
New Fossils of Stegosaurs from the Upper Jurassic of the Eastern Iberian Peninsula (Spain)


Abstract
:1. Introduction
Institutional Abbreviations
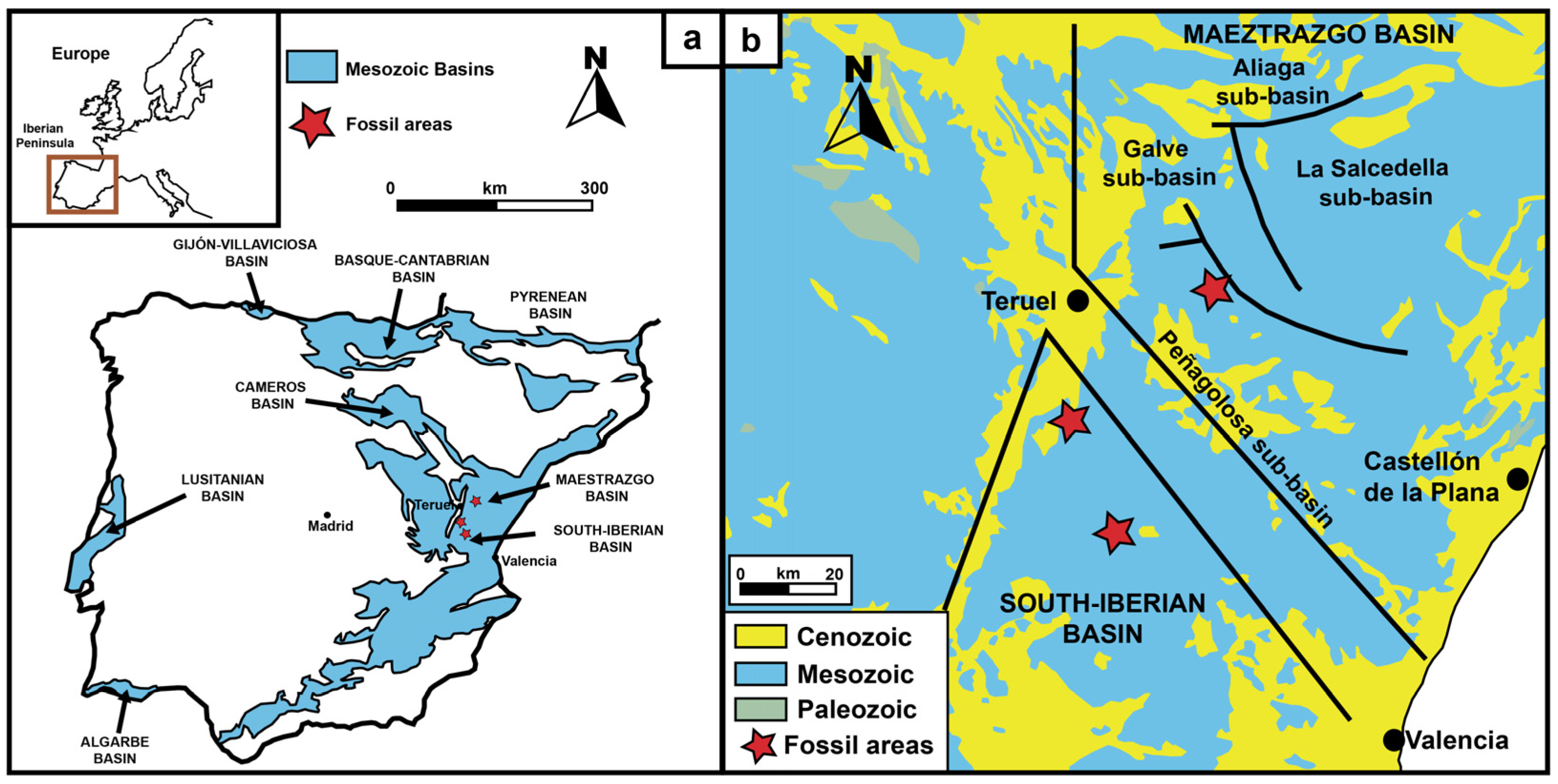
2. Geographical and Geological Context
3. Materials and Methods
4. Results and Discussion
4.1. Systematic Paleontology (CT-61-1 and MPA D-110)
4.1.1. Descriptions
- CT-61-1 (Figure 2) is a left humerus. It is robust, short and has an expanded distal end. The lateral and medial margins are concave and almost straight, respectively, in anterior and posterior views. The humeral head is not preserved. Regarding the deltopectoral crest, it is mostly destroyed, with only its distal region preserved. On the anterior surface, an oblique ridge extends from the distal base of the deltopectoral crest towards the medial (ulnar) condyle. A robust supinator ridge can be observed in the lateral surface of the distal end. The condylar region is rough and partially preserved. The anterior half of the medial condyle is not preserved. The medial condyle is slightly expanded posteriorly. The lateral (radial) condyle is robust and sub-circular in ventral view.
