1. Introduction
The Australian monsoonal tropics, encompassing the vast tropical savanna landscapes of the northern third of the continent (
Figure 1), is a centre of exceptional but largely unrecognized ant diversity. Many taxa that are formally recognized as single, widespread species are in fact hyperdiverse species complexes [
1,
2]. For example,
Melophorus rufoniger Heterick, Castalanelli and Shattuck was recently described as a single species occurring throughout mainland Australia but most commonly in the monsoonal tropics [
3]. However, a subsequent analysis that integrated genetic, morphological and distributional information revealed that at least 30 species within the taxon occur in the Top End (high rainfall northern third) of the Northern Territory (NT) alone. It was concluded that the total
M. rufoniger fauna included up to 100 species from monsoonal Australia, none of which are described [
4].
The
Monomorium nigrius Forel group is another case in point. It is an intractably diverse assemblage of very small, brownish-black species with 11-segmented antenna occurring throughout mainland Australia but with its centre of diversity in the monsoonal north [
5,
6]. In a recent revision of the Australian
Monomorium fauna the group was described as representing a single species,
M. fieldi Forel [
7], despite morphological variation that is obviously interspecific.
Six general morphotypes, each with multiple species, can be recognized based on variation in body size, length of antennal scapes, shape of the propodeum and petiolar node, and pilosity (the ‘
fieldi’, ‘
donisthorpei’, ‘sp. 50’, ‘sp. 14’, ‘sp. 13’ and ‘sp. 9’ morphotypes, using the species nomenclature of [
8]:
Figure 2). A key to the morphotypes is as follows:
Antennal scapes relatively long, reaching occipital margin or nearly so ….……………………………………………………………………………………………………………....... 1
Antennal scapes relatively short, failing to reach occipital margin by a distance greater than their maximum width ……….………………………………………………………. 4
Mesosoma with particularly long hairs; metanotal notch deep and propodeum rather prominently rounded; petiole often unusually broad in profile (
Figure 2a,b) …‘...
fieldi’
Not as above …………………………………………………………………………...….…………………………………………………………………………………………………………2
Relatively large and robust, often with a squarish head; metanotal notch feeble and propodeum not at all prominently rounded (
Figure 2c,d) ……..…….……… ‘
donisthorpei’
Smaller, with a rectangular head; metanotal notch more pronounced and propodeum more rounded (
Figure 2e,f) …………………………………………………….…… ‘sp. 50’
Petiolar node very small, lower than long in profile .…..……………………….….………………………………………………………………………………………………………….. 5
Petiolar node as high as long in profile (
Figure 2g,h) ..…………………………………………………………………………………………………………………………………. ‘sp. 14’
Tiny species, propodeum short and obliquely angled in profile (
Figure 2i,j) .……………………………………………………………………………………………………….. ‘sp. 13’
Larger species; propodeum more broadly rounded in profile (
Figure 2k,l) …..……………………………………………………………………………………………………… ‘sp. 9’
The different morphotypes commonly occur in close sympatry; indeed, nine species shown to be differentiated genetically and morphologically, and representing all six morphotypes, have been recorded from a savanna woodland site (Territory Wildlife Park) near Darwin [
8]. Remarkably, seven of these species were recorded from a single 10 × 10 m plot and six in another. If so many species of the
M. nigrius group can occur at such a small spatial scale, and the species group occurs throughout most of mainland Australia, then how many species are there in total? A recent compilation of the known ant fauna of the Top End of the NT lists 17 species from the group [
9], but otherwise there has been no attempt at a broader quantification of total diversity within the taxon.
In this paper, we present an integrated genetic (CO1), morphological and distributional analysis of the M. nigrius group in order to provide an estimate of the total number of species within it and to document spatial patterns of species richness and turnover. We specifically address the following questions:
What are indicative levels of total diversity based on available morphological, CO1 and distributional information?
What is the extent of congruence between the six general morphotypes and CO1 phylogeny?
How diverse is the M. nigrius-group fauna within Australia, and what are the spatial patterns of species richness and turnover?
