

Effects of Invasive Plant Diversity on Soil Microbial Communities



Abstract
:1. Introduction
2. Materials and Methods
2.1. Native and Invasive Species Preparation
2.2. Experimental Design
2.3. Soil Sample Collection and Physico-Chemical Analysis
2.4. Microbial Sample Collection and Sequencing
2.5. Bioinformatic Analysis
2.6. Statistical Analysis
3. Results
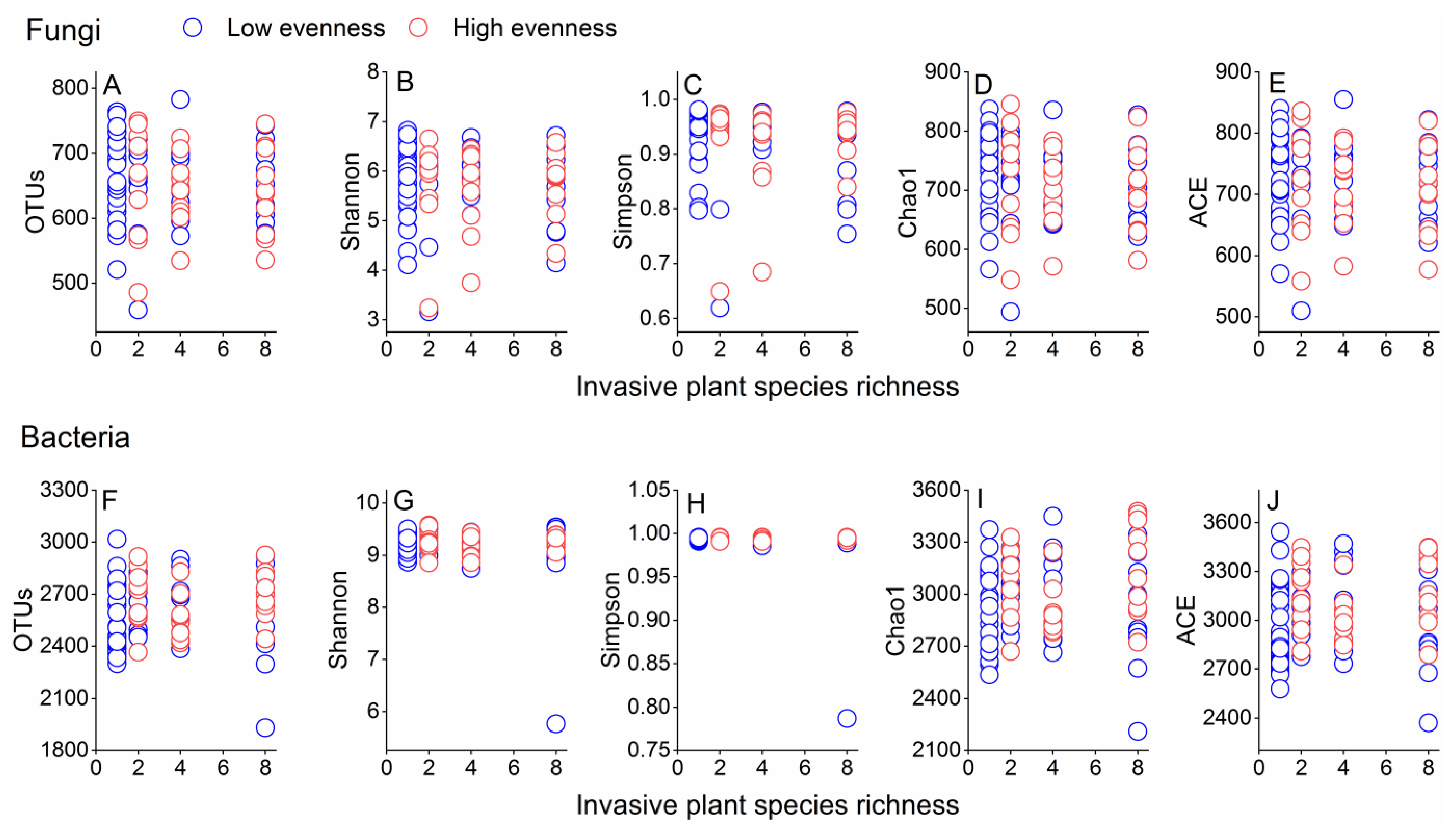
3.1. Effects of Invasive Species Diversity on Alpha Diversity of Soil Microbial Communities
3.2. Effects of Invasive Species Diversity on Soil Physico-Chemical Properties
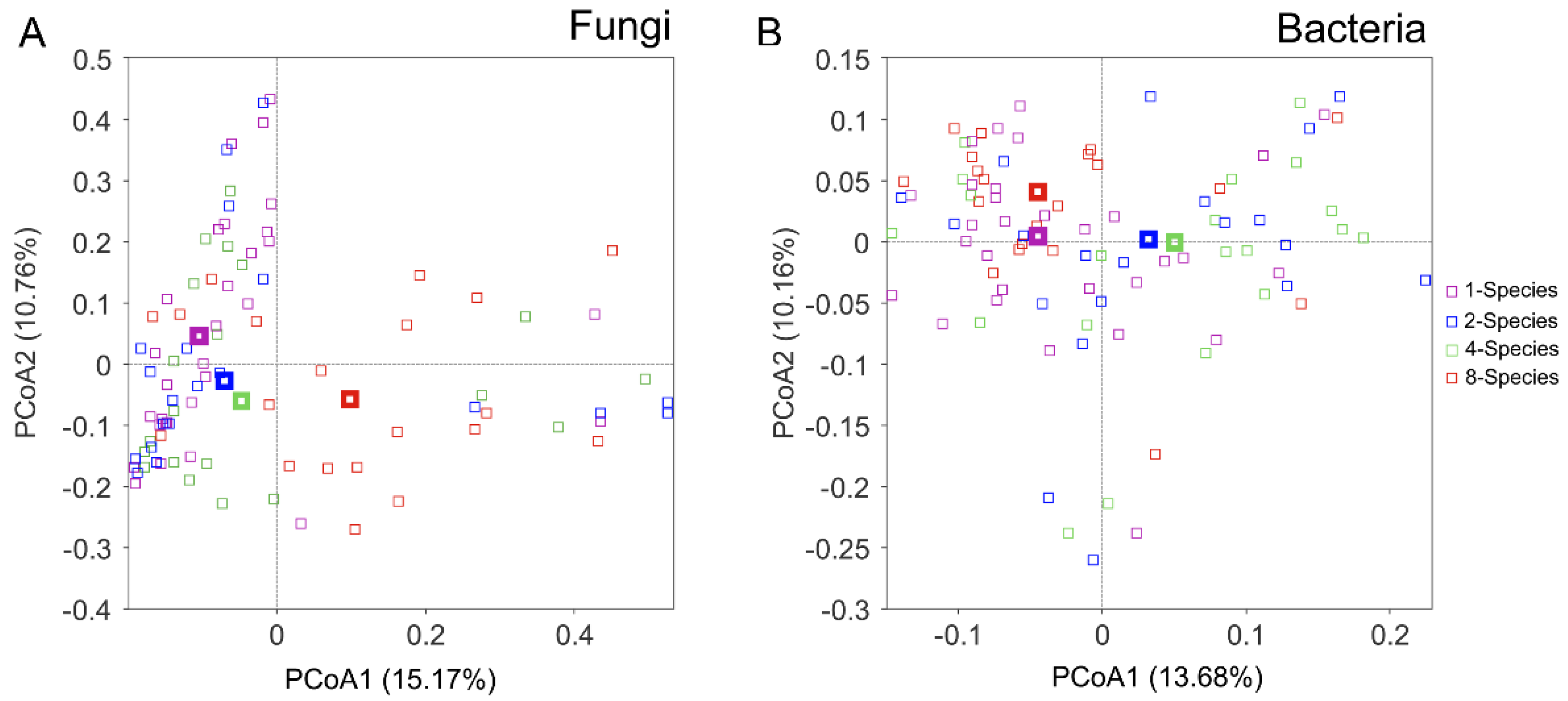
3.3. Effects of Invasive Species Diversity on Soil Microbial Community Composition
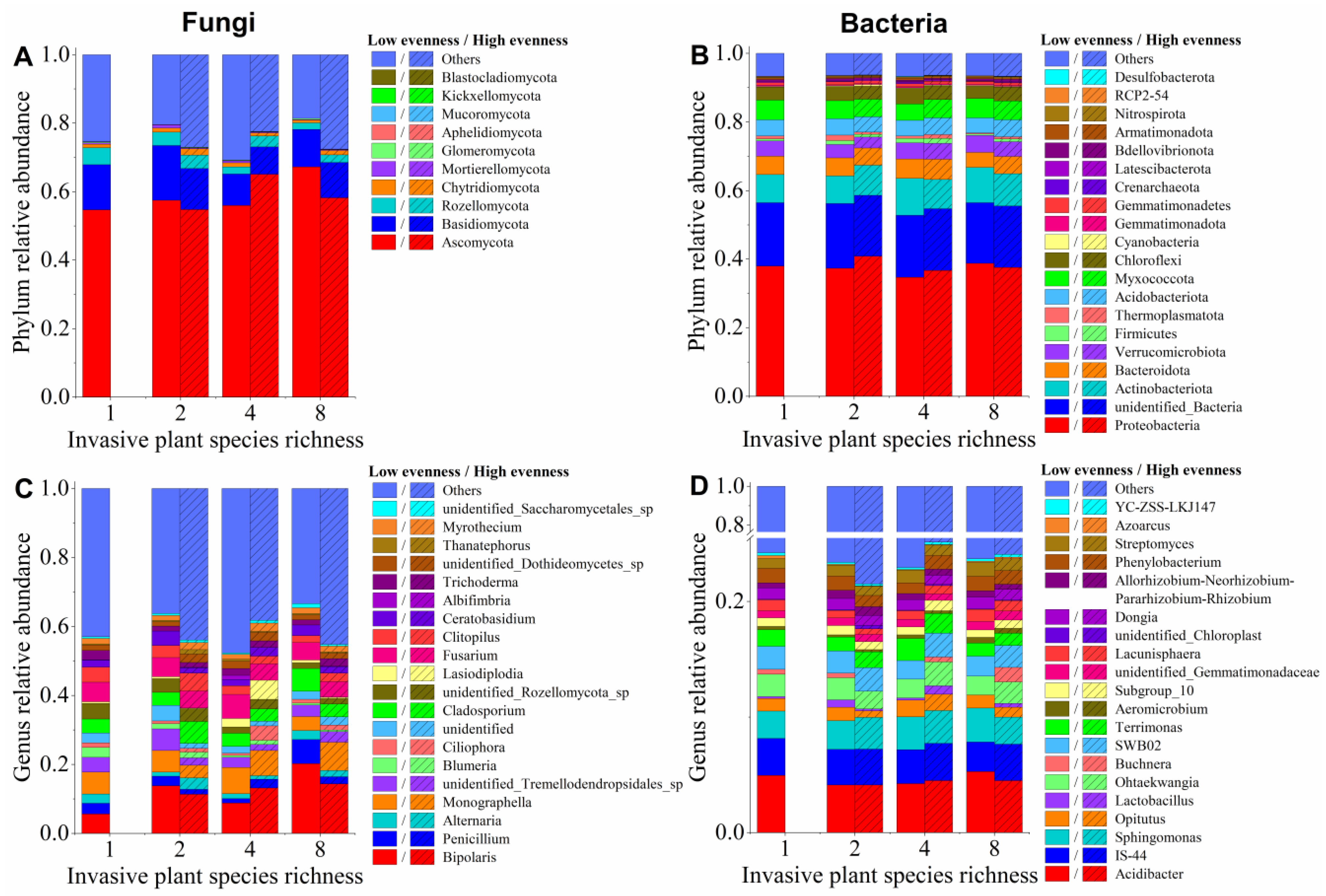
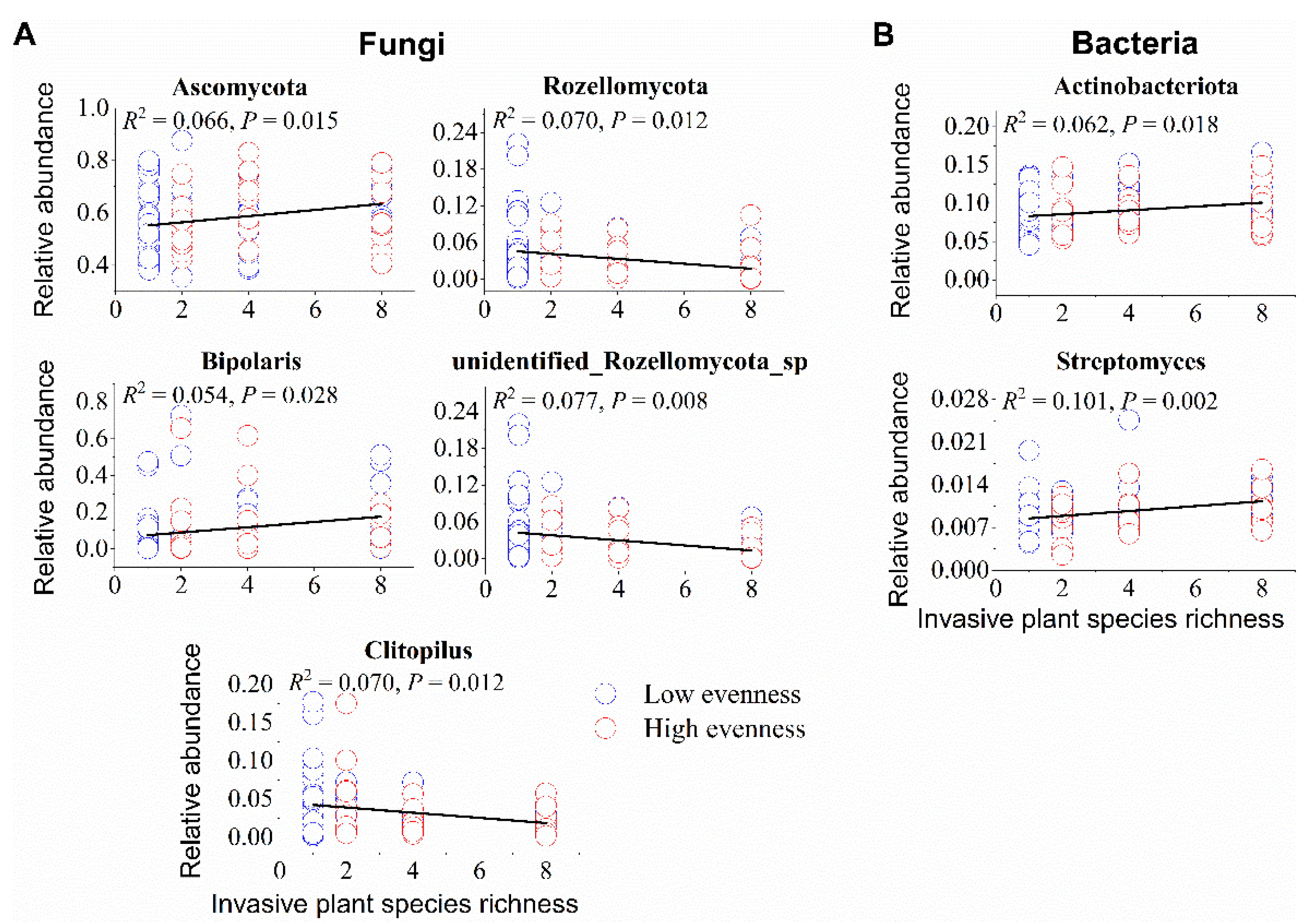
3.4. Effects of Invasive Species Diversity on Relative Abundances of Soil Microbial Taxa
4. Discussion
4.1. Effects of Invasive Species Diversity on Alpha Diversity of Soil Microbial Communities
4.2. Effects of Invasive Species Diversity on Soil Microbial Taxa and Community Composition
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1-species | 2-species | ||||||||||||||||||||||||||||||||||||||
| High evenness | Low evenness | ||||||||||||||||||||||||||||||||||||||
| Invasive species | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |||||||||
| Bidens pilosa | 24 | 12 | 12 | 22 | 2 | ||||||||||||||||||||||||||||||||||
| Bidens frondosa | 24 | 12 | 12 | 22 | 22 | ||||||||||||||||||||||||||||||||||
| Ageratum conyzoides | 24 | 12 | 12 | 2 | 22 | ||||||||||||||||||||||||||||||||||
| Sesbania cannabina | 24 | 12 | 12 | 2 | 22 | ||||||||||||||||||||||||||||||||||
| Talinum paniculatum | 24 | 12 | 12 | 2 | 2 | ||||||||||||||||||||||||||||||||||
| Celosia argentea | 24 | 12 | 12 | 2 | 22 | ||||||||||||||||||||||||||||||||||
| Solidago canadensis | 24 | 12 | 12 | 22 | 2 | ||||||||||||||||||||||||||||||||||
| Mirabilis jalapa | 24 | 12 | 12 | 22 | 22 | ||||||||||||||||||||||||||||||||||
| Alternanthera philoxeroides | 24 | 12 | 12 | 22 | 2 | ||||||||||||||||||||||||||||||||||
| Aster subulatus | 24 | 12 | 12 | 2 | 22 | ||||||||||||||||||||||||||||||||||
| 4-species | 8-species | ||||||||||||||||||||||||||||||||||||||
| High evenness | Low evenness | High evenness | Low evenness | ||||||||||||||||||||||||||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
| 6 | 6 | 6 | 6 | 18 | 3 | 2 | 1 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 1 | 3 | 2 | 1 | 13 | 1 | 1 | 2 | ||||||||||||||||
| 6 | 6 | 6 | 6 | 3 | 3 | 18 | 1 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 13 | 1 | 1 | 2 | 1 | 1 | 2 | 3 | ||||||||||||||||
| 6 | 6 | 6 | 6 | 18 | 1 | 2 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 13 | 3 | 1 | 1 | 1 | 2 | 2 | 1 | ||||||||||||||||
| 6 | 6 | 6 | 6 | 2 | 3 | 2 | 18 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 1 | 1 | 1 | 13 | 3 | 1 | 1 | ||||||||||||||||
| 6 | 6 | 6 | 6 | 2 | 1 | 2 | 18 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 2 | 1 | 13 | 2 | 2 | 2 | 3 | 2 | ||||||||||||||||
| 6 | 6 | 6 | 6 | 1 | 3 | 18 | 18 | 2 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 2 | 1 | 2 | 2 | 13 | 3 | 1 | 1 | |||||||||||||||
| 6 | 6 | 6 | 6 | 18 | 2 | 2 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 13 | |||||||||||||||
| 6 | 6 | 6 | 6 | 1 | 3 | 2 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 1 | 3 | 1 | 1 | 1 | 2 | 13 | 1 | |||||||||||||||||
| 6 | 6 | 6 | 6 | 1 | 18 | 1 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 1 | 2 | 1 | 1 | 3 | 1 | 1 | 13 | |||||||||||||||||
| 6 | 6 | 6 | 6 | 18 | 1 | 3 | 1 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 1 | 2 | 13 | 2 | 3 | 2 | 1 | 1 | ||||||||||||||||
| Bidens pilosa | Bidens frondosa | Ageratum conyzoides | Sesbania cannabina | Talinum paniculatum | Celosia argentea | Solidago canadensis | Mirabilis jalapa | Alternanthera philoxeroides | Aster subulatus | |
|---|---|---|---|---|---|---|---|---|---|---|
| Fungi | ||||||||||
| Phylum | ||||||||||
| Ascomycota | −0.375 * | 0.265 | −0.328 | −0.465 ** | −0.248 | 0.052 | −0.241 | 0.337 | −0.474 ** | −0.340 |
| Basidiomycota | 0.195 | 0.124 | 0.080 | 0.531 ** | 0.179 | −0.114 | 0.202 | −0.219 | 0.367 * | −0.182 |
| Rozellomycota | 0.265 | −0.225 | 0.541 ** | 0.391 * | 0.066 | 0.065 | 0.099 | 0.186 | 0.481 ** | 0.174 |
| Chytridiomycota | −0.178 | −0.281 | 0.314 | −0.166 | −0.007 | 0.243 | −0.106 | −0.152 | 0.229 | 0.044 |
