
Response of Phytoplankton Community Structure to Vegetation Restoration after Removal of Purse Seine in Shengjin Lake

Abstract
:1. Introduction
2. Materials and Methods
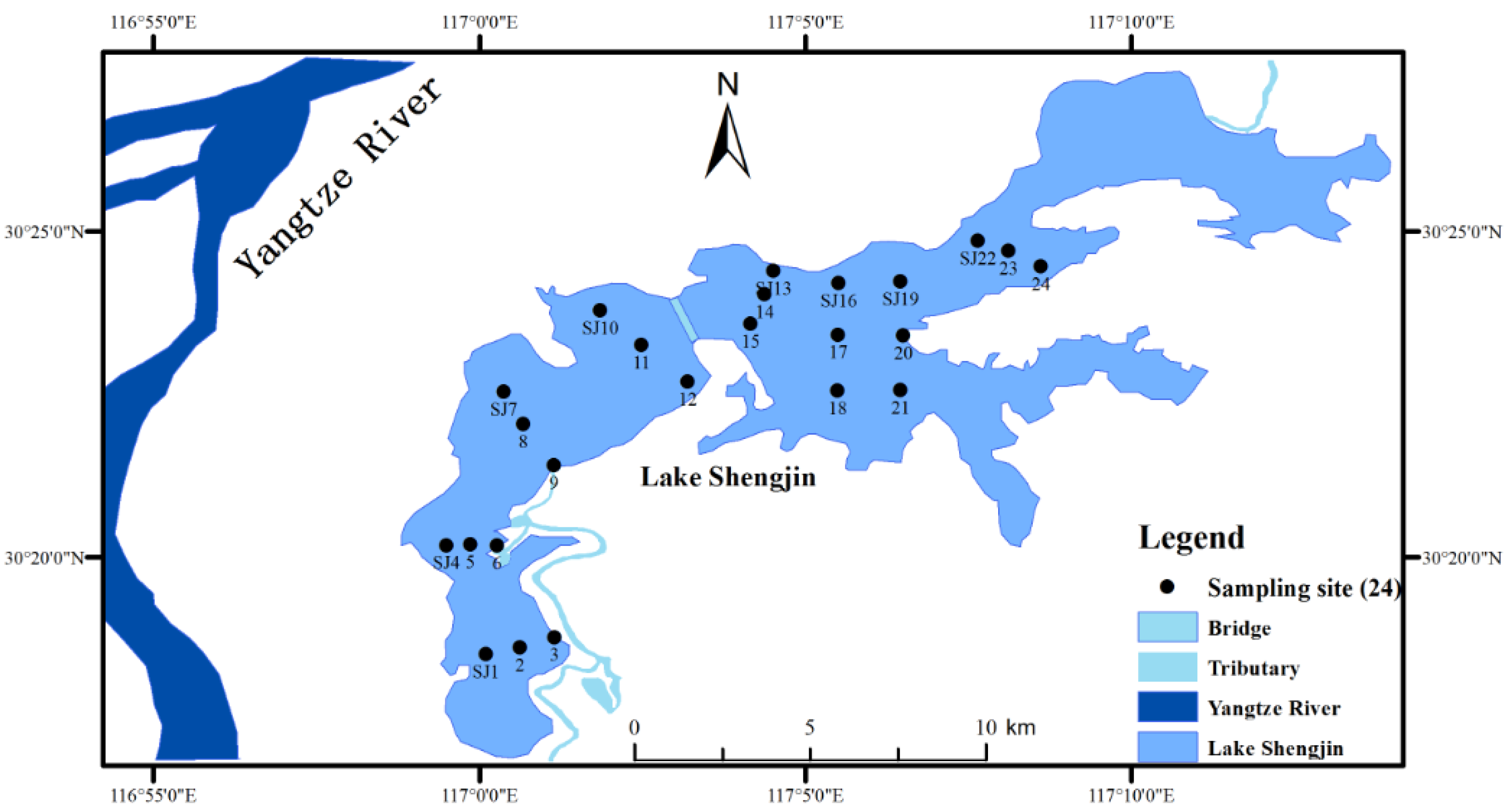
2.1. Description of Study Area
2.2. Data Collection
2.3. Data Analysis
3. Results
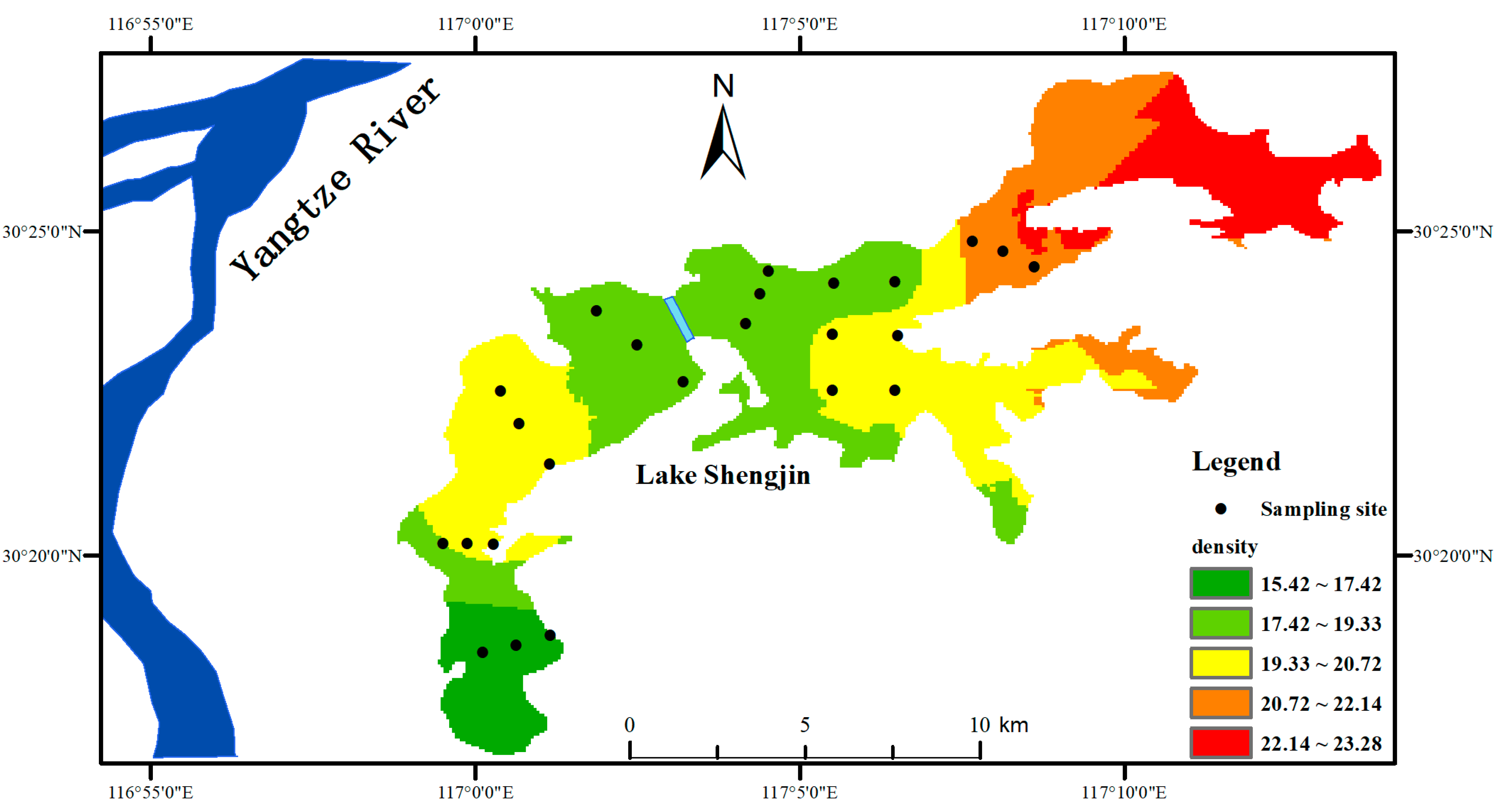
3.1. Environmental Characteristics Parameters
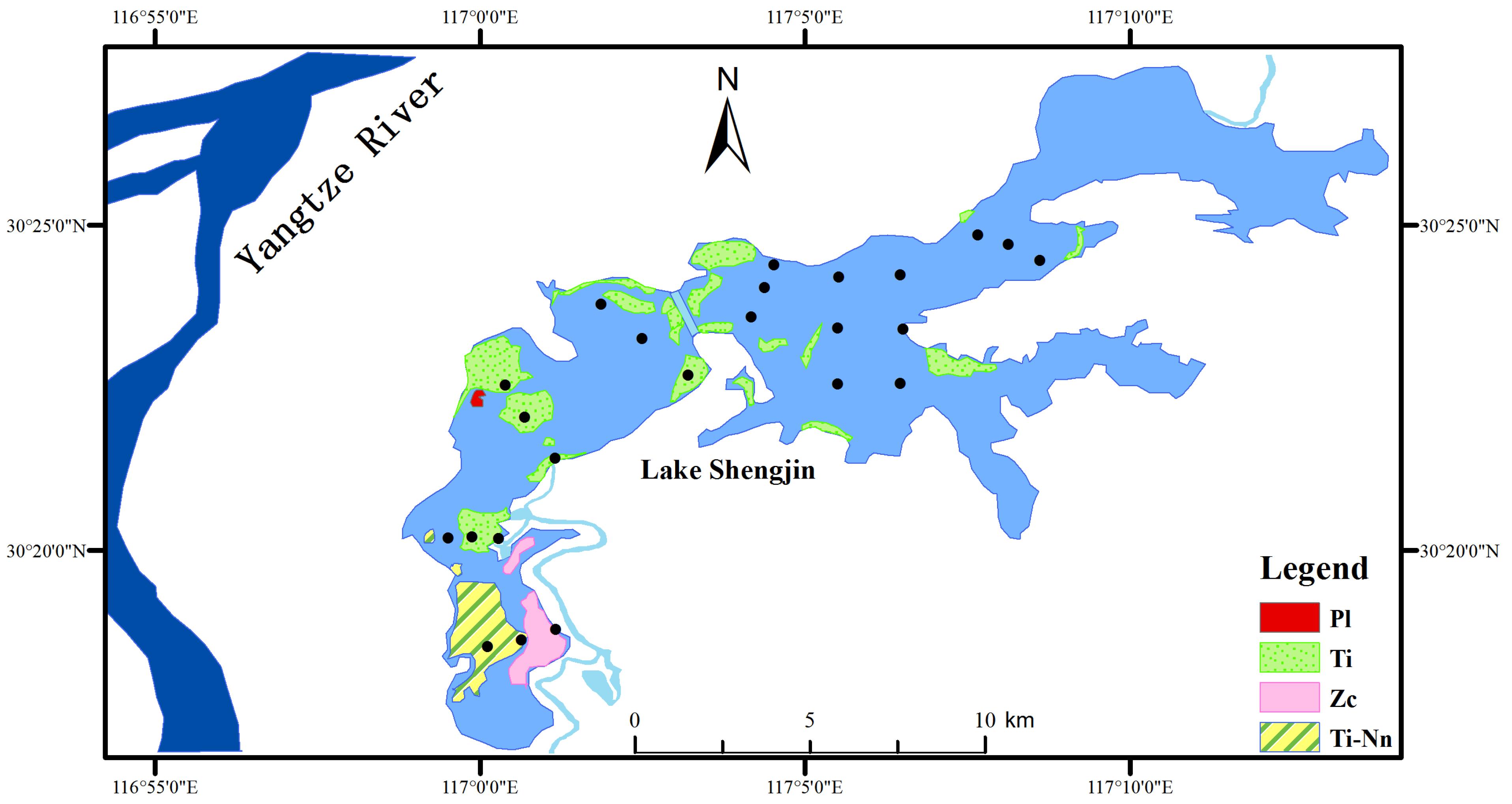
3.2. Distribution of Aquatic Vegetation
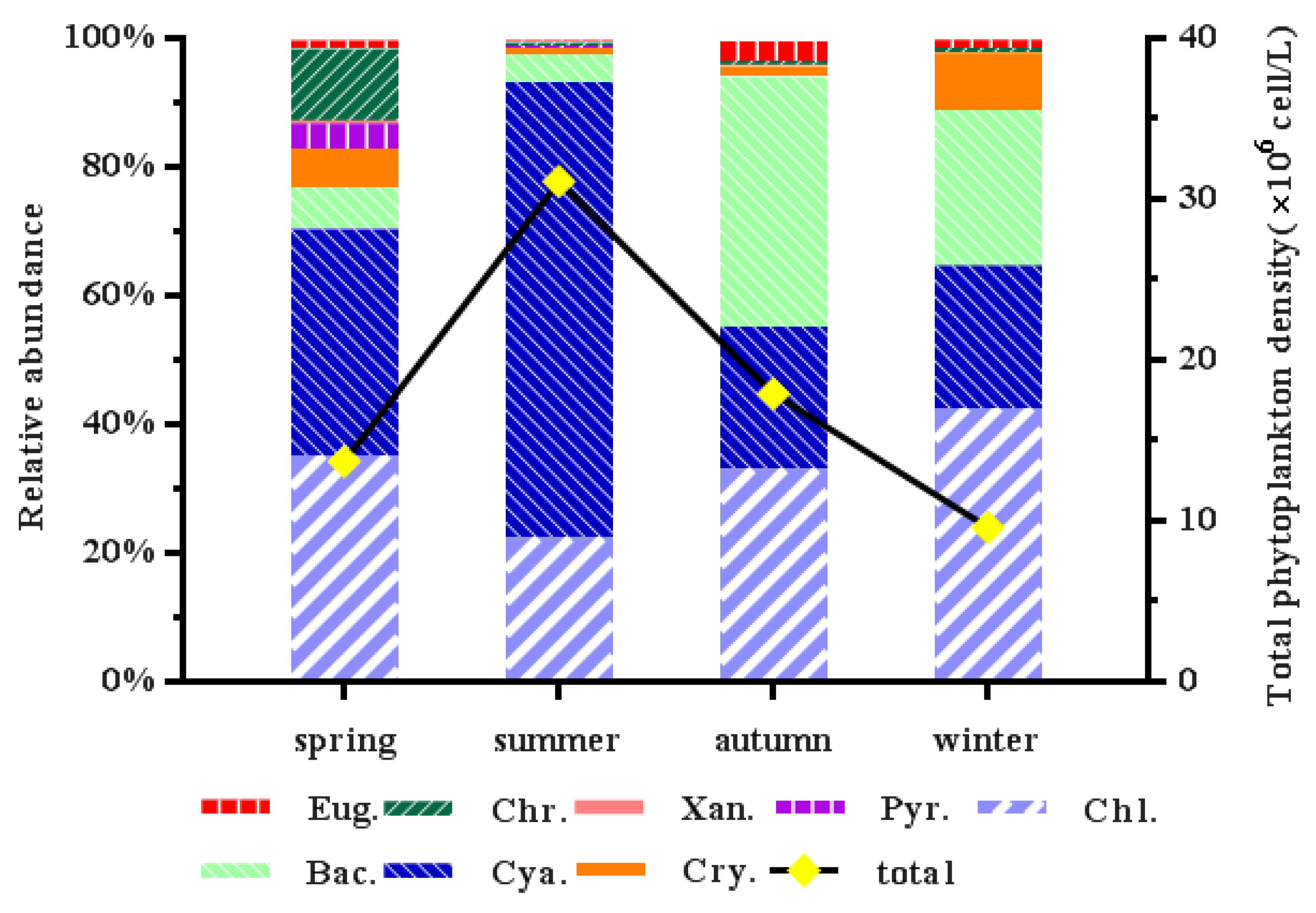
3.3. Phytoplankton Community Diversity and Dominant Species
3.4. Spatial and Temporal Variation of Phytoplankton Communities
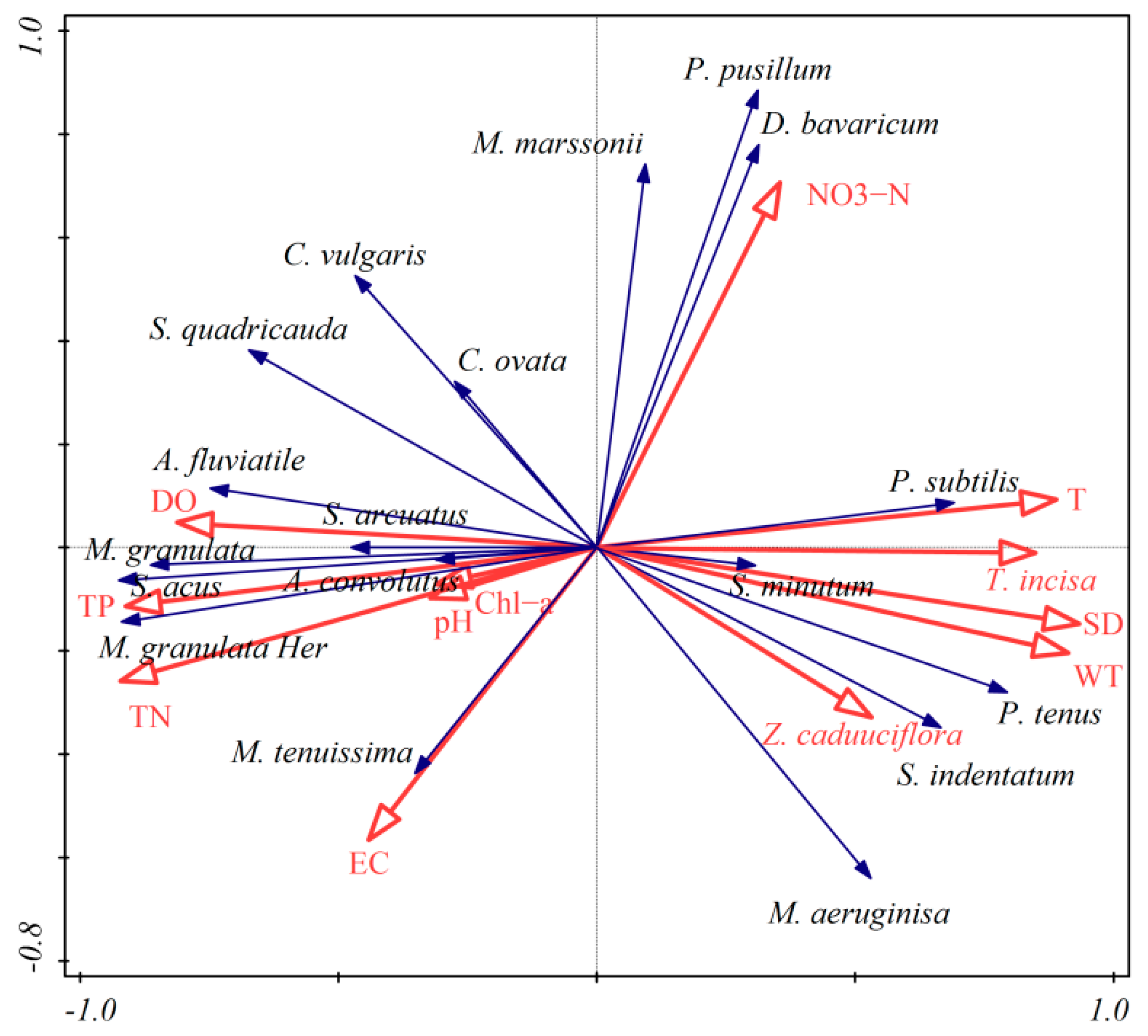
3.5. Relationship between Phytoplankton Communities and Environmental Parameters
4. Discussion
4.1. Aquatic Vegetation Restored after Removal of Purse Seine
4.2. Phytoplankton Community Composition and Spatial and Temporal Variation
4.3. Response Mechanisms of Phytoplankton Communities to Aquatic Vegetation Restoration
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kallfa, A.K.; Babani, F.; Kraja, A.Y. Evaluation of the Trophic Level of Kune and Vain Lagoons in Albania, Using Phytoplankton as a Bioindicator. J. Sustain. Dev. Energy Water Environ. Syst. 2014, 2, 61–67. [Google Scholar] [CrossRef]
