
Insight into the Taxonomic Resolution of Apiospora: Introducing Novel Species and Records from Bamboo in China and Thailand

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, Fungal Isolation and Morphological Examination
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Phylogenetic Analyses
2.4. Host and Geographical Distribution of Apiospora Species
3. Results
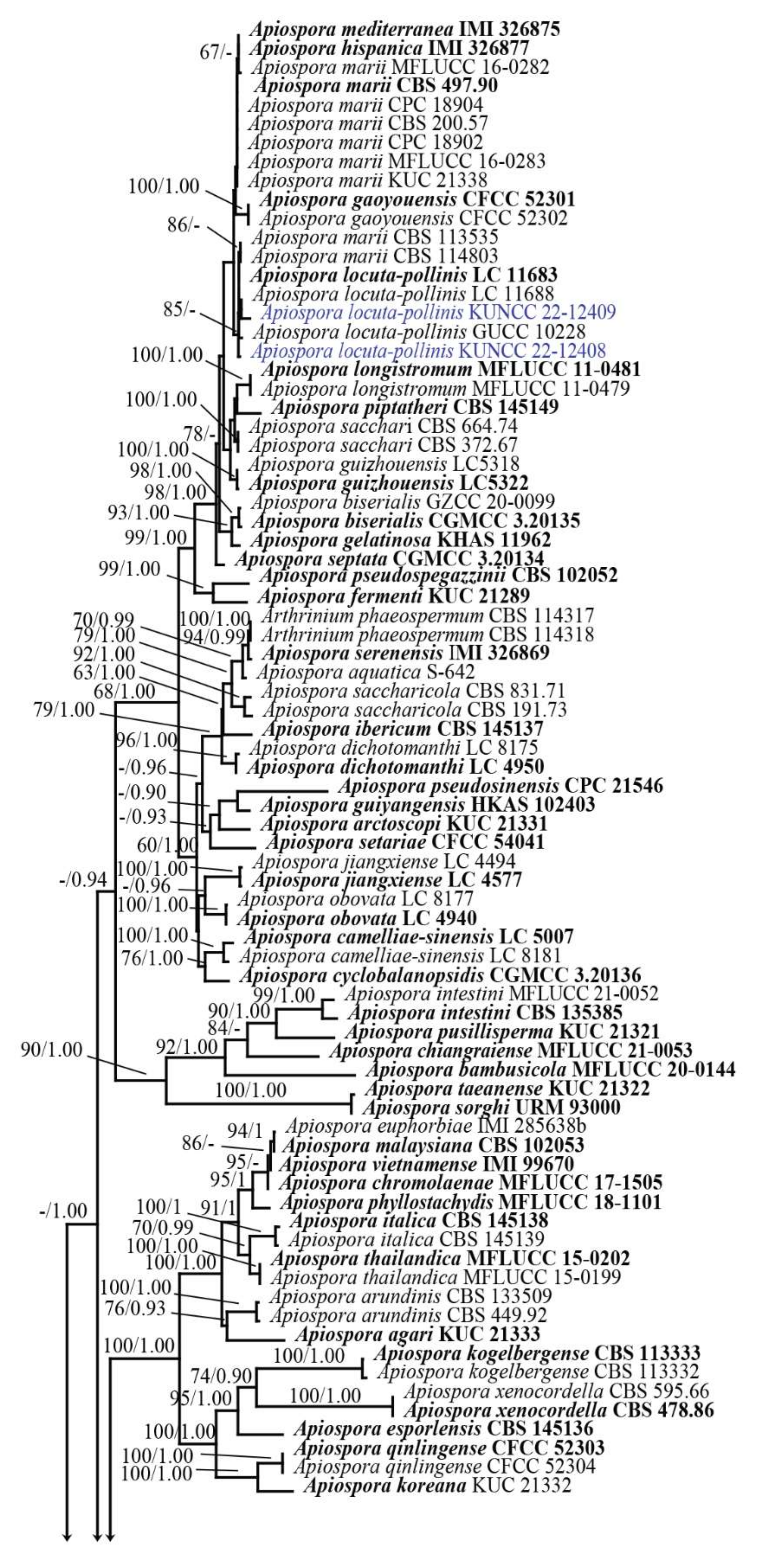
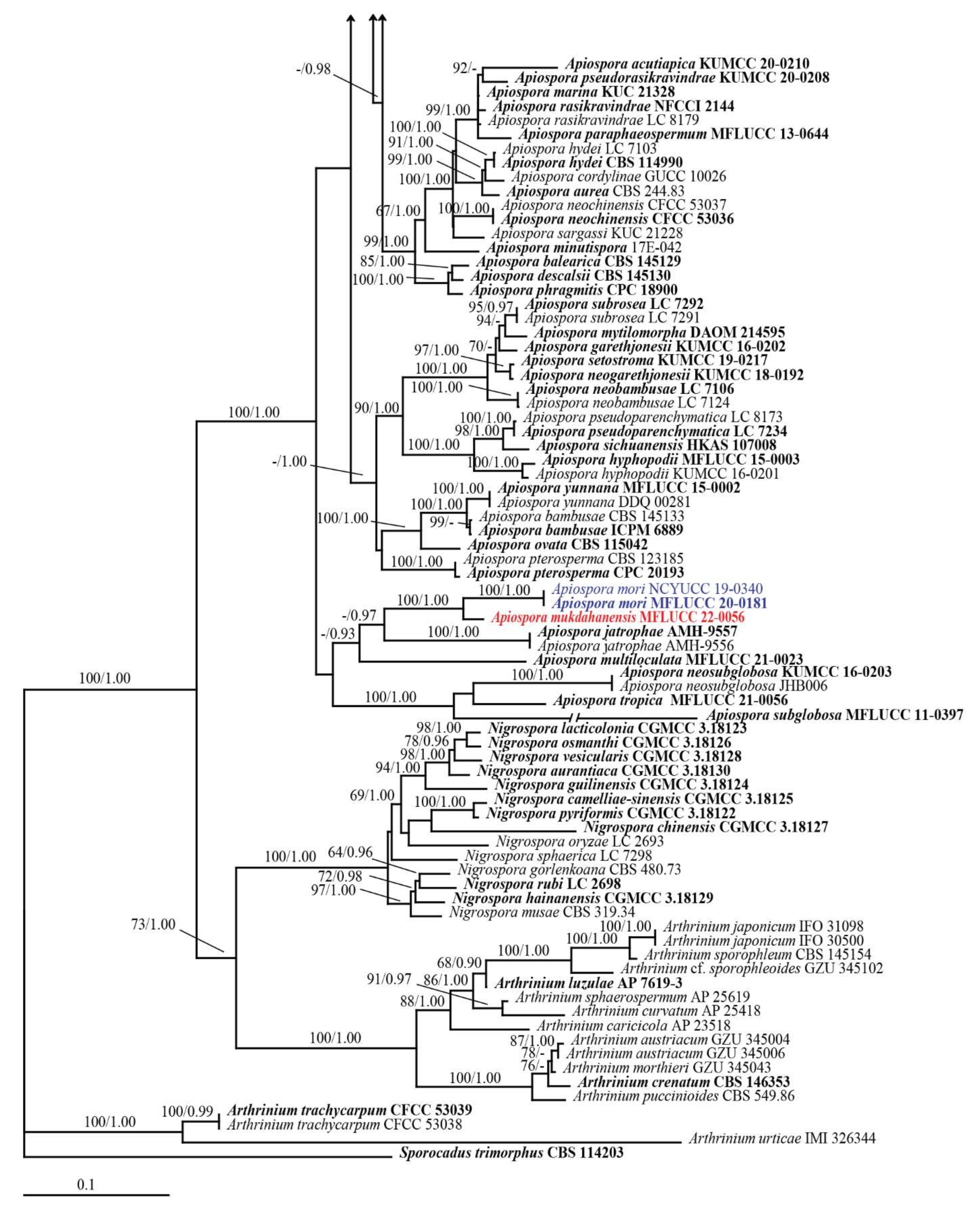
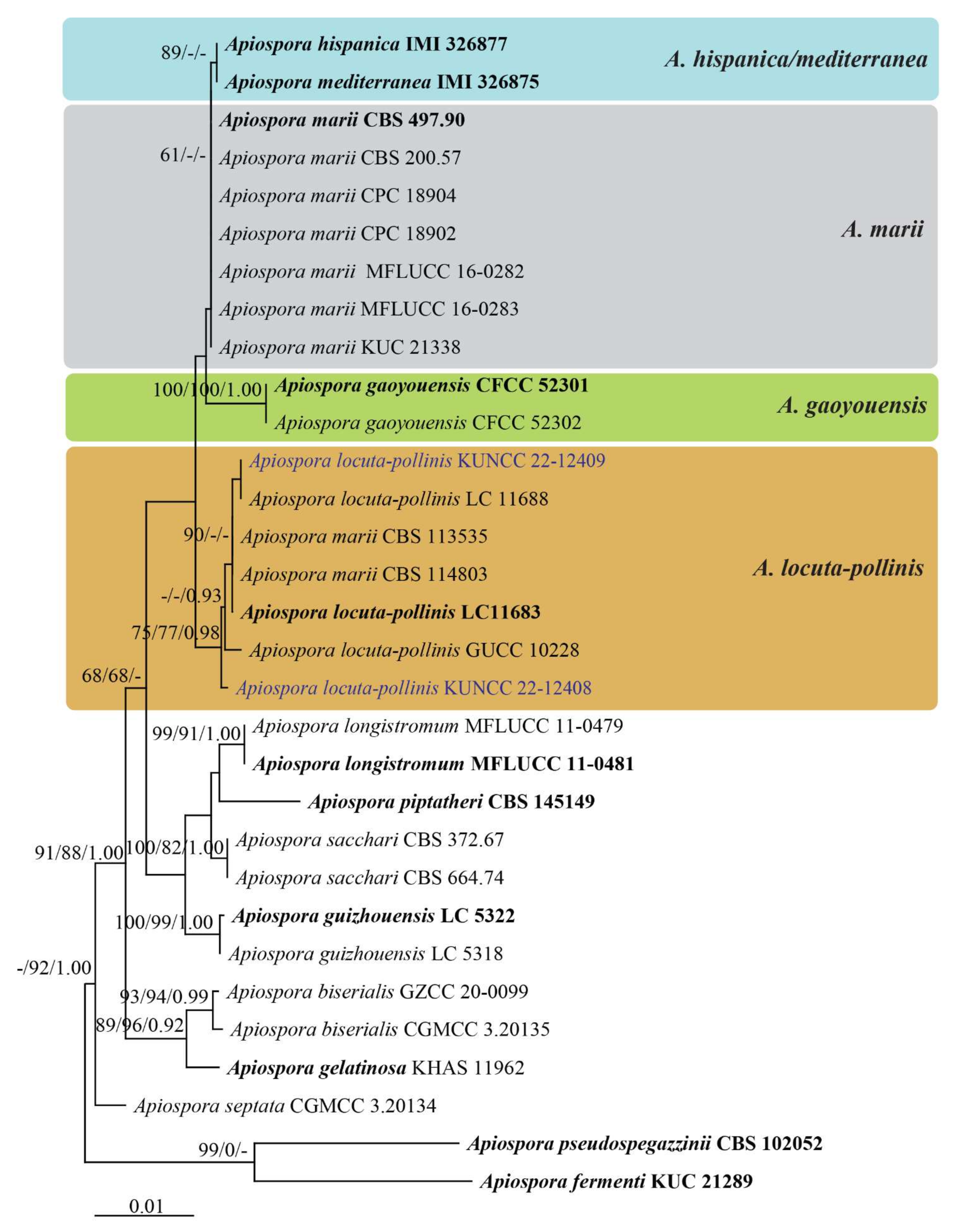
3.1. Phylogenetic Analyses
3.2. Taxonomy
3.2.1. Apiospora mori (Tennakoon, C.H. Kuo and K.D. Hyde) Monkai and Phookamsak, comb. nov.
3.2.2. Apiospora mukdahanensis Monkai and Phookamsak, sp. nov.
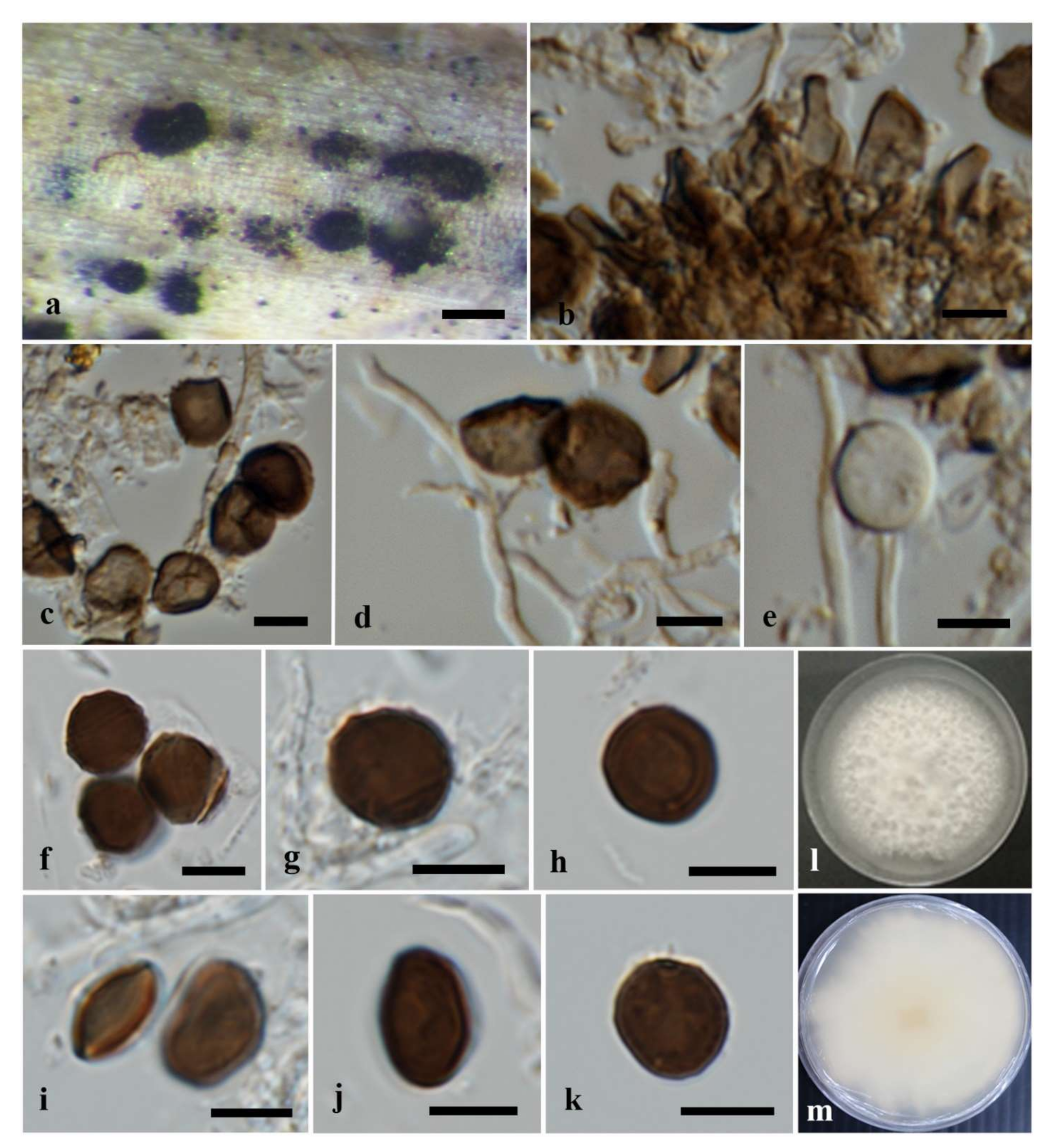
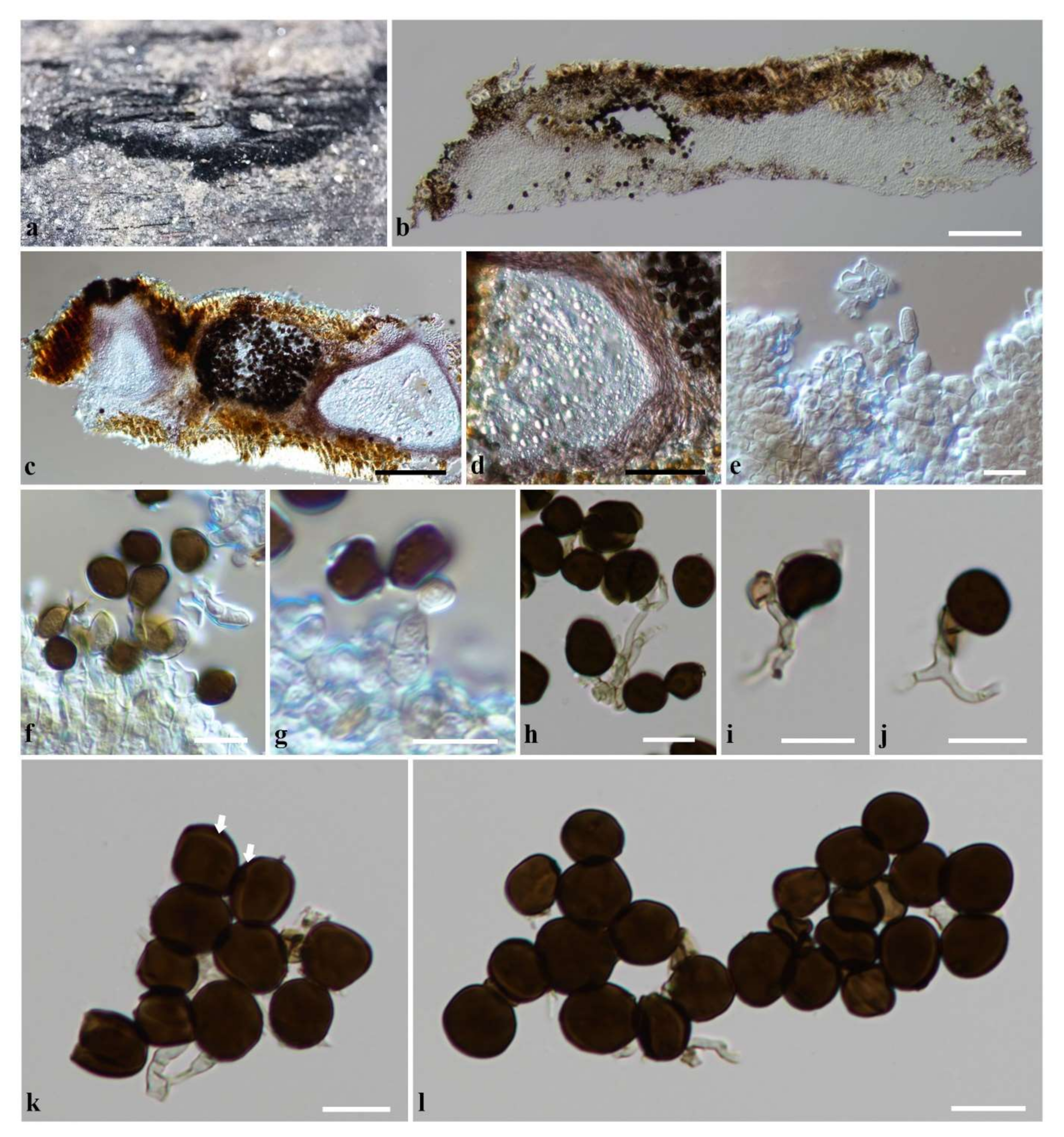
3.2.3. Apiospora locuta-pollinis (F. Liu and L. Cai) Pintos and P. Alvarado, Fungal Systematics and Evolution 7: 206 (2021)
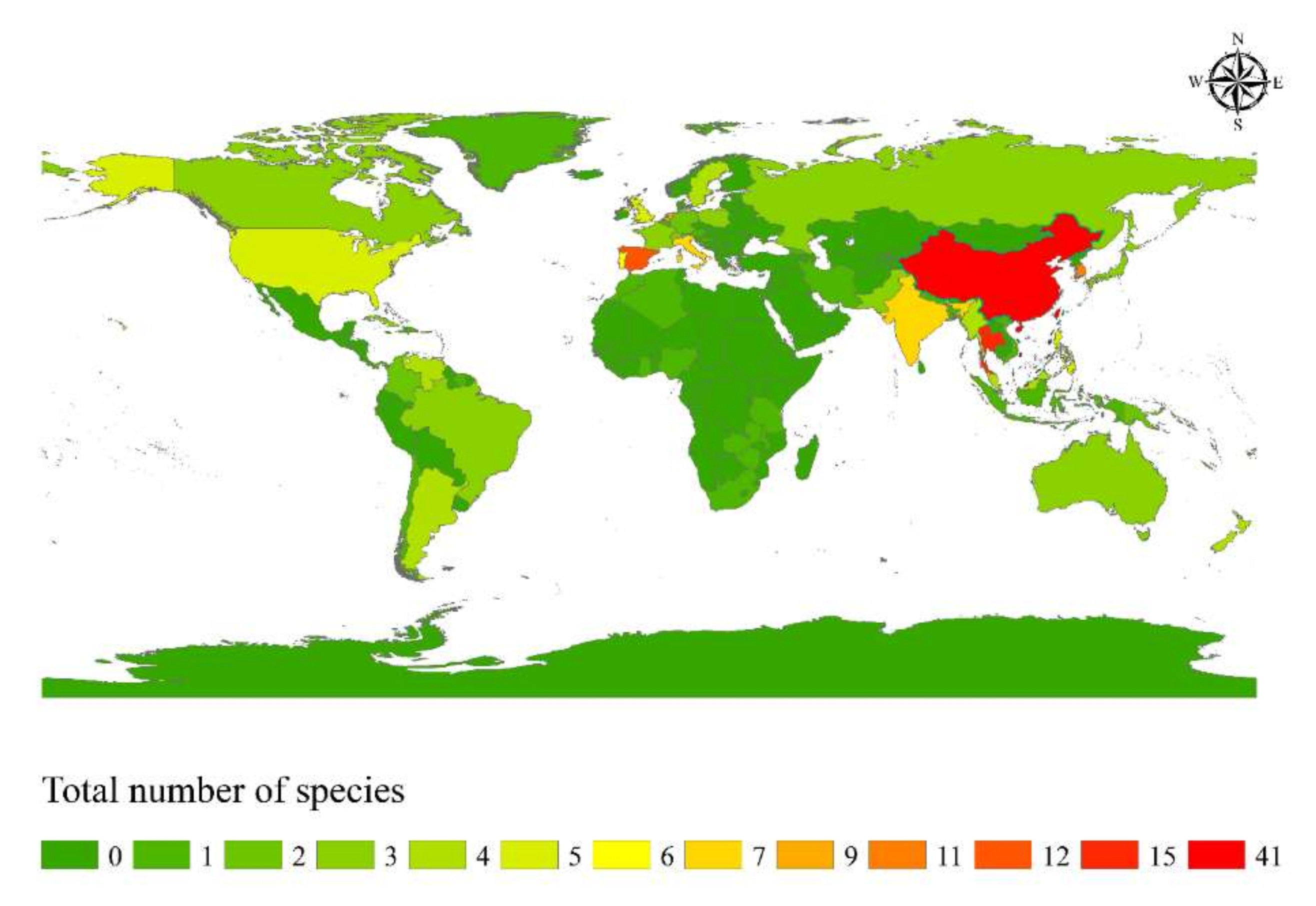
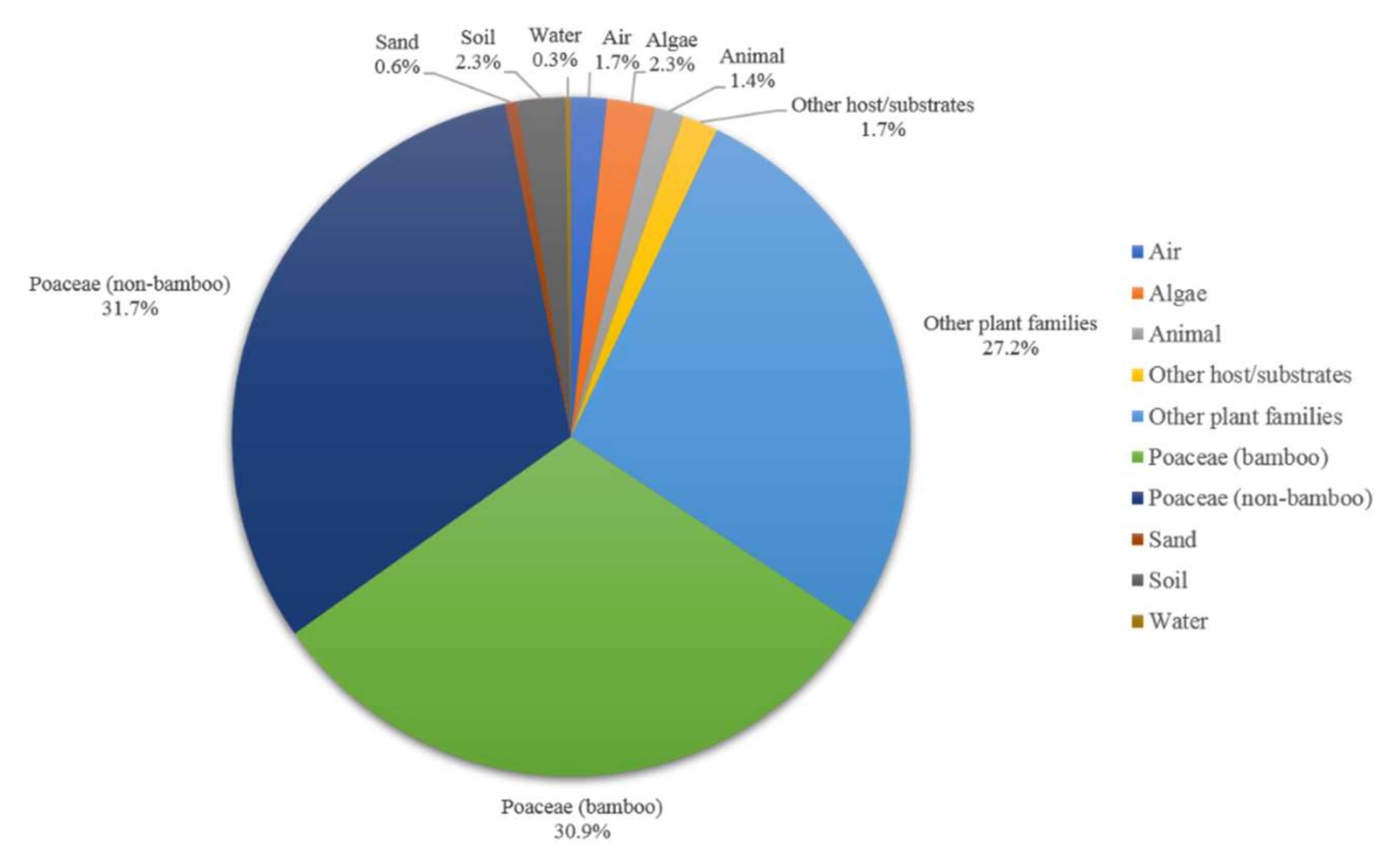
3.3. Host and Geographical Distribution of Apiospora Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Bao, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Pintos, Á.; Alvarado, P. Phylogenetic delimitation of Apiospora and Arthrinium. Fungal Syst. Evol. 2021, 7, 197–221. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Groenewald, J.Z. A phylogenetic re-evaluation of Arthrinium. IMA Fungus 2013, 4, 133–154. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Tan, X.M.; Liu, F.; Cai, L. Eight new Arthrinium species from China. MycoKeys 2018, 1, 1–24. [Google Scholar] [CrossRef]
- Pintos, Á.; Alvarado, P.; Planas, J.; Jarling, R. Six new species of Arthrinium from Europe and notes about A. caricicola and other species found in Carex spp. hosts. MycoKeys 2019, 49, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, R.; Kulkarni, G.; Sonawane, M.S.; Shouche, Y.S. A new endophytic species of Arthrinium (Apiosporaceae) from Jatropha podagrica. Mycoscience 2014, 55, 118–123. [Google Scholar] [CrossRef]
- Dai, D.Q.; Jiang, H.B.; Tang, L.Z.; Bhat, D.J. Two new species of Arthrinium (Apiosporaceae, Xylariales) associated with bamboo from Yunnan, China. Mycosphere 2016, 7, 1332–1345. [Google Scholar] [CrossRef]
- Dai, D.Q.; Phookamsak, R.; Wijayawardene, N.N.; Li, W.J.; Bhat, D.J.; Xu, J.C.; Taylor, J.E.; Hyde, K.D.; Chukeatirote, E. Bambusicolous fungi. Fungal Divers. 2017, 82, 1–105. [Google Scholar] [CrossRef]
- Jiang, N.; Li, J.; Tian, C.M. Arthrinium species associated with bamboo and reed plants in China. Fungal Syst. Evol. 2018, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Senanayake, I.C.; Bhat, J.D.; Cheewangkoon, R.; Xie, N. Bambusicolous Arthrinium Species in Guangdong Province, China. Front. Microbiol. 2020, 11, 602773. [Google Scholar] [CrossRef]