2. Materials and Methods
This study was based on pinned specimens of the
M. nigrius group held in the ant collection at the CSIRO laboratory in Darwin, which holds by far the most extensive collection of the taxon. For CO1 analysis we used the 40
M. nigrius-group sequences from [
8] and obtained sequences from an additional 361 specimens collected from throughout mainland Australia (
Supplementary Table S1). One of these (from urban Sydney) is a perfect morphological match with the New Zealand species
M. antipodum Forel [
10], and we refer to it as
M.?
antipodum. We also sequenced a specimen of the closely related
M. carinatum Heterick group to be used as the outgroup for building a CO1 tree. Geographic coverage of samples within mainland Australia was extremely patchy (
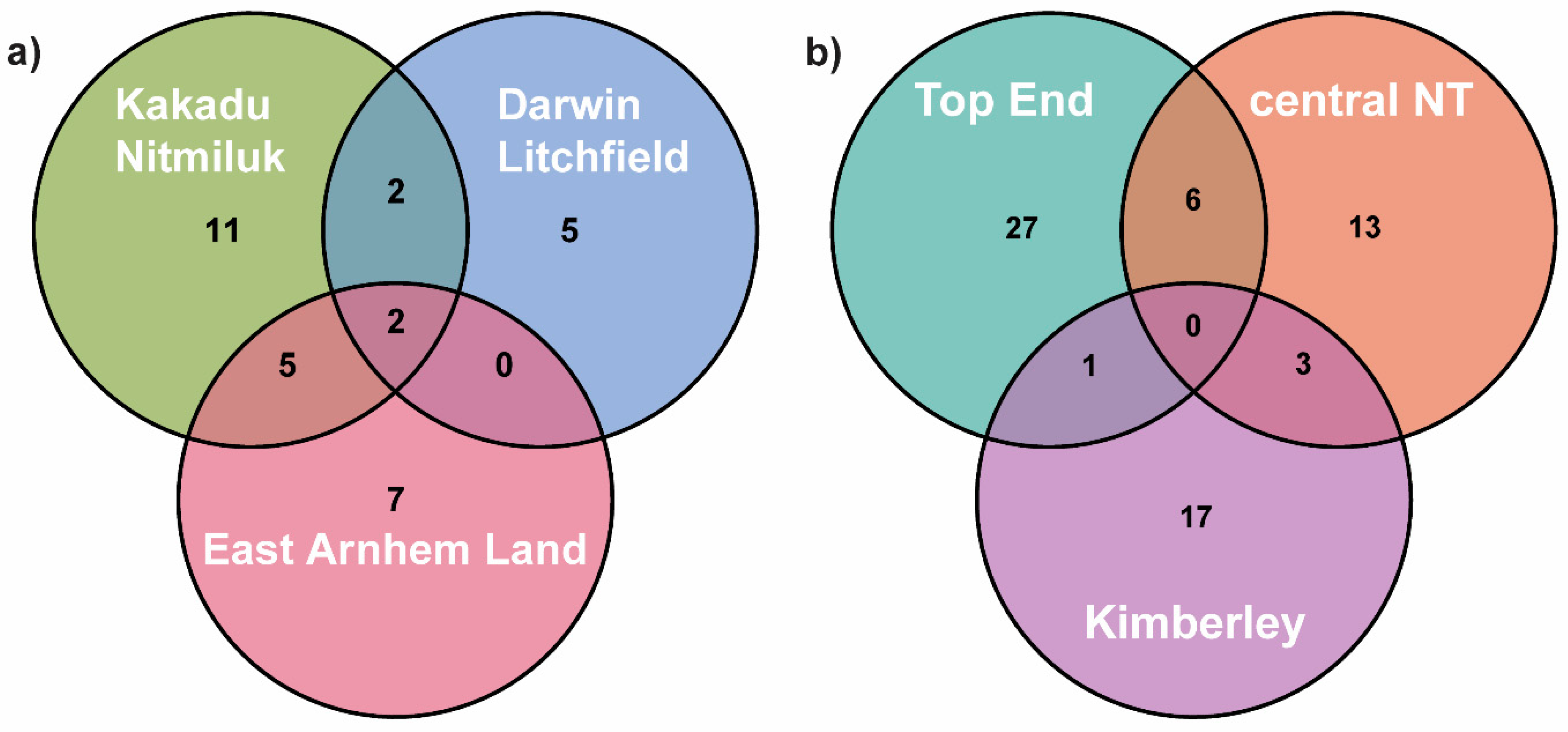
Figure 1). The most intensively sampled region was the Top End of the NT (1000–2000 mm mean annual rainfall), but even here large areas are unrepresented. Other regions of relatively high sampling intensity within monsoonal Australia are the Sturt Plateau region of central Northern Territory (550–800 mm), the Kimberley region of far northern Western Australia (500–1700 mm) and far North Queensland (north of the Townsville region) (
Figure 1). Vast areas of central and southern Australia are not represented.