- Gao, S.R.; Pan, L.J.; Sun, F.Y. Assessment of environmental water pollution and eutrophication by aquatic organisms. Environ. Sci. Manag. 2006, 31, 174–176. [Google Scholar]
- Wei, Y.Q.; Sun, J.; Zhang, G.C.; Wang, Z.X.; Wang, F. Environmental factors controlling the dynamics of phytoplankton communities during spring and fall seasons in the southern Sunda Shelf. Environ. Sci. Pollut. Res. 2020, 27, 23222–23233. [Google Scholar] [CrossRef] [PubMed]
- Miao, X.M.; Wang, S.K.; Liu, M.Q.; Ma, J.; Hu, J.; Li, T.L.; Chen, L.J. Changes in the phytoplankton community structure of the Backshore Wetland of Expo Garden, Shanghai from 2009 to 2010. Aquacult. Fish. 2019, 4, 198–204. [Google Scholar] [CrossRef]
- Liu, C.Q.; Liu, L.S.; Shen, H.T. Seasonal variations of phytoplankton community structure in relation to physico-chemical factors in Lake Baiyangdian, China. Procedia Environ. Sci. 2010, 2, 1622–1631. [Google Scholar] [CrossRef] [Green Version]
- Loza, V.; Perona, E.; Mateo, P. Specific responses to nitrogen and phosphorus enrichment in cyanobacteria: Factors influencing changes in species dominance along eutrophic gradients. Water Res. 2014, 48, 622–631. [Google Scholar] [CrossRef]
