultraphytoplankton in the western part of the North Pacific subtropical gyre during a strong La Niña condition: Relations with the hydrological conditions. Biogeosciences 2013, 10, 5947–5965. [Google Scholar] [CrossRef] [Green Version]
- Estrada, M.; Berdalet, E. Phytoplankton in a turbulent world. Sci. Mar. 1997, 61, 125–140. [Google Scholar] [CrossRef]
- Thomas, W.H.; Gibson, C.H. Quantified small-scale turbulence inhibits a red tide dinoflagellate, Gonyaulax polyedra Stein. Deep. Sea Res. Part. A Oceanogr. Res. Pap. 1990, 37, 1583–1593. [Google Scholar] [CrossRef]
- Smayda, T.J. Harmful algal blooms: Their ecophysiology and general relevance to phytoplankton blooms in the sea. Limnol. Oceanogr. 1997, 42, 1137–1153. [Google Scholar] [CrossRef]
- Sullivan, J.M.; Swift, E.; Donaghay, P.L.; Rines, J.E.B. Small-scale turbulence affects the division rate and morphology of two red-tide dinoflagellates. Harmful Algae 2003, 2, 183–199. [Google Scholar] [CrossRef]
- Dugdale, R.C.; Menzel, D.W.; Ryther, J.H. Nitrogen fixation in the Sargasso Sea. Deep Sea Res. 1961, 7, 297–300. [Google Scholar] [CrossRef]
- Laroche, J.; Breitbarth, E. Importance of the diazotrophs as a source of new nitrogen in the ocean. J. Sea Res. 2005, 53, 67–91. [Google Scholar] [CrossRef]
- Karl, D.; Michaels, A.; Bergman, B.; Capone, D.; Carpenter, E.; Letelier, R.; Lipschultz, F.; Paerl, H.; Sigman, D.; Stal, L. Dinitrogen fixation in the world’s oceans. Biogeochemistry 2002, 57–58, 47–98. [Google Scholar] [CrossRef]
- Toby, T.; Emilio, M.; Poulton, A.J.; Bowie, A.R.; Harbour, D.S.; Woodward, E.M.S. Large-scale latitudinal distribution of Trichodesmium spp. in the Atlantic Ocean. J. Plankton Res. 2003, 58, 405–416. [Google Scholar] [CrossRef]
- Capone, D.G.; Zehr, J.P.; Paerl, H.W.; Bergman, B.; Carpenter, E.J. Trichodesmium, a Globally Significant Marine Cyanobacterium. Science 1997, 276, 1221–1229. [Google Scholar] [CrossRef] [Green Version]
- Campbell, L.; Carpenter, E.J.; Montoya, J.P.; Kustka, A.B.; Capone, D.G. Picoplankton community structure within and outside a Trichodesmium bloom in the southwestern Pacific Ocean. Vie Milieu 2005, 55, 185–195. [Google Scholar]
- Stal, L.J.; Severin, I.; Bolhuis, H. The Ecology of Nitrogen Fixation in Cyanobacterial Mats; Springer: New York, NY, USA, 2010; Volume 675, pp. 31–45. [Google Scholar] [CrossRef]
- Zehr, J.P.; Ward, B.B. Nitrogen Cycling in the Ocean: New Perspectives on Processes and Paradigms. Appl. Environ. Microbiol. 2002, 68, 1015–1024. [Google Scholar] [CrossRef] [Green Version]
- Bonnet, S.; Biegala, I.C.; Dutrieux, P.; Slemons, L.O.; Capone, D.G. Nitrogen fixation in the western equatorial Pacific: Rates, diazotrophic cyanobacterial size class distribution, and biogeochemical significance. Glob. Biogeochem. Cycles 2009, 23, 45–62. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.S.; Wang, C.S.; Wang, X.G.; Liu, Z.S.; Gao, X.H. The large-scale distribution of Trichodesmium in the tropical and subtropical Pacific Ocean and the southern Indian Ocean. Acta Oceanol. Sin. 2010, 32, 148–154. [Google Scholar] [CrossRef]