Many localities represent multiple sites. The four major biogeographic regions of relatively high collection effort in northern Australia are indicated. Total annual rainfall in the monsoonal zone ranges from approximately 2000 mm on the Tiwi Islands in the Top End to 500 mm on the southern boundary with the central arid zone.
DNA extraction (from foreleg or whole-body tissue) and CO1 sequencing were conducted through the Barcode of Life Data (BOLD) System (for extraction details, see
http://ccdb.ca/resources, accessed on 4 January 2022). Each sequenced specimen was assigned a unique identification code that combines the batch within which it was processed, its number within the batch and the year of sequencing (e.g., MONO244-16). All specimens are labeled with their respective BOLD identification numbers in the Darwin collection.
DNA sequences were checked and edited in MEGA [
11]. Sequences were aligned using the UPGMB clustering method in MUSCLE [
12], and then translated into (invertebrate) proteins to check for stop codons and nuclear paralogues. The aligned sequences were trimmed accordingly, resulting in 822 base pairs.
To explore overall CO1 diversity in the samples, the mean genetic pairwise distances between sequences were calculated in MEGA. This was done using the Kimura-2 parameter (K2P) model [
13] to ensure that results were comparable with those of most other studies of insect DNA barcoding, with 500 bootstrap replicates and the ‘pairwise deletion’ option of missing data (to remove all ambiguous positions for each sequences pair). Analysis involved all nucleotide sequences, excluding those of the outgroup. Codon positions included were 1st + 2nd + 3rd.
The level of CO1 variation within ant species is typically 1–3% [
14] but there is no specific level of CO1 divergence that can be used to define a species. For species delimitation we adopted the species concept based on reproductive isolation and evolutionary independence as evidenced by morphological differentiation between sister (i.e., most closely related) clades (considering all available samples from the same collections as those of sequenced specimens) and sympatric distribution. We thus delimited species based on the integration of morphological variation, CO1 clustering and distance, and geographic distribution [
15]. We compared our species delimitations using such an integrated approach with two statistical methods using CO1 data alone. We used the MEGA genetic distances to produce a tree file with IQ-TREE [
16] and then ran this into two models. The first was the Poisson Tree Processes (PTP) model, which infers species boundaries using the number of substitutions within and between species in a maximum likelihood tree [
17]. The second was the Bayesian implementation of the PTP model (bPTP), which adds Bayesian values to delimited species on the input tree [
17]. We subjected trees including all specimens, as well as a tree of each major clade within the full tree separately, to PTP and bPTP algorithms on the web server (
http://species.h-its.org/ptp/, accessed on 8 October 2021), using the settings of 500,000 MCMC generations, 100 thinning and 0.1 burn in. We elected to increase the number of MCMC generations from 100,000 to 500,000 to increase the rate of convergence for the MCMC chain. Nevertheless, we did not reach convergence for several clades, and thus discarded these results.
We imaged representative specimens using a Leica DMC4500 camera mounted on a Leica M205C dissecting microscope. We took image montages using the Leica Application suite v. 4.13 and stacked them in Zerene stacker.