- Ma, J.R.; Qin, B.Q.; Paerl, H.W.; Brookes, J.D.; Wu, P.; Zhou, J.; Deng, J.M. Green algal over cyanobacterial dominance promoted with nitrogen and phosphorus additions in a mesocosm study at Lake Taihu, China. Environ. Sci. Pollut. Res. 2015, 22, 5041–5049. [Google Scholar] [CrossRef]
- Wu, B.; Zhao, Q.; Ma, F.K. Study on Restoration of the Relationship between Blocking Lakes and Yangtze River. Environ. Sci. Surv. 2019, 38, 10–14. [Google Scholar]
- Cui, Y.H.; Wang, J. Influence of the sluice on water level and area of Yangtze river-connected lakes: A case study in Shengjin Lake. J. Water Resour. Water Eng. 2018, 29, 47–52. [Google Scholar]
- Song, X.W.; Zhou, L.Z. Effects of habitat changes on spatiotemporal pattern of the wintering waterbird community at Shengjin Lake. J. Anhui Agric. Univ. 2019, 46, 610–617. [Google Scholar]
- Chen, J.Y.; Zhou, L.Z.; Zhou, B.; Xu, R.X.; Xu, W.B. Seasonal dynamics of wintering waterbirds in two shallow lakes along Yangtze River in Anhui Province. Zool. Res. 2011, 32, 540–548. [Google Scholar] [CrossRef]
- Cao, L.; Zhang, Y.; Barter, M.; Lei, G. Anatidae in eastern China during the non-breeding season: Geographical distributions and protection status. Biol. Conserv. 2009, 143, 650–659. [Google Scholar] [CrossRef]