- Feng, Y.; Liu, J.K.J.; Lin, C.G.; Chen, Y.Y.; Xiang, M.M.; Liu, Z.Y. Additions to the genus Arthrinium (Apiosporaceae) from bamboos in China. Front. Microbiol. 2021, 12, 661281. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.G.; Karunarathna, S.C.; Mapook, A.; Promputtha, I.; Xu, J.C.; Bao, D.F.; Tibpromma, S. One new species and two new host records of Apiospora from bamboo and maize in Northern Thailand with thirteen new combinations. Life 2021, 11, 1071. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Luo, F.; Zhang, H.; Fang, X.; Zhu, T.; Li, S. First report of Arthrinium kogelbergense causing blight disease of Bambusa intermedia in Sichuan Province, China. Plant Dis. 2021, 105, 214. [Google Scholar] [CrossRef] [PubMed]
- Mavragani, D.C.; Abdellatif, L.; McConkey, B.; Hamel, C.; Vujanovic, V. First report of damping-off of durum wheat caused by Arthrinium sacchari in the semi-arid Saskatchewan fields. Plant Dis. 2007, 91, 469. [Google Scholar] [CrossRef]
- Aiello, D.; Gulisano, S.; Gusella, G.; Polizzi, G.; Guarnaccia, V. First report of fruit blight caused by Arthrinium xenocordella on Pistacia vera in Italy. Plant Dis. 2018, 102, 1853. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Liu, F.; Zhou, X.; Liu, X.Z.; Liu, S.J.; Cai, L. Culturable mycobiota from Karst caves in China, with descriptions of 20 new species. Persoonia 2017, 39, 1–31. [Google Scholar] [CrossRef]
- Singh, S.M.; Yadav, L.S.; Singh, P.N.; Hepat, R.; Sharma, R.; Singh, S.K. Arthrinium rasikravindrii sp. nov. from Svalbard, Norway. Mycotaxon 2013, 122, 449–460. [Google Scholar] [CrossRef]
- Das, K.; Lee, S.Y.; Choi, H.W.; Eom, A.H.; Cho, Y.J.; Jung, H.Y. Taxonomy of Arthrinium minutisporum sp. nov., Pezicula neosporulosa, and Acrocalymma pterocarpi: New records from soil in Korea. Mycobiology 2020, 48, 450–463. [Google Scholar] [CrossRef]
- Luo, Z.L.; Hyde, K.D.; Liu, J.K.; Maharachchikumbura, S.S.N.; Jeewon, R.; Bao, D.F.; Bhat, D.J.; Lin, C.G.; Li, W.L.; Yang, J.; et al. Freshwater Sordariomycetes. Fungal Divers. 2019, 99, 451–660. [Google Scholar] [CrossRef] [Green Version]
- Suryanarayanan, T.S. Fungal endosymbionts of seaweeds. In Biology of Marine Fungi; Raghukumar, C., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 53–70. [Google Scholar]
- Hong, J.-H.; Jang, S.; Heo, Y.M.; Min, M.; Lee, H.; Lee, Y.; Lee, H.; Kim, J.J. Investigation of marine-derived fungal diversity and their exploitable biological activities. Mar. Drugs 2015, 13, 4137–4155. [Google Scholar] [CrossRef]
