
The Expanding Role of DNA Barcodes: Indispensable Tools for Ecology, Evolution, and Conservation



Abstract
:1. Introduction
1.1. Accurate and Reliable Identification of Species in Taxonomy, Ecology, Evolution, and Conservation
1.2. Generating, Applying, and Sharing DNA Barcodes
1.2.1. Sequencing Technology
1.2.2. Novel Computational Resources and Software
1.2.3. National and International Sequencing Consortia
1.2.4. Building the Plant DNA Barcode Library
1.3. The Purpose and Structure of This Review
2. Current Applications
2.1. Improving Taxonomy and Species Identification
2.1.1. Defining Species Boundaries
2.1.2. Regional Biodiversity Assessments
2.2. Quantify Species Diversity
2.2.1. Species Richness and Phylogenetic Diversity
2.2.2. BCI as an Exemplar Tropical Field Site for DNA Barcoding
2.3. Determining Community Structure and Species Interactions
2.3.1. Community Evolution and Assembly
2.3.2. Herbivory and Food Webs
2.3.3. Symbiotic Relationships and Plant-Pollinator Interactions
2.4. Protecting Species
2.4.1. Forensics and Monitoring Traffic in Endangered Plants
2.4.2. Tracking Biological Invasions
2.4.3. Conservation Assessment
3. Looking Forward: The Expanding Technological Spectrum of DNA Barcodes
3.1. Metabarcoding
3.2. Super- and Ultrabarcoding
3.3. Macrogenetics
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; DeWaard, J.R. Biological Identifications through DNA Barcodes. Proc. R. Soc. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeSalle, R.; Goldstein, P. Review and Interpretation of Trends in DNA Barcoding. Front. Ecol. Evol. 2019, 7, 302. [Google Scholar] [CrossRef] [Green Version]
- Grant, D.M.; Brodnicke, O.B.; Evankow, A.M.; Ferreira, A.O.; Fontes, J.T.; Hansen, A.K.; Jensen, M.R.; Gba, T.; Kalaycı, E.; Leeper, A.; et al. The Future of DNA Barcoding: Reflections from Early Career Researchers. Diversity 2021, 13, 313. [Google Scholar] [CrossRef]
- Kress, W.J.; Erickson, D.L. DNA Barcodes: Methods and Protocols; Humana Press: Totowa, NJ, USA, 2012; volume 858, pp. 3–8. [Google Scholar]
- Kress, W.J. Plant DNA Barcodes: Applications Today and in the Future. Front. Plant Syst. Evol. 2017, 55, 291–307. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, M.J.; Szabo, C.; Ford, C.S.; Yarom, Y.; Croxford, A.E.; Camp, A.; Gooding, P. Replacing Sanger with Next Generation Sequencing to Improve Coverage and Quality of Reference DNA Barcodes for Plants. Sci. Rep. 2017, 7, 46040. [Google Scholar] [CrossRef] [Green Version]
- Cruaud, P.; Rasplus, J.-Y.; Rodriguez, L.J.; Cruaud, A. High-Throughput Sequencing of Multiple Amplicons for Barcoding and Integrative Taxonomy. Sci. Rep. 2017, 7, 41948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gostel, M.R.; Zúñiga, J.D.; Kress, W.J.; Funk, V.A.; Puente-Lelievre, C. Microfluidic Enrichment Barcoding (MEBarcoding): A New Method for High Throughput Plant DNA Barcoding. Sci. Rep. 2020, 10, 8701. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, C.; Sun, Y.; Chen, X.; Dong, W.; Yang, X.; Zhou, S. Method for Quick DNA Barcode Reference Library Construction. Ecol. Evol. 2021, 11, 11627–11638. [Google Scholar] [CrossRef]
- Page, R.D.M. DNA Barcoding and Taxonomy: Dark Taxa and Dark Texts. Philos. Trans. R. Soc. B 2016, 371. [Google Scholar] [CrossRef] [Green Version]
- Nic Lughadha, E.M.; Govaerts, R.H.A.; Belyaeva, I.; Black, N.; Lindon, H.; Allkin, R.; Magill, R.E.; Nicolson, N. Counting Counts: Revised Estimates of Numbers of Accepted Species of Flowering Plants, Seed Plants, Vascular Plants and Land Plants with a Review of Other Recent. Phytotaxa 2016, 272, 82–88. [Google Scholar] [CrossRef] [Green Version]