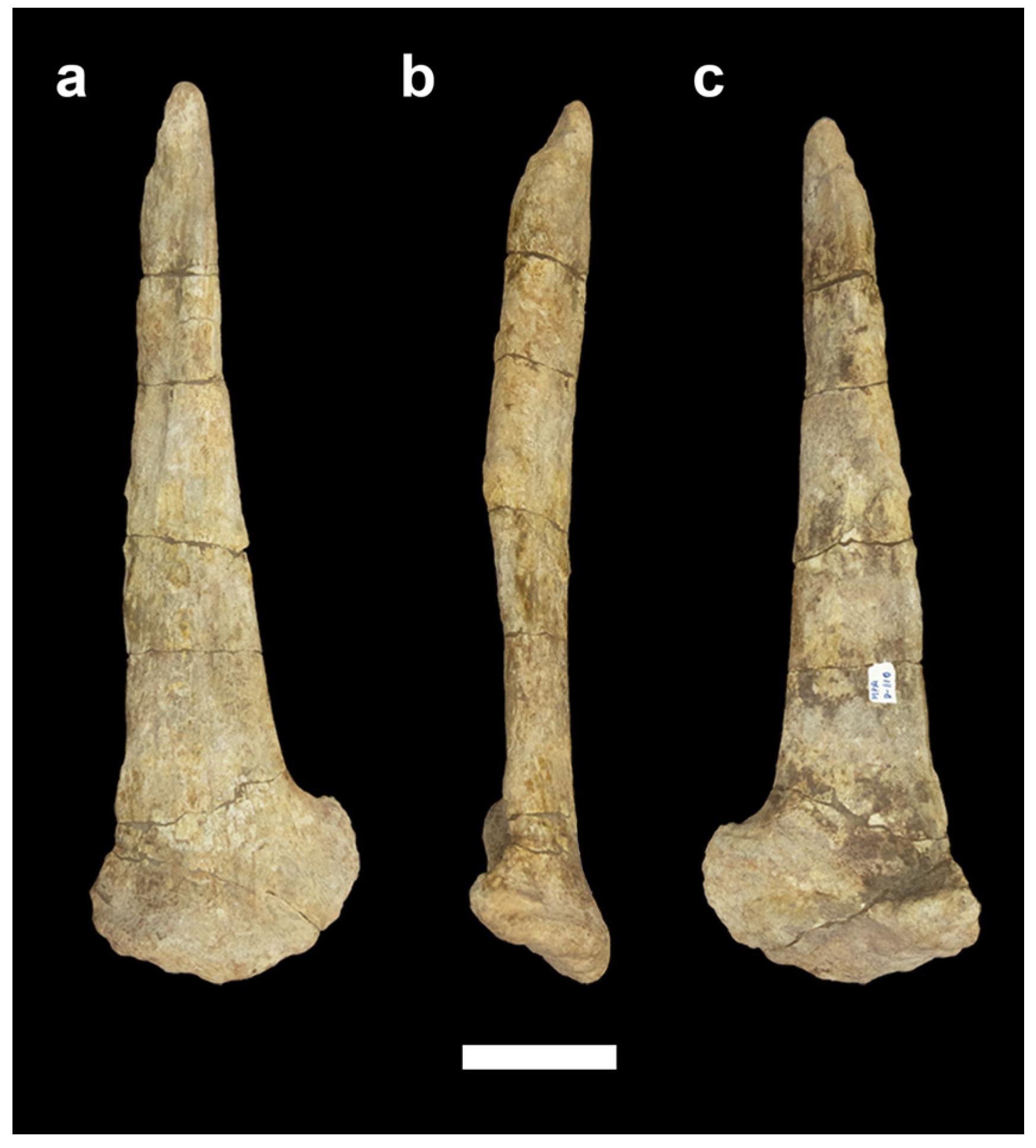
- MPA D-110 (Figure 3) is a proximal fragment of a slender dermal spine. The base of the spine is wider anteroposteriorly than dorsoventrally. In lateral and medial views, the basal region is moderately expanded with respect to the shaft. The surface of the base is slightly concave and C-shaped. Regarding the spine, the medial surface is slightly concave, especially in the proximal region. On the other hand, the lateral surface is slightly convex. The point of maximum width is located slightly above the beginning of the spine. Furthermore, approximately at this point, the spine begins to twist along its length. Anterior and posterior margins are convex and narrow. The cross-section is oval-shaped.
4.1.2. Comparisons and Discussions
- The left humerus CT-61-1 (Figure 2) shows a robust and strongly expanded transversally distal region that demonstrates thyreophoran affinities [53,54]. Furthermore, the presence of a pronounced anteroproximal expansion of the lateral condyle and an oblique ridge on the anterior surface that extends from the deltopectoral crest to the medial condyle support stegosaurian affinities [54]. In comparison to other stegosaurs, CT-61-1 shows an oblique crest less pronounced than Stegosaurus (Plate 33 [55]) or Kentrosaurus (Figure 4b [54]), being more similar to Dacentrurus armatus (Plate 17 [2]; Figure 9a–e [9]) and ‘Miragaia’ (Figure S3 [18]).