- Heo, Y.M.; Kim, K.; Ryu, S.M.; Kwon, S.L.; Park, M.Y.; Kang, J.E.; Hong, J.H.; Lim, Y.W.; Kim, C.; Kim, B.S.; et al. Diversity and ecology of marine Algicolous Arthrinium species as a source of bioactive natural products. Mar. Drugs 2018, 16, 508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, M.S.; Oh, S.-Y.; Lee, S.; Eimes, J.A.; Lim, Y.W. Fungal diversity and enzyme activity associated with sailfin sandfish egg masses in Korea. Fungal Ecol. 2018, 34, 1–9. [Google Scholar] [CrossRef]
- Kwon, S.L.; Park, M.S.; Jang, S.; Lee, Y.M.; Heo, Y.M.; Hong, J.H.; Lee, H.; Jang, Y.; Park, J.H.; Kim, C.; et al. The genus Arthrinium (Ascomycota, Sordariomycetes, Apiosporaceae) from marine habitats from Korea, with eight new species. IMA Fungus 2021, 12, 13. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.L.; Cho, M.; Lee, Y.M.; Kim, C.; Lee, S.M.; Ahn, B.J.; Lee, H.; Kim, J.J. Two unrecorded Apiospora species isolated from marine substrates in Korea with eight new combinations (A. piptatheri and A. rasikravindrae). Mycobiology 2022, 50, 46–54. [Google Scholar] [CrossRef]
- He, Y.; Zhang, Z. Diversity of organism in the Usnea longissima lichen. Afr. J. Microbiol. Res. 2012, 6, 4797–4804. [Google Scholar]
- Crous, P.W.; Wingfield, M.J.; Le Roux, J.J.; Richardson, D.M.; Strasberg, D.; Shivas, R.G.; Alvarado, P.; Edwards, J.; Moreno, G.; Sharma, R. Fungal Planet description sheets: 371–399. Pers. Mol. Phylogeny Evol. Fungi 2015, 35, 264. [Google Scholar] [CrossRef]
- Rai, M.K. Mycosis in man due to Arthrinium phaeospermum var. indicum. First case report. Mycoses 1989, 32, 472–475. [Google Scholar]
- Zhao, Y.M.; Deng, C.R.; Chen, X. Arthrinium phaeospermum causing dermatomycosis, a new record of China. Acta Mycol. Sin. 1990, 9, 232–235. [Google Scholar]
- De Hoog, G.S.; Guarro, J.; Gené, J.; Figueras, M.J. Atlas of Clinical Fungi, 2nd ed.; CBS: Utrecht, The Netherlands, 2000; 1126p. [Google Scholar]
- Shrestha, P.; Ibáñez, A.B.; Bauer, S.; Glassman, S.I.; Szaro, T.M.; Bruns, T.D.; Taylor, J.W. Fungi isolated from Miscanthus and sugarcane: Biomass conversion, fungal enzymes, and hydrolysis of plant cell wall polymers. Biotechnol. Biofuels 2015, 8, 1. [Google Scholar] [CrossRef] [Green Version]
- Kunze, G. Zehn neue Pilzgattungen. Mykol. Hefte 1817, 1, 1–18. [Google Scholar]
- Ellis, M.B. Dematiaceous Hyphomycetes: IV. Mycol. Pap. 1963, 29, 1–33. [Google Scholar]
- Seifert, K.; Morgan-Jones, G.; Gams, W.; Kendrick, B. The Genera of Hyphomycetes. [CBS Biodiversity Series 9]; CBSKNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2011; pp. 1–1997. [Google Scholar]
