

Climate Change Is Driving Shifts in Dragonfly Species Richness across Europe via Differential Dynamics of Taxonomic and Biogeographic Groups



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data
2.2.1. Species Distribution Data
2.2.2. Species Classification
2.3. Data Analysis
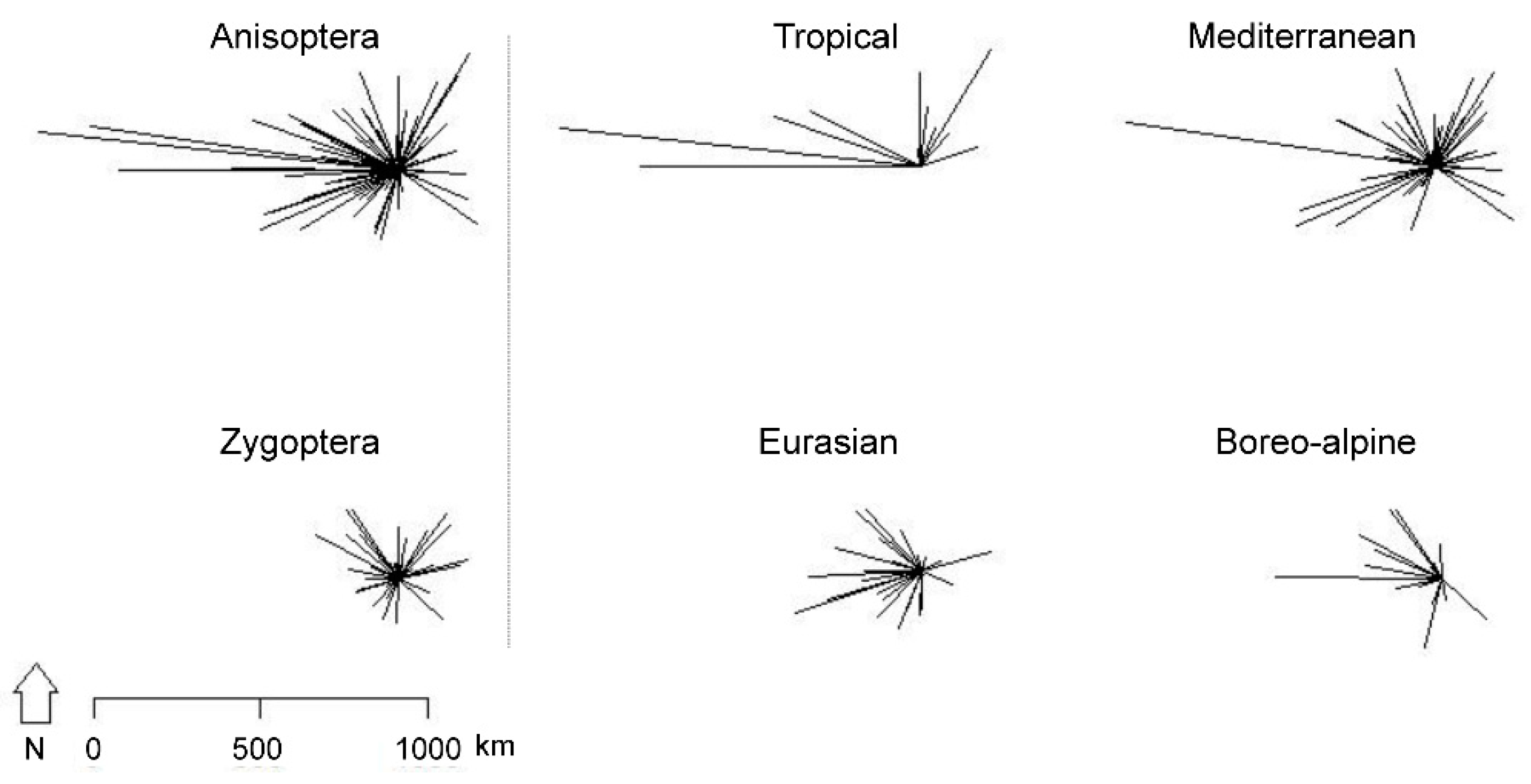
2.3.1. Range Shifts
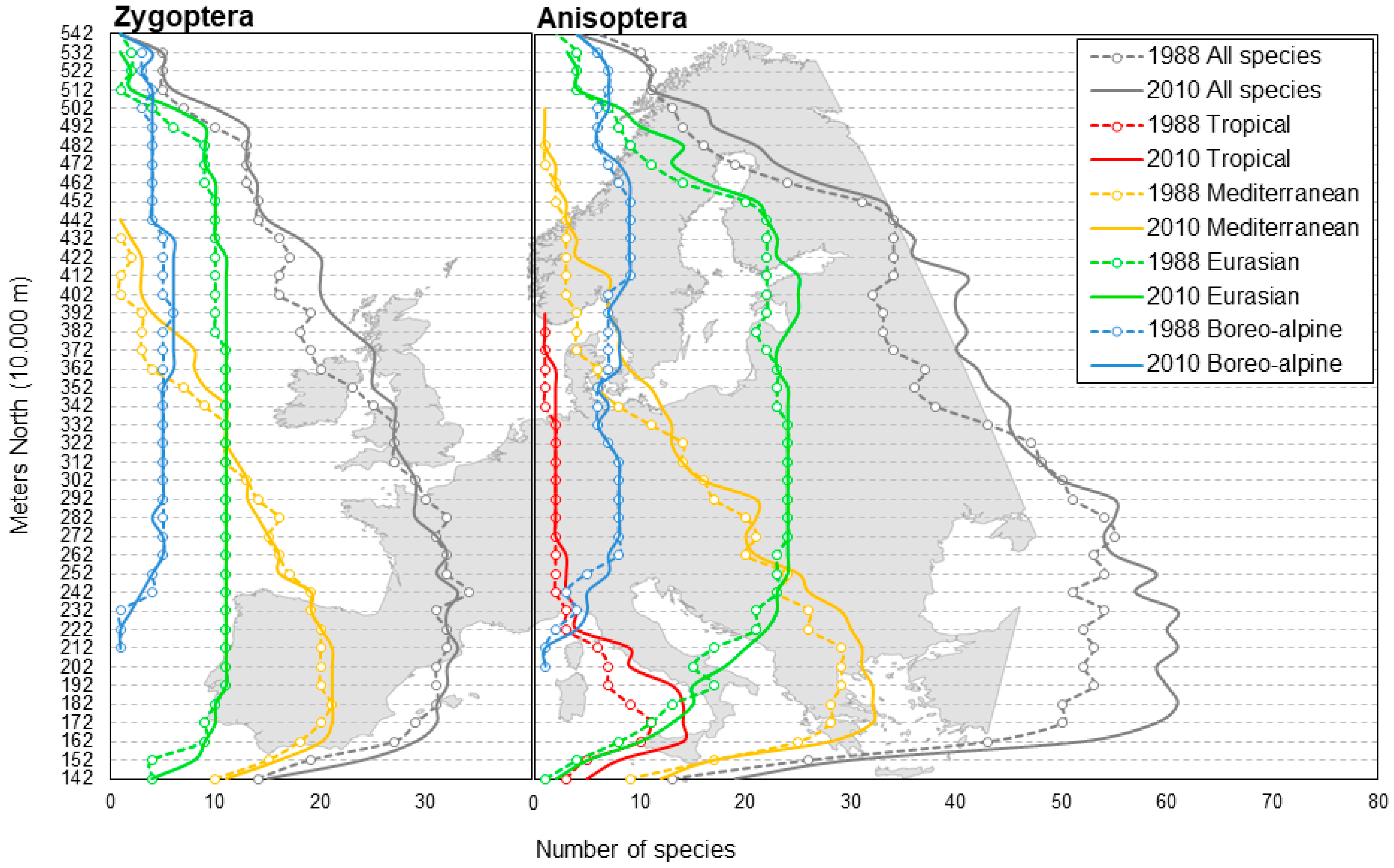
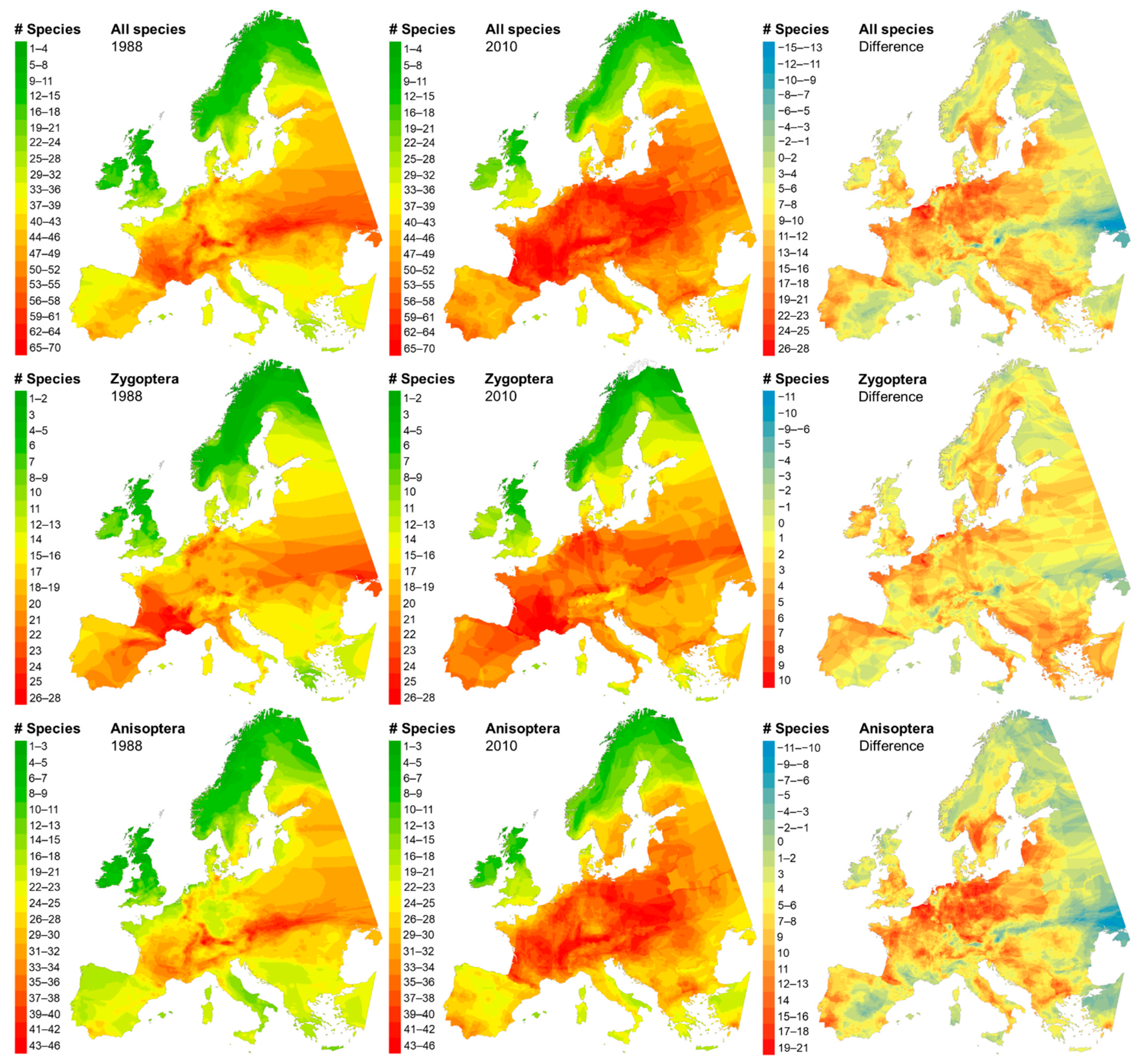
2.3.2. Species Richness
2.4. Statistical Analysis
3. Results
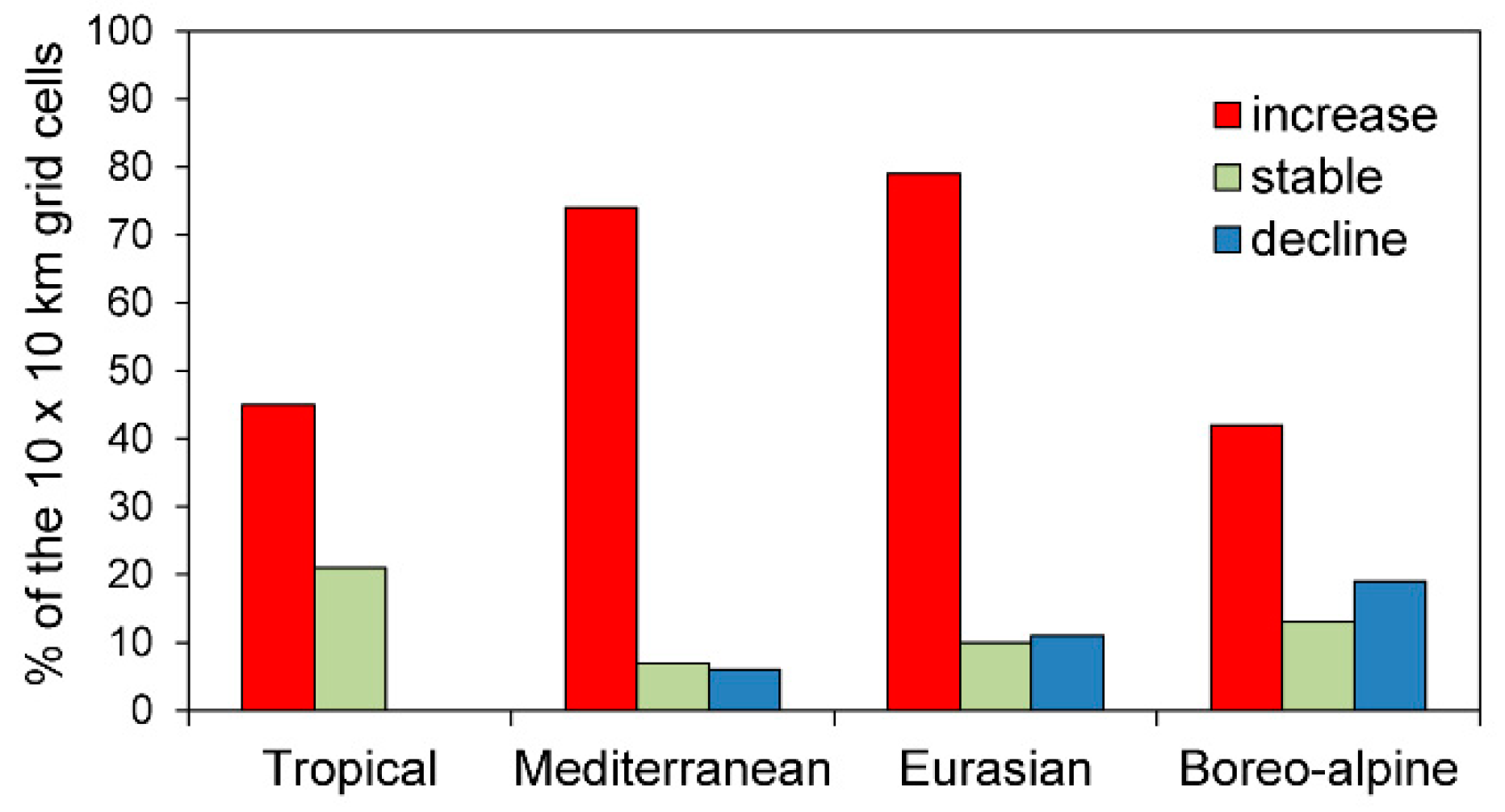
3.1. Range Shift Pattern
3.2. Species Richness Pattern
4. Discussion
4.1. Data Quality
4.2. Are Increases in Ranges of True Dragonflies Greater Than Those of Damselflies?
4.3. Have the Ranges of Southern Species Increased More Than the Ranges of Continental and Northern Species?
4.4. Will Northern Species and High-Elevation Species Experience Reduced Overall Ranges as Their Realized Climatic Envelopes Shrink Because of Global Warming?
4.5. Are Northwards Range Shifts Greater Than Movements at Other Range Margins Reflecting a Directional Poleward Shift Rather Than Non-Directional Range Expansion?
4.6. Are Geographical Shifts in Species Richness Patterns Driven Mainly by Southern Species Rather Than by Species from a More Continental and Northern Origin?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Suborder | Species | Biogeographic Group | Range Shift Trend |
|---|---|---|---|
| Zygoptera | Calopteryx haemorrhoidalis Vander Linden, 1825 | Mediterranean | Expansion |
| Zygoptera | Calopteryx splendens Harris, 1780 | Eurasian | Expansion |
| Zygoptera | Calopteryx virgo Linnaeus, 1758 | Eurasian | Expansion |
| Zygoptera | Calopteryx xanthostoma Charpentier, 1825 | Mediterranean | Contraction |
| Zygoptera | Ceriagrion georgifreyi Schmidt, 1953 | Mediterranean | Expansion |
| Zygoptera | Ceriagrion tenellum De Villers, 1789 | Mediterranean | Expansion |
| Zygoptera | Chalcolestes viridis Vander Linden, 1825 | Mediterranean | Expansion |
| Zygoptera | Coenagrion armatum Charpentier, 1840 | Boreo-alpine | Expansion |
| Zygoptera | Coenagrion caerulescens Fonscolombe, 1838 | Mediterranean | Expansion |
| Zygoptera | Coenagrion hastulatum Charpentier, 1825 | Boreo-alpine | Expansion |
| Zygoptera | Coenagrion hylas Trybom, 1889 | Boreo-alpine | Stable |
| Zygoptera | Coenagrion intermedium Lohmann, 1990 | Mediterranean | Stable |
| Zygoptera | Coenagrion johanssoni Wallengren, 1894 | Boreo-alpine | Expansion |
| Zygoptera | Coenagrion lunulatum Charpentier, 1840 | Boreo-alpine | Contraction |
| Zygoptera | Coenagrion mercuriale Charpentier, 1840 | Mediterranean | Expansion |
| Zygoptera | Coenagrion ornatum Selys, 1850 | Mediterranean | Expansion |
| Zygoptera | Coenagrion puella Linnaeus, 1758 | Mediterranean | Expansion |
| Zygoptera | Coenagrion pulchellum Vander Linden, 1825 | Eurasian | Expansion |
| Zygoptera | Coenagrion scitulum Rambur, 1842 | Mediterranean | Expansion |
| Zygoptera | Enallagma cyathigerum Charpentier, 1840 | Eurasian | Expansion |
| Zygoptera | Epallage fatime Charpentier, 1840 | Mediterranean | Expansion |
| Zygoptera | Erythromma lindenii Selys, 1840 | Mediterranean | Expansion |
| Zygoptera | Erythromma najas Hansemann, 1823 | Eurasian | Expansion |
| Zygoptera | Erythromma viridulum Charpentier, 1840 | Mediterranean | Expansion |
| Zygoptera | Ischnura elegans Vander Linden, 1820 | Eurasian | Expansion |
| Zygoptera | Ischnura fountaineae * Morton, 1905 | Mediterranean | Expansion |
| Zygoptera | Ischnura genei Rambur, 1842 | Mediterranean | Stable |