- The presence of a base that is not hollow with rounded edges, the absence of a longitudinal groove on the lateral and medial surfaces, a straight spine in the lateral and medial views and the absence of a groove posteriorly for a following spine indicates that MPA D-110 (Figure 3) belongs to a stegosaur [12]. MPA D-110 presents a slightly concave base with a moderate expansion very similar to the slender spine of the holotype of Dacentrurus armatus (Figure 10d,e [9]) and three spines from different localities of the Villar del Arzobispo Formation (Figure 2a,b [6]; Figure 1 [27]; Figure 4d–f [30]). This condition is different from the massive spines with strongly expanded and concave bases of several specimens referred to Dacentrurus or ‘Miragaia’ from Portugal and the UK (Figure 10f–m [9]; Figure 7a–c [13]; Figure 3.32 and 3.33 [16]; Figure 68 [19]). It is also different from the massive spines associated with the most caudal vertebrae related to Dacentrurus (Figure 3.34 [16]; Figure 4g–i [30]). The cross-section of MPA D-110 is oval, as is the spine of the specimen from Barranco del Curro (Figure 4f [30]). This is different from the sub-circular cross-section of the spine of the holotype of Dacentrurus armatus (Figure 1t [12]), the spine from Losilla (Figure 1 [27]) or the spine from La Canaleta [6] and from typical spines with a rhomboidal cross-section found in the Upper Jurassic of Europe, all of which are associated with Dacentrurus (Figure 10f–m [9]; Figures 5j and 7a–c [13]; Figure 3.33 [16]) or ‘Miragaia’ (Figure 68a [19]). It is also different from the sub-circular cross-section with a narrow anterior edge observed in Stegosaurus [55].
4.2. Systematic Paleontology (Cañada París specimen, MPA-653 and MPA D-1086)
4.2.1. Descriptions
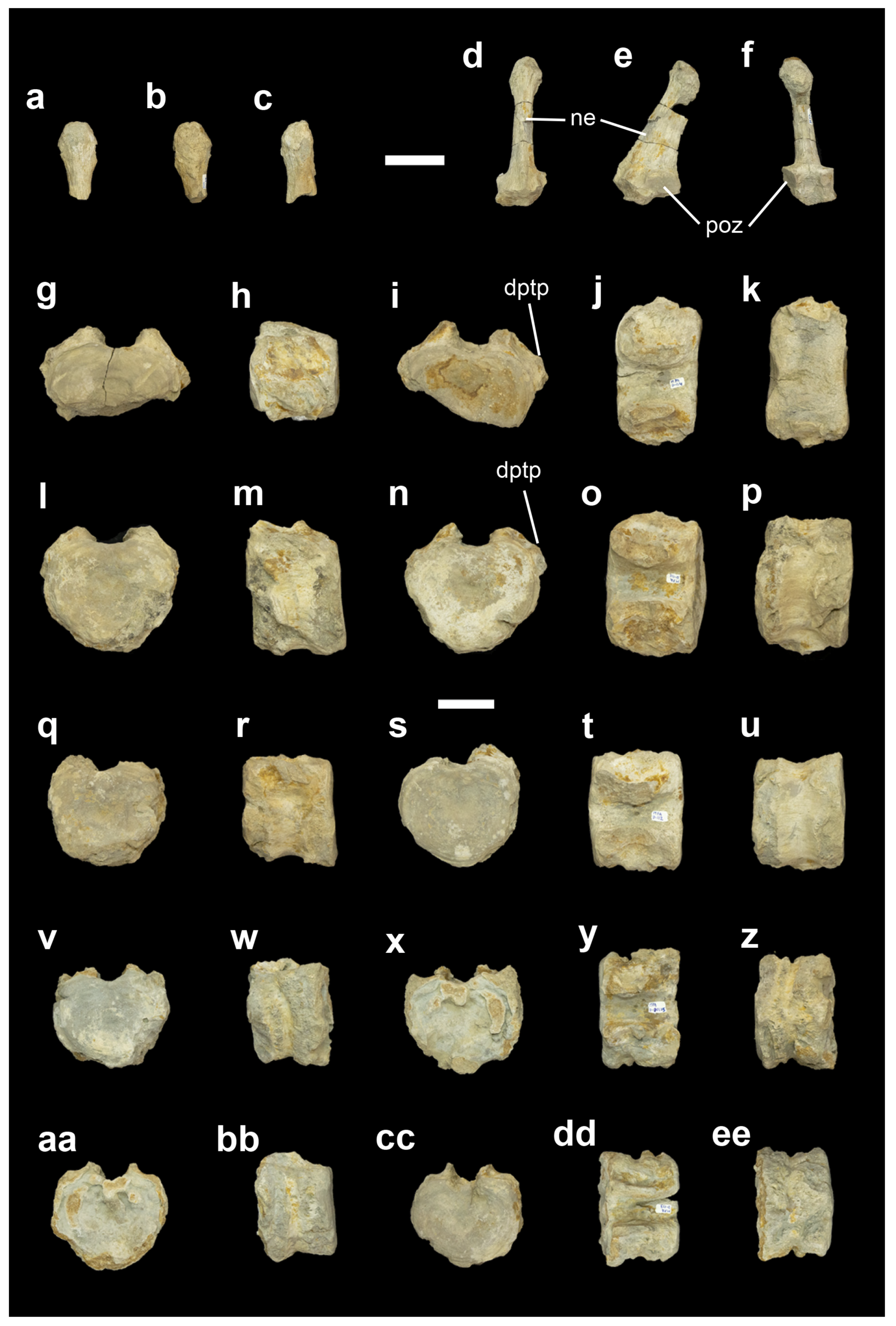
- MPA D-108 (Figure 4a–c) and MPA D-109 (Figure 4d–f) are two neural arch fragments. In particular, MPA D-108 consists of the apical region of a neural spine. The top of the neural spine has a circular morphology, and it is anteroposteriorly and mediolaterally expanded. Owing to these features, MPA D-108 belongs to the caudal series. On the other hand, MPA D-109 is an almost complete neural arch. The postzygapophyses are located above the prezygapophyses. The articulation surfaces of the postzygapophyses are anteroposteriorly directed and oval-shaped. With regard to the neural spine, it is short, and its lateral and medial surfaces are straight. The apex is anteroposteriorly and mediolaterally expanded and presents a circular morphology. Owing to the morphology of the postzygapophyses and the size of the postzygapophyses and the neural spine, we identified MPA D-109 as a middle caudal neural arch, more posterior than MPA D-108.MPA D-114 (Figure 4g–k), MPA D-111 (Figure 4l–p), MPA D-112 (Figure 4q–u), MPA D-115 (Figure 4v–z) and MPA D-113 (Figure 4aa–ee) are vertebral centra. MPA D-115 and MPA D-133 are contiguous vertebral centra. All centra are amphicoelous and wider than they are tall and long. As we move backward through the series, the centra tend to reduce their size, to become longer and to present a more marked heart-shaped morphology of the articulation facets. The lateral surfaces are concave with transverse processes located on the upper half. In the proximal region of the transverse processes of MPA D-114 and MPA D-111, a dorsal process is observed. In ventral view, all centra have chevron facets but lack a keel or groove. Owing to the presence of heart-shaped articulation facets, transverse processes (two of them with dorsal processes) and chevron facets, the vertebral centra from Cañada París were identified as caudal vertebrae from the anterior and middle region.