- Kane, N.; Ecology, Q.C.-M. Botany without Borders: Barcoding in Focus. Mol. Ecol. 2008, 17, 5175–5176. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, Y.; Henry, R.J.; Rossetto, M.; Wang, Y.; Chen, S. Plant DNA Barcoding: From Gene to Genome. Biol. Rev. 2015, 90, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Kane, N.; Sveinsson, S.; Dempewolf, H.; Yang, J.Y.; Zhang, D.; Engels, J.M.M.; Cronk, Q. Ultra-Barcoding in Cacao (Theobroma Spp.; Malvaceae) Using Whole Chloroplast Genomes and Nuclear Ribosomal DNA. Am. J. Bot. 2012, 99, 320–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, Y.; Liu, C.; Yang, J.; Jin, L.; Yang, Z.; Yang, J.B. Ultra-Barcoding Discovers a Cryptic Species in Paris Yunnanensis (Melanthiaceae), a Medicinally Important Plant. Front. Plant Sci. 2020, 11, 411. [Google Scholar] [CrossRef] [Green Version]
- Coissac, E.; Hollingsworth, P.M.; Lavergne, S.; Taberlet, P. From Barcodes to Genomes: Extending the Concept of DNA Barcoding. Mol. Ecol. 2016, 25, 1423–1428. [Google Scholar] [CrossRef] [Green Version]
- Bohmann, K.; Mirarab, S.; Bafna, V.; Gilbert, M.T.P. Beyond DNA Barcoding: The Unrealized Potential of Genome Skim Data in Sample Identification. Mol. Ecol. 2020, 2521–2534. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The Barcode of Life Data System: Barcoding. Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Young, R.G.; Yu, J.; Cote, M.J.; Hanner, R.H. The Molecular Data Organization for Publication (MDOP) R Package to Aid the Upload of Data to Shared Databases. Biodivers. Data J. 2020, 8, e50630. [Google Scholar] [CrossRef]
- Fontes, J.T.; Vieira, P.E.; Ekrem, T.; Soares, P.; Costa, F.O. BAGS: An Automated Barcode, Audit & Grade System for DNA Barcode Reference Libraries. Mol. Ecol. Resour. 2021, 21, 573–583. [Google Scholar] [CrossRef]
- Young, R.; Gill, R.; Gillis, D.; Hanner, R. Molecular Acquisition, Cleaning and Evaluation in R (MACER)—A Tool to Assemble Molecular Marker Datasets from BOLD and GenBank. Biodivers. Data J. 2021, 9. [Google Scholar] [CrossRef]
- Oliveira, R.R.M.; Nunes, G.L.; de Lima, T.G.L.; Oliveira, G.; Alves, R. PIPEBAR and OverlapPER: Tools for a Fast and Accurate DNA Barcoding Analysis and Paired-End Assembly. BMC Bioinform. 2018, 19, 297. [Google Scholar] [CrossRef] [PubMed]
- Sahlin, K.; Lim, M.C.W.; Prost, S. NGSpeciesID: DNA Barcode and Amplicon Consensus Generation from Long-Read Sequencing Data. Ecol. Evol. 2021, 11, 1392–1398. [Google Scholar] [CrossRef]
- Bik, H.M. Just Keep It Simple? Benchmarking the Accuracy of Taxonomy Assignment Software in Metabarcoding Studies. Mol. Ecol. Resour. 2021, 21, 2187–2189. [Google Scholar] [CrossRef]
- Hleap, J.S.; Littlefair, J.E.; Steinke, D.; Hebert, P.D.N.; Cristescu, M.E. Assessment of Current Taxonomic Assignment Strategies for Metabarcoding Eukaryotes. Mol. Ecol. Resour. 2021, 21, 2190–2203. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing Taxonomic Classification of Marker-Gene Amplicon Sequences with QIIME 2’s Q2-Feature-Classifier Plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Murali, A.; Bhargava, A.; Wright, E.S. IDTAXA: A Novel Approach for Accurate Taxonomic Classification of Microbiome Sequences. Microbiome 2018, 6, 140. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson-Palme, J.; Richardson, R.T.; Meola, M.; Wurzbacher, C.; Tremblay, É.D.; Thorell, K.; Kanger, K.; Eriksson, K.M.; Bilodeau, G.J.; Johnson, R.M.; et al. MetaxA2 Database Builder: Enabling Taxonomic Identification from Metagenomic or Metabarcoding Data Using Any Genetic Marker. Bioinformatics 2018, 34, 4027–4033. [Google Scholar] [CrossRef]