| Zygoptera | Ischnura graellsii Rambur, 1842 | Mediterranean | Expansion |
| Zygoptera | Ischnura pumilio Charpentier, 1825 | Mediterranean | Expansion |
| Zygoptera | Lestes barbarus Fabricius, 1798 | Mediterranean | Expansion |
| Zygoptera | Lestes dryas Kirby, 1890 | Eurasian | Expansion |
| Zygoptera | Lestes macrostigma Eversmann, 1836 | Mediterranean | Contraction |
| Zygoptera | Lestes sponsa Hansemann, 1823 | Eurasian | Expansion |
| Zygoptera | Lestes virens Charpentier, 1825 | Eurasian | Expansion |
| Zygoptera | Nehalennia speciosa Charpentier, 1840 | Boreo-alpine | Contraction |
| Zygoptera | Platycnemis acutipennis Selys, 1841 | Mediterranean | Expansion |
| Zygoptera | Platycnemis latipes Rambur, 1842 | Mediterranean | Contraction |
| Zygoptera | Platycnemis pennipes Pallas, 1771 | Eurasian | Contraction |
| Zygoptera | Pyrrhosoma nymphula Sulzer, 1776 | Eurasian | Expansion |
| Zygoptera | Sympecma fusca Vander Linden, 1820 | Mediterranean | Contraction |
| Zygoptera | Sympecma paedisca Brauer, 1877 | Boreo-alpine | Expansion |
| Anisoptera | Aeshna affinis Vander Linden, 1820 | Mediterranean | Expansion |
| Anisoptera | Aeshna caerulea Ström, 1783 | Boreo-alpine | Expansion |
| Anisoptera | Aeshna crenata Hagen, 1856 | Boreo-alpine | Expansion |
| Anisoptera | Aeshna cyanea Müller, 1764 | Eurasian | Contraction |
| Anisoptera | Aeshna grandis Linnaeus, 1758 | Eurasian | Expansion |
| Anisoptera | Aeshna isoceles (Müller, 1767) | Mediterranean | Expansion |
| Anisoptera | Aeshna juncea Linnaeus, 1758 | Boreo-alpine | Expansion |
| Anisoptera | Aeshna mixta Latreille, 1805 | Mediterranean | Expansion |
| Anisoptera | Aeshna serrata Hagen, 1856 | Eurasian | Expansion |
| Anisoptera | Aeshna subarctica Walker, 1908 | Boreo-alpine | Expansion |
| Anisoptera | Aeshna viridis Eversmann, 1836 | Eurasian | Expansion |
| Anisoptera | Anax imperator Leach, 1815 | Mediterranean | Expansion |
| Anisoptera | Anax parthenope Selys, 1839 | Mediterranean | Expansion |
| Anisoptera | Boyeria cretensis Peters, 1991 | Mediterranean | Expansion |
| Anisoptera | Boyeria irene Fonscolombe, 1838 | Mediterranean | Expansion |
| Anisoptera | Brachythemis impartita Karsch, 1890 | Tropical | Expansion |
| Anisoptera | Brachytron pratense Müller, 1764 | Eurasian | Expansion |
| Anisoptera | Caliaeschna microstigma Schneider, 1845 | Mediterranean | Expansion |
| Anisoptera | Cordulegaster bidentata Selys, 1843 | Mediterranean | Expansion |
| Anisoptera | Cordulegaster boltonii Donovan, 1807 | Eurasian | Contraction |
| Anisoptera | Cordulegaster helladica Lohmann, 1993 | Mediterranean | Expansion |
| Anisoptera | Cordulegaster heros Theischinger, 1979 | Mediterranean | Expansion |
| Anisoptera | Cordulegaster insignis Schneider, 1845 | Mediterranean | Contraction |
| Anisoptera | Cordulegaster picta Selys, 1854 | Mediterranean | Expansion |
| Anisoptera | Cordulegaster trinacriae Waterston, 1976 | Mediterranean | Expansion |
| Anisoptera | Cordulia aenea Linnaeus, 1758 | Eurasian | Expansion |
| Anisoptera | Crocothemis erythraea Brullé, 1832 | Tropical | Expansion |
| Anisoptera | Diplacodes lefebvrii Rambur, 1842 | Tropical | Expansion |
| Anisoptera | Epitheca bimaculata Charpentier, 1825 | Eurasian | Expansion |
| Anisoptera | Gomphus flavipes Selys, 1837 | Eurasian | Expansion |
| Anisoptera | Gomphus graslinii Rambur, 1842 | Mediterranean | Expansion |
| Anisoptera | Gomphus pulchellus Selys, 1840 | Mediterranean | Expansion |
| Anisoptera | Gomphus schneiderii Selys, 1850 | Mediterranean | Expansion |
| Anisoptera | Gomphus simillimus Selys, 1840 | Mediterranean | Expansion |