- MPA-1086 (Figure 5a–f) and MPA-653 (Figure 5g–l) are vertebral centra. Both are amphicoelous and wider than they are tall and long. The articulation facets are heart-shaped. The lateral surfaces are concave with transverse processes located in the upper half of the centrum. MPA-653 presents a deep concavity in the left lateral located immediately under the transverse process, whereas, in the right lateral, the concavity is more superficial. In ventral view, neither centra have a keel or a groove, but they do have chevron facets. The dimensions of the centra, the shape of the articular facets and the presence of transverse processes and chevron facets led us to identify them as middle caudal centra.
4.2.2. Comparisons and Discussions
4.3. Systematic Paleontology (Barrihonda-El Humero specimen)
4.3.1. Description
4.3.2. Comparisons and Discussions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seeley, H.G. On the base of a large lacertian cranium from the Potton sands, presumably dinosaurian. Q. J. Geol. Soc Lond. 1874, 30, 690–692. [Google Scholar] [CrossRef] [Green Version]
- Owen, R. Monographs of the fossil Reptilia of the Mesozoic Formations. Pt. II. Genera Bothriospondylus, Cetiosaurus, Omosaurus. Palaeontogr. Soc. 1875, 29, 15–94. [Google Scholar]
- Maidment, S.C.R.; Norman, D.B.; Barrett, P.M.; Upchurch, P. Systematics and phylogeny of Stegosauria (Dinosauria: Ornithischia). J. Syst. Palaeontol. 2008, 6, 367–407. [Google Scholar] [CrossRef]
- Maidment, S.C.R. Stegosauria: A historical review of the body fossil record and phylogenetic relationship. Swiss J. Geosci. 2010, 103, 199–210. [Google Scholar] [CrossRef]
- Pereda-Suberbiola, X.; Galton, P.M.; Torcida, F.; Huerta, P.; Izquierdo, L.A.; Montero, D.; Pérez, G.; Urién, V. First stegosaurian dinosaur remains from the Early Cretaceous of Burgos (Spain), with a review of cretaceous stegosaurs. Rev. Esp. Paleontol. 2003, 18, 143–150. [Google Scholar] [CrossRef]
- Pereda-Suberbiola, X.; Galton, P.M.; Ruiz-Omeñaca, J.I.; Canudo, J.I. Dermal spines of stegosaurian dinosaurs from the Lower Cretaceous (Hauterivian–Barremian) of Galve (Teruel, Aragón, Spain). Geogaceta 2005, 38, 35–38. [Google Scholar]
- Allain, R.; Vullo, R.; Rozada, L.; Anquetin, J.; Bourgeais, R.; Goedert, J.; Lasseron, M.; Martin, J.E.; Pérez-García, A.; Peyre de Fabrègues, C.; et al. Vertebrate paleobiodiversity of the Early Cretaceous (Berriasian) Angeac-Charente Lagerstätte (southwestern France): Implications for continental faunal turnover at the J/K boundary. Geodiversitas 2022, 25, 683–752. [Google Scholar] [CrossRef]
- Nopzca, F. Notes on British dinosaurs IV: Stegosaurus priscus sp. nov. Geol. Mag. 1911, 8, 145–153. [Google Scholar]
- Galton, P.M. British plated dinosaurs (Ornithischia, Stegosauridae). J. Vertebr. Paleontol. 1985, 5, 211–254. [Google Scholar] [CrossRef]
- Hoffstetter, R. Quelques observations sur les Stégosaurinés. Bull. Mus. Natl. Hist. Nat. Paris 1957, 29, 537–547. [Google Scholar]
- Galton, P.M. A partial skeleton of the stegosaurian dinosaur Lexovisaurus from the uppermost Lower Callovian (Middle Jurassic) of Normandy, France. Geol. Palaeontol. 1990, 24, 185–199. [Google Scholar]