- McNeill, J.; Barrie, F.R.; Buck, W.R.; Demoulin, V.; Greuter, W.; Hawksworths, D.L.; Herendeen, P.S.; Knapp, S.; Marhold, K.; Prado, J.; et al. International code of nomenclature for algae, fungi and plants (Melbourne Code) adopted by the Eighteenth International Botanical Congress Melbourne, Australia, July 2011. Regnum Veg. 2012, 154, 1–140. [Google Scholar]
- Senanayake, I.C.; Maharachchikumbura, S.S.N.; Hyde, K.D.; Bhat, J.D.; Jones, E.B.G.; McKenzie, E.H.C.; Dai, D.Q.; Daranagama, D.A.; Dayarathne, M.C.; Goonasekara, I.D.; et al. Towards unraveling relationships in Xylariomycetidae (Sordariomycetes). Fungal Divers. 2015, 73, 73–144. [Google Scholar] [CrossRef]
- Réblová, M.; Miller, A.N.; Rossman, A.Y.; Seifert, K.; Crous, P.; Hawksworth, D.; Adel-Wahab, M.A.; Cannon, P.F.; Daranagama, D.A.; De Beer, Z.W.; et al. Recommendations for competing sexual-asexually typified generic names in Sordariomycetes (except Diaporthales, Hypocreales, and Magnaporthales). IMA Fungus 2016, 7, 131–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.B.; Hyde, K.D.; Doilom, M.; Karunarathna, S.C.; Xu, J.C.; Phookamsak, R. Arthrinium setostromum (Apiosporaceae, Xylariales), a novel species associated with dead bamboo from Yunnan, China. Asian J. Mycol. 2019, 2, 254–268. [Google Scholar] [CrossRef]
- Phukhamsakda, C.; Nilsson, R.H.; Bhunjun, C.S.; de Farias, A.R.G.; Sun, Y.R.; Wijesinghe, S.N.; Raza, M.; Bao, D.-F.; Lu, L.; Tibpromma, S.; et al. The numbers of fungi: Contributions from traditional taxonomic studies and challenges of metabarcoding. Fungal Divers. 2022, 114, 327–386. [Google Scholar] [CrossRef]
- Species Fungorum. Available online: http://www.speciesfungorum.org (accessed on 1 September 2022).
- Senanayake, I.; Rathnayaka, A.; Marasinghe, D.; Calabon, M.; Gentekaki, E.; Lee, H.; Hurdeal, V.; Pem, D.; Dissanayake, L.; Wijesinghe, S.; et al. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Index Fungorum. Available online: http://www.indexfungorum.org (accessed on 1 September 2022).
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, J.; Buyck, B.; Cai, L.; Dai, Y.C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I.; et al. The Faces of Fungi database: Fungal names linked with morphology, phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: Cambridge, MA, USA, 1990; Volume 18, p. 7. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Bhunjun, C.S.; Niskanen, T.; Suwannarach, N.; Wannathes, N.; Chen, Y.J.; McKenzie, E.H.; Maharachchikumbura, S.S.N.; Buyck, B.; Zhao, C.-L.; Fan, Y.-G.; et al. The numbers of fungi: Are the most speciose genera truly diverse? Fungal Divers. 2022, 114, 387–462. [Google Scholar] [CrossRef]