- Chang, J.; Chiang, K.P.; Gong, G.C. Seasonal variation and cross-shelf distribution of the nitrogen-fixing cyanobacterium, Trichodesmium, in southern East China Sea. Cont. Shelf Res. 2000, 20, 479–492. [Google Scholar] [CrossRef]
- Karl, D.M.; Tien, G. Temporal variability in dissolved phosphorus concentrations in the subtropical North Pacific Ocean. Mar. Chem. 1997, 56, 77–96. [Google Scholar] [CrossRef]
- Capone, D.; Carpente, E. Nitrogen fixation by marine cyanobacteria: Historical and global perspectives. Bull. Linstitut Océanogr. 1999, 19, 235–256. [Google Scholar]
- Polat, S.; Koray, T. Planktonic dinoflagellates of the northern Levantine Basin, northeastern Mediterranean Sea. Eur. J. Protistol. 2007, 43, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, E.J.; Montoya, J.P.; Burns, J.A.; Mulholland, M.R.; Capone, D.G. Extensive bloom of a N2-fixing diatom/cyanobacterial association in the Tropical Atlantic Ocean. Mar. Ecol. Prog. Ser. 1999, 185, 273–283. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum | Genera | Species | Ratio of Richness (%) |
|---|---|---|---|
| Bacillariophyta | 37 | 105 | 31.63 |
| Dinophyta | 28 | 221 | 66.57 |
| Cyanophyta | 2 | 4 | 1.20 |
| Chrysophyta | 1 | 2 | 0.60 |
| Species | Percentage (%) | Frequency (fi) | Dominance (Y) | Maximum (103 Cells m−3) | Average (103 Cells m−3) | Maximum Stations |
|---|---|---|---|---|---|---|
| T. thiebautii | 65.67 | 0.952 | 0.6254 | 8895.529 | 374.425 | A35 |
| Trichodesmium erythraeum | 19.46 | 0.889 | 0.1730 | 2128.430 | 110.979 | A36 |
| Trichodesmium hildebrandtii | 12.07 | 0.746 | 0.0900 | 2782.329 | 68.807 | A35 |
| C. kofoidii | 0.18 | 0.952 | 0.0017 | 5.619 | 1.037 | A28 |
| Prorocentrum leniculatum | 0.15 | 1.000 | 0.0015 | 3.300 | 0.845 | A36 |
| Richelia intracellularis | 0.14 | 0.698 | 0.0010 | 7.947 | 0.805 | A24 |
| P. foromsa | 0.12 | 0.794 | 0.0010 | 13.276 | 0.688 | B19 |
| Coscinodiscus marginato-lineatus | 0.10 | 0.968 | 0.0009 | 2.844 | 0.542 | B18 |
| Thalassiosira subtilis | 0.09 | 0.968 | 0.0009 | 3.433 | 0.512 | B07 |
| Prorocentrum compressum | 0.08 | 1.000 | 0.0007 | 1.836 | 0.425 | B07 |
| T. longissima | 0.08 | 0.937 | 0.0007 | 6.542 | 0.445 | B18 |
| Thalassiosira minima | 0.07 | 1.000 | 0.0007 | 2.540 | 0.415 | B19 |
| Group A | Group B | Group C | Group D | Group E | |
|---|---|---|---|---|---|
| PO43− | 0.37 ± 0.07 | 0.13 ± 0.08 | 0.27 ± 0.10 | 0.16 ± 0.06 | 0.13 ± 0.05 |
| NO2− | 0.23 ± 0.12 | 0.05 ± 0.01 | 0.12 ± 0.04 | 0.06 ± 0.01 | 0.09 ± 0.02 |
| NO3− | 2.66 ± 0.87 | 0.70 ± 0.52 | 2.15 ± 1.07 | 2.06 ± 0.79 | 0.66 ± 0.47 |
| NH4+ | 0.87 ± 0.21 | 1.32 ± 0.43 | 1.51 ± 0.56 | 0.86 ± 0.09 | 1.49 ± 0.62 |
| SiO32− | 1.83 ± 0.29 | 2.00 ± 0.17 | 2.23 ± 1.07 | 2.62 ± 0.60 | 1.57 ± 0.45 |