| Anisoptera | Gomphus vulgatissimus Linnaeus, 1758 | Eurasian | Expansion |
| Anisoptera | Leucorrhinia albifrons Burmeister, 1839 | Boreo-alpine | Contraction |
| Anisoptera | Leucorrhinia caudalis Charpentier, 1840 | Boreo-alpine | Expansion |
| Anisoptera | Leucorrhinia dubia Vander Linden, 1825 | Eurasian | Expansion |
| Anisoptera | Leucorrhinia pectoralis Charpentier, 1825 | Eurasian | Expansion |
| Anisoptera | Leucorrhinia rubicunda Linnaeus, 1758 | Boreo-alpine | Expansion |
| Anisoptera | Libellula depressa Linnaeus, 1758 | Eurasian | Expansion |
| Anisoptera | Libellula fulva Müller, 1764 | Eurasian | Expansion |
| Anisoptera | Libellula quadrimaculata Linnaeus, 1758 | Eurasian | Expansion |
| Anisoptera | Lindenia tetraphylla Vander Linden, 1825 | Mediterranean | Contraction |
| Anisoptera | Macromia splendens Pictet, 1843 | Mediterranean | Expansion |
| Anisoptera | Onychogomphus costae Selys, 1885 | Mediterranean | Expansion |
| Anisoptera | Onychogomphus forcipatus Linnaeus, 1758 | Mediterranean | Expansion |
| Anisoptera | Onychogomphus uncatus Charpentier, 1840 | Mediterranean | Expansion |
| Anisoptera | Ophiogomphus cecilia Fourcroy, 1785 | Eurasian | Expansion |
| Anisoptera | Orthetrum albistylum Selys, 1848 | Mediterranean | Expansion |
| Anisoptera | Orthetrum brunneum Fonscolombe, 1837 | Mediterranean | Expansion |
| Anisoptera | Orthetrum cancellatum Linnaeus, 1758 | Mediterranean | Expansion |
| Anisoptera | Orthetrum chrysostigma Burmeister, 1839 | Tropical | Expansion |
| Anisoptera | Orthetrum coerulescens Fabricius, 1798 | Mediterranean | Expansion |
| Anisoptera | Orthetrum nitidinerve Selys, 1841 | Mediterranean | Expansion |
| Anisoptera | Orthetrum sabina * Drury, 1773 | Tropical | Expansion |
| Anisoptera | Orthetrum taeniolatum * Schneider, 1845 | Tropical | Expansion |
| Anisoptera | Orthetrum trinacria Selys, 1841 | Tropical | Expansion |
| Anisoptera | Oxygastra curtisii Dale, 1834 | Mediterranean | Contraction |
| Anisoptera | Pantala flavescens Fabricius, 1798 | Tropical | Expansion |
| Anisoptera | Paragomphus genei Selys, 1841 | Tropical | Expansion |
| Anisoptera | Selysiothemis nigra Vander Linden, 1825 | Mediterranean | Expansion |
| Anisoptera | Somatochlora alpestris Selys, 1840 | Boreo-alpine | Expansion |
| Anisoptera | Somatochlora arctica Zetterstedt, 1840 | Boreo-alpine | Expansion |
| Anisoptera | Somatochlora flavomaculata Vander Linden, 1825 | Eurasian | Expansion |
| Anisoptera | Somatochlora meridionalis Nielsen, 1935 | Mediterranean | Expansion |
| Anisoptera | Somatochlora metallica Vander Linden, 1825 | Eurasian | Contraction |
| Anisoptera | Somatochlora sahlbergi Trybom, 1889 | Boreo-alpine | Expansion |
| Anisoptera | Sympetrum danae Sulzer, 1776 | Eurasian | Expansion |
| Anisoptera | Sympetrum depressiusculum Selys, 1841 | Eurasian | Expansion |
| Anisoptera | Sympetrum flaveolum Linnaeus, 1758 | Eurasian | Expansion |
| Anisoptera | Sympetrum fonscolombii Selys, 1840 | Tropical | Expansion |
| Anisoptera | Sympetrum meridionale Selys, 1841 | Mediterranean | Expansion |
| Anisoptera | Sympetrum pedemontanum O.F.Müller, 1766 | Eurasian | Expansion |
| Anisoptera | Sympetrum sanguineum Müller, 1764 | Eurasian | Expansion |
| Anisoptera | Sympetrum sinaiticum Dumont, 1977 | Mediterranean | Expansion |
| Anisoptera | Sympetrum striolatum Charpentier, 1840 | Eurasian | Expansion |
| Anisoptera | Sympetrum vulgatum Linnaeus, 1758 | Eurasian | Expansion |
| Anisoptera | Trithemis annulate Palisot de Beauvois, 1807 | Tropical | Expansion |