4. Discussion
We have revealed remarkable hyperdiversity within the Monomorium nigrius group, recognizing 97 species from limited geographic coverage of sequenced specimens. PTP analysis of CO1 clustering suggests that this figure is conservative. Given (1) the high levels of spatial turnover, (2) the fact that much of the taxon’s range remains unsampled, (3) that nearly one-third of the species are known from single records, and (4) many additional species (that are too old to yield sequences) are held in the Darwin collection, the sequenced specimens are likely to represent just a fraction of total diversity within the group.
We acknowledge that our sampling is limited when viewed at a continental scale, but we do not believe that this significantly affects our species delimitations. More than one-third (34) of our 97 recognized species occurs in a single region (Top End of the NT), and this region has been the most intensively sampled. The Top End fauna has very limited overlap with those of the region immediately south (central NT) that connects the Top End with the rest of Australia, which has also been intensively sampled. This indicates that further sampling would not show that species from other regions that we have recognized as different from those from the Top End are in fact conspecific. A detailed examination of sister relationships among our recognized species (
Figure S1) further supports our view that our species delimitations are not an artefact of limited sampling. For example, we recognize 20 species (all of the ‘
fieldi’ morphotype) in clade A. A1 is represented by ten specimens, all of which occur in southwestern western Australia. Its two sister species (A2 and A3) are from the Top End of the NT (furthest north) rather than from central or southern NT (which connect the Top End to southern Australia). These three species belong to a clade that includes two other, clearly distinct species, one (A4;
Figure 4a) known only from the Top End and the other (A5;
Figure 4b) occurring both in the Top End and central NT. The sister to this clade is another species from southwestern Australia (A6;
Figure 4c), which obviously cannot be the same as A1. Similar reasoning can be applied to other species.
The 34 species that we recognize from the Top End of the NT is twice as many as listed by [
9], and it does not include several species on that list (
Figure 11). Many of the species appear to have narrow ranges. For example, species E1, H1 and L are all known only from the same one site near Darwin [
8], and species G1 is known only within a 20 km range from that site. Given the limited spatial coverage of samples (
Figure 1), it is likely that the total Top End fauna comprises at least 50 species.
We recorded far fewer species in the two other high-rainfall regions of northern Australia: the Kimberley with 21 species, and far North Queensland with only seven. However, these figures are proportionate to sampling effort (
Figure 1,
Table S1) and there is no reason to believe that the faunas of these regions are substantially less diverse than in the Top End. Given the very little species overlap among them, the three regions collectively can be expected to have around 150 species. The high diversity (22 species) of the Sturt Plateau subregion of central NT is presumably repeated throughout the semi-arid north of the continent, suggesting that over 200 species occur just in monsoonal Australia. Diversity is also high in semi-arid central and southern Australia, with virtually no overlap with the monsoonal fauna, and so a reasonable estimate of the total
M. nigrius-group fauna is a phenomenally high 300 species.
An analysis of other (especially nuclear) genes is required for testing the robustness of the deeper clade structure within our CO1 tree. However, the high concordance between CO1 clade structure and our six previously recognized morphotypes suggests that they have a strong phylogenetic basis. Clade A contained all specimens of the ‘
fieldi’ morphotype, and so this is likely to represent a phylogenetically robust species complex. However, all other morphotypes are shown in the CO1 tree as polyphyletic. The ‘
donisthorpei’ morphotype occurs in two (disjunct) clades, one including ‘
donisthorpei’ in the strict sense (clade C, with three recognized species), and the other including sp. 37 from [
8] (clade D, with five recognized species). Despite their close morphological affinity (
Figure 5) they likely represent separate species complexes. Notably, not only did two of the three ‘
donisthorpei’ species from clade C occur at the same site but they did so in the same 10 × 10 m plot (plot 3 in [
8]). One of these (C2;
Figure 5b) is known only from the Top End, whereas the other (C1;
Figure 2c,d) occurs also in the Kimberley region.
The ‘sp. 14’, ‘sp. 13’ and ‘sp. 9’ morphotypes appear to be particularly diverse phylogenetically. The ‘sp. 14’ morphotype occurs as three (G, I and J) of the ten major clades and is distributed throughout mainland Australia. Clade J consists exclusively of species from outside the monsoonal zone, ranging throughout southern semi-arid Australia from central Queensland to southwestern Western Australia. The only other sequenced specimens from southern Australia are from the ‘fieldi’ morphotype, one species (A6) of which is from southwestern Western Australia, and a distantly related species (A20) occurring throughout semi-arid southeastern Australia. Both are shown in the CO1 tree as being most closely related to (different) species from central NT.