| Mortierellomycota | −0.151 | −0.037 | 0.293 | 0.352 | −0.102 | −0.278 | 0.331 | −0.256 | 0.370 * | 0.598 ** |
| Glomeromycota | 0.126 | 0.206 | 0.145 | −0.220 | 0.237 | −0.312 | 0.066 | 0.382 * | −0.173 | −0.045 |
| Mucoromycota | −0.001 | −0.078 | −0.126 | 0.492 ** | 0.174 | 0.069 | 0.102 | −0.100 | 0.018 | 0.148 |
| Monoblepharomycota | 0.255 | −0.122 | 0.052 | −0.189 | 0.549 ** | 0.063 | −0.005 | −0.077 | −0.100 | −0.079 |
| Genus | ||||||||||
| Bipolaris | −0.309 | 0.419 * | −0.352 | −0.389 * | −0.426 * | −0.350 | −0.139 | 0.197 | −0.429 * | −0.282 |
| Alternaria | 0.026 | −0.366 * | −0.225 | −0.090 | 0.094 | 0.380 * | −0.043 | 0.161 | −0.281 | 0.068 |
| Monographella | −0.181 | −0.242 | 0.514 ** | −0.155 | 0.174 | 0.297 | −0.045 | −0.083 | −0.213 | −0.155 |
| unidentified_Tremellodendropsidales_sp | 0.038 | −0.007 | −0.162 | 0.323 | 0.101 | −0.076 | 0.128 | −0.340 | 0.398 * | −0.196 |
| Blumeria | 0.028 | −0.182 | −0.142 | 0.388 * | 0.270 | 0.153 | −0.106 | 0.516 ** | 0.417 * | 0.160 |
| unidentified | −0.026 | −0.058 | −0.148 | 0.288 | 0.075 | −0.089 | 0.156 | −0.304 | 0.366 * | −0.191 |
| Cladosporium | −0.020 | −0.317 | −0.230 | 0.051 | 0.006 | 0.101 | −0.116 | 0.280 | −0.187 | −0.078 |
| unidentified_Rozellomycota_sp | 0.279 | −0.237 | 0.588 ** | 0.445 * | 0.048 | 0.097 | 0.056 | 0.298 | 0.476 ** | 0.124 |
| Clitopilus | 0.668 ** | 0.195 | 0.293 | 0.570 ** | 0.157 | −0.035 | 0.187 | 0.113 | 0.256 | 0.137 |
| Albifimbria | −0.124 | −0.164 | −0.114 | −0.153 | 0.037 | 0.266 | 0.090 | −0.096 | −0.123 | 0.346 |
| Trichoderma | −0.050 | 0.093 | 0.440 * | 0.112 | 0.329 | −0.164 | −0.034 | −0.178 | 0.137 | 0.121 |
| unidentified_Dothideomycetes_sp | 0.335 | −0.038 | 0.049 | −0.051 | 0.289 | −0.089 | −0.181 | 0.104 | −0.058 | −0.167 |
| Bacteria | ||||||||||
| Phylum | ||||||||||
| Proteobacteria | −0.013 | 0.207 | −0.139 | 0.197 | 0.267 | −0.062 | 0.225 | 0.058 | 0.060 | −0.498 ** |
| unidentified_Bacteria | 0.242 | −0.147 | 0.221 | 0.229 | −0.186 | −0.024 | 0.027 | 0.062 | 0.488 ** | −0.113 |
| Actinobacteriota | −0.431 * | 0.061 | −0.180 | −0.478 ** | −0.358 * | −0.117 | −0.271 | 0.152 | −0.277 | 0.169 |
| Verrucomicrobiota | −0.001 | −0.100 | 0.212 | −0.209 | 0.142 | 0.056 | −0.115 | −0.295 | −0.357 * | −0.055 |
| Firmicutes | 0.006 | −0.019 | −0.008 | −0.185 | −0.133 | −0.055 | −0.176 | −0.064 | −0.182 | 0.506 ** |
| Thermoplasmatota | 0.149 | −0.269 | 0.394 * | 0.068 | 0.013 | 0.294 | −0.203 | −0.066 | 0.159 | 0.527 ** |
| Acidobacteriota | 0.132 | −0.400 * | −0.139 | 0.109 | 0.055 | 0.158 | 0.233 | −0.186 | −0.060 | 0.172 |
| Myxococcota | −0.029 | −0.169 | −0.056 | 0.490 ** | 0.244 | 0.421 * | −0.021 | −0.028 | −0.165 | −0.228 |
| Gemmatimonadota | −0.005 | −0.192 | 0.349 | −0.353 | −0.219 | −0.022 | 0.055 | −0.190 | 0.106 | 0.065 |
| Gemmatimonadetes | −0.024 | 0.363 * | −0.019 | −0.438 * | −0.241 | −0.270 | 0.339 | −0.178 | 0.007 | 0.076 |
| Latescibacterota | 0.228 | −0.286 | −0.155 | 0.332 | 0.406 * | 0.218 | 0.152 | −0.081 | −0.017 | 0.023 |
| Bdellovibrionota | 0.045 | −0.092 | 0.249 | 0.221 | 0.188 | 0.191 | 0.020 | −0.038 | −0.047 | −0.377 * |
| Nitrospirota | −0.106 | −0.036 | 0.042 | −0.123 | −0.063 | −0.065 | 0.381 * | −0.287 | 0.005 | −0.383 * |
| RCP2-54 | −0.005 | −0.025 | 0.121 | 0.333 | −0.034 | 0.107 | 0.118 | 0.172 | 0.374 * | −0.164 |
| Desulfobacterota | 0.072 | 0.097 | 0.146 | −0.221 | −0.056 | −0.025 | −0.092 | −0.039 | −0.138 | 0.723 ** |
| Genus | ||||||||||
| Acidibacter | −0.086 | −0.271 | 0.227 | 0.010 | 0.386 * | 0.113 | 0.037 | −0.116 | −0.328 | −0.067 |
| IS-44 | 0.341 | −0.424 * | −0.105 | 0.376 * | 0.354 | 0.116 | −0.180 | 0.125 | 0.035 | 0.094 |
| Sphingomonas | −0.284 | 0.313 | −0.165 | −0.302 | −0.239 | −0.341 | 0.373 * | −0.099 | 0.102 | −0.296 |
| Opitutus | 0.089 | 0.071 | 0.320 | −0.309 | 0.048 | −0.014 | −0.237 | −0.189 | −0.325 | 0.171 |
| Lactobacillus | −0.059 | −0.068 | −0.010 | −0.033 | −0.065 | −0.097 | −0.045 | −0.130 | −0.111 | 0.469 ** |
| Ohtaekwangia | 0.146 | −0.278 | −0.272 | 0.430 * | 0.084 | 0.142 | −0.021 | 0.037 | 0.088 | −0.062 |
| SWB02 | 0.184 | −0.178 | −0.121 | 0.337 | 0.066 | 0.214 | 0.158 | −0.025 | −0.057 | 0.063 |
| Terrimonas | 0.192 | −0.019 | 0.368 * | 0.007 | 0.190 | −0.172 | 0.100 | 0.235 | 0.044 | 0.165 |
| Aeromicrobium | −0.237 | 0.331 | −0.242 | −0.237 | −0.240 | −0.144 | −0.343 | −0.199 | −0.283 | −0.202 |
| Subgroup_10 | −0.053 | −0.186 | 0.018 | −0.188 | −0.071 | −0.131 | 0.252 | −0.076 | −0.010 | 0.445 * |