| Anisoptera | Trithemis festiva Rambur, 1842 | Tropical | Expansion |
| Anisoptera | Trithemis kirbyi * Selys, 1891 | Tropical | Expansion |
| Anisoptera | Zygonyx torridus Kirby, 1889 | Tropical | Expansion |
References
- Brown, J.; Lomolino, M. Biogeography; Sinauer Associates: Sunderland, MA, USA, 1998. [Google Scholar]
- Lenoir, J.; Svenning, J.C. Climate-Related Range Shifts—A Global Multidimensional Synthesis and New Research Directions. Ecography 2015, 38, 15–28. [Google Scholar] [CrossRef]
- Thomas, C.D.; Bodsworth, E.J.; Wilson, R.J.; Simmons, A.D.; Davies, Z.G.; Musche, M.; Conradt, L. Ecological and Evolutionary Processes at Expanding Range Margins. Nature 2001, 411, 577–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lancaster, L.T.; Dudaniec, R.Y.; Hansson, B.; Svensson, E.I. Latitudinal Shift in Thermal Niche Breadth Results from Thermal Release during a Climate-Mediated Range Expansion. J. Biogeogr. 2015, 42, 1953–1963. [Google Scholar] [CrossRef]
- Channell, R.; Lomollno, M.V. Dynamic Biogeography and Conservation of Endangered Species. Nature 2000, 403, 84–86. [Google Scholar] [CrossRef] [PubMed]
- Fahrig, L. Effects of Habitat Fragmentation on Biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 59–71. [Google Scholar] [CrossRef] [Green Version]
- Wiens, J.J. Climate-Related Local Extinctions Are Already Widespread among Plant and Animal Species. PLoS Biol. 2016, 14, e2001104. [Google Scholar] [CrossRef] [Green Version]
- Opdam, P.; Wascher, D. Climate Change Meets Habitat Fragmentation: Linking Landscape and Biogeographical Scale Levels in Research and Conservation. Biol. Conserv. 2004, 117, 285–297. [Google Scholar] [CrossRef]
- Chaudhary, C.; Richardson, A.J.; Schoeman, D.S.; Costello, M.J. Global Warming Is Causing a More Pronounced Dip in Marine Species Richness around the Equator. Proc. Natl. Acad. Sci. USA 2021, 118, e2015094118. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; de Siqueira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction Risk from Climate Change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef] [Green Version]
- Flenner, I.; Sahlén, G. Dragonfly Community Re-Organisation in Boreal Forest Lakes: Rapid Species Turnover Driven by Climate Change? Insect Conserv. Divers. 2008, 1, 169–179. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2007: The Physical Science Basis: Contribution of Working Group I to the Fourth Assessment; Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2007. [Google Scholar]
- Gienapp, P.; Teplitsky, C.; Alho, J.S.; Mills, J.A.; Merilä, J. Climate Change and Evolution: Disentangling Environmental and Genetic Responses. Mol. Ecol. 2008, 17, 167–178. [Google Scholar] [CrossRef]
- Davis, M.B.; Shaw, R.G. Range Shifts and Adaptive Responses to Quaternary Climate Change. Science 2001, 292, 673–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, H.; Stoks, R.; Nilsson-Örtman, V.; Ingvarsson, P.K.; Johansson, F. Large-Scale Patterns in Genetic Variation, Gene Flow and Differentiation in Five Species of European Coenagrionid Damselfly Provide Mixed Support for the Central-Marginal Hypothesis. Ecography 2013, 36, 744–755. [Google Scholar] [CrossRef]
- Kellermann, V.; Loeschcke, V.; Hoffmann, A.A.; Kristensen, T.N.; Fløjgaard, C.; David, J.R.; Svenning, J.C.; Overgaard, J. Phylogenetic Constraints in Key Functional Traits Behind Species’ Climate Niches: Patterns of Desiccation and Cold Resistance across 95 Drosophila Species. Evolution 2012, 66, 3377–3389. [Google Scholar] [CrossRef] [PubMed]