- Kahlke, T.; Ralph, P. BASTA–Taxonomic Classification of Sequences and Sequence Bins Using Last Common Ancestor Estimations. Methods Ecol. Evol. 2018, 10, 100–103. [Google Scholar] [CrossRef] [Green Version]
- Wood, D.E.; Salzberg, S.L. Kraken: Ultrafast Metagenomic Sequence Classification Using Exact Alignments. Genome Biol. 2014, 15, R46. [Google Scholar] [CrossRef] [Green Version]
- Somervuo, P.; Koskela, S.; Pennanen, J.; Nilsson, R.H.; Ovaskainen, O. Unbiased Probabilistic Taxonomic Classification for DNA Barcoding. Bioinformatics 2016, 32, 2920–2927. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Meher, P.K.; Sahu, T.K.; Gahoi, S.; Tomar, R.; Rao, A.R. FunbarRF: DNA Barcode-Based Fungal Species Prediction Using Multiclass Random Forest Supervised Learning Model. BMC Genet. 2019, 20, 2. [Google Scholar] [CrossRef] [PubMed]
- Curd, E.E.; Gold, Z.; Kandlikar, G.S.; Gomer, J.; Ogden, M.; O’Connell, T.; Pipes, L.; Schweizer, T.M.; Rabichow, L.; Lin, M.; et al. Anacapa Toolkit: An Environmental DNA Toolkit for Processing Multilocus Metabarcode Datasets. Methods Ecol. Evol. 2019, 10, 1469–1475. [Google Scholar] [CrossRef] [Green Version]
- Hobern, D. Bioscan: Dna Barcoding to Accelerate Taxonomy and Biogeography for Conservation and Sustainability. Genome 2021, 64, 161–164. [Google Scholar] [CrossRef]
- Seberg, O.; Droege, G.; Barker, K.; Coddington, J.A.; Funk, V.; Gostel, M.; Petersen, G.; Smith, P.P. Global Genome Biodiversity Network: Saving a Blueprint of the Tree of Life—A Botanical Perspective. Ann. Bot. 2016, 118, 393–399. [Google Scholar] [CrossRef] [Green Version]
- Lewin, H.A.; Robinson, G.E.; Kress, W.J.; Baker, W.J.; Coddington, J.; Crandall, K.A.; Durbin, R.; Edwards, S.V.; Forest, E.; Thomas, M.; et al. Earth BioGenome Project: Sequencing Life for the Future of Life. Proc. Natl. Acad Sci. USA 2018, 115, 4325–4333. [Google Scholar] [CrossRef] [Green Version]
- Lewin, H.; Richards, S.; Aiden, E.L.; Allende, M.L.; Archibald, J.M.; Bálint, M.; Barker, K.B.; Baumgartner, B.; Belov, K.; Bertorelle, G.; et al. The Earth BioGenome Project 2020: Starting the Clock. Proc. Natl. Acad Sci. USA 2022, 119, e2115635118. [Google Scholar] [CrossRef] [PubMed]
- Kress, W.J.; Erickson, D.L. DNA Barcodes: Genes, Genomics, and Bioinformatics. Proc. Natl. Acad. Sci. USA 2008, 105, 2761–2762. [Google Scholar] [CrossRef] [Green Version]
- Bezeng, B.S.; Davies, T.J.; Daru, B.H.; Kabongo, R.M.; Maurin, O.; Yessoufou, K.; Van Der Bank, H.; Van Der Bank, M. Ten Years of Barcoding at the African Centre for DNA Barcoding. Genome 2017, 60, 629–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.Q.; Lv, Q.; Zhang, A.B. Sixteen Years of DNA Barcoding in China: What Has Been Done? What Can Be Done? Front. Ecol. Evol. 2020, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- The Darwin Tree of Life Project Consortium. Sequence Locally, Think Globally: The Darwin Tree of Life Project. Proc. Natl. Acad Sci. USA 2022, 119, e2115642118. [Google Scholar] [CrossRef]
- Anderson-Teixeira, K.J.; Davies, S.J.; Bennett, A.C.; Gonzalez-Akre, E.B.; Muller-Landau, H.C.; Joseph Wright, S.; Abu Salim, K.; Almeyda Zambrano, A.M.; Alonso, A.; Baltzer, J.L.; et al. CTFS-ForestGEO: A Worldwide Network Monitoring Forests in an Era of Global Change. Glob. Chang. Biol. 2015, 21, 528–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kress, W.J.; Lopez, I.C.; Erickson, D.L. Generating Plant DNA Barcodes for Trees in Long-Term Forest Dynamics Plots. Methods Mol. Biol. 2012, 858, 441–458. [Google Scholar] [CrossRef] [PubMed]