| Lacunisphaera | −0.109 | −0.113 | 0.225 | −0.065 | 0.194 | 0.078 | −0.061 | −0.307 | −0.241 | −0.380 * |
| Dongia | −0.073 | −0.065 | −0.052 | 0.075 | 0.244 | −0.118 | 0.229 | −0.044 | 0.216 | −0.342 |
| Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium | −0.324 | 0.401 * | −0.002 | 0.080 | −0.343 | −0.288 | −0.062 | 0.154 | 0.239 | −0.358 * |
| Streptomyces | −0.268 | −0.017 | −0.089 | −0.524 ** | −0.363 * | −0.199 | −0.410 * | −0.160 | −0.372 * | −0.070 |
| Azoarcus | 0.383 * | 0.007 | −0.210 | 0.247 | 0.359 * | 0.344 | −0.224 | 0.077 | 0.288 | −0.307 |
| YC-ZSS-LKJ147 | 0.059 | −0.290 | 0.315 | −0.314 | −0.232 | −0.010 | −0.034 | −0.252 | 0.039 | −0.112 |
| Fungi | Bacteria | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| OTUs | Shannon | Simpson | Chao1 | ACE | OTUs | Shannon | Simpson | Chao1 | ACE | |
| pH | 0.069 | 0.092 | 0.101 | 0.070 | 0.065 | −0.009 | 0.060 | 0.044 | 0.030 | −0.005 |
| Organic matter | 0.004 | 0.066 | 0.009 | −0.013 | −0.004 | 0.054 | −0.154 | −0.191 | 0.044 | 0.063 |
| Total N | 0.012 | 0.002 | −0.012 | −0.009 | −0.010 | −0.008 | −0.063 | −0.046 | −0.011 | 0.028 |
| NH4_N | −0.080 | −0.043 | −0.066 | −0.082 | −0.079 | 0.006 | 0.073 | 0.052 | −0.047 | −0.052 |
| NO3_N | −0.275 ** | −0.106 | −0.033 | −0.239 * | −0.241 * | −0.067 | 0.042 | 0.065 | −0.140 | −0.124 |
| Total P | −0.252 * | −0.191 | −0.123 | −0.265 ** | −0.240 * | 0.039 | 0.074 | 0.073 | 0.029 | 0.051 |
| Available P | −0.225 * | −0.207 * | −0.182 | −0.188 | −0.178 | −0.225 * | −0.068 | 0.017 | −0.261 * | −0.250 * |
| Available K | −0.039 | −0.097 | −0.139 | −0.025 | −0.015 | −0.158 | −0.034 | −0.020 | −0.208 * | −0.203 * |
References
- van Kleunen, M.; Dawson, W.; Essl, F.; Pergl, J.; Winter, M.; Weber, E.; Kreft, H.; Weigelt, P.; Kartesz, J.; Nishino, M.; et al. Global exchange and accumulation of non-native plants. Nature 2015, 525, 100–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambertini, M.; Leape, J.; Marton-Lefèvre, J.; Mittermeier, R.A.; Rose, M.; Robinson, J.G.; Stuart, S.N.; Waldman, B.; Genovesi, P. Invasives: A major conservation threat. Science 2011, 333, 404–405. [Google Scholar] [CrossRef] [PubMed]
- Atwater, D.Z.; Callaway, R.M. Extended consequences of selection by exotic invaders on natives. New Phytol. 2022, 233, 588–591. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Shi, J.; Xie, Q.; Jiang, Y.; Yu, N.; Wang, E. Nutrient exchange and regulation in arbuscular mycorrhizal symbiosis. Mol. Plant 2017, 10, 1147–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pysek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.T.; Dawson, W.; Essl, F.; Foxcroft, L.C.; Genovesi, P.; et al. Scientists’ warning on invasive alien species. Biol. Rev. 2020, 95, 1511–1534. [Google Scholar] [CrossRef]
- Simberloff, D.; Martin, J.L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; Garcia-Berthou, E.; Pascal, M.; et al. Impacts of biological invasions: What’s what and the way forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef] [Green Version]
- Vila, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarosik, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pysek, P. Ecological impacts of invasive alien plants: A meta-analysis of their effects on species, communities and ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef]
- Xu, H.; Liu, Q.; Wang, S.; Yang, G.; Xue, S. A global meta-analysis of the impacts of exotic plant species invasion on plant diversity and soil properties. Sci. Total Environ. 2022, 810, 152286. [Google Scholar] [CrossRef]
- Pisani, D.; Pazienza, P.; Perrino, E.V.; Caporale, D.; De Lucia, C. The economic valuation of ecosystem services of biodiversity components in protected areas: A review for a framework of analysis for the Gargano National Park. Sustainability 2021, 13, 11726. [Google Scholar] [CrossRef]
- Stinca, A.; Musarella, C.M.; Rosati, L.; Laface, V.L.A.; Licht, W.; Fanfarillo, E.; Wagensommer, R.P.; Galasso, G.; Fascetti, S.; Esposito, A.; et al. Italian vascular flora: New findings, updates and exploration of floristic similarities between regions. Diversity 2021, 13, 600. [Google Scholar] [CrossRef]
- Byun, C.; Lee, E.J. Ecological application of biotic resistance to control the invasion of an invasive plant, Ageratina altissima. Ecol. Evol. 2017, 7, 2181–2192. [Google Scholar] [CrossRef] [PubMed]