- Samarakoon, M.C.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Stadler, M.; Jones, E.B.G.; Promputtha, I.; Suwannarach, N.; Camporesi, E.; Bulgakov, T.S.; Liu, J.K. Taxonomy, phylogeny, molecular dating and ancestral state reconstruction of Xylariomycetidae (Sordariomycetes). Fungal Divers. 2022, 112, 1–88. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Proceedings of the Nucleic Acids Symposium Series, London, UK, 2–6 September 1999; pp. 95–98. [Google Scholar]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES science gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; IEEE: New Orleans, LA, USA, 2010; pp. 1–8. [Google Scholar]
- Nylander, J.A. MrModeltest 2. Program Distributed by the Author; Department of Systematic Zoology, Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhaxybayeva, O.; Gogarten, J.P. Bootstrap, Bayesian probability and maximum likelihood mapping: Exploring new tools for comparative genome analyses. BMC Genom. 2002, 3, 4. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Hoehna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Swofford, D.L. PAUP* Phylogenetic Analysis Using Parsimony * (and Other Methods); Version 4.0.; Sinauer Associates: Sunderland, UK, 2002. [Google Scholar]
- Hillis, D.M.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J. FigTree: Tree Figure Drawing Tool; Institute of Evolutionary Biology, University of Edinburgh: Edinburgh, Scotland, 2012. [Google Scholar]
- Tennakoon, D.S.; Kuo, C.H.; Maharachchikumbura, S.S.N.; Thambugala, K.M.; Gentekaki, E.; Phillips, A.J.L.; Bhat, D.J.; Wanasinghe, D.N.; de Silva, N.I.; Promputtha, I.; et al. Taxonomic and phylogenetic contributions to Celtis formosana, Ficus ampelas, F. septica, Macaranga tanarius and Morus australis leaf litter inhabiting microfungi. Fungal Divers. 2021, 108, 1–215. [Google Scholar] [CrossRef]
- Zhao, Y.Z.; Zhang, Z.F.; Cai, L.; Peng, W.J.; Liu, F. Four new filamentous fungal species from newly-collected and hive-stored bee pollen. Mycosphere 2018, 9, 1089–1116. [Google Scholar] [CrossRef]
- Gerin, D.; Nigro, F.; Faretra, F.; Pollastro, S. Identification of Arthrinium marii as causal agent of olive tree dieback in Apulia (southern Italy). Plant Dis. 2020, 104, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Lobovikov, M.; Paudel, S.; Ball, L.; Piazza, M.; Guardia, M.; Ren, H.; Russo, L.; Wu, J.Q. World Bamboo Resources: A Thematic Study Prepared in the Framework of the Global Forest Resources Assessment 2005; Food and Agriculture Organization of The United Nations: Rome, Italy, 2007; Volume 18. [Google Scholar]
- Farr, D.F.; Rossman, A.Y. Fungal Databases, U.S. National Fungus Collections, ARS, USDA. Available online: https://nt.ars-grin.gov/fungaldatabases/ (accessed on 1 September 2022).