- Bota-Sierra, C.A.; García-Robledo, C.; Escobar, F.; Novelo-Gutiérrez, R.; Londoño, G.A. Environment, Taxonomy and Morphology Constrain Insect Thermal Physiology along Tropical Mountains. Funct. Ecol. 2022, 36, 1924–1935. [Google Scholar] [CrossRef]
- Hickling, R.; Roy, D.B.; Hill, J.K.; Fox, R.; Thomas, C.D. The Distributions of a Wide Range of Taxonomic Groups Are Expanding Polewards. Glob. Chang. Biol. 2006, 12, 450–455. [Google Scholar] [CrossRef]
- Hickling, R.; Roy, D.B.; Hill, J.K.; Thomas, C.D. A Northward Shift of Range Margins in British Odonata. Glob. Chang. Biol. 2005, 11, 502–506. [Google Scholar] [CrossRef]
- de Knijf, G.; Anselin, A. When South Goes North: Mediterranean Dragonflies (Odonata) Conquer Flanders (North-Belgium). BioRisk 2010, 5, 141–153. [Google Scholar] [CrossRef] [Green Version]
- Ott, J. Dragonflies and Climatic Change—Recent Trends in Germany and Europe. BioRisk 2010, 5, 253–286. [Google Scholar] [CrossRef]
- Olsen, K.; Svenning, J.-C.; Balslev, H. Niche Breadth Predicts Geographical Range Size and Northern Range Shift in European Dragonfly Species (Odonata). Diversity 2022, 14, 719. [Google Scholar] [CrossRef]
- Flenner, I.; Richter, O.; Suhling, F. Rising Temperature and Development in Dragonfly Populations at Different Latitudes. Freshw. Biol. 2010, 55, 397–410. [Google Scholar] [CrossRef]
- Johansson, F. Latitudinal Shifts in Body Size of Enallagma Cyathigerum (Odonata). J. Biogeogr. 2003, 30, 29–34. [Google Scholar] [CrossRef]
- Dingemanse, N.J.; Kalkman, V.J. Changing Temperature Regimes Have Advanced the Phenology of Odonata in the Netherlands. Ecol. Entomol. 2008, 33, 394–402. [Google Scholar] [CrossRef]
- Braune, E.; Richter, O.; Söndgerath, D.; Suhling, F. Voltinism Flexibility of a Riverine Dragonfly along Thermal Gradients. Glob. Chang. Biol. 2008, 14, 470–482. [Google Scholar] [CrossRef]
- Hassall, C.; Thompson, D.J.; French, G.C.; Harvey, I.F. Historical Changes in the Phenology of British Odonata Are Related to Climate. Glob. Chang. Biol. 2007, 13, 933–941. [Google Scholar] [CrossRef]
- Richter, O.; Suhling, F.; Müller, O.; Kern, D. A Model for Predicting the Emergence of Dragonflies in a Changing Climate. Freshw. Biol. 2008, 53, 1868–1880. [Google Scholar] [CrossRef]
- Johansson, F.; Śniegula, S.; Brodin, T. Emergence Patterns and Latitudinal Adaptations in Development Time of Odonata in North Sweden and Poland. Odonatologica 2010, 39, 97–106. [Google Scholar]
- Stoks, R.; de Block, M.; McPeek, M.A. Physiological Costs of Compensatory Growth in a Damselfly. Ecology 2006, 87, 1566–1574. [Google Scholar] [CrossRef]
- de Block, M.; Stoks, R. Compensatory Growth and Oxidative Stress in a Damselfly. Proc. R. Soc. B Biol. Sci. 2008, 275, 781–785. [Google Scholar] [CrossRef] [Green Version]
- Sternberg, K. Colours, Colour Change, Colour Patterns and ‘Cuticular Windows’ as Light Traps. Their Thermoregulatoric and Ecological Significance in Some Aeshna Species (Odonata: Aeshnidae). Zool. Anz. 1996, 235, 77–88. [Google Scholar]
- Corbet, P.S. Dragonflies: Behavior and Ecology of Odonata; Harley Books: Colchester, UK, 2004. [Google Scholar]
- Kalkman, V.J.; Boudot, J.-P.; Bernard, R.; Conze, K.-J.; de Knijf, G.; Dyatlova, E.; Ferreira, S.; Jović, M.; Ott, J.; Riservato, E.; et al. European Red List of Dragonflies; Publications Office of the European Union: Luxembourg, 2010. [Google Scholar]
- Clausnitzer, V.; Kalkman, V.J.; Ram, M.; Collen, B.; Baillie, J.E.M.; Bedjanič, M.; Darwall, W.R.T.; Dijkstra, K.D.B.; Dow, R.; Hawking, J.; et al. Odonata Enter the Biodiversity Crisis Debate: The First Global Assessment of an Insect Group. Biol. Conserv. 2009, 142, 1864–1869. [Google Scholar] [CrossRef]