- Hoffberg, S.L.; Mauricio, R.; Hall, R.J. Control or re-treat? Model-based guidelines for managing established plant invasions. Biol. Invasions 2018, 20, 1387–1402. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, Y.; Brunel, C.; van Kleunen, M. Soil-microorganism-mediated invasional meltdown in plants. Nat. Ecol. Evol. 2020, 4, 1612–1621. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.L.; Bai, J.H.; Tebbe, C.C.; Huang, L.B.; Jia, J.; Wang, W.; Wang, X.; Yu, L.; Zhao, Q.Q. Plant invasion reconstructs soil microbial assembly and functionality in coastal salt marshes. Mol. Ecol. 2022, 31, 4478–4494. [Google Scholar] [CrossRef] [PubMed]
- Torres, N.; Herrera, I.; Fajardo, L.; Bustamante, R.O. Meta-analysis of the impact of plant invasions on soil microbial communities. BMC Evol. Biol. 2021, 21, 172. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Wang, J.; He, J.Z. Plant diversity enhances soil fungal diversity and microbial resistance to plant invasion. Appl. Environ. Microbiol. 2021, 87, e00251-00221. [Google Scholar] [CrossRef]
- Li, Y.-P.; Feng, Y.-L.; Chen, Y.-J.; Tian, Y.-H. Soil microbes alleviate allelopathy of invasive plants. Sci. Bull. 2015, 60, 1083–1091. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.-J.; Li, Q.; Chen, F.-X.; Xu, H.-Y.; Inderjit; Wan, F.-H. Arbuscular mycorrhizal fungi facilitate growth and competitive ability of an exotic species Flaveria bidentis. Soil Biol. Biochem. 2017, 115, 275–284. [Google Scholar] [CrossRef]
- van der Putten, W.H.; Klironomos, J.N.; Wardle, D.A. Microbial ecology of biological invasions. ISME J. 2007, 1, 28–37. [Google Scholar] [CrossRef] [Green Version]
- Stefanowicz, A.M.; Stanek, M.; Nobis, M.; Zubek, S. Species-specific effects of plant invasions on activity, biomass, and composition of soil microbial communities. Biol. Fertil. Soils 2016, 52, 841–852. [Google Scholar] [CrossRef]
- Jacobs, K.; Conradie, T.; Jacobs, S. Microbial communities in the Fynbos region of South Africa: What happens during woody alien plant invasions. Diversity 2020, 12, 254. [Google Scholar] [CrossRef]
- Kuebbing, S.E.; Classen, A.T.; Simberloff, D. Two co-occurring invasive woody shrubs alter soil properties and promote subdominant invasive species. J. Appl. Ecol. 2014, 51, 124–133. [Google Scholar] [CrossRef] [Green Version]
- Gibbons, S.M.; Lekberg, Y.; Mummey, D.L.; Sangwan, N.; Ramsey, P.W.; Gilbert, J.A. Invasive plants rapidly reshape soil properties in a grassland ecosystem. mSystems 2017, 2, e00178-00116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, T.; Du, X.; Peng, Y.; Guo, W.; Zhao, C.; Losapio, G. Invasive species allelopathy decreases plant growth and soil microbial activity. PLoS ONE 2021, 16, e0246685. [Google Scholar] [CrossRef] [PubMed]
- Ehrenfeld, J.G. Effects of exotic plant invasions on soil nutrient cycling processes. Ecosystems 2003, 6, 503–523. [Google Scholar] [CrossRef]
- Gaggini, L.; Rusterholz, H.-P.; Baur, B. The invasive plant Impatiens glandulifera affects soil fungal diversity and the bacterial community in forests. Appl. Soil Ecol. 2018, 124, 335–343. [Google Scholar] [CrossRef]
- Bell, J.K.; Siciliano, S.D.; Lamb, E.G. A survey of invasive plants on grassland soil microbial communities and ecosystem services. Sci. Data 2020, 7, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Li, B.; Wu, J.; Hu, S. Invasive plants differentially affect soil biota through litter and rhizosphere pathways: A meta-analysis. Ecol. Lett. 2019, 22, 200–210. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Zhou, J.; Liu, J.; Jiang, K.; Xiao, H.; Du, D. Responses of the soil fungal communities to the co-invasion of two invasive species with different cover classes. Plant Biol. 2018, 20, 151–159. [Google Scholar] [CrossRef]
- Liao, C.; Peng, R.; Luo, Y.; Zhou, X.; Wu, X.; Fang, C.; Chen, J.; Li, B. Altered ecosystem carbon and nitrogen cycles by plant invasion: A meta-analysis. New Phytol. 2008, 177, 706–714. [Google Scholar] [CrossRef]
- Vogelsang, K.M.; Bever, J.D. Mycorrhizal densities decline in association with nonnative plants and contribute to plant invasion. Ecology 2009, 90, 399–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, M.A.; Beaulieu, M.-E.; Reshi, Z.A.; Qureshi, S.; Khasa, D.P. A cross-city molecular biogeographic investigation of arbuscular mycorrhizas in Conyza canadensis rhizosphere across native and non-native regions. Ecol. Process. 2015, 4, 7. [Google Scholar] [CrossRef] [Green Version]