- Chen, K.; Wu, X.Q.; Huang, M.X.; Han, Y.Y. First report of brown culm streak of Phyllostachys praecox caused by Arthrinium arundinis in Nanjing, China. Plant Dis. 2014, 98, 1274. [Google Scholar] [CrossRef]
- Bagherabadi, S.; Zafari, D.; Anvar, F.G. First report of leaf spot caused by Arthrinium arundinis on rosemary in Iran. J. Plant Pathol. 2014, 96, 4–126. [Google Scholar]
- Larrondo, J.V.; Calvo, M.A. Two new species of Arthrinium from Spain. Mycologia 1990, 82, 396–398. [Google Scholar] [CrossRef]
- Larrondo, J.V.; Calvo, M.A. New contributions to the study of the genus Arthrinium. Mycologia 1992, 84, 475–478. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Culture Accession No. | Genbank Accession No. | |||
|---|---|---|---|---|---|
| ITS | LSU | TUB2 | TEF1-α | ||
| Apiospora acutiapica | KUMCC 20-0210 | MT946343 | MT946339 | MT947366 | MT947360 |
| Apiospora agari | KUC 21333 | MH498520 | N/A | MH498478 | MH544663 |
| Apiospora aquatica | S-642 | MK828608 | MK835806 | N/A | N/A |
| Apiospora arctoscopi | KUC 21331 | MH498529 | N/A | MH498487 | MN868918 |
| Apiospora arundinis | CBS 133509 | KF144886 | KF144930 | KF144976 | KF145018 |
| Apiospora arundinis | CBS 449.92 | KF144887 | KF144931 | KF144977 | KF145019 |
| Apiospora aurea | CBS 244.83 | AB220251 | KF144935 | KF144981 | KF145023 |
| Apiospora balearica | CBS 145129 | MK014869 | MK014836 | MK017975 | MK017946 |
| Apiospora bambusae | CBS 145133 | MK014875 | MK014842 | MK017981 | MK017952 |
| Apiospora bambusae | ICMP 6889 | MK014874 | MK014841 | MK017980 | MK017951 |
| Apiospora bambusicola | MFLUCC 20-0144 | MW173030 | MW173087 | N/A | MW183262 |
| Apiospora biserialis | CGMCC 3.20135 * | MW481708 | MW478885 | MW522955 | MW522938 |
| Apiospora biserialis | GZCC 20_0099 * | MW481709 | MW478886 | MW522956 | MW522939 |
| Apiospora camelliae-sinensis | LC 5007 | KY494704 | KY494780 | KY705173 | KY705103 |
| Apiospora camelliae-sinensis | LC 8181 | KY494761 | KY494837 | KY705229 | KY705157 |
| Apiospora chiangraiense | MFLUCC 21-0053 | MZ542520 | MZ542524 | MZ546409 | N/A |
| Apiospora chromolaenae | MFLUCC 17-1505 | MT214342 | MT214436 | N/A | MT235802 |
| Apiospora cordylinae | GUCC 10026 | MT040105 | N/A | MT040147 | MT040126 |
| Apiospora cyclobalanopsidis | CGMCC 3.20136 | MW481713 | MW478892 | MW522962 | MW522945 |
| Apiospora descalsii | CBS 145130 | MK014870 | MK014837 | MK017976 | MK017947 |
| Apiospora dichotomanthi | LC 4950 | KY494697 | KY494773 | KY705167 | KY705096 |
| Apiospora dichotomanthi | LC 8175 | KY494755 | KY494831 | KY705223 | KY705151 |
| Apiospora esporlensis | CBS 145136 | MK014878 | MK014845 | MK017983 | MK017954 |
| Apiospora euphorbiae | IMI 285638b | AB220241 | AB220335 | AB220288 | N/A |
| Apiospora fermenti | KUC 21289 * | MF615226 | N/A | MF615231 | MH544667 |
| Apiospora gaoyouensis | CFCC 52301 * | MH197124 | N/A | MH236789 | MH236793 |
| Apiospora gaoyouensis | CFCC 52302 * | MH197125 | N/A | MH236790 | MH236794 |
| Apiospora garethjonesii | KUMCC 16-0202 | KY356086 | KY356091 | N/A | N/A |
| Apiospora gelatinosa | KHAS 11962 * | MW481706 | MW478888 | MW522958 | MW522941 |
| Apiospora guiyangensis | HKAS 102403 | MW240647 | MW240577 | MW775604 | MW759535 |
| Apiospora guizhouensis | LC 5318 * | KY494708 | KY494784 | KY705177 | KY705107 |
| Apiospora guizhouensis | LC 5322 * | KY494709 | KY494785 | KY705178 | KY705108 |
| Apiospora hispanica | IMI 326877 * | AB220242 | AB220336 | AB220289 | N/A |
| Apiospora hydei | CBS 114990 | KF144890 | KF144936 | KF144982 | KF145024 |
| Apiospora hydei | LC 7103 | KY494715 | KY494791 | KY705183 | KY705114 |
| Apiospora hyphopodii | MFLUCC 15-0003 | KR069110 | N/A | N/A | N/A |
| Apiospora hyphopodii | KUMCC 16-0201 | KY356088 | KY356093 | N/A | N/A |
| Apiospora ibericum | CBS 145137 | MK014879 | MK014846 | MK017984 | MK017955 |
| Apiospora intestini | CBS 135835 | KR011352 | KR149063 | KR011350 | KR011351 |
| Apiospora intestini | MFLUCC 21-0052 | MZ542521 | MZ542525 | MZ546410 | MZ546406 |
| Apiospora italica | CBS 145138 | MK014880 | MK014847 | MK017985 | MK017956 |
| Apiospora italica | CBS 145139 | MK014881 | MK014848 | MK017986 | MK017957 |
| Apiospora jatrophae | AMH-9556 | HE981191 | N/A | N/A | N/A |
| Apiospora jatrophae | AMH-9557 | JQ246355 | N/A | N/A | N/A |
| Apiospora jiangxiense | LC 4494 | KY494690 | KY494766 | KY705160 | KY705089 |
| Apiospora jiangxiense | LC 4577 | KY494693 | KY494769 | KY705163 | KY705092 |
| Apiospora kogelbergense | CBS 113332 | KF144891 | KF144937 | KF144983 | KF145025 |
| Apiospora kogelbergense | CBS 113333 | KF144892 | KF144938 | KF144984 | KF145026 |
| Apiospora koreana | KUC 21332 | MH498524 | N/A | MH498482 | MH544664 |
| Apiospora locuta-pollinis | GUCC 10228 * | MT040124 | N/A | MT040166 | MT040145 |
| Apiospora locuta-pollinis | KUNCC 22-12408 * | OP377736 | OP377743 | N/A | OP381090 |
| Apiospora locuta-pollinis | KUNCC 22-12409 * | OP377737 | OP377744 | N/A | OP381091 |
| Apiospora locuta-pollinis | LC 11683 * | MF939595 | N/A | MF939622 | MF939616 |
| Apiospora locuta-pollinis | LC 11688 * | MF939596 | N/A | MF939623 | MF939618 |
| Apiospora longistromum | MFLUCC 11-0479 * | KU940142 | KU863130 | N/A | N/A |
| Apiospora longistromum | MFLUCC 11-0481 * | KU940141 | KU863129 | N/A | N/A |
| Apiospora malaysiana | CBS 102053 | KF144896 | KF144942 | KF144988 | KF145030 |
| Apiospora marii | CBS 113535 * | KF144898 | KF144944 | KF144990 | KF145032 |
| Apiospora marii | CBS 114803 * | KF144899 | KF144945 | KF144991 | KF145033 |
| Apiospora marii | CPC 18902 * | KF144901 | KF144948 | ||
| Apiospora marii | CPC 18904 * | KF144902 | KF144949 | KF144994 | KF145036 |
| Apiospora marii | CBS 200.57 * | KF144900 | KF144946 | KF144992 | KF145034 |
| Apiospora marii | CBS 497.90 * | AB220252 | KF144947 | KF144993 | KF145035 |
| Apiospora marii | KUC 21338 * | MH498549 | N/A | MH498507 | MH544681 |
| Apiospora marii | MFLUCC 16-0282 * | MH109526 | N/A | N/A | MH206166 |
| Apiospora marii | MFLUCC 16-0283 * | MH109527 | N/A | N/A | MH220419 |
| Apiospora marina | KUC 21328 | MH498538 | N/A | MH498496 | MH544669 |