- Galton, P.M. Notes on plated dinosaurs (Ornithischia: Stegosauria), mostly on dermal armor from Middle and Upper Jurassic of England (also France, Iberia), with a revised diagnosis for Loricatosaurus priscus (Callovian, England). Neues Jahr. Geol. Paläontol. Abh. 2016, 282, 1–25. [Google Scholar] [CrossRef]
- Galton, P.M. Postcranial remains of stegosaurian dinosaur Dacentrurus from Upper Jurassic of France and Portugal. Geol. Palaeontol. 1991, 25, 299–327. [Google Scholar]
- Casanovas-Cladellas, M.L.; Santafé-Llopis, J.V.; Santisteban, C.; Pereda-Suberbiola, X. Estegosaurios (Dinosauria) del Jurásico Superior-Cretácico Inferior de la comarca de Los Serranos (Valencia, España). Rev. Esp. Paleontol. 1999, 14, 57–63. [Google Scholar] [CrossRef]
- Cobos, A.; Royo-Torres, R.; Luque, L.; Alcalá, L.; Mampel, L. An Iberian stegosaurs paradise: The Villar del Arzobispo Formation (Tithonian–Berriasian) in Teruel (Spain). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 293, 223–236. [Google Scholar] [CrossRef]
- Escaso, F. Historia Evolutiva de los Ornithischia (Dinosauria) del Jurásico Superior de Portugal. Ph.D. Thesis, Universidad Autónoma de Madrid, Madrid, Spain, 2014. [Google Scholar]
- Escaso, F.; Ortega, F.; Dantas, P.; Malafaia, E.; Pimentel, N.L.; Pereda-Suberbiola, X.; Sanz, J.L.; Kullberg, J.C.; Kullberg, M.C.; Barriga, F. New evidence of shared dinosaur across Upper Jurassic Proto-North Atlantic: Stegosaurus from Portugal. Naturwissenschaften 2007, 94, 367–374. [Google Scholar] [CrossRef]
- Mateus, O.; Maidment, S.C.R.; Christiansen, N.A. A new long–necked “sauropod–mimic” stegosaur and the evolution of the plated dinosaurs. Proc. R. Soc. Biol. Sci. 2009, 276, 1815–1821. [Google Scholar] [CrossRef] [Green Version]
- Costa, F.; Mateus, O. Dacentrurine stegosaurs (Dinosauria): A new specimen of Miragaia longicollum from the Late Jurassic of Portugal resolves taxonomical validity and shows the occurrence of the clade in North America. PLoS ONE 2019, 14, e0224263. [Google Scholar] [CrossRef]
- Cobos, A.; Gascó, F. New vertebral remains of the stegosaurian dinosaur Dacentrurus from Riodeva (Teruel, Spain). Geogaceta 2013, 53, 17–20. [Google Scholar]
- Cobos, A.; Alcalá, L.; Royo-Torres, R. The Dinosaur Route in El Castellar (Teruel, Spain): Palaeontology as a Factor of Territorial Development and Scientific Education in a Sparsely Inhabited Area. Geoheritage 2020, 12, 52. [Google Scholar] [CrossRef]
- Ortega, F.; Malafaia, E.; Escaso, F.; Pérez-García, A.; Dantas, P. Faunas de répteis do Jurássico Superior de Portugal. Paleolusitana 2009, 1, 43–56. [Google Scholar]
- Raven, T.J.; Maidment, S.C.R. A new phylogeny of Stegosauria (Dinosauria, Ornithischia). Palaeontology 2017, 60, 401–408. [Google Scholar] [CrossRef] [Green Version]
- Lapparent, A.F.; Zbyszewski, G. Les dinosauriens du Portugal. Dir. Gén. Min. Serv. Géol. 1957, 2, 1–63. [Google Scholar]
- Escaso, F.; Ortega, F.; Dantas, P.; Malafaia, E.; Silva, B.; Sanz, J.L. Elementos postcraneales de Dacentrurus (Dinosauria: Stegosauria) del Jurásico Superior de Moçafaneira (Torres Vedras, Portugal). In Proceedings of the Cantera Paleontológica, Cuenca, Spain, 18-21 April 2007. [Google Scholar]
- Ruiz-Omeñaca, J.; Piñuela, L.; Garcia-Ramos, J.; Pereda-Suberbiola, X. A dacentrurine stegosaur from the Late Jurassic of Asturias (Northern Spain). In Proceedings of the 69th Annual Meeting Society of Vertebrate Paleontology, Bristol, UK, 23–26 September 2009. [Google Scholar]