- Li, C.L.; Yang, Y.; Wang, Z.; Zhou, L.Z. The relationship between seasonal water level fluctuation and habitat availability for wintering waterbirds at Shengjin Lake, China. Bird Conserv. Int. 2018, 29, 100–114. [Google Scholar] [CrossRef]
- Xu, H.Q. Study on phytoplankton community structure characteristics in the Shengjin Lake of Anhui Province. Master’s Thesis, Anhui University, Hefei, China, 2011. [Google Scholar]
- Fox, A.D.; Cao, L.; Zhang, Y.; Barter, M.; Meng, F.J.; Wang, S.L. Declines in the tuber-feeding waterbird guild at Shengjin Lake National Nature Reserve, China—A barometer of submerged macrophyte collapse. Aquat. Conserv. Mar. Freshw. Ecosyst. 2011, 21, 82–91. [Google Scholar] [CrossRef]
- Zhang, K.; Jiang, F.Y.; Chen, H.; Wu, Q.L.; Zhou, Z.Z. Temporal and spatial variations in zooplankton communities in relation to environmental factors in four floodplain lakes located in the middle reach of the Yangtze River, China. Environ. Pollut. 2019, 251, 277–284. [Google Scholar] [CrossRef]
- Bian, G.G.; Zhao, W.D.; Da, L. The research and application of the allelopathic effects of submerged plants inhibit the growth of algae. Environ. Dev. 2012, 24, 59–64. [Google Scholar]
