

Efficacy of Killing Large Carnivores to Enhance Moose Harvests: New Insights from a Long-Term View

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
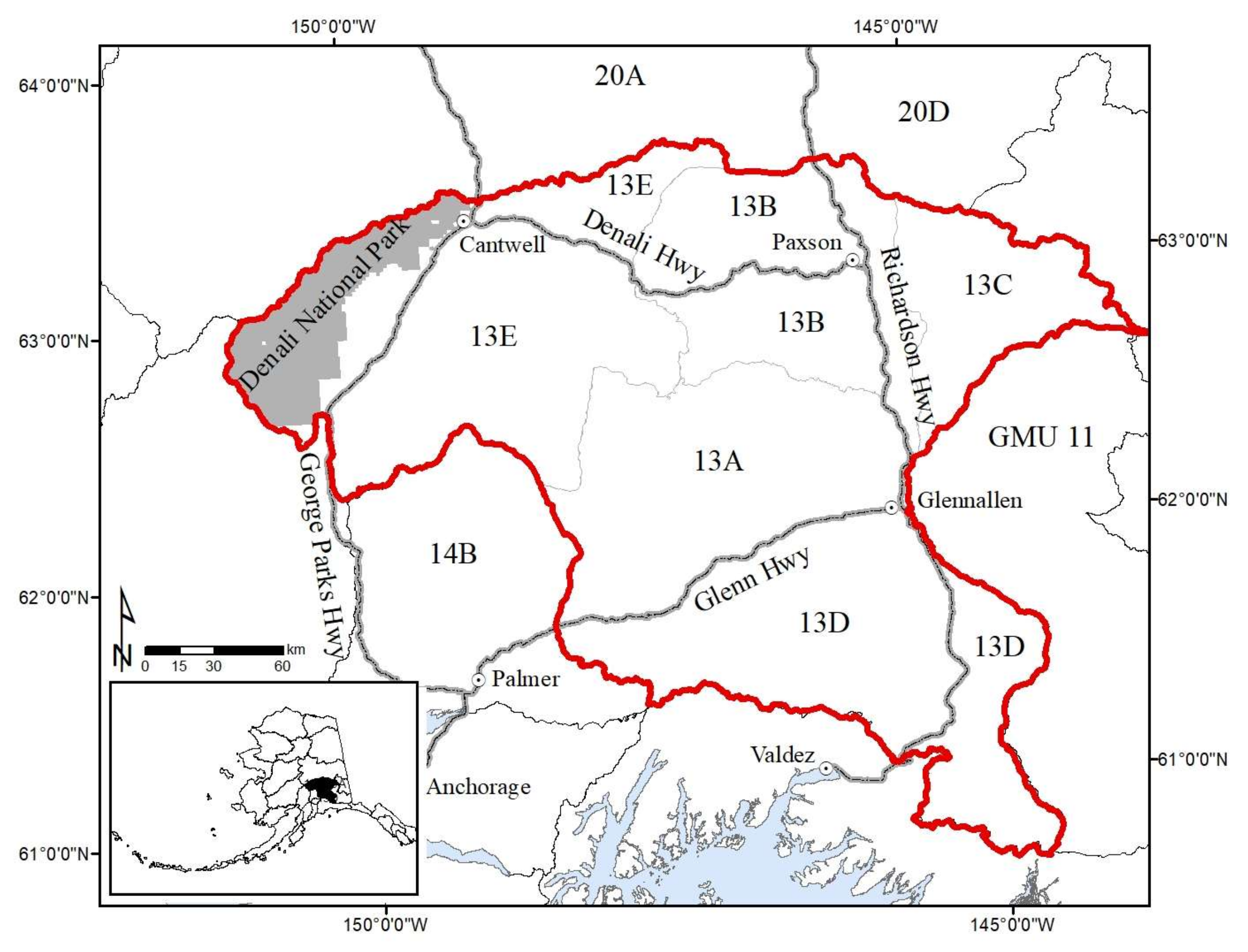
2. Study Area
3. Materials and Methods
3.1. Harvest Data
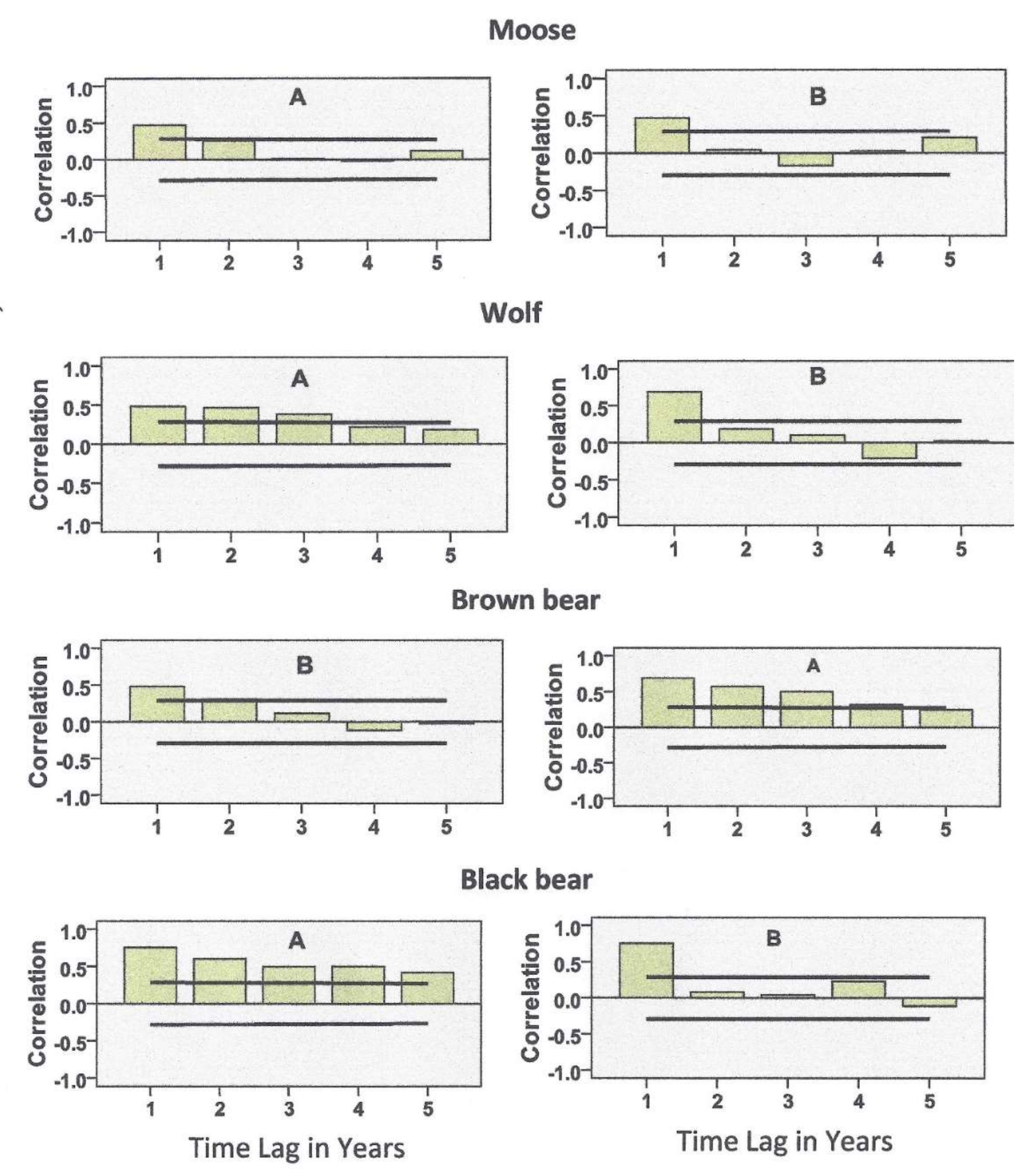
3.2. Statistical Analyses
4. Results
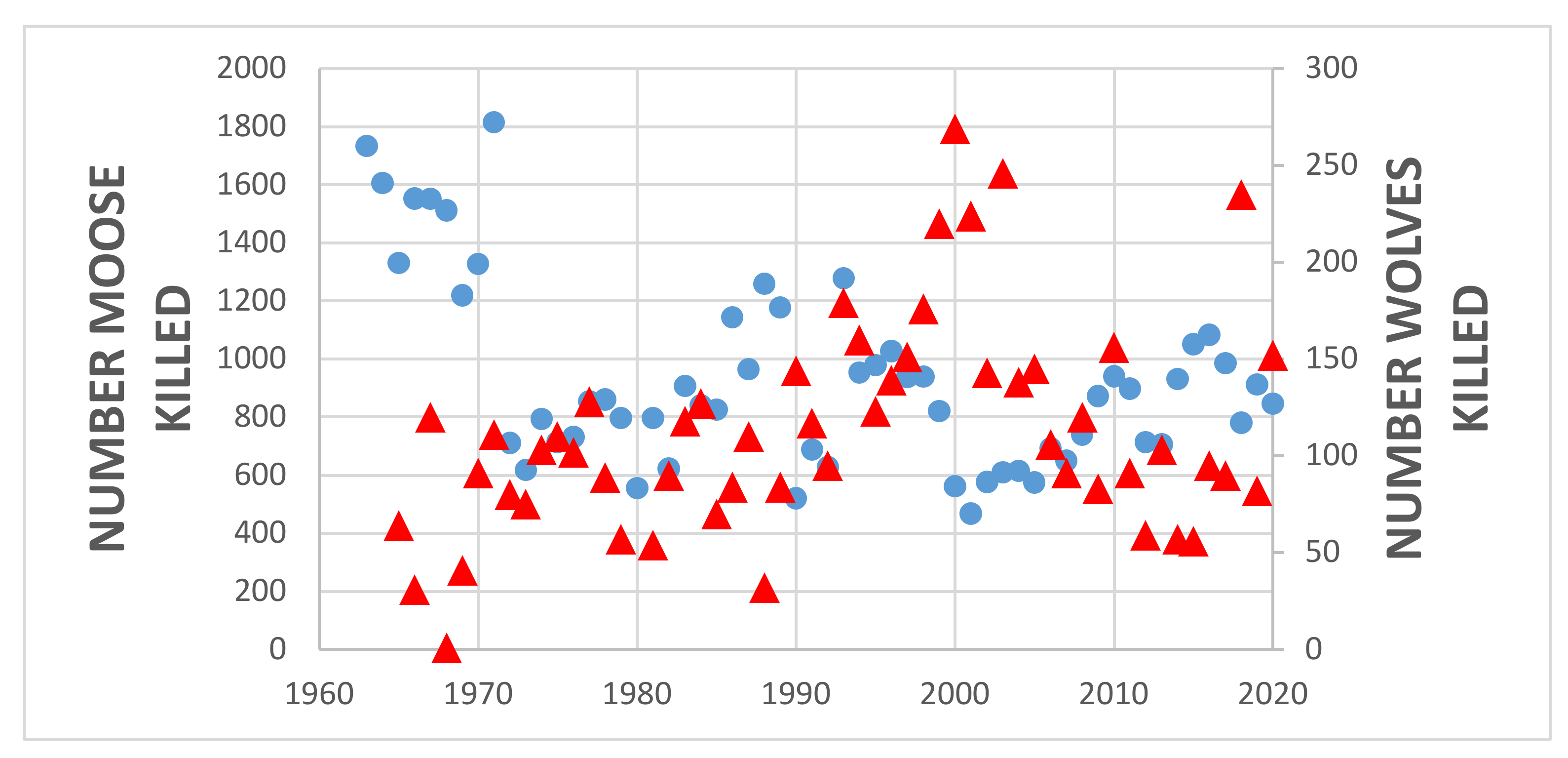
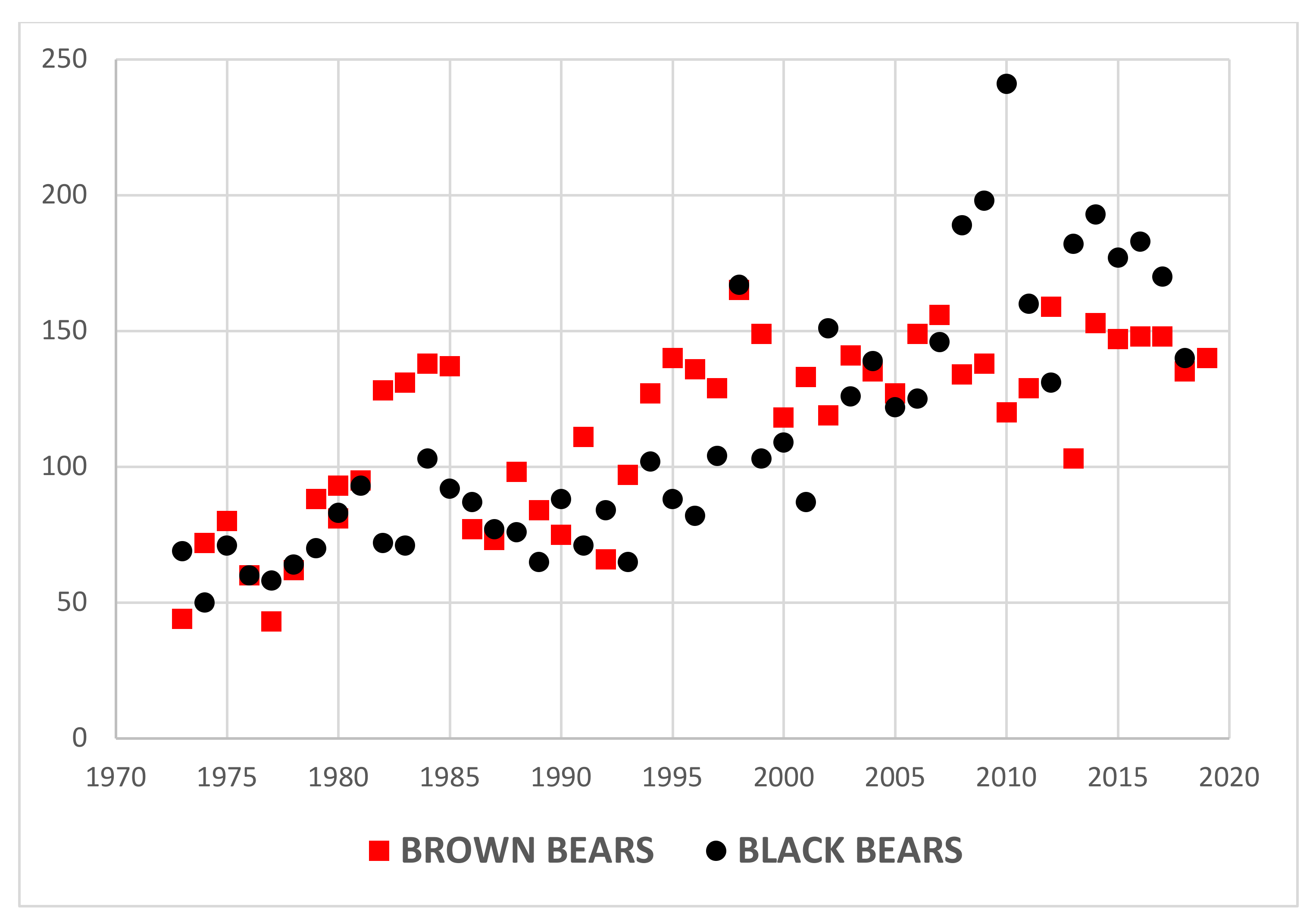
4.1. Game Management Unit 13
4.2. GMU 13D and Subunits A, B, C, and E
4.3. Moose Harvest Means Comparisons, Subunit A, B, C, and E
5. Discussion
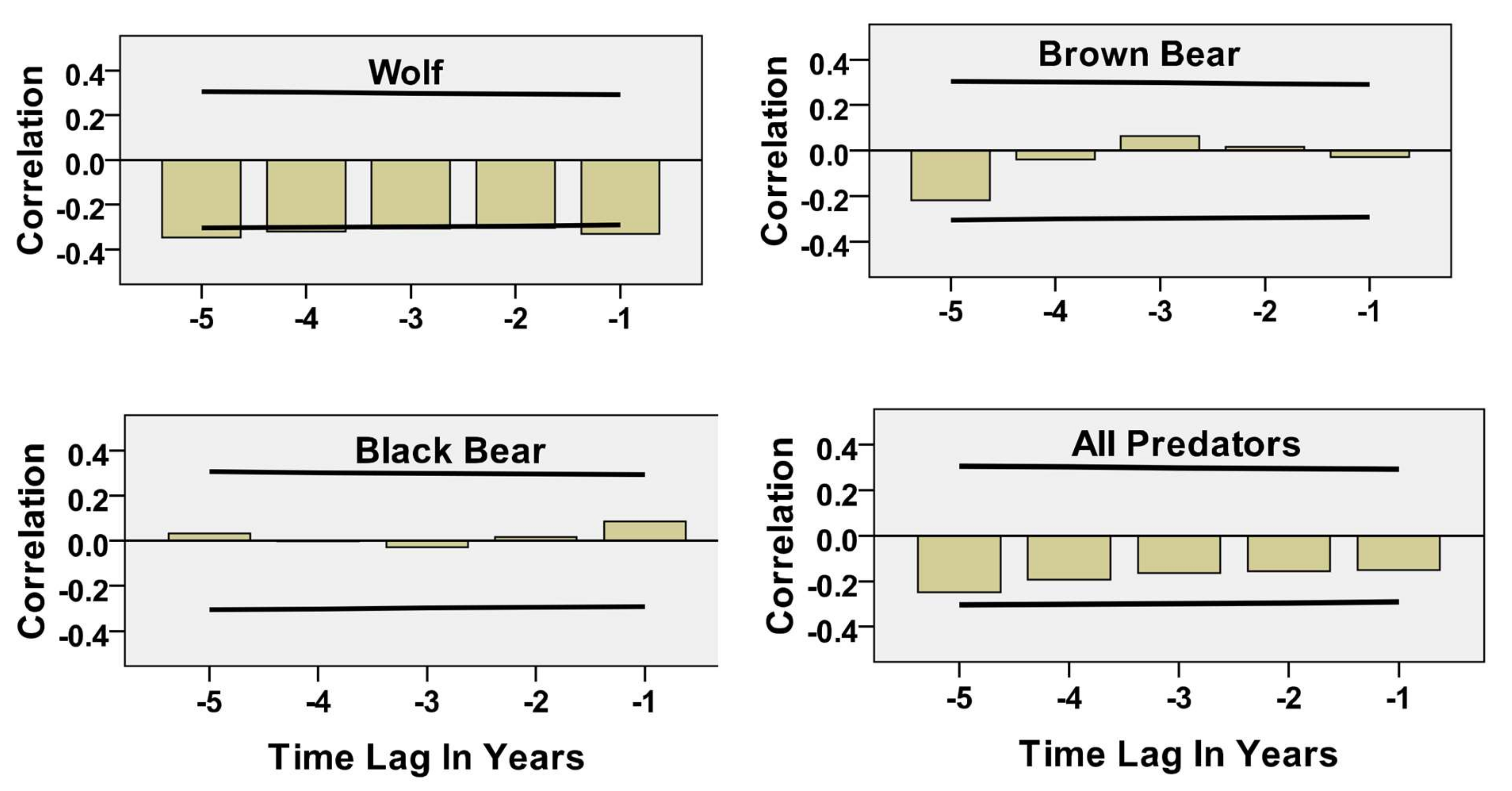
5.1. Predator Kill and Moose Harvest
5.2. Management Considerations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bowyer, R.T.; Person, D.E.; Pierce, E.M. Detecting Dop-Down versus Bottom-Up Regulation of Ungulates by Large Carnivores: Implications for Conservation of Biodiver-Sity. In Large Carnivores and the Conservation of Biodiversity; Ray, J.C., Redford, K.H., Steneck, R.S., Berger, J., Eds.; Island Press: Washington, DC, USA, 2005; pp. 342–361. [Google Scholar]
- Gasaway, W.C.; Boertje, R.D.; Grandgaard, D.V.; Kelleyhouse, D.G.; Stephenson, R.O.; Larsen, D.G. The role of predation in limiting moose at low densities in Alaska and Yukon and implications for conservation. Wildl. Monogr. 1992, 120, 3–59. [Google Scholar]