- Řezáčová, V.; Řezáč, M.; Gryndler, M.; Hršelová, H.; Gryndlerová, H.; Michalová, T. Plant invasion alters community structure and decreases diversity of arbuscular mycorrhizal fungal communities. Appl. Soil Ecol. 2021, 167, 104039. [Google Scholar] [CrossRef]
- Custer, G.F.; van Diepen, L.T.A. Plant invasion has limited impact on soil microbial α-diversity: A meta-analysis. Diversity 2020, 12, 112. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, J.; Hu, B.; Zheng, W.-L.; Li, M.; Shen, Z.-X.; Yu, F.-H.; Schmid, B.; Li, M.-H. Richness, not evenness, of invasive plant species promotes invasion success into native plant communities via selection effects. Oikos 2022, 2022, e08966. [Google Scholar] [CrossRef]
- Wilsey, B.J.; Teaschner, T.B.; Daneshgar, P.P.; Isbell, F.I.; Polley, H.W. Biodiversity maintenance mechanisms differ between native and novel exotic-dominated communities. Ecol. Lett. 2009, 12, 432–442. [Google Scholar] [CrossRef] [Green Version]
- Kuebbing, S.E.; Souza, L.; Sanders, N.J. Effects of co-occurring non-native invasive plant species on old-field succession. For. Ecol. Manag. 2014, 324, 196–204. [Google Scholar] [CrossRef]
- Kuebbing, S.E.; Classen, A.T.; Sanders, N.J.; Simberloff, D. Above- and below-ground effects of plant diversity depend on species origin: An experimental test with multiple invaders. New Phytol. 2015, 208, 727–735. [Google Scholar] [CrossRef]
- Zak, D.R.; Tilman, D.; Parmenter, R.R.; Rice, C.W.; Fisher, F.M.; Vose, J.; Milchunas, D.; Martin, C.W. Plant production and soil microorganisms in late-successional ecosystems: A continental-scale study. Ecology 1994, 75, 2333–2347. [Google Scholar] [CrossRef]
- Waldrop, M.P.; Holloway, J.M.; Smith, D.B.; Goldhaber, M.B.; Drenovsky, R.E.; Scow, K.M.; Dick, R.; Howard, D.; Wylie, B.; Grace, J.B. The interacting roles of climate, soils, and plant production on soil microbial communities at a continental scale. Ecology 2017, 98, 1957–1967. [Google Scholar] [CrossRef]
- McTee, M.R.; Lekberg, Y.; Mummey, D.; Rummel, A.; Ramsey, P.W. Do invasive plants structure microbial communities to accelerate decomposition in intermountain grasslands? Ecol. Evol. 2017, 7, 11227–11235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broughton, L.C.; Gross, K.L. Patterns of diversity in plant and soil microbial communities along a productivity gradient in a Michigan old-field. Oecologia 2000, 125, 420–427. [Google Scholar] [CrossRef] [PubMed]
- Wilsey, B.J.; Chalcraft, D.R.; Bowles, C.M.; Willig, M.R. Relationships among indices suggest that richness is an incomplete surrogate for grassland biodiversity. Ecology 2005, 86, 1178–1184. [Google Scholar] [CrossRef]
- Wang, X.-Y.; Ge, Y.; Gao, S.; Chen, T.; Wang, J.; Yu, F.-H. Evenness alters the positive effect of species richness on community drought resistance via changing complementarity. Ecol. Indic. 2021, 133, 108464. [Google Scholar] [CrossRef]
- Wang, J.; Yu, F.-H. Effects of functional diversity and functional dominance on complementary light use depend on evenness. J. Veg. Sci. 2018, 29, 726–736. [Google Scholar] [CrossRef]
- Sun, Y.-Q.; Wang, J.; Shen, C.; He, J.-Z.; Ge, Y. Plant evenness modulates the effect of plant richness on soil bacterial diversity. Sci. Total Environ. 2019, 662, 8–14. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.Y.H.; Reich, P.B. Forest productivity increases with evenness, species richness and trait variation: A global meta-analysis. J. Ecol. 2012, 100, 742–749. [Google Scholar] [CrossRef]
- Filstrup, C.T.; King, K.B.S.; McCullough, I.M. Evenness effects mask richness effects on ecosystem functioning at macro-scales in lakes. Ecol. Lett. 2019, 22, 2120–2129. [Google Scholar] [CrossRef]
- Lamb, E.G.; Kennedy, N.; Siciliano, S.D. Effects of plant species richness and evenness on soil microbial community diversity and function. Plant Soil 2011, 338, 483–495. [Google Scholar] [CrossRef]
- Kirwan, L.; LÜScher, A.; SebastiÀ, M.T.; Finn, J.A.; Collins, R.P.; Porqueddu, C.; Helgadottir, A.; Baadshaug, O.H.; Brophy, C.; Coran, C.; et al. Evenness drives consistent diversity effects in intensive grassland systems across 28 European sites. J. Ecol. 2007, 95, 530–539. [Google Scholar] [CrossRef]
- Sadiqi, S.; Hamza, M.; Ali, F.; Alam, S.; Shakeela, Q.; Ahmed, S.; Ayaz, A.; Ali, S.; Saqib, S.; Ullah, F.; et al. Molecular characterization of bacterial isolates from soil samples and evaluation of their antibacterial potential against MDRS. Molecules 2022, 27, 6281. [Google Scholar] [CrossRef]