- Graham, C.H.; Hijmans, R.J. A Comparison of Methods for Mapping Species Ranges and Species Richness. Glob. Ecol. Biogeogr. 2006, 15, 578–587. [Google Scholar] [CrossRef]
- Suhling, F.; Sahlén, G.; Gorb, S.; Kalkman, V.; Dijkstra, K.-D.; van Tol, J. Order Odonata. In Ecology and General Biology: Thorp and Covich’s Freshwater Invertebrates; Thorp, J., Rogers, D., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 893–932. [Google Scholar]
- Kalkman, V.J.; Clausnitzer, V.; Dijkstra, K.-D.B.; Orr, A.G.; Paulson, D.R.; van Tol, J. Global Diversity of Dragonflies (Odonata) in Freshwater. In Freshwater Animal Diversity Assessment; Balian, E., Lévêque, C., Segers, H., Martens, K., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 351–363. [Google Scholar]
- de Lattin, G. Grundriss Der Zoogeographie; Fischer: Stuttgart, Germany, 1967. [Google Scholar]
- Husemann, M.; Schmitt, T.; Zachos, F.E.; Ulrich, W.; Habel, J.C. Palaearctic Biogeography Revisited: Evidence for the Existence of a North African Refugium for Western Palaearctic Biota. J. Biogeogr. 2014, 41, 81–94. [Google Scholar] [CrossRef]
- Simonsen, T.J.; Huemer, P. Phylogeography of Hepialus Humuli (L.) (Lepidoptera: Hepialidae) in Europe: Short Distance vs. Large Scale Postglacial Expansions from Multiple Alpine Refugia and Taxonomic Implications. Insect Syst. Evol. 2014, 45, 209–250. [Google Scholar] [CrossRef]
- Ursenbacher, S.; Carlsson, M.; Helfer, V.; Tegelström, H.; Fumagalli, L. Phylogeography and Pleistocene Refugia of the Adder (Vipera Berus) as Inferred from Mitochondrial DNA Sequence Data. Mol. Ecol. 2006, 15, 3425–3437. [Google Scholar] [CrossRef]
- Brochmann, C.; Gabrielsen, T.M.; Nordal, I.; Landvik, J.Y.; Elven, R. Glacial Survival or Tabula Rasa? The History of North Atlantic Biota Revisited. Taxon 2003, 52, 417–450. [Google Scholar] [CrossRef]
- Schmitt, T.; Rákosy, L.; Abadjiev, S.; Müller, P. Multiple Differentiation Centres of a Non-Mediterranean Butterfly Species in South-Eastern Europe. J. Biogeogr. 2007, 34, 939–950. [Google Scholar] [CrossRef]
- Baselga, A.; Lobo, J.M.; Svenning, J.C.; Aragón, P.; Araújo, M.B. Dispersal Ability Modulates the Strength of the Latitudinal Richness Gradient in European Beetles. Glob. Ecol. Biogeogr. 2012, 21, 1106–1113. [Google Scholar] [CrossRef]
- Castillo-Pérez, E.U.; Suárez-Tovar, C.M.; González-Tokman, D.; Schondube, J.E.; Córdoba-Aguilar, A. Insect Thermal Limits in Warm and Perturbed Habitats: Dragonflies and Damselflies as Study Cases. J. Therm. Biol. 2022, 103, 103164. [Google Scholar] [CrossRef]
- Dumont, H.J.; Desmet, K. Trans-Sahara and Trans-Mediterranean Migratory Activity of Hemianax Ephippiger (Burmeister) in 1988 and 1989 (Anisoptera: Aeshnidae). Odonatologica 1990, 19, 181–185. [Google Scholar]
- Hassall, C.; Thompson, D.J. The Effects of Environmental Warming on Odonata: A Review. Int. J. Odonatol. 2008, 11, 131–153. [Google Scholar] [CrossRef]
- Askew, R.R. The Dragonflies of Europe; Harley Books (BH & A Harley Ltd.): Colchester, UK, 1988. [Google Scholar]
- Dijkstra, K.-D.; Lewington, R. Field Guide to the Dragonflies of Britain and Europe; British Wildlife Publishing: Dorset, UK, 2006. [Google Scholar]
- ESRI. ArcGIS Desktop [GIS Software], version 10.2; Environmental Systems Research Institute: Redlands, CA, USA, 2010. [Google Scholar]
- Sternberg, K. Die Postglaziale Besiedlung Mitteleuropas Durch Libellen, Mit Besonderer Berucksichtigung Sudwestdeutschlands (Insecta, Odonata). The Postglacial Colonization of Central Europe by Dragonflies, with Special Reference to Southwestern Germany (Insecta, Odonata). J. Biogeogr. 1998, 25, 319–337. [Google Scholar] [CrossRef]