| temperature | 28.24 ± 0.45 | 23.50 ± 0.46 | 27.29 ± 1.49 | 22.72 ± 1.08 | 26.89 ± 1.48 |
| salinity | 35.06 ± 0.18 | 34.98 ± 0.11 | 34.54 ± 0.24 | 34.80 ± 0.08 | 34.62 ± 0.31 |
| chl a | 0.41 ± 0.11 | 0.27 ± 0.46 | 0.34 ± 0.08 | 0.31 ± 0.08 | 0.27 ± 0.08 |
| Group A | Group B | Group C | Group D | Group E | |
|---|---|---|---|---|---|
| Phytoplankton | 211,796 ± 129,387 | 91,379 ± 60,250 | 1,454,796 ± 3,127,957 | 175,003 ± 211,356 | 77,120 ± 101,440 |
| Diatoms | 12,176 ± 12,454 | 4188 ± 2124 | 6664 ± 3171 | 8109 ± 2944 | 4022 ± 1794 |
| Dinoflagellates | 9820 ± 7323 | 6136 ± 2530 | 7306 ± 2106 | 5748 ± 3264 | 5834 ± 3357 |
| Cyanobacteria | 189,548 ± 114,153 | 80,735 ± 55,865 | 1,440,161 ± 3,126,653 | 160,503 ± 210,271 | 67,168 ± 100,680 |
| Chrysophyceae | 252 ± 268 | 321 ± 496 | 665 ± 560 | 643 ± 361 | 96 ± 157 |
| T. thiebautii | 96,835 ± 102,200 | 44,784 ± 34,253 | 1,093,840 ± 2,302,347 | 23,845 ± 21,450 | 37,684 ± 64,710 |
| T. erythraeum | 37,156 ± 53,088 | 33,982 ± 41,793 | 162,062 ± 292,659 | 119,011 ± 173,439 | 23,111 ± 35,473 |
| T. hildebrandtii | 55,556 ± 49,123 | 1486 ± 2972 | 183,590 ± 649,917 | 15,922 ± 33,384 | 4509 ± 14,216 |
| C. kofoidii | 965 ± 908 | 1312 ± 830 | 1251 ± 921 | 918 ± 1059 | 1261 ± 1652 |
| P. leniculatum | 782 ± 629 | 177 ± 190 | 867 ± 612 | 495 ± 371 | 1443 ± 993 |
| R. intracellularis | 0 ± 0 | 483 ± 278 | 669 ± 690 | 1725 ± 2011 | 1865 ± 2174 |
| P. foromsa | 2389 ± 3473 | 55 ± 52 | 334 ± 261 | 83 ± 58 | 9 ± 16 |
| C. marginato-lineatus | 632 ± 812 | 545 ± 318 | 719 ± 583 | 259 ± 255 | 519 ± 434 |
| T. subtilis | 705 ± 978 | 473 ± 223 | 598 ± 498 | 536 ± 685 | 199 ± 163 |
| P. compressum | 524 ± 448 | 373 ± 388 | 482 ± 403 | 266 ± 183 | 428 ± 353 |
| T. longissima | 988 ± 1594 | 64 ± 63 | 388 ± 377 | 230 ± 110 | 65 ± 99 |
| T. minima | 583 ± 679 | 413 ± 309 | 403 ± 426 | 513 ± 306 | 300 ± 209 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Z.; Sun, J.; Chen, D.; Wang, S.; Yu, H.; Chen, H.; Wang, M. Effects of Ocean Currents in the Western Pacific Ocean on Net-Phytoplankton Community Compositions. Diversity 2021, 13, 428. https://doi.org/10.3390/d13090428
Chen Z, Sun J, Chen D, Wang S, Yu H, Chen H, Wang M. Effects of Ocean Currents in the Western Pacific Ocean on Net-Phytoplankton Community Compositions. Diversity. 2021; 13(9):428. https://doi.org/10.3390/d13090428
Chicago/Turabian StyleChen, Zhuo, Jun Sun, Dawei Chen, Shihao Wang, Hao Yu, Hongtao Chen, and Min Wang. 2021. "Effects of Ocean Currents in the Western Pacific Ocean on Net-Phytoplankton Community Compositions" Diversity 13, no. 9: 428. https://doi.org/10.3390/d13090428
APA StyleChen, Z., Sun, J., Chen, D., Wang, S., Yu, H., Chen, H., & Wang, M. (2021). Effects of Ocean Currents in the Western Pacific Ocean on Net-Phytoplankton Community Compositions. Diversity, 13(9), 428. https://doi.org/10.3390/d13090428