- Porazinska, D.L.; Farrer, E.C.; Spasojevic, M.J.; Bueno de Mesquita, C.P.; Sartwell, S.A.; Smith, J.G.; White, C.T.; King, A.J.; Suding, K.N.; Schmidt, S.K. Plant diversity and density predict belowground diversity and function in an early successional alpine ecosystem. Ecology 2018, 99, 1942–1952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergmann, G.T.; Bates, S.T.; Eilers, K.G.; Lauber, C.L.; Caporaso, J.G.; Walters, W.A.; Knight, R.; Fierer, N. The under-recognized dominance of Verrucomicrobia in soil bacterial communities. Soil Biol. Biochem. 2011, 43, 1450–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, W.; Yang, M.; Liu, Y.; Huang, H.; Ye, C.; Zheng, J.; Guo, C.; Hao, M.; He, X.; Zhu, S. Fertilizer N application rate impacts plant-soil feedback in a sanqi production system. Sci. Total Environ. 2018, 633, 796–807. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Schmitz, M.; Flynn, D.F.B.; Mwangi, P.N.; Schmid, R.; Scherer-Lorenzen, M.; Weisser, W.W.; Schmid, B. Consistent effects of biodiversity on ecosystem functioning under varying density and evenness. Folia Geobot. 2013, 48, 335–353. [Google Scholar] [CrossRef] [Green Version]
- Peay, K.G.; Baraloto, C.; Fine, P.V.A. Strong coupling of plant and fungal community structure across western Amazonian rainforests. ISME J. 2013, 7, 1852–1861. [Google Scholar] [CrossRef]
- Dassen, S.; Cortois, R.; Martens, H.; de Hollander, M.; Kowalchuk, G.A.; van der Putten, W.H.; de Deyn, G.B. Differential responses of soil bacteria, fungi, archaea and protists to plant species richness and plant functional group identity. Mol. Ecol. 2017, 26, 4085–4098. [Google Scholar] [CrossRef]
- Yang, N.; Zhang, Y.; Li, J.; Li, X.; Ruan, H.; Bhople, P.; Keiblinger, K.; Mao, L.; Liu, D. Interaction among soil nutrients, plant diversity and hypogeal fungal trophic guild modifies root-associated fungal diversity in coniferous forests of Chinese Southern Himalayas. Plant Soil 2022. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Cajthaml, T.; Põlme, S.; Hiiesalu, I.; Anslan, S.; Harend, H.; Buegger, F.; Pritsch, K.; Koricheva, J.; et al. Tree diversity and species identity effects on soil fungi, protists and animals are context dependent. ISME J. 2016, 10, 346–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldrop, M.P.; Zak, D.R.; Blackwood, C.B.; Curtis, C.D.; Tilman, D. Resource availability controls fungal diversity across a plant diversity gradient. Ecol. Lett. 2006, 9, 1127–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prober, S.M.; Leff, J.W.; Bates, S.T.; Borer, E.T.; Firn, J.; Harpole, W.S.; Lind, E.M.; Seabloom, E.W.; Adler, P.B.; Bakker, J.D.; et al. Plant diversity predicts beta but not alpha diversity of soil microbes across grasslands worldwide. Ecol. Lett. 2015, 18, 85–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, N.H.; Williams, L.J.; Vincent, J.B.; Stefanski, A.; Cavender-Bares, J.; Messier, C.; Paquette, A.; Gravel, D.; Reich, P.B.; Kennedy, P.G. Ectomycorrhizal fungal diversity and saprotrophic fungal diversity are linked to different tree community attributes in a field-based tree experiment. Mol. Ecol. 2016, 25, 4032–4046. [Google Scholar] [CrossRef]
- Fierer, N.; McCain, C.M.; Meir, P.; Zimmermann, M.; Rapp, J.M.; Silman, M.R.; Knight, R. Microbes do not follow the elevational diversity patterns of plants and animals. Ecology 2011, 92, 797–804. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Xu, T.; Fu, W.; Hu, Y.; Hu, H.; You, L.; Chen, B. Soil organic carbon and total nitrogen predict large-scale distribution of soil fungal communities in temperate and alpine shrub ecosystems. Eur. J. Soil Biol. 2021, 102, 103270. [Google Scholar] [CrossRef]
- Hooper, D.U.; Bignell, D.E.; Brown, V.K.; Brussard, L.; Dangerfield, J.M.; Wall, D.H.; Wardle, D.A.; Coleman, D.C.; Giller, K.E.; Lavelle, P.; et al. Interactions between aboveground and belowground biodiversity in terrestrial ecosystems: Patterns, mechanisms, and feedbacks. BioScience 2000, 50, 1049–1061. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Põlme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.; Villarreal Ruiz, L.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Fungal biogeography. Global diversity and geography of soil fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef] [Green Version]