- Van Ballenberghe, V.; Ballard, W.B. Limitation and regulation of moose populations: The role of predation. Can. J. Zool. 1994, 72, 2071–2077. [Google Scholar] [CrossRef]
- Bowyer, R.T.; Kie, J.G.; Person, D.K.; Monteith, K.L. Metrics of predation: Perils of predator-prey ratios. Acta Theol. 2013, 58, 329–340. [Google Scholar] [CrossRef]
- Bowyer, R.T.; Stewart, K.M.; Bleich, V.C.; Whiting, J.C.; Monteith, K.L.; Blum, M.E.; LaSharr, T.N. Metrics of harvest for ungulate populations: Misconceptions, lurking variables, and prudent management. Alces 2020, 56, 15–38. [Google Scholar]
- Ballard, W.B.; Spraker, T.H.; Taylor, K.P. Causes of neonatal moose calf mortality in southcentral Alaska. J. Wildl. Manag. 1981, 45, 335–342. [Google Scholar] [CrossRef]
- Ballard, W.B.; Miller, S.D.; Whitman, J.S. Brown and black bear predation on moose in southcentral Alaska. Alces 1990, 26, 1–8. [Google Scholar]
- Ballard, W.B.; Whitman, J.S.; Reed, D.J. Dynamics of moose in south-central Alaska. Wildl. Monogr. 1991, 114, 1–49. [Google Scholar]
- Schwartz, C.C.; Franzmann, A.W. Interrelationship of black bears to moose and forest succession in the northern coniferous forest. Wildl. Monogr. 1991, 113, 3–58. [Google Scholar]
- Bowyer, R.T.; Van Ballenberghe, V.; Kie, J.G. Timing and synchrony of parturition in Alaskan moose: Long-term versus proximal effects of climate. J. Mammal. 1998, 79, 1332–1344. [Google Scholar] [CrossRef] [Green Version]