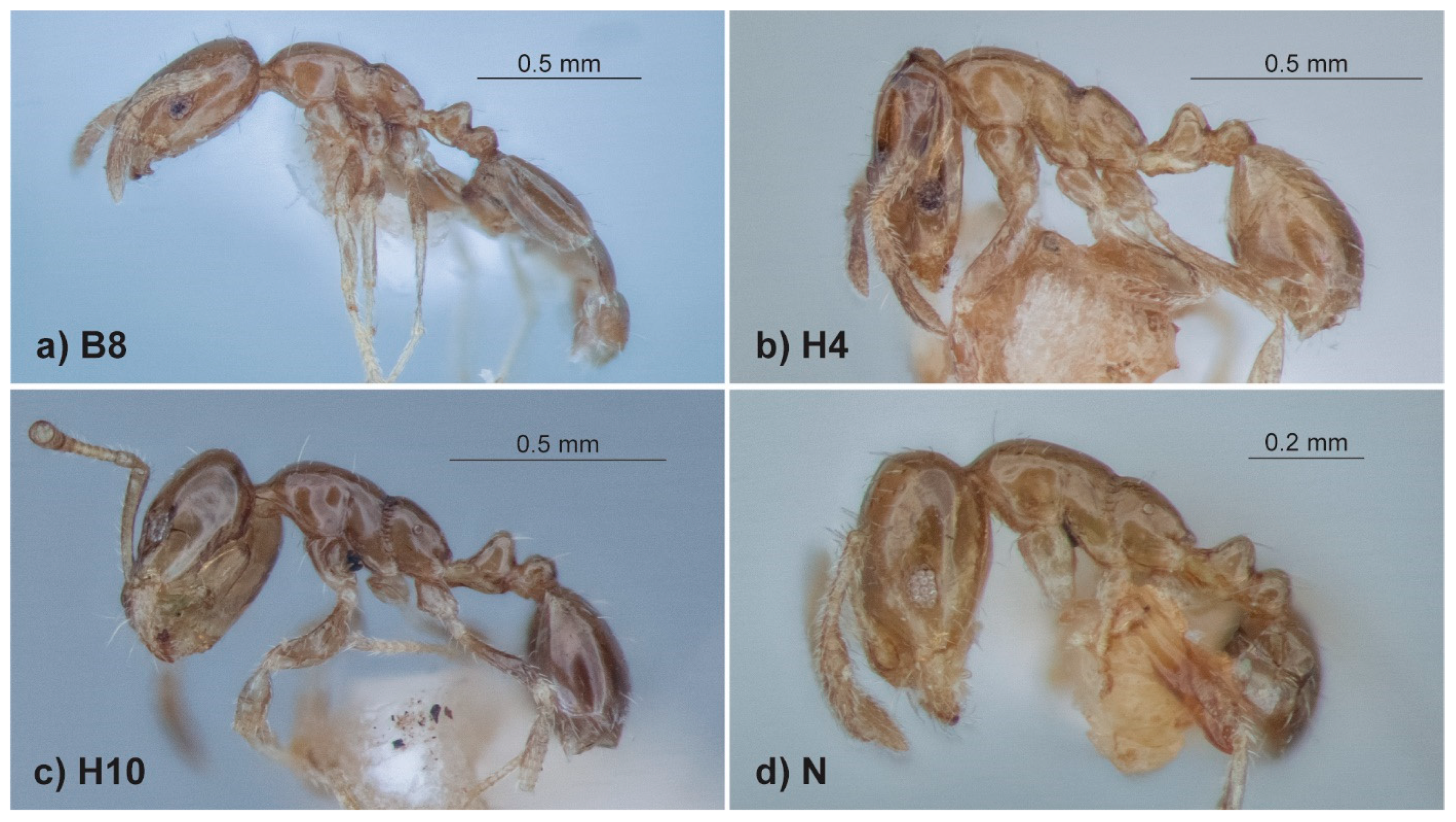
The ‘sp. 13’ morphotype is heavily concentrated in just one (H) of the ten major clades, but it is also represented in clade B (B8) and in two other locations on the CO1 tree (sp. M and sp. N;
Figure 3). This strongly indicates that despite being highly distinctive and relatively uniform (
Figure 2i,j and
Figure 8), the morphotype has evolved multiple times. The ‘sp. 9’ morphotype occurs in three locations (clades F and K and sp. L) on the CO1 tree and these are associated with conspicuous morphological differences, suggesting that they represent three separate species complexes. Species L (sp. 9 from [
8]) is unique among known species within the
M. nigrius group in being somewhat polymorphic (with head size and shape showing considerable allometric variation) and having an anterior clypeal margin that is only weakly convex (
Figure 2k). The two species from clade K (
Figure 9c,d) are unusual in having a mesosoma that is conspicuously sculptured postero-laterally; the unsequenced sp. 64 (
Figure 11c) shares this trait and presumably belongs to this complex.