| Apiospora mediterranea | IMI 326875 * | AB220243 | AB220337 | AB220290 | N/A |
| Apiospora minutispora | 17E-042 | LC517882 | N/A | LC518888 | LC518889 |
| Apiospora mori | NCYUCC 19-0340 | MW114314 | MW114394 | N/A | N/A |
| Apiospora mori | MFLUCC 20-0181 | MW114313 | MW114393 | N/A | N/A |
| Apiospora mukdahanensis | MFLUCC 22-0056 | OP377735 | OP377742 | N/A | OP381089 |
| Apiospora multiloculata | MFLUCC 21-0023 | OL873137 | OL873138 | OL874718 | N/A |
| Apiospora mytilomorpha | DAOM 214595 | KY494685 | N/A | N/A | N/A |
| Apiospora neobambusae | LC 7106 | KY494718 | KY494794 | KY705186 | KY806204 |
| Apiospora neobambusae | LC 7124 | KY494727 | KY494803 | KY705195 | KY806206 |
| Apiospora neochinensis | CFCC 53036 | MK819291 | N/A | MK818547 | MK818545 |
| Apiospora neochinensis | CFCC 53037 | MK819292 | N/A | MK818548 | MK818546 |
| Apiospora neogarethjonesii | KUMCC 18-0192 | MK070897 | MK070898 | N/A | N/A |
| Apiospora neosubglobosa | JHB006 | KY356089 | KY356094 | N/A | N/A |
| Apiospora neosubglobosa | KUMCC 16-0203 | KY356090 | KY356095 | N/A | N/A |
| Apiospora obovata | LC 4940 | KY494696 | KY494772 | KY705166 | KY705095 |
| Apiospora obovata | LC 8177 | KY494757 | KY494833 | KY705225 | KY705153 |
| Apiospora ovata | CBS 115042 | KF144903 | KF144950 | KF144995 | KF145037 |
| Apiospora paraphaeospermum | MFLUCC 13-0644 | KX822128 | KX822124 | N/A | N/A |
| Apiospora phragmitis | CPC 18900 | KF144909 | KF144956 | KF145001 | KF145043 |
| Apiospora phyllostachydis | MFLUCC 18-1101 | MK351842 | MH368077 | MK291949 | MK340918 |
| Apiospora piptatheri | CBS 145149 * | MK014893 | MK014860 | N/A | MK017969 |
| Apiospora pseudoparenchymatica | LC 7234 | KY494743 | KY494819 | KY705211 | KY705139 |
| Apiospora pseudoparenchymatica | LC 8173 | KY494753 | KY494829 | KY705221 | KY705149 |
| Apiospora pseudorasikravindrae | KUMCC 20-0208 | MT946344 | N/A | MT947367 | MT947361 |
| Apiospora pseudosinensis | CPC 21546 | KF144910 | KF144957 | N/A | KF145044 |
| Apiospora pseudospegazzinii | CBS 102052 * | KF144911 | KF144958 | KF145002 | KF145045 |
| Apiospora pterosperma | CBS 123185 | KF144912 | KF144959 | KF145003 | N/A |
| Apiospora pterosperma | CPC 20193 | KF144913 | KF144960 | KF145004 | KF145046 |
| Apiospora pusillisperma | KUC 21321 | MH498533 | N/A | MH498491 | MN868930 |
| Apiospora qinlingense | CFCC 52303 | MH197120 | N/A | MH236791 | MH236795 |
| Apiospora qinlingense | CFCC 52304 | MH197121 | N/A | MH236792 | MH236796 |
| Apiospora rasikravindrae | NFCCI 2144 | JF326454 | N/A | N/A | N/A |
| Apiospora rasikravindrae | LC 8179 | KY494759 | KY494835 | KY705227 | KY705155 |
| Apiospora sacchari | CBS 372.67 * | KF144918 | KF144964 | KF145007 | KF145049 |
| Apiospora sacchari | CBS 664.74 * | KF144919 | KF144965 | KF145008 | KF145050 |
| Apiospora saccharicola | CBS 191.73 | KF144920 | KF144966 | KF145009 | KF145051 |
| Apiospora saccharicola | CBS 831.71 | KF144922 | KF144969 | KF145012 | KF145054 |
| Apiospora sargassi | KUC 21228 | KT207746 | N/A | KT207644 | MH544677 |
| Apiospora septata | CGMCC 3.20134 * | MW481711 | MW478890 | MW522960 | MW522943 |
| Apiospora serenensis | IMI 326869 | AB220250 | AB220344 | AB220297 | N/A |
| Apiospora setariae | CFCC 54041 | MT492004 | N/A | MT497466 | MW118456 |
| Apiospora setostroma | KUMCC 19-217 | MN528012 | MN528011 | N/A | MN527357 |
| Apiospora sichuanensis | HKAS 107008 | MW240648 | MW240578 | MW775605 | MW759536 |
| Apiospora sorghi | URM 93000 | MK371706 | N/A | MK348526 | N/A |
| Apiospora subglobosa | MFLUCC 11-0397 | KR069112 | KR069113 | N/A | N/A |
| Apiospora subrosea | LC 7291 | KY494751 | KY494827 | KY705219 | KY705147 |
| Apiospora subrosea | LC 7292 | KY494752 | KY494828 | KY705220 | KY705148 |
| Apiospora taeanense | KUC 21322 | MH498515 | N/A | MH498473 | MH544662 |
| Apiospora thailandica | MFLUCC 15-1999 | KU940146 | KU863134 | N/A | N/A |
| Apiospora thailandica | MFLUCC 15-0202 | KU940145 | KU863133 | N/A | N/A |
| Apiospora tropica | MFLUCC 21–0056 | OK491657 | OK491653 | OK560922 | N/A |
| Apiospora vietnamense | IMI 99670 | KX986096 | KX986111 | KY019466 | N/A |
| Apiospora xenocordella | CBS 478.86 | KF144925 | KF144970 | KF145013 | KF145055 |
| Apiospora xenocordella | CBS 595.66 | KF144926 | KF144971 | N/A | N/A |
| Apiospora yunnana | MFLUCC 15-1002 | KU940147 | KU863135 | N/A | N/A |
| Apiospora yunnana | DDQ00281 | KU940148 | KU863136 | N/A | N/A |
| Arthrinium austriacum | GZU 345004 | MW208928 | N/A | N/A | N/A |
| Arthrinium austriacum | GZU 345006 | MW208929 | MW208860 | N/A | N/A |
| Arthrinium caricicola | CBS 145127 | MK014871 | MK014838 | MK017977 | MK017948 |
| Arthrinium cf. sporophleoides | GZU 345102 | MW208944 | MW208866 | N/A | N/A |
| Arthrinium crenatum | CBS 146353 | MW208931 | MW208861 | MW221923 | MW221917 |
| Arthrinium curvatum | AP 25418 | MK014872 | MK014839 | MK017978 | MK017949 |
| Arthrinium japonicum | IFO 30500 | AB220262 | AB220356 | AB220309 | N/A |
| Arthrinium japonicum | IFO 31098 | AB220264 | AB220358 | AB220311 | N/A |
| Arthrinium luzulae | AP 7619-3 | MW208937 | MW208863 | N/A | N/A |
| Arthrinium morthieri | GZU 345043 | MW208938 | MW208864 | N/A | N/A |
| Arthrinium phaeospermum | CBS 114317 | KF144906 | KF144953 | KF144998 | KF145040 |
| Arthrinium phaeospermum | CBS 114318 | KF144907 | KF144954 | KF144999 | KF145041 |
| Arthrinium puccinioides | CBS 549.86 | AB220253 | AB220347 | AB220300 | N/A |
| Arthrinium sphaerospermum | AP 25619 | MW208943 | MW208865 | N/A | N/A |
| Arthrinium sporophleum | CBS 145154 | MK014898 | MK014865 | MK018001 | MK017973 |
| Arthrinium trachycarpum | CFCC 53038 | MK301098 | N/A | MK303394 | MK303396 |
| Arthrinium trachycarpum | CFCC 53039 | MK301099 | N/A | MK303395 | MK303397 |
| Arthrinium urticae | IMI 326344 | AB220245 | AB220339 | AB220292 | N/A |
| Nigrospora aurantiaca | CGMCC 3.18130 | KX986064 | KX986098 | KY019465 | KY019295 |
| Nigrospora camelliae-sinensis | CGMCC 3.18125 | KX985986 | KX986103 | KY019460 | KY019293 |
| Nigrospora chinensis | CGMCC 3.18127 | KX986023 | KX986107 | KY019462 | KY019422 |