- Ballard, W.B.; Whitman, J.S.; Gardner, G.L. Ecology of an exploited wolf population in south-central Alaska. Wildl. Monogr. 1987, 98, 3–54. [Google Scholar]
- Keech, M.A.; Boertje, R.D.; Bowyer, R.T.; Dale, B.W. Effects of birth weight on growth of young moose: Do low-weight neonates compensate? Alces 1999, 35, 51–57. [Google Scholar]
- McCullough, D.R. Male harvest in relation to female removals in a black-tailed deer population. J. Wildl. Manag. 2001, 65, 46–58. [Google Scholar] [CrossRef]
- Bartmann, R.M.; White, G.C.; Carpenter, H.H. Compensatory mortality in a Colorado mule deer population. Wildl. Monogr. 1992, 121, 3–39. [Google Scholar]
- Gilbert, S.L.; Hundertmark, K.J.; Lindberg, M.S.; Person, D.K.; Boyce, M.S. The importance of environmental variability and transient population dynamics for a northern ungulate. Front. Ecol. Evol. 2020, 8, 1–14. [Google Scholar] [CrossRef]
- Testa, J.W. Population dynamics and life history trade-offs of moose (Alces alces) in south-central Alaska. Ecology 2004, 85, 1439–1452. [Google Scholar] [CrossRef]
- Hundermark, K.J.; Schwartz, C.C. Considerations for intensive management of moose in Alaska. Alces 1996, 32, 15–24. [Google Scholar]
- Boertje, R.D.; Keech, M.A.; Young, D.D.; Kellie, K.A.; Seaton, T.C. Managing for elevated yield of moose in interior Alaska. J. Wildl. Manag. 2009, 73, 314–327. [Google Scholar] [CrossRef]