- Fierer, N. Embracing the unknown: Disentangling the complexities of the soil microbiome. Nat. Rev. Microbiol. 2017, 15, 579–590. [Google Scholar] [CrossRef]
- de Vries, F.T.; Manning, P.; Tallowin, J.R.B.; Mortimer, S.R.; Pilgrim, E.S.; Harrison, K.A.; Hobbs, P.J.; Quirk, H.; Shipley, B.; Cornelissen, J.H.C.; et al. Abiotic drivers and plant traits explain landscape-scale patterns in soil microbial communities. Ecol. Lett. 2012, 15, 1230–1239. [Google Scholar] [CrossRef]
- Liu, S.; Yu, H.; Yu, Y.; Huang, J.; Zhou, Z.; Zeng, J.; Chen, P.; Xiao, F.; He, Z.; Yan, Q. Ecological stability of microbial communities in Lake Donghu regulated by keystone taxa. Ecol. Indic. 2022, 136, 108695. [Google Scholar] [CrossRef]
- Banerjee, S.; Schlaeppi, K.; van der Heijden, M.G.A. Keystone taxa as drivers of microbiome structure and functioning. Nat. Rev. Microbiol. 2018, 16, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Pratt, R.G. Johnsongrass, yellow foxtail, and broadleaf signalgrass as new hosts for six species of Bipolaris, Curvularia, and Exserohilum pathogenic to bermudagrass. Plant Dis. 2006, 90, 528. [Google Scholar] [CrossRef] [PubMed]
- Kumar, J.; Schäfer, P.; Hückelhoven, R.; Langen, G.; Baltruschat, H.; Stein, E.; Nagarajan, S.; Kogel, K.-H. Bipolaris sorokiniana, a cereal pathogen of global concern: Cytological and molecular approaches towards better control. Mol. Plant Pathol. 2002, 3, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Flory, S.L.; Kleczewski, N.; Clay, K. Ecological consequences of pathogen accumulation on an invasive grass. Ecosphere 2011, 2, 120. [Google Scholar] [CrossRef]
- Stricker, K.B.; Harmon, P.F.; Goss, E.M.; Clay, K.; Luke Flory, S. Emergence and accumulation of novel pathogens suppress an invasive species. Ecol. Lett. 2016, 19, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Merzaeva, O.V.; Shirokikh, I.G. Colonization of plant rhizosphere by actinomycetes of different genera. Microbiology 2006, 75, 226–230. [Google Scholar] [CrossRef]
- Kaushik, R.; Pandit, M.K.; Meyerson, L.A.; Chaudhari, D.S.; Sharma, M.; Dhotre, D.; Shouche, Y.S. Contrasting composition, diversity and predictive metabolic potential of the rhizobacterial microbiomes associated with native and invasive Prosopis congeners. Curr. Microbiol. 2021, 78, 2051–2060. [Google Scholar] [CrossRef]
- Franco-Correa, M.; Quintana, A.; Duque, C.; Suarez, C.; Rodríguez, M.X.; Barea, J.-M. Evaluation of actinomycete strains for key traits related with plant growth promotion and mycorrhiza helping activities. Appl. Soil Ecol. 2010, 45, 209–217. [Google Scholar] [CrossRef]
- Stefan, L.; Hartmann, M.; Engbersen, N.; Six, J.; Schöb, C. Positive effects of crop diversity on productivity driven by changes in soil microbial composition. Front. Microbiol. 2021, 12, 660749. [Google Scholar] [CrossRef] [PubMed]
- Olanrewaju, O.S.; Babalola, O.O. Streptomyces: Implications and interactions in plant growth promotion. Appl. Microbiol. Biotechnology 2019, 103, 1179–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Wan, F.; Zhao, M. Distinct soil microbial communities under Ageratina adenophora invasions. Plant Biol. 2022, 24, 430–439. [Google Scholar] [CrossRef] [PubMed]




| Richness (R) | Evenness (E) | R’ E | ||||
|---|---|---|---|---|---|---|
| Variable | F | P | F | P | F | P |
| (A) Soil fungal communities | ||||||
| OTUs | 0.515 | 0.477 | 0.337 | 0.564 | 0.298 | 0.588 |
| Shannon | 0.555 | 0.461 | 0.007 | 0.933 | 0.292 | 0.592 |
| Simpson | 0.286 | 0.596 | 0.033 | 0.857 | 0.781 | 0.381 |
| Chao1 | 0.284 | 0.597 | 0.170 | 0.682 | 0.357 | 0.553 |
| ACE | 0.341 | 0.563 | 0.128 | 0.722 | 0.316 | 0.577 |
| (B) Soil bacterial communities | ||||||
| OTUs | 0.585 | 0.449 | 1.344 | 0.252 | 0.008 | 0.931 |
| Shannon | 0.568 | 0.456 | 1.856 | 0.180 | 0.768 | 0.385 |
| Simpson | 2.155 | 0.150 | 2.204 | 0.144 | 1.433 | 0.237 |
| Chao1 | 1.668 | 0.204 | 2.688 | 0.108 | 0.587 | 0.447 |
| ACE | 1.232 | 0.274 | 2.529 | 0.118 | 0.150 | 0.701 |
| (C) Soil physico-chemical properties | ||||||
| pH | 0.012 | 0.913 | 0.004 | 0.951 | 0.448 | 0.506 |
| Organic matter | 0.019 | 0.891 | 0.724 | 0.399 | 1.317 | 0.257 |
| Total N | 1.508 | 0.227 | 1.257 | 0.268 | 2.303 | 0.136 |
| NH4_N | 0.021 | 0.886 | 1.835 | 0.182 | 0.843 | 0.363 |
| NO3_N | 0.005 | 0.944 | 2.427 | 0.126 | 2.755 | 0.104 |
| Total P | 0.475 | 0.495 | 0.277 | 0.602 | 0.037 | 0.849 |