- Beschovski, V.; Marinov, M. Fauna, Ecology, and Zoogeography of Dragonflies (Insecta: Odonata) of Bulgaria. In Biogeography and Ecology of Bulgaria; Fet, V., Popov, A., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 199–231. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2022. [Google Scholar]
- Lemon, J. Plotrix: A Package in the Red Light District of R. R-News 2006, 6, 8–12. [Google Scholar]
- Pulliam, H.R. On the Relationship between Niche and Distribution. Ecol. Lett. 2000, 3, 349–361. [Google Scholar] [CrossRef]
- Rondinini, C.; Wilson, K.A.; Boitani, L.; Grantham, H.; Possingham, H.P. Tradeoffs of Different Types of Species Occurrence Data for Use in Systematic Conservation Planning. Ecol. Lett. 2006, 9, 1136–1145. [Google Scholar] [CrossRef]
- Gaston, K.J. The Structure and Dynamics of Geographic Ranges; Oxford University Press: Oxford, UK, 2003. [Google Scholar]
- Jetz, W.; Sekercioglu, C.H.; Watson, J.E.M. Ecological Correlates and Conservation Implications of Overestimating Species Geographic Ranges. Conserv. Biol. 2008, 22, 110–119. [Google Scholar] [CrossRef]
- Hurlbert, A.H.; White, E.P. Disparity between Range Map- and Survey-Based Analyses of Species Richness: Patterns, Processes and Implications. Ecol. Lett. 2005, 8, 319–327. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Field, R.; Cornell, H.V.; Currie, D.J.; Guégan, J.F.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T.; O’Brien, E.M.; et al. Energy, Water, and Broad-Scale Geographic Patterns of Species Richness. Ecology 2003, 84, 3105–3117. [Google Scholar] [CrossRef] [Green Version]
- Grewe, Y.; Hof, C.; Dehling, D.M.; Brandl, R.; Brändle, M. Recent Range Shifts of European Dragonflies Provide Support for an Inverse Relationship between Habitat Predictability and Dispersal. Glob. Ecol. Biogeogr. 2013, 22, 403–409. [Google Scholar] [CrossRef]
- Hof, C.; Brändle, M.; Dehling, D.M.; Munguía, M.; Brandl, R.; Araújo, M.B.; Rahbek, C. Habitat Stability Affects Dispersal and the Ability to Track Climate Change. Biol. Lett. 2012, 8, 639–643. [Google Scholar] [CrossRef] [Green Version]
- Hof, C.; Brändle, M.; Brandl, R. Lentic Odonates Have Larger and More Northern Ranges than Lotic Species. J. Biogeogr. 2006, 33, 63–77. [Google Scholar] [CrossRef]
- Kalkman, V.J.; Boudot, J.-P.; Futahashi, R.; Abbott, J.C.; Bota-Sierra, C.A.; Guralnick, R.; Bybee, S.M.; Ware, J.; Belitz, M.W. Diversity of Palaearctic Dragonflies and Damselflies (Odonata). Diversity 2022, 14, 966. [Google Scholar] [CrossRef]
- Hurlbert, A.H.; Jetz, W. Species Richness, Hotspots, and the Scale Dependence of Range Maps in Ecology and Conservation. Proc. Natl. Acad. Sci. USA 2007, 104, 13384–13389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boudot, J.-P.; Kalkman, V.J. Atlas of the European Dragonflies and Damselflies; KNNV-Publishing: Utrecht, The Netherlands, 2015. [Google Scholar]
- Riservato, E.; Boudot, J.; Ferreira, S.; Jović, M.; Kalkman, V.; Schneider, W.; Samraoui, B.; Cuttelod, A. The Status and Distribution of Dragonflies of the Mediterranean Basin; IUCN: Gland, Switzerland; Malaga, Spain, 2009. [Google Scholar]
- Termaat, T.; Kalkman, V.; Bouwman, J. Changes in the Range of Dragonflies in the Netherlands and the Possible Role of Temperature Change. BioRisk 2010, 5, 155–173. [Google Scholar] [CrossRef]
- Hill, J.K.; Thomas, C.D.; Huntley, B. Climate and Habitat Availability Determine 20th Century Changes in a Butterfly’s Range Margin. Proc. R. Soc. B Biol. Sci. 1999, 266, 1197–1206. [Google Scholar] [CrossRef] [Green Version]