- Scheffer, M.; Hosper, S.H.; Meijer, M.L. Alternative equilibria in shallow lakes. Trends Ecol. Evol. 1993, 8, 275–279. [Google Scholar] [CrossRef]
- Carlos, I.; Guillermo, G.; Nestor, M.; Mariana, M.; Elena, R.; Erik, J. Horizontal dynamics of zooplankton in subtropical Lake Blanca (Uruguay) hosting multiple zooplankton predators and aquatic plant refuges. Hydrobiologia 2007, 584, 179–189. [Google Scholar] [CrossRef]
- Ellen, D.; Wouter, J.B. Impact of submerged macrophytes including charophytes on phyto- and zooplankton communities: Allelopathy versus other mechanisms. Aquat. Bot. 2002, 72, 261–274. [Google Scholar] [CrossRef]
- Hu, W.H.; Yu, G.Q.; Liu, Z.Y. The ecology environment and its protection of Shengjin Lake wetland. Territ. Nat. Resour. Study. 2004, 04, 70–71. [Google Scholar]
- State Environmental Protection Bureau (SEPB). Methods of Monitoring and Analysis for Water and Wastewater, 4th ed.; SEPB: Beijing, China, 2002. [Google Scholar]
- GB 3838-2002; Environmental Quality Standards for Surface Water. Ministry of Ecology and Environment of the People’s Republic of China: Beijing, China, 2002.