| Available P | 0.058 | 0.811 | 2.977 | 0.091 | 0.801 | 0.375 |
| Available K | 0.023 | 0.882 | 0.884 | 0.352 | 3.272 | 0.077 |
| Richness | Evenness | pH | Organic matter (g/kg) | Total N (mg/g) | NH4_N (mg/kg) | NO3_N (mg/kg) | Total P (mg/g) | Available P (mg/kg) | Available K (mg/kg) |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Low | 7.27 ± 0.04 | 285.05 ± 17.48 | 0.77 ± 0.07 | 10.45 ± 1.20 | 33.76 ± 3.24 | 3.45 ± 0.06 | 122.05 ± 7.49 | 163.27 ± 8.90 |
| 2 | Low | 7.35 ± 0.05 | 303.00 ± 37.82 | 0.78 ± 0.08 | 11.90 ± 2.29 | 37.29 ± 6.68 | 3.43 ± 0.14 | 125.64 ± 17.72 | 182.28 ± 31.35 |
| High | 7.34 ± 0.06 | 328.04 ± 27.28 | 0.58 ± 0.06 | 8.74 ± 1.33 | 35.72 ± 7.62 | 3.48 ± 0.11 | 122.13 ± 15.25 | 189.38 ± 24.27 | |
| 4 | Low | 7.34 ± 0.05 | 275.48 ± 37.52 | 0.67 ± 0.09 | 9.96 ± 1.69 | 36.20 ± 6.91 | 3.32 ± 0.10 | 132.27 ± 12.46 | 180.98 ± 27.74 |
| High | 7.26 ± 0.09 | 303.12 ± 18.88 | 0.65 ± 0.06 | 8.21 ± 0.61 | 31.32 ± 3.71 | 3.47 ± 0.12 | 105.33 ± 11.21 | 161.78 ± 13.50 | |
| 8 | Low | 7.27 ± 0.04 | 296.57 ± 27.73 | 0.66 ± 0.05 | 11.12 ± 2.22 | 43.31 ± 6.00 | 3.39 ± 0.20 | 139.36 ± 13.88 | 191.89 ± 18.65 |
| High | 7.29 ± 0.05 | 278.40 ± 11.35 | 0.70 ± 0.05 | 11.11 ± 1.48 | 26.67 ± 4.40 | 3.37 ± 0.20 | 115.02 ± 16.12 | 150.62 ± 16.54 |
| df | F | R2 | P | ||
|---|---|---|---|---|---|
| (A) Comparison among different richness of alien invasive plant species | |||||
| Fungi | |||||
| 1 vs. 2 species | 1 (48) | 0.704 | 0.014 | 0.872 | |
| 1 vs. 4 species | 1 (48) | 1.135 | 0.023 | 0.291 | |
| 1 vs. 8 species | 1 (48) | 3.048 | 0.060 | 0.001 | |
| 2 vs. 4 species | 1 (38) | 0.747 | 0.019 | 0.807 | |
| 2 vs. 8 species | 1 (38) | 1.803 | 0.045 | 0.030 | |
| 4 vs. 8 species | 1 (38) | 1.967 | 0.049 | 0.016 | |
| Bacteria | |||||
| 1 vs. 2 species | 1 (48) | 1.333 | 0.027 | 0.113 | |
| 1 vs. 4 species | 1 (48) | 1.767 | 0.036 | 0.017 | |
| 1 vs. 8 species | 1 (48) | 1.149 | 0.023 | 0.242 | |
| 2 vs. 4 species | 1 (38) | 0.763 | 0.020 | 0.812 | |
| 2 vs. 8 species | 1 (38) | 1.595 | 0.040 | 0.026 | |
| 4 vs. 8 species | 1 (38) | 1.791 | 0.045 | 0.021 | |
| (B) Comparison among different evenness of alien invasive plant species | |||||
| Fungi | |||||
| Low vs. high evenness of the 2 species | 1 (18) | 0.910 | 0.048 | 0.517 | |
| Low vs. high evenness of the 4 species | 1 (18) | 0.637 | 0.034 | 0.929 | |
| Low vs. high evenness of the 8 species | 1 (18) | 0.737 | 0.039 | 0.844 | |
| Bacteria | |||||
| Low vs. high evenness of the 2 species | 1 (18) | 0.694 | 0.037 | 0.930 | |
| Low vs. high evenness of the 4 species | 1 (18) | 0.814 | 0.043 | 0.593 | |
| Low vs. high evenness of the 8 species | 1 (18) | 0.958 | 0.051 | 0.501 | |
| Variable | Richness (R) | Evenness (E) | R × E | |||
|---|---|---|---|---|---|---|
| F | P | F | P | F | P | |
| (A) Fungi | ||||||
| Phylum | ||||||
| Ascomycota | 4.896 | 0.033 | 0.119 | 0.732 | 0.894 | 0.349 |
| Rozellomycota | 7.365 | 0.010 | 0.093 | 0.762 | 0.180 | 0.673 |
| Genus | ||||||
| Bipolaris | 5.203 | 0.028 | 0.022 | 0.883 | 0.275 | 0.602 |
| unidentified_Rozellomycota_sp | 7.835 | 0.008 | 0.043 | 0.836 | 0.011 | 0.918 |
| Clitopilus | 6.076 | 0.018 | 0.606 | 0.440 | 0.222 | 0.639 |
| (B) Bacteria | ||||||
| Phylum | ||||||
| Actinobacteriota | 4.920 | 0.033 | 1.838 | 0.182 | 0.844 | 0.363 |
| Bdellovibrionota | 0.252 | 0.619 | 0.210 | 0.649 | 7.782 | 0.008 |
| Armatimonadota | 1.065 | 0.309 | 4.358 | 0.042 | 0.018 | 0.894 |
| Genus | ||||||
| Sphingomonas | 1.465 | 0.234 | 0.067 | 0.796 | 4.576 | 0.038 |
| Buchnera | 0.314 | 0.579 | 0.918 | 0.343 | 7.560 | 0.008 |
| Streptomyces | 8.822 | 0.005 | 6.024 | 0.018 | 0.126 | 0.724 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Wang, X.; Wang, W.; Wang, J.; Yu, F. Effects of Invasive Plant Diversity on Soil Microbial Communities. Diversity 2022, 14, 992. https://doi.org/10.3390/d14110992
Wang X, Wang X, Wang W, Wang J, Yu F. Effects of Invasive Plant Diversity on Soil Microbial Communities. Diversity. 2022; 14(11):992. https://doi.org/10.3390/d14110992
Chicago/Turabian StyleWang, Xiaoyan, Xue Wang, Wei Wang, Jiang Wang, and Feihai Yu. 2022. "Effects of Invasive Plant Diversity on Soil Microbial Communities" Diversity 14, no. 11: 992. https://doi.org/10.3390/d14110992
APA StyleWang, X., Wang, X., Wang, W., Wang, J., & Yu, F. (2022). Effects of Invasive Plant Diversity on Soil Microbial Communities. Diversity, 14(11), 992. https://doi.org/10.3390/d14110992