- Boudot, J.-P.; Kalkman, V.J.; Azpilicueta Amorin, M.; Bogdanović, T.; Cordero Rivera, A.; Degabriele, G.; Dommanget, J.-L.; Ferreira, S.; Garrigos, B.; Jović, M.; et al. Atlas of the Odonata of the Mediterranean and North Africa. Libellula Suppl. 2009, 9, 1–256. [Google Scholar]
- de Knijf, G.; Flenker, U.; Vanappelghem, C.; Manci, C.O.; Kalkman, V.J.; Demolder, H. The Status of Two Boreo-Alpine Species, Somatochlora Alpestris and S. Arctica, in Romania and Their Vulnerability to the Impact of Climate Change (Odonata: Corduliidae). Int. J. Odonatol. 2011, 14, 111–126. [Google Scholar] [CrossRef]
- Bernard, R.; Wildermuth, H. Nehalennia Speciosa (Charpentier, 1840) in Europe: A Case of a Vanishing Relict (Zygoptera: Coenagrionidae). Odonatologica 2005, 34, 335–378. [Google Scholar]
- Hampe, A.; Petit, R.J. Conserving Biodiversity under Climate Change: The Rear Edge Matters. Ecol. Lett. 2005, 8, 461–467. [Google Scholar] [CrossRef] [Green Version]
- Simonsen, T.J.; Olsen, K.; Djernæs, M. The African-Iberian Connection in Odonata: MtDNA and NcDNA Based Phylogeography of Aeshna Cyanea (MüLler, 1764) (Odonata: Aeshnidae) in Western Palaearctic. Arthropod Syst. Phylogeny 2020, 78, 309–320. [Google Scholar] [CrossRef]
- Boudot, J.-P.; Clausnitzer, V.; Samraoui, B.; Suhling, F.; Dijkstra, K.-D.B.; Schneider, W. Paragomphus genei. The IUCN Red List of Threatened Species 2016: E.T59976A83861834. Available online: https://www.iucnredlist.org/species/59976/83861834 (accessed on 26 August 2022).
- Jiménez-Alfaro, B.; Girardello, M.; Chytrý, M.; Svenning, J.-C.; Willner, W.; Gégout, J.-C.; Agrillo, E.; Campos, J.A.; Jandt, U.; Kącki, Z.; et al. History and Environment Shape Species Pools and Community Diversity in European Beech Forests. Nat. Ecol. Evol. 2018, 2, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Aguesse, P. Les Odonates de l’Europe Occidentale, Du Nord de l’Afrique et Des Îles Atlantiques. In Faune de l’Europe et du Bassin Méditerranéen; Masson et Cie: Paris, France, 1968; Volume 4, pp. 1–258. [Google Scholar]
- Pilgrim, E.M.; von Dohlen, C.D. Molecular and Morphological Study of Species-Level Questions within the Dragonfly Genus Sympetrum (Odonata: Libellulidae). Ann. Entomol. Soc. Am. 2007, 100, 688–702. [Google Scholar] [CrossRef]
- IUCN The IUCN Red List of Threatened Species. Version 2015-3. Available online: www.iucnredlist.org (accessed on 22 August 2016).





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olsen, K.; Svenning, J.-C.; Balslev, H. Climate Change Is Driving Shifts in Dragonfly Species Richness across Europe via Differential Dynamics of Taxonomic and Biogeographic Groups. Diversity 2022, 14, 1066. https://doi.org/10.3390/d14121066
Olsen K, Svenning J-C, Balslev H. Climate Change Is Driving Shifts in Dragonfly Species Richness across Europe via Differential Dynamics of Taxonomic and Biogeographic Groups. Diversity. 2022; 14(12):1066. https://doi.org/10.3390/d14121066
Chicago/Turabian StyleOlsen, Kent, Jens-Christian Svenning, and Henrik Balslev. 2022. "Climate Change Is Driving Shifts in Dragonfly Species Richness across Europe via Differential Dynamics of Taxonomic and Biogeographic Groups" Diversity 14, no. 12: 1066. https://doi.org/10.3390/d14121066
APA StyleOlsen, K., Svenning, J. -C., & Balslev, H. (2022). Climate Change Is Driving Shifts in Dragonfly Species Richness across Europe via Differential Dynamics of Taxonomic and Biogeographic Groups. Diversity, 14(12), 1066. https://doi.org/10.3390/d14121066