- Xu, Y.G.; Li, J.A.; Qin, J.H.; Li, Q. Seasonal patterns of water quality and phytoplankton dynamics in surface waters in Guangzhou and Foshan, China. Sci. Total Environ. 2017, 590–591, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Guesdon, S.; Stachowski-Haberkorn, S.; Lambert, C.; Beker, B.; Brach-Papa, C.; Auger, D.; Béchemin, C. Effect of local hydroclimate on phytoplankton groups in the Charente estuary. Estuar. Coast. Shelf Sci. 2016, 181, 325–337. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.S.; Sheng, J.P.; Li, Z.; Hong, Z. Seasonal variation of phytoplankton community in Xiaojiang backwater area during the preliminary operation stage of the Three Gorges Reservoir. Environ. Sci. 2010, 31, 1492–1497. [Google Scholar]
- Hu, H.J.; Wei, Y.X. The Freshwater Algae of China—Systematics, Taxonomy and Ecology; Science Press: Beijing, China, 2006. [Google Scholar]
- Zhang, Z.S.; Huang, X.F. Research Methods of Freshwater Plankton; Science Press: Beijing, China, 1991; Volume 10, pp. 336–339. [Google Scholar]
- Wu, C.; Zhang, Q.G. Research Progress on diversity and quantitative analysis methods of freshwater plankton. Anhui Agric. Sci. Bull. 2009, 15, 41–42. [Google Scholar]
- Lampitt, R.S.; Wishner, K.F.; Turley, C.M.; Angel, M.V. Marine snow studies in the Northeast Atlantic Ocean: Distribution, composition and role as a food source for migrating plankton. Mar. Biol. 1993, 116, 689–702. [Google Scholar] [CrossRef]
- Jiang, Z.B.; Liu, J.J.; Chen, J.F.; Chen, Q.Z.; Yan, X.J.; Xuan, J.L.; Zeng, J.N. Responses of summer phytoplankton community to drastic environmental changes in the Changjiang (Yangtze River) estuary during the past 50 years. Water Res. 2014, 54, 1–11. [Google Scholar] [CrossRef]
- Wendroth, O. Multivariate Analysis of Ecological Data using CANOCO. Vadose Zone J. 2004, 3, 1057. [Google Scholar] [CrossRef]
- Chen, B.P.; Wu, L.Q. Ecological security and sustainable utilization of wetland in Shengjin Lake National Nature Reserve. J. Chizhou Univ. 2006, 20, 78–80, 95. [Google Scholar]
- Xu, L.L.; Xu, W.B.; Sun, Q.Y. Flora and Vegetation in Shengjin Lake. Plant Sci. J. 2008, 27, 264–270. [Google Scholar]
- Liu, J.J.; Zhou, Z.Z.; Tian, H.X. Vascular plant community types and flora in Shengjin Lake National Nature Reserve. J. Biol. 2016, 33, 40–46. [Google Scholar]
- Yang, L. Effects of Lake Wetland Degradation on foraging activities of overwintering white headed crane (Grus monacha). Ph.D. Thesis, Anhui University, Hefei, China, 2015. [Google Scholar]
- Wu, F.Y.; Wang, L.; Xu, M. Community dynamics of phytoplankton and related affecting factors in Shengjin Lake. J. Biol. 2016, 33, 34–39. [Google Scholar]
- Wang, L.; Wang, C.; Deng, D.G.; Zhao, X.X.; Zhou, Z.Z. Temporal and spatial variations in phytoplankton: Correlations with environmental factors in Shengjin Lake, China. Environ. Sci. Pollut. Res. 2015, 22, 14144–14156. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, C.; Zhao, X.X.; Zhou, Z.Z. Seasonal and spatial variation of chlorophyll-a concentration in Shengjin Lake. J. Biol. 2016, 33, 49–52. [Google Scholar]
- Ma, J.R.; Deng, J.M.; Qin, B.Q. Progress and prospects on Cyanobacteria bloom-forming mechanism in lakes. Acta Ecol. Sin. 2013, 33, 3020–3030. [Google Scholar]