| Nigrospora gorlenkoana | CBS 480.73 | KX986048 | KX986109 | KY019456 | KY019420 |
| Nigrospora guilinensis | CGMCC 3.18124 | KX985983 | KX986113 | KY019459 | KY019292 |
| Nigrospora hainanensis | CGMCC 3.18129 | KX986091 | KX986112 | KY019464 | KY019415 |
| Nigrospora lacticolonia | CGMCC 3.18123 | KX985978 | KX986105 | KY019458 | KY019291 |
| Nigrospora musae | CBS 319.34 | MH855545 | KX986110 | KY019455 | KY019419 |
| Nigrospora oryzae | LC 2693 | KX985944 | KX986101 | KY019471 | KY019299 |
| Nigrospora osmanthi | CGMCC 3.18126 | KX986010 | KX986106 | KY019461 | KY019421 |
| Nigrospora pyriformis | CGMCC 3.18122 | KX985940 | KX986100 | KY019457 | KY019290 |
| Nigrospora rubi | LC 2698 | KX985948 | KX986102 | KY019475 | KY019302 |
| Nigrospora sphaerica | LC 7298 | KX985937 | KX986097 | KY019606 | KY019401 |
| Nigrospora vesicularis | CGMCC 3.18128 | KX986088 | KX986099 | KY019463 | KY019294 |
| Sporocadus trimorphus | CBS 114203 | MH553977 | MH554196 | MH554636 | MH554395 |
| Characteristics | Apiospora locuta-pollinis Strains | ||
|---|---|---|---|
| Type Strain (LC 11683) | KUN-HKAS 124566 | KUN-HKAS 124567 | |
| Host/substrate | On PDA, MEA isolated from hive-stored pollen | Decaying stem of bamboo | Dead stem of bamboo |
| Conidiomata | ND | 150–230 high × 450–830 µm long | 177–243 high × 446–682 µm long |
| Pycnidial wall | ND | (15–)20–40(–50) µm wide | (12–)17–40(–84) µm wide |
| Conidiophore mother cell | ND | (3.5–)4.5–9(–12) × 3.5–5.5 µm ( = 7.8 × 4.8 µm) | (4.5–)5.5–8 × (3.5–)5.5–8 µm ( = 5.7 × 5.5 µm) |
| Conidiophores | Reduced to conidiogenous cells | Pale brown, septate, branched, flexuous, (4–)5–10(–20) × (1.5–)2–3.5 µm ( = 8.9 × 2.2 µm) | Hyaline to light brown, septate, branched, flexuous, (8–)11–15(–24) × (1.5–)2.5–4.5 µm ( = 13.8 × 2.9 µm) |
| Conidiogenous cells | Pale brown, subglobose to ampulliform to doliiform, 3–7.5 × 3–6 μm ( = 4.9 × 3.8 μm) | Hyaline to light brown, cylindrical to subcylindrical or ampulliform, (3.5–)4.5–6.5(–9) × 2.5–4.5 µm ( = 6 × 3.5 µm) | Hyaline to light brown, subglobose to ampulliform or doliiform, (2–)3–6 × (1–)2–4.5 µm ( = 3.2 × 2.2 µm) |
| Conidia | Pale brown to brown with hyaline equatorial rim, globose to subglobose, 5.5–9 × 4.5–8 μm ( = 7.1 × 6.4 μm), or ellipsoidal, 8–15 × 5–9.5 μm ( = 10.7 × 7.1 μm) | Dark brown with a paler equatorial slit, globose to obovoid, or ellipsoidal, (8–)9–13 × (7–)9–12 µm ( = 10.5 × 10 µm) | Dark brown with a paler equatorial slit, globose to obovoid, or ellipsoidal, (8–)9–11 × (5.5–)8–11 µm ( = 9.6 × 8 µm) |
| Sterile cells | Pale brown or brown, ellipsoidal to clavate, 11.5–21 × 3.5–8 μm ( = 15.7 × 5.7 μm) | brown, leaf-like, attached to the conidiophore | ND |
| References | Zhao et al. [61] | This study | This study |
| Characters | Apiospora Species | ||||
|---|---|---|---|---|---|
| A. marii | A. mediterranea | A. hispanica | A. gaoyouensis | A. locuta-pollinis | |
| Host/substrate | On MEA | On MEA, isolated from Pharmaceutical excipient, atmosphere and grass | On MEA, isolated from Beach sand | On leaves and culms of Phragmites australis | On PDA, MEA isolated from hive-stored pollen |
| Conidiomata | NR | NR | NR | Scattered to gregarious, superficial on leaf and culms, 1–15 long × 0.5–5 mm wide | NR |
| Conidiophore mother cell | Ampulliform | Ampulliform | Globose to subglobose | NR | NR |
| Conidiophores | Basauxic, mononematous, macronematous, brownish with transverse septa of the same color | Basauxic, macronematous, mononematous, nonseptate, colorless | Basauxic, mononematous, macronematous, brownish, without transverse septa | Reduced to conidiogenous cells | Reduced to conidiogenous cells |
| Conidiogenous cells | NR | Integrated and terminal | NR | Aggregated in clusters on hyphae, smooth | Pale brown, smooth, subglobose to ampulliform to doliiform, 3–7.5 × 3–6 μm |
| Conidia | Lateral or terminal, dark brown with hyaline rim, 7.2–7.5 × 6.1–6.5 μm | Lateral or terminal, brown with hyaline rim, smooth, lenticular in shape, 9–9.5 × 7.5–9 μm | Lateral or terminal, dark brown with hyaline rim, 7.5–8.5 × 6.2–7.6 μm | Brown with pale equatorial slit, smooth, granular, globose to elongate ellipsoid in surface view, 5–8 μm diam., lenticular in side view, 4–8 μm diam., with central basal scar | Pale brown to brown with hyaline equatorial rim, smooth, globose to subglobose, 5.5–9 × 4.5–8 μm, or ellipsoidal, 8–15× 5–9.5 μm |
| Sterile cells | NR | Pale brown, 7–7.5 × 6.5–7 μm | Irregularly shaped, much smaller than conidia, occasionally globose | Brown, elongated cells seldom intermingled among conidia | Pale brown or brown, smooth, ellipsoidal to clavate, 11.5–21 × 3.5–8 μm |
| References | Larrondo and Calvo [67] | Larrondo and Calvo [68] | Larrondo and Calvo [68] | Jiang and Tian [9] | Zhao et al. [61] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monkai, J.; Phookamsak, R.; Tennakoon, D.S.; Bhat, D.J.; Xu, S.; Li, Q.; Xu, J.; Mortimer, P.E.; Kumla, J.; Lumyong, S. Insight into the Taxonomic Resolution of Apiospora: Introducing Novel Species and Records from Bamboo in China and Thailand. Diversity 2022, 14, 918. https://doi.org/10.3390/d14110918
Monkai J, Phookamsak R, Tennakoon DS, Bhat DJ, Xu S, Li Q, Xu J, Mortimer PE, Kumla J, Lumyong S. Insight into the Taxonomic Resolution of Apiospora: Introducing Novel Species and Records from Bamboo in China and Thailand. Diversity. 2022; 14(11):918. https://doi.org/10.3390/d14110918
Chicago/Turabian StyleMonkai, Jutamart, Rungtiwa Phookamsak, Danushka S. Tennakoon, Darbhe Jayarama Bhat, Sheng Xu, Qinxian Li, Jianchu Xu, Peter E. Mortimer, Jaturong Kumla, and Saisamorn Lumyong. 2022. "Insight into the Taxonomic Resolution of Apiospora: Introducing Novel Species and Records from Bamboo in China and Thailand" Diversity 14, no. 11: 918. https://doi.org/10.3390/d14110918
APA StyleMonkai, J., Phookamsak, R., Tennakoon, D. S., Bhat, D. J., Xu, S., Li, Q., Xu, J., Mortimer, P. E., Kumla, J., & Lumyong, S. (2022). Insight into the Taxonomic Resolution of Apiospora: Introducing Novel Species and Records from Bamboo in China and Thailand. Diversity, 14(11), 918. https://doi.org/10.3390/d14110918