- Casanovas-Cladellas, M.L.; Santafé-Llopis, J.V.; Pereda-Suberbiola, X.; Santisteban, C. Presencia por primera vez en España de Dinosaurios estegosaurios (Cretácico Inferior de Aldea de Losilla, Valencia). Rev. Esp. Paleontol. 1995, 10, 83–89. [Google Scholar] [CrossRef]
- Santafé-Llopis, J.V. Los estegosaurios y su presencia en el Mesozoico de “Los Serranos” (Valencia). Zubía 1996, 14, 105–111. [Google Scholar]
- Suñer, M.; Martín, M. Un nuevo yacimiento del tránsito Jurásico-Cretácico de Alpuente (Los Serranos, Valencia, España): Resultados preliminares. Paleolusitana 2009, 1, 441–447. [Google Scholar]
- Company, J.; Pereda-Suberbiola, X.; Ruiz-Omeñaca, J.I. New stegosaurian (Ornithischia, Thyreophora) remains from Jurassic-Cretaceous transition beds of Valencia province (Southwestern Iberian Range, Spain). J. Iber. Geol. 2010, 36, 243–252. [Google Scholar] [CrossRef]
- Royo-Torres, R.; Cobos, A.; Luque, L.; Aberasturi, A.; Espílez, E.; Fierro, I.; González, A.; Mampel, L.; Alcalá, L. High European sauropod dinosaur diversity during Jurassic-Cretaceous transition in Riodeva (Teruel, Spain). Palaeontology 2009, 52, 1009–1027. [Google Scholar] [CrossRef]
- Alcalá, L.; Cobos, A.; Espílez, E.; Gascó, F.; Mampel, L.; Martín-Escorza, C.; Royo-Torres, R. Icnitas de dinosaurios en la Formación Villar del Arzobispo de Ababuj (Teruel, España). Geogaceta 2012, 51, 35–38. [Google Scholar]
- Cobos, A.; Alcalá, L.; Mampel, L. Stegosaurian footprints from the Jurassic Cretaceous transition in Teruel (Spain). In Proceedings of the 11th Symposium on Mesozoic Terrestrial Ecosystems, Gwangju, Republic of Korea, 15–18 August 2012. [Google Scholar]
- Mampel, L.; Cobos, A.; Alcalá, L.; Espílez, E.; Royo-Torres, R.; González, A.; Gascó, F. Icnitas de dinosaurios en Aguilar de Alfambra (Teruel, España). Rev. Inst. Estud. Turol. 2011, 93, 43–54. [Google Scholar]
- Casanovas-Cladellas, M.L.; Santafé-Llopis, J.V.; Santisteban, C. Dacentrurus armatus (Stegosauria, Dinosauria) del Cretácico Inferior de Los Serranos (Valencia, España). Rev. Esp. Paleontol. 1995, 10, 273–283. [Google Scholar] [CrossRef]
- Royo-Torres, R.; Cobos, A.; Alcalá, L. A Giant European Dinosaur and a New Sauropod Clade. Science 2006, 314, 1925–1927. [Google Scholar] [CrossRef]
- Campos-Soto, S.; Benito, M.I.; Cobos, A.; Caus, E.; Quijada, E.; Suarez-Gonzalez, P.; Mas, R.; Royo-Torres, R.; Alcalá, L. Revisiting the age and palaeoenvironments of the Upper Jurassic—Lower Cretaceous? dinosaur-bearing sedimentary record of Eastern Spain: Implications for Iberian palaeogeography. J. Iber. Geol. 2019, 45, 471–510. [Google Scholar] [CrossRef]
- Mas, R.; Alonso, A.; Meléndez, N. La Formación Villar del Arzobispo: Un ejemplo de llanuras de mareas siliciclásticas asociadas a plataformas carbonatadas. Jurásico terminal. (NE de Valencia y E de Cuenca). Publ. Geol. Univ. Autón. Barc. 1984, 20, 175–188. [Google Scholar]
- Mas, R.; García, A.; Salas, R.; Meléndez, A.; Alonso, A.; Aurell, M.; Bádenas, B.; Benito, M.I.; Carenas, J.F.; García-Hidalgo, J.; et al. Segunda fase de rifting: Jurásico Superior Cretácico Inferior. In Geología de España; Vera, J.A., Ed.; SGE-IGME: Madrid, Spain, 2004; pp. 503–510. [Google Scholar]
- Santisteban, C. El entorno geológico de los yacimientos con dinosaurios del Jurásico Superior y Cretácico Inferior del Levante Peninsular I. In Dinosaurios del Levante Peninsular, 2nd ed.; Poza, B., Galobart, A., Suñer, M., Nieto, E., Eds.; Institut Català de Paleontologia: Sabadell, Spain, 2008; pp. 34–39. [Google Scholar]