- Young, D.D., Jr.; Boertje, R.D.; Seaton, C.T.; Kellie, K.A. Intensive management of moose at high density: Impediments, achievements, and recommendations. Alces 2006, 42, 41–48. [Google Scholar]
- National Research Council. Wolves, Bears, and Their Prey in Alaska: Biological and Social Challenges in Wildlife Management; National Academy Press: Washington, DC, USA, 1997. [Google Scholar]
- Clark, J.T.; Horn, J.S.; Hebblewhite, M.; Luis, A.D. Stochastic predation exposes prey to predator pit and local extinction. Oikos 2021, 130, 300–309. [Google Scholar] [CrossRef]
- Keech, M.A.; Lindberg, M.S.; Boertje, R.D.; Valkenburg, P.; Taras, P.D.; Boudreau, T.A.; Beckmen, K.B. Effects of predator treatments, individual traits, and environment on moose survival in Alaska. J. Wildl. Manag. 2011, 75, 1361–1380. [Google Scholar] [CrossRef]
- Boertje, R.D.; Valkenburg, P.; McNay, M.E. Increases in moose, caribou, and wolves following wolf control in Alaska. J. Wildl. Manag. 1996, 60, 474–489. [Google Scholar] [CrossRef]
- Boertje, R.D.; Frye, G.G.; Young, D.D. Lifetime sex-specific moose mortality during an intentional population reduction. J. Wildl. Manag. 2020, 84, 6–19. [Google Scholar] [CrossRef]
- Ripple, W.J.; Miller, S.D.; Schoen, J.W.; Rabinowitch, S.P. Large carnivores under assault in Alaska. PLoS Biol. 2019, 17, 332–346. [Google Scholar] [CrossRef] [PubMed]