Our M. ?antipodum sample is an outlier on the CO1 tree. It was collected from suburban Sydney and the only other specimens in the Darwin collection that match it morphologically are from suburban Brisbane. Together, this strongly suggests that it is introduced and is indeed M. antipodum from New Zealand, where it is commonly associated with human settlements (Don 2007).
5. Conclusions
What are the implications of our findings for total richness within the ant fauna of monsoonal Australia? Two decades ago, the fauna was estimated to contain approximately 1500 species, which at the time seemed remarkably high [
6]. In that analysis, the
Melophorus rufoniger group (then referred to as the
M. aeneovirens group, before
M. rufoniger was described) was estimated to contain ten species and the
Monomorium nigrius group twenty, estimates that have now been shown to be an order of magnitude too low. The more recent analysis of the ant fauna of the Top End of the NT recognized 901 native species, with a remarkable 60% of these apparently endemic to the region [
9]. Subsequent surveys in the Top End (e.g., [
18]) have recorded over 100 additional species. Detailed analyses of the
Melophorus rufoniger and
Monomorium nigrius groups have revealed that richness in these taxa is at least twice as high as was then recognized, and that levels of regional endemism are far higher than 60%. Our unpublished CO1 data show that this is true for many other species groups within
Melophorus and
Monomorium, as well as in other genera such as
Tetramorium,
Rhytidoponera,
Meranoplus,
Camponotus and
Iridomyrmex. Given the highly patchy sampling within the Top End (
Figure 1), we estimate that its total ant fauna comprises at least 1300 species.
Ant diversity and endemism are also exceptionally high in the Kimberley region of far northern Western Australia [
19,
20,
21]. Other biogeographical regions in monsoonal Australia have been even less intensively sampled, but their levels of species richness and endemism are also likely to be extremely high (see [
21] for a broader discussion of this). This means that the total monsoonal fauna likely numbers in the several thousand.
Such diversity is truly remarkable for a tropical savanna landscape. For example, although the ant fauna of the similarly sized Brazilian savanna (‘cerrrado’) is considered particularly diverse [
22], it is estimated to comprise only approximately 700 species (R. Feitosa, personal communication). Peak ant diversity globally is generally considered to occur in lowland tropical rainforest and especially in Amazonia [
23]. However, our analysis suggests that monsoonal Australia may in fact be the true global centre of ant diversity. How can such remarkable richness—and that of arid Australia more generally, be explained? It is presumably a product of historical processes given that the contemporary Australian environment is not so dramatically different from elsewhere in the world. The remarkable diversity occurs within species complexes rather than at the genus level, indicating that it was generated over recent evolutionary time. One explanation [
24] is that it is a product of the Pleistocene glaciations that caused massive movement of sand across Australia during these times of peak aridity, when up to 85% of the continent was covered by desert dunes. Such dunes are hostile for most ant species, whose distributions would have retracted to isolated refugia scattered within the vast sand-dominated landscape, allowing for speciation on a mass scale.
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}