- Santisteban, C.; Santos-Cubedo, A. Relación entre playas aterrazadas y cauces encajados, en depósitos deltaicos de la Formación Villar del Arzobispo (Cuenca Íbero-Levantina). In Proceedings of the V Congreso Jurásico de España, Colunga, Spain, 8–11 September 2010. [Google Scholar]
- Aurell, M.; Bádenas, B.; Bello, J.; Delvene, G.; Meléndez, G.; Pérez-Urresti, I.; Ramajo, J. El Calloviense y el Jurásico Superior en la Cordillera Ibérica Nororiental y la Zona de Enlace con la Cordillera Costero-Catalana, en los sectores Sierra de Arcos, Calanda y Xerta-Paüls. Cuad. Geol. Ibér. 1999, 25, 73–110. [Google Scholar]
- Aurell, M.; Bádenas, B.; Gasca, J.M.; Canudo, J.I.; Liesa, C.L.; Soria, A.R.; Moreno-Azanza, M.; Najes, L. Stratigraphy and evolution of the Galve sub-basin (Spain) in the middle Tithonian-early Barremian: Implications for the setting and age of some dinosaur fossil sites. Cretac. Res. 2016, 65, 138–162. [Google Scholar] [CrossRef] [Green Version]
- Campos-Soto, S.; Benito, M.I.; Mas, R.; Caus, E.; Cobos, A.; Suarez-Gonzalez, P.; Quijada, I.E. Revisiting the Late Jurassic Early Cretaceous of the NW South Iberian Basin: New ages and sedimentary environments. J. Iber. Geol. 2016, 42, 69–94. [Google Scholar] [CrossRef] [Green Version]
- Pacios, D.; Campos-Soto, S.; Suarez-Gonzalez, P.; Benito, M.I.; Cobos, A.; Caus, E. Revisión cartográfica y estratigráfica del Jurásico Superior-Cretácico Inferior de Villel (Teruel). Geogaceta 2018, 63, 19–22. [Google Scholar]
- Aurell, M.; Mas, R.; Meléndez, A.; Salas, R. El tránsito Jurásico-Cretácico en la Cordillera Ibérica: Relación tectónicasedimentación y evolución paleogeográfica. Cuad. Geol. Ibér. 1994, 18, 369–396. [Google Scholar]
- Campos-Soto, S.; Cobos, A.; Esmeralda, C.; Benito, M.I.; Fernández-Labrador, L.; Suarez-Gonzalez, P.; Quijada, I.E.; Mas, R.; Royo-Torres, R.; Alcalá, L. Jurassic Coastal Park: A great diversity of palaeoenvironments for the dinosaurs of the Villar del Arzobispo Formation (Teruel, eastern Spain). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 485, 154–177. [Google Scholar] [CrossRef]
- Bengtson, P. Open nomenclature. Palaeontology 1988, 31, 223–227. [Google Scholar]
- Owen, R. Report on British fossil reptiles. Rep. Br. Assoc. Adv. Sci. 1842, 11, 60–204. [Google Scholar]
- Seeley, H.G. The classification of the Dinosauria. Rep. Br. Assoc. Adv. Sci. 1887, 58, 698–699. [Google Scholar] [CrossRef] [Green Version]
- Nopcsa, F. Die Dinosaurier der Seibenburgischen Landesteile Ungarns. Mitt. Jahrb. Kn. Ung. Geol. Reichsanst. 1915, 23, 1–26. [Google Scholar]
- Marsh, O.C. A new Order of extinct Reptilia (Stegosauria) from the Jurassic of the Rocky Mountains. Am. J. Sci. 1877, 14, 34–35. [Google Scholar] [CrossRef] [Green Version]
- Galton, P.M.; Upchurch, P. Stegosauria. In The Dinosauria, 2nd ed.; Weishampel, D.B., Dodson, P., Osmólska, H., Eds.; University of California Press: Berkeley, CA, USA, 2004; pp. 343–362. [Google Scholar]
- Rauhut, O.W.M.; Carballido, J.L.; Pol, D. First Osteological Record of a Stegosaur (Dinosauria, Ornithischia) from the Upper Jurassic of South America. J. Vertebr. Paleontol. 2021, 40, e1862133. [Google Scholar] [CrossRef]
- Ostrom, J.H.; McIntosh, J.S. Stegosaur plates. In Marsh’s Dinosaurs. The Collections from Como Bluff; Yale University Press: New Haven, CT, USA, 1966; pp. 249–362. [Google Scholar]