- Boutin, S. Predation and moose population dynamics: A critique. J. Wildl. Manag. 1992, 56, 116–127. [Google Scholar] [CrossRef]
- Alaska Department of Fish and Game. Intensive Management Protocol; Division of Wildlife Conservation, 2011; p. 44. Available online: https://www.adfg.alaska.gov/static/research/programs/intensivemanagement/pdfs/intensive_management_protocol.pdf (accessed on 2 June 2022).
- Alaska Department of Fish and Game. Board of Game Bear Conservation, Harvest, and Management Policy 2016-214-BOG. 2021. Available online: http://www.adfg.alaska.gov/static/regulations/regprocess/gameboard/pdfs/findings/16214.pdf (accessed on 8 August 2022).
- Miller, S.D.; Schoen, J.W.; Schwartz, C.C. Trends in brown bear reduction efforts in Alaska, 1980–2017. Ursus 2017, 28, 135–149. [Google Scholar] [CrossRef]
- Weixelman, D.A.; Bowyer, R.T.; Van Ballenberghe, V. Diet selection by Alaskan moose during winter: Effects of fire and forest succession. Alces 1998, 34, 213–238. [Google Scholar]
- Schmidt, J.I.; Ver Hoef, J.M.; Maier, J.A.; Bowyer, R.T. Catch per unit effort for moose: A new approach using Weibull regression. J. Wildl. Manag. 2005, 69, 1112–1124. [Google Scholar] [CrossRef]