- Silva, C.; Torgan, L.C.; Schneck, F. Temperature and surface runoff affect the community of periphytic diatoms and have distinct effects on functional groups: Evidence of a mesocosms experiment. Hydrobiologia 2019, 839, 37–50. [Google Scholar] [CrossRef]
- Ma, H.T.; Zhang, F.; Zhang, H.J. The inhibition effect of two common limnetic submerged macrophytes on Scenedesmus obliquus under co-culture condition. J. Biol. 2020, 37, 72–75. [Google Scholar]
- Xie, X.L.; Zhou, R.; Deng, Z.F. Overcompensation and competitive effects of Microcystis aeruginosa and Scenedesmus obliquus after low temperature and light stresses. Acta Ecol. Sin. 2014, 34, 1224–1234. [Google Scholar]
- Figueredo, C.C.; Giani, A. Seasonal variation in the diversity and species richness of phytoplankton in a tropical eutrophid reservoir. Hydrobiologia 2001, 445, 165–174. [Google Scholar] [CrossRef]
- Zhu, Q.C.; Yang, W.D.; Cheng, H. Test of Potamogeton crispus Decomposing in Water. Mod. Agric. Sci. Technol. 2013, 23, 224–226, 229. [Google Scholar]
- Liu, C.G.; Qiu, J.Q.; Wang, W. Advances on theory of biomanipulation in control of eutrophicated lakes. J. Agro-Environ. Scie. 2004, 23, 198–201. [Google Scholar]
- Wang, L.X.; Wu, G.R. Inhibitory effect of Hydrilla verticillata on Microcystis aeruginosa. J. Lake Sci. 2004, 16, 337–342. [Google Scholar]
- Hu, H.Y.; Men, Y.J.; Li, F.M.; Men, Y. Research progress on phyto-allelopathic algae control. Ecol. Environ. Sci. 2006, 15, 153–157. [Google Scholar]
- Park, M.H.; Chung, I.M.; Ahmad, A.; Hwang, S.J. Growth inhibition of unicellular and colonial Microcystis strains (Cyanophyceae) by compounds isolated from rice (Oryza Sativa) hulls. Aquat. Bot. 2008, 90, 309–314. [Google Scholar] [CrossRef]
- Elisabeth, M. Allelopathy of Aquatic Autotrophs. Crit. Rev. Plant Sci. 2003, 22, 313–339. [Google Scholar] [CrossRef] [Green Version]
- Friebe, A.; Roth, U.; Kück, P.; Schnabl, H.; Schulz, M. Effects of 2, 4-dihydroxy-1, 4-benzoxazin-3-ones on the activity of plasma membrane H +− ATPase. Phytochemistry 1997, 44, 979–983. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Factors | Spring (April) | Summer (July) | Autumn (October) | Winter (November) | Seasonal p-Value | Spatial p-Value | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| UL | ML | UL | ML | UL | ML | UL | ML | |||
| WT (°C) | 27.66 ± 0.73 | 27.25 ± 0.31 | 34.92 ± 0.48 | 34.43 ± 0.15 | 17.35 ± 2.65 | 16.27 ± 0.26 | 8.57 ± 0.25 | 8.13 ± 0.11 | ** | * |
| pH | 8.47 ± 0.45 | 9.03 ± 0.15 | 8.6 ± 0.18 | 8.77 ± 0.1 | 8.64 ± 0.36 | 8.43 ± 0.13 | 8.77 ± 0.04 | 8.52 ± 0.18 | - | - |
| DO (mg/L) | 7.98 ± 1.02 | 8.39 ± 0.29 | 7.67 ± 0.39 | 7.72 ± 0.16 | 9.73 ± 0.17 | 9.45 ± 0.08 | 11.75 ± 0.11 | 11.77 ± 0.15 | * | * |
| Cond (Us/cm) | 116.05 ± 6.82 | 111.21 ± 2.16 | 190.57 ± 6.5 | 177.04 ± 3.36 | 247.5 ± 8.5 | 234.5 ± 7.76 | 157.73 ± 2.44 | 154.38 ± 3.52 | * | - |
| SD (m) | 1.08 ± 0.1 | 0.86 ± 0.12 | 1.37 ± 0.56 | 1.14 ± 0.19 | 0.20 ± 0.06 | 0.19 ± 0.06 | 0.16 ± 0.01 | 0.13 ± 0.02 | ** | * |
| WL (m) | 2.54 ± 0.45 | 3.66 ± 0.38 | 4.19 ± 0.30 | 5.57 ± 0.62 | 0.45 ± 0.25 | 0.97 ± 0.27 | 1.08 ± 0.23 | 1.43 ± 0.13 | ** | * |