- Marsh, O.C. Principal characters of American Jurassic dinosaurs. Part III. Am. J. Sci. 1880, 19, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Lucas, F.A. Paleontological notes: The generic name Omosaurus. Science 1902, 402, 435. [Google Scholar] [CrossRef]
- Maidment, S.C.R.; Brassey, C.; Barrett, P.M. The Postcranial Skeleton of an Exceptionally Complete Individual of the Plated Dinosaur Stegosaurus stenops (Dinosauria: Thyreophora) from the Upper Jurassic Morrison Formation of Wyoming, U.S.A. PLoS ONE 2015, 10, e0138352. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, K.; Miles, C.A.; Cloward, K. New primitive Stegosaur from the Morrison Formation, Wyoming. In The Armored Dinosaurs; Carpenter, K., Ed.; Indiana University Press: Bloomington, IN, USA, 2001; pp. 55–75. [Google Scholar]
- Galton, P.M.; Carpenter, K. The plated dinosaur Stegosaurus longispinus Gilmore, 1914 (Dinosauria: Ornithischia; Upper Jurassic, western USA), type species of Alcovasaurus n. gen. Neues Jahrb. Geol. Paläontol. Abh. 2016, 279, 185–208. [Google Scholar] [CrossRef]
- Gilmore, C.W. Osteology of the armored dinosaurian in the United States National Museum, with special reference to the genus Stegosaurus. U. S. Nat. Mus. Bull. 1914, 89, 1–143. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CT-61 Site | L | PW | DW | |||
| Left humerus CT-61-1 | 628 * | 237 | 267 | |||
| Puntal de la Magdalena | L | BH | BW | |||
| Dermal spine MPA D-110 | 285 * | 55 | 89 | |||
| Cañada París site | CL | AAFH | AAFW | PAFH | PAFW | NAH |
| Anterior caudal centrum MPA D-114 | 59 | - | 107 | - | 104 | - |
| Anterior caudal centum MPA D-111 | 71 | 86 | 104 | 85 | 106 | - |
| Middle caudal centrum MPA D-112 | 71 | 76 | 85 | 78 | 94 | - |
| Middle caudal centrum MPA D-115 | 66 | 69 | 94 | 73 | 93 | - |
| Middle caudal centrum MPA D-113 | 61 | 73 | 91 | 67 | 89 | - |
| Caudal neural arch MPA D-108 | - | - | - | - | - | 68 * |
| Caudal neural arch MPA D-109 | - | - | - | - | - | 124 |
| CL | AAFH | AAFW | PAFH | PAFW | NAH | |
| Middle caudal centrum MPA D-653 | 66 | 72 | 94 | 73 | 96 | - |
| Middle caudal centrum MPA D-1086 | 65 | 70 | 82 | 68 | 84 | - |
| Barrihonda-El Humero site | CL | AAFH | AAFW | PAFH | PAFW | NAH |
| Anterior caudal vertebra MAP-4682 | 49 | 82 | 121 | 79 | 115 | 170 |
| Anterior caudal vertebra MAP-4680 | 47 | 65 | 108 | 84 | 112 | 98 * |
| Middle caudal vertebra MAP-4683 | 53 | 79 | 105 | 85 | 103 | - |
| Middle caudal vertebra MAP-4681 | 53 | 79 | 91 | 88 | 105 | 147 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Fenollosa, S.; Suñer, M.; Cobos, A. New Fossils of Stegosaurs from the Upper Jurassic of the Eastern Iberian Peninsula (Spain). Diversity 2022, 14, 1047. https://doi.org/10.3390/d14121047
Sánchez-Fenollosa S, Suñer M, Cobos A. New Fossils of Stegosaurs from the Upper Jurassic of the Eastern Iberian Peninsula (Spain). Diversity. 2022; 14(12):1047. https://doi.org/10.3390/d14121047
Chicago/Turabian StyleSánchez-Fenollosa, Sergio, Maite Suñer, and Alberto Cobos. 2022. "New Fossils of Stegosaurs from the Upper Jurassic of the Eastern Iberian Peninsula (Spain)" Diversity 14, no. 12: 1047. https://doi.org/10.3390/d14121047
APA StyleSánchez-Fenollosa, S., Suñer, M., & Cobos, A. (2022). New Fossils of Stegosaurs from the Upper Jurassic of the Eastern Iberian Peninsula (Spain). Diversity, 14(12), 1047. https://doi.org/10.3390/d14121047