- Schmidt, J.I.; Ver Hoef, J.M.; Bowyer, R.T. Antler size of Alaskan moose Alces alces gigas: Effects of population density, hunter harvest and use of guides. Wildl. Biol. 2007, 13, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Maier, J.A.K.; Ver Hoef, J.M.; McGuire, D.A.; Bowyer, R.T.; Saperstein, L.; Maier, A. Distribution and density of moose in relation to landscape characteristics: Effects of scale. Can. J. For. Res. 2005, 35, 2233–2243. [Google Scholar] [CrossRef]
- Alaska Department of Fish and Game. Annual Report to the Alaska Board of Game on Intensive Management for Moose with Wolf Predation Control in Unit 13; Division of Wildlife Conservation, February 2022; p. 14. Available online: https://www.adfg.alaska.gov/static/applications/web/nocache/research/programs/intensivemanagement/pdfs/2022_gmu_13_intensive_management_annual_report.pdf38D732401CD0CA2E43CBC5FFBF8CABDF/2022_gmu_13_intensive_management_annual_report.pdf (accessed on 2 June 2022).
- Ballard, W.B.; Miller, S.D. Effects of reducing brown bear density on moose calf survival in southcentral Alaska. Alces 1990, 26, 9–13. [Google Scholar]
- Miller, S.D. Brown Bears in Alaska: A Statewide Management Overview; Wildlife Technical Bulletin 11; Alaska Department of Fish and Game, Division of Wildlife Conservation: Juneau, AK, USA, 1993. Available online: http://www.adfg.alaska.gov/static/home/library/pdfs/wildlife/research_pdfs/miller_1993_adfg_tech_bulletin_11_brown_bears_in_alaska.pdf (accessed on 18 August 2022).