| Turb | 4.06 ± 1.63 | 8.06 ± 1.08 | 5.54 ± 1.95 | 5.48 ± 1.68 | 41 ± 24.2 | 56.33 ± 44 | 203.72 ± 77 | 232 ± 89.37 | ** | * |
| TP (mg/L) | 0.009 ± 0.1 | 0.013 ± 0.01 | 0.027 ± 0.01 | 0.036 ± 0.02 | 0.072 ± 0.06 | 0.077 ± 0.04 | 0.051 ± 0.01 | 0.073 ± 0.09 | ** | * |
| TN (mg/L) | 0.97 ± 0.21 | 1.15 ± 0.36 | 0.57 ± 0.52 | 0.92 ± 0.35 | 1.07 ± 0.14 | 1.33 ± 0.82 | 1.12 ± 0.51 | 1.18 ± 1.08 | * | * |
| NO3-N (mg/L) | 2.12 ± 0.99 | 0.82 ± 0.33 | 0.15 ± 0.05 | 0.06 ± 0.04 | 0.013 ± 0.01 | 0.14 ± 0.17 | 0.18 ± 0.02 | 0.21 ± 0.06 | - | - |
| NH3-N (mg/L) | 0.09 ± 0.05 | 0.13 ± 0.1 | 0.09 ± 0.05 | 0.13 ± 0.09 | 0.89 ± 0.36 | 0.81 ± 0.23 | 0.24 ± 0.07 | 0.28 ± 0.14 | * | - |
| Chl a (μg/L) | 2.47 ± 4.72 | 3.42 ± 3.15 | 1.32 ± 0.71 | 2.58 ± 1.05 | 1.52 ± 1.1 | 0.87 ± 0.41 | 3.08 ± 0.78 | 2.92 ± 0.71 | * | * |
| Species | Dominance | ||||
|---|---|---|---|---|---|
| Spring (April) | Summer (July) | Autumn (October) | Winter (January) | ||
| Cyanophyta | P. subtilis | 0.012 | 0.228 | 0.032 | 0.019 |
| P. tenuis | 0.003 | 0.035 | 0.014 | 0.01 | |
| M. tenuissima | 0.002 | 0.005 | 0.011 | 0.033 | |
| M. marssonii | 0.064 | 0.031 | 0.002 | 0.004 | |
| M. aeruginosa | - | 0.031 | - | 0.004 | |
| Chlorophyta | S. minutum | 0.0007 | 0.035 | 0.006 | 0.017 |
| S. indentatum | 0.005 | 0.025 | 0.009 | - | |
| Chlorella vulgaris | 0.015 | 0.011 | 0.059 | 0.019 | |
| S. arcuatus | 0.008 | 0.0006 | 0.035 | 0.002 | |
| A. convolutus | 0.006 | 0.017 | 0.012 | 0.043 | |
| S. quadricauda | 0.016 | - | 0.0031 | 0.081 | |
| A. fluviatile | 0.1 | 0.001 | - | 0.065 | |
| Bacillariophyta | M. granulata Her | 0.002 | - | 0.134 | 0.034 |
| M. granulata | 0.005 | 0.003 | 0.051 | 0.018 | |
| S. acus | 0.013 | 0.002 | 0.019 | 0.073 | |
| Cryptophyta | C. ovata | 0.015 | 0.009 | 0.01 | 0.064 |
| Pyrrophyta | P. pusillum | 0.032 | 0.008 | - | - |
| Chrysophyta | D. bavaricum | 0.079 | 0.01 | 0.0002 | - |
| Sort Axis | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| Eigenvalue | 0.329 | 0.211 | 0.126 | 0.101 |
| Species-environment correlation | 0.852 | 0.757 | 0.682 | 0.714 |
| Cumulative percentage of species | 22.9 | 31.5 | 36.3 | 38.6 |
| Cumulative percentage of species-environment relationships | 43.4 | 71.9 | 83.8 | 91.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, X.; Chen, J.; Guo, W.; Zhou, Z.; Ye, X. Response of Phytoplankton Community Structure to Vegetation Restoration after Removal of Purse Seine in Shengjin Lake. Diversity 2022, 14, 178. https://doi.org/10.3390/d14030178
Zheng X, Chen J, Guo W, Zhou Z, Ye X. Response of Phytoplankton Community Structure to Vegetation Restoration after Removal of Purse Seine in Shengjin Lake. Diversity. 2022; 14(3):178. https://doi.org/10.3390/d14030178
Chicago/Turabian StyleZheng, Xudong, Jingwen Chen, Wenli Guo, Zhongze Zhou, and Xiaoxin Ye. 2022. "Response of Phytoplankton Community Structure to Vegetation Restoration after Removal of Purse Seine in Shengjin Lake" Diversity 14, no. 3: 178. https://doi.org/10.3390/d14030178
APA StyleZheng, X., Chen, J., Guo, W., Zhou, Z., & Ye, X. (2022). Response of Phytoplankton Community Structure to Vegetation Restoration after Removal of Purse Seine in Shengjin Lake. Diversity, 14(3), 178. https://doi.org/10.3390/d14030178