- Miller, S.D.; White, G.C.; Sellers, R.A.; Reynolds, H.V.; Schoen, J.W.; Titus, K.; Barnes, V.G., Jr.; Smith, R.B.; Nelson, R.R.; Ballard, W.B.; et al. Brown and black bear density estimation in Alaska using radiotelemetry and replicated mark-resight techniques. Wildl. Monogr. 1997, 133, 1–55. [Google Scholar]
- Miller, S.D.; Sellers, R.A.; Keay, J.A. Effects of hunting on brown bear cub survival and litter size in Alaska. Ursus 2003, 14, 130–152. [Google Scholar]
- Brockman, C.M.; Guttery, M.R.; Dale, R.W.; Schwanke, R.A.; Tobey, R.W.; Koons, D.N. Effect of harvest on a brown bear population in Alaska. J. Wildl. Manag. 2020, 84, 865–876. [Google Scholar] [CrossRef]
- Robbins, W.F. Black Bear Management Report of Sur-Vey-Inventory Activities 1 July 2010–30 June 2013; Harper, P., Ed.; Unit 13 Black Bear; Species Management Report ADF&G/DWC/SMR-2014-5; Alaska Department of Fish and Game: Juneau, AK, USA, 2008; Chapter 12; pp. 1–8. Available online: http://www.adfg.alaska.gov/static/home/library/pdfs/wildlife/mgt_rpts/08_blb.pdf (accessed on 2 June 2022).
- Derrick, T.R.; Thomas, J.M. Time-series analysis: The cross-correlation function. In Innovative Analyses of Human Movement; Stergiou, N., Ed.; Human Kinetics Publishers: Champaign, IL, USA, 2004; pp. 189–205. [Google Scholar]
- Neter, J.; Wasserman, W.; Kutner, M.H. Applied Linear Statistical Models: Regression, Analysis of Variance, and Experimental Designs, 2nd ed.; Irwin: Homewood, IL, USA, 1985. [Google Scholar]
- Nelson, C.R. Applied Time Series Analysis for Managerial Forecasting; Holden-Day Inc.: San Francisco, CA, USA, 1973. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer Science: New York, NY, USA, 2002. [Google Scholar]
- Arnold, T.W. Uninformative parameters and model selection using Akaike’s information criterion. J. Wildl. Manag. 2010, 74, 1175–1178. [Google Scholar] [CrossRef]
- Thalmann, J.C.; Bowyer, R.T.; Aho, K.A.; Weckerly, F.W.; McCullough, D.R. Antler and body size in black-tailed deer: An analysis of cohort effects. Adv. Ecol. 2015, 2015, 156041. [Google Scholar] [CrossRef]
- Zweiers, F.W.; von Storch, H. Taking Serial Correlation into Account in Tests of the Mean. J. Clim. 1994, 8, 336351. [Google Scholar]
- McCullough, D.R. The George Reserve Deer Herd: Ecology of a K-Selected Species; University of Michigan Press: Ann Arbor, MI, USA, 1979. [Google Scholar]
- Westgate, M.J.; Likens, G.E.; Lindenmayer, D.B. Adaptive management of biological systems: A review. Biol. Conserv. 2013, 158, 128–139. [Google Scholar] [CrossRef]
- Mitchell, C.D.; Chaney, R.; Aho, K.; Bowyer, R.T. Population characteristics, morphometry, and growth of harvested gray wolves and coyotes in Alaska. Arctic 2022, 75, 242–256. [Google Scholar] [CrossRef]
- Robbins, W.F. Moose Management Report and Plan, Game Management Unit 13: Report Period 1 July 2010–30 June 2015; Species Management Report and Plan ADF&G/DWC/SMR&P-2018-47; Alaska Department of Fish and Game: Juneau, AK, USA, 2018. Available online: http://www.adfg.alaska.gov/index.cfm?adfg=librarypublications.wildlifemanagement (accessed on 9 August 2022).
- Testa, J.W. Interaction of top-down and bottom-up life history trade-offs in moose (Alces alces). Ecology 2004, 85, 453–1455. [Google Scholar] [CrossRef]
- Monteith, K.L.; Bleich, V.C.; Stephenson, T.R.; Pierce, B.M.; Conner, M.M.; Kie, J.G.; Bowyer, R.T. Life-history characteristics of mule deer: Effects of nutrition in a variable environment. Wildl. Monogr. 2014, 186, 1–56. [Google Scholar] [CrossRef]
- Hobbs, N.T.; Swift, D.M. Estimates of habitat carrying capacity incorporating explicit nutritional constraints. J. Wildl. Manag. 1985, 49, 814–822. [Google Scholar] [CrossRef]
- Beck, J.L.; Peek, J.M.; Strand, E.K. Estimates of elk summer range nutritional carrying capacity constrained by probabilities of habitat selection. J. Wildl. Manag. 2006, 70, 283–294. [Google Scholar] [CrossRef]
- Boertje, R.D.; Frye, G.G.; Young, D.D., Jr. Lifetime, known-age moose reproduction in a nutritionally stressed population. J. Wildl. Manag. 2019, 83, 6–19. [Google Scholar] [CrossRef]
- Paragi, T.F.; Seaton, C.T.; Kellie, K.A.; Boertje, R.D.; Kieland, K.; Young, D.D., Jr.; Keech, M.A.; DuBois, S.D. Browse removal, plant, condition, and twinning rates before and after short-term changes in moose density. Alces 2015, 51, 1–21. [Google Scholar]
- Hayes, F.P.; Milspaugh, J.J.; Bergman, E.J.; Callaway, R.M.; Bishop, C.J. Effects of willow nutrition and morphology on calving success of moose. J. Wildl. Manag. 2022, 86, e22175. [Google Scholar] [CrossRef]
- Stewart, K.M.; Bowyer, R.T.; Ruess, R.W.; Dick, B.L.; Kie, J.G. Herbivore optimization in North American elk: Consequences for theory and management. Wildl. Monogr. 2006, 167, 1–24. [Google Scholar] [CrossRef]
- Gaudry, W.; Gaillard, J.M.; Saïd, S.; Mårell, A.; Balzinger, C.; Rocquencourt, A.; Bonenfant, C. Population density and plant availability interplay to shape browsing intensity by roe deer in a deciduous forest. For. Ecol. Manag. 2002, 515, 120153. [Google Scholar] [CrossRef]
- Bowyer, R.T.; Pierce, B.M.; Duffy, L.K.; Haggstrom, A. Sexual segregation in moose: Effects of habitat manipulation. Alces 2001, 37, 109–122. [Google Scholar]
- Oehlers, S.A.; Bowyer, R.T.; Huettmann, F.; Person, D.K.; Kessler, W.B. Sex and scale: Implications for habitat selection by Alaskan moose Alces alces gigas. Wildl. Biol. 2011, 17, 67–84. [Google Scholar] [CrossRef] [Green Version]
- Bowyer, R.T. Sexual Segregation in Ungulates: Ecology, Behavior, and Conservation, Institute of Arctic Biology, University of Alaska Fairbanks: Fairbanks, AK, USA, 2022; in press.
- Theberge, J.B. Potentials for misinterpreting impacts of wolf predation through prey: Predator ratios. Wildl. Soc. Bull. 1990, 18, 188–192. [Google Scholar]
- Person, D.K.; Bowyer, R.T.; Van Ballenberghe, V. Density dependence of ungulates and functional responses of wolves: Effects on predator–prey ratios. Alces 2001, 37, 253–273. [Google Scholar]
- Monteith, K.L.; Long, R.A.; Stephenson, T.R.; Bleich, V.C.; Bowyer, R.T.; LaSharr, T. Horn size and nutrition in mountain sheep: Can ewe handle the truth? J. Wildl. Manag. 2018, 82, 67–84. [Google Scholar] [CrossRef]
- Treves, A.; Karanth, K.U. Human-carnivore conflict and perspectives on carnivore management worldwide. Conserv. Biol. 2003, 17, 1491–1499. [Google Scholar] [CrossRef]
- Laliberte, A.S.; Ripple, W.J. Range contractions of North American carnivores and ungulates. BioScience 2004, 64, 123–138. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, C.C.; Swenson, J.E.; Miller, S.D. Large carnivores, moose, and humans: A changing paradigm of predator management in the 21st Century. Alces 2011, 39, 41–63. [Google Scholar]
- Gilbert, S.; Haynes, L.T.; Lindberg, M.S.; Albert, D.M.; Kissing, M.; Lynch, L.; Person, D. Potential futures for coastal wolves and their ecosystem services in Alaska, with implications for management of a social-ecological system. Front. Ecol. Evolut. 2022, 10, 809371. [Google Scholar] [CrossRef]
- Bruskotter, J.T. The predator pendulum revisited: Social conflict over wolves and their management in the western United States. Wildl. Soc. Bull. 2013, 37, 674–679. [Google Scholar] [CrossRef]
- Kellert, S.R.; Black, M.; Reid Rush, C.; Bath, A.J. Human culture and large carnivore conservation in North America. Conserv. Biol. 1996, 10, 977–990. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miller, S.D.; Person, D.K.; Bowyer, R.T. Efficacy of Killing Large Carnivores to Enhance Moose Harvests: New Insights from a Long-Term View. Diversity 2022, 14, 939. https://doi.org/10.3390/d14110939
Miller SD, Person DK, Bowyer RT. Efficacy of Killing Large Carnivores to Enhance Moose Harvests: New Insights from a Long-Term View. Diversity. 2022; 14(11):939. https://doi.org/10.3390/d14110939
Chicago/Turabian StyleMiller, Sterling D., David K. Person, and R. Terry Bowyer. 2022. "Efficacy of Killing Large Carnivores to Enhance Moose Harvests: New Insights from a Long-Term View" Diversity 14, no. 11: 939. https://doi.org/10.3390/d14110939
APA StyleMiller, S. D., Person, D. K., & Bowyer, R. T. (2022). Efficacy of Killing Large Carnivores to Enhance Moose Harvests: New Insights from a Long-Term View. Diversity, 14(11), 939. https://doi.org/10.3390/d14110939