- Erickson, D.L.; Jones, F.A.; Swenson, N.G.; Pei, N.; Bourg, N.; Chen, W.; Davies, S.J.; Ge, X.; Hao, Z.; Howe, R.W.; et al. Comparative Evolutionary Diversity and Phylogenetic Structure across Multiple Forest Dynamics Plots: A Mega-Phylogeny Approach. Front. Genet. 2014, 5, 358. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Liu, J.; Shan, Y.-X.; Ge, X.-J.; Burgess, K.S. The Use of DNA Barcodes to Estimate Phylogenetic Diversity in Forest Communities of Southern China. Ecol. Evol. 2019, 9, 5372–5379. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Qian, X.; Zheng, Z.; Shi, M.; Chang, X.; Li, X.; Liu, J.; Tu, T.; Zhang, D. DNA Barcoding the Flowering Plants from the Tropical Coral Islands of Xisha (China). Ecol. Evol. 2018, 8, 10587–10593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, B.A.; Musili, P.M.; Kurukura, S.; Hassan, A.A.; Goheen, J.R.; Kress, W.J.; Kuzmina, M.; Pringle, R.M.; Kartzinel, T.R. Plant DNA-Barcode Library and Community Phylogeny for a Semi-Arid East African Savanna. Mol. Ecol. Resour. 2019, 19, 838–846. [Google Scholar] [CrossRef]
- Bringloe, T.T.; Sjøtun, K.; Saunders, G.W. A DNA Barcode Survey of Marine Macroalgae from Bergen (Norway). Mar. Biol. Res. 2019, 15, 580–589. [Google Scholar] [CrossRef]
- Hernández-León, S.; Little, D.P.; Acevedo-Sandoval, O.; Gernandt, D.S.; Rodríguez-Laguna, R.; Saucedo-García, M.; Arce-Cervantes, O.; Razo-Zárate, R.; Espitia-López, J. Plant Core DNA Barcode Performance at a Local Scale: Identification of the Conifers of the State of Hidalgo, Mexico. Syst. Biodivers. 2018, 16, 791–806. [Google Scholar] [CrossRef]
- Nitta, J.H.; Ebihara, A.; Smith, A.R. A Taxonomic and Molecular Survey of the Pteridophytes of the Nectandra Cloud Forest Reserve, Costa Rica. PLoS ONE 2020, 15, e0241231. [Google Scholar] [CrossRef]
- Chen, S.; Yao, H.; Han, J.; Liu, C.; Song, J.; Shi, L.; Zhu, Y.; Ma, X.; Gao, T.; Pang, X.; et al. Validation of the ITS2 Region as a Novel DNA Barcode for Identifying Medicinal Plant Species. PLoS ONE 2010, 5, e8613. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhao, J.; Erickson, D.L.; Xia, N.; Kress, W.J. Testing DNA Barcodes in Closely Related Species of Curcuma (Zingiberaceae) from Myanmar and China. Mol. Ecol. Resour. 2014, 15, 337–348. [Google Scholar] [CrossRef]
- Wang, D.Y.; Wang, Q.; Wang, Y.L.; Xiang, X.G.; Huang, L.Q.; Jin, X.H. Evaluation of DNA Barcodes in Codonopsis (Campanulaceae) and in Some Large Angiosperm Plant Genera. PLoS ONE 2017, 12, e0170286. [Google Scholar] [CrossRef]
- Nieves, K.G.; van Ee, B.W. DNA Barcoding of Adiantum (Pteridaceae: Vittarioideae) in Puerto Rico. Caribb. J. Sci. 2021, 51, 287–313. [Google Scholar] [CrossRef]
- Dal Forno, M.; Lawrey, J.D.; Moncada, B.; Bungartz, F.; Grube, M.; Schuettpelz, E.; Lücking, R. DNA Barcoding of Fresh and Historical Collections of Lichen-Forming Basidiomycetes in the Genera Cora and Corella (Agaricales: Hygrophoraceae): A Success Story? Diversity 2022, 14. [Google Scholar]
- Sánchez-C, D.; Richardson, J.E.; Hart, M.; Serrano, J.; Cárdenas, D.; Gonzalez, M.A.; Cortés-B, R. A Plea to DNA Barcode Type Specimens: An Example from Micropholis (Sapotaceae). Taxon 2021, 71, 154–167. [Google Scholar] [CrossRef]
- Braukmann, T.W.A.; Kuzmina, M.L.; Sills, J.; Zakharov, E.V.; Hebert, P.D.N. Testing the Efficacy of DNA Barcodes for Identifying the Vascular Plants of Canada. PLoS ONE 2017, 12, e0169515. [Google Scholar] [CrossRef]
- Jones, L.; Twyford, A.D.; Ford, C.R.; Rich, T.C.G.; Davies, H.; Forrest, L.L.; Hart, M.L.; McHaffie, H.; Brown, M.R.; Hollingsworth, P.M.; et al. Barcode UK: A Complete DNA Barcoding Resource for the Flowering Plants and Conifers of the United Kingdom. Mol. Ecol. Resour. 2021, 21, 2050–2062. [Google Scholar] [CrossRef] [PubMed]
- Mosa, K.A.; Gairola, S.; Jamdade, R.; El-Keblawy, A.; Al Shaer, K.I.; Al Harthi, E.K.; Shabana, H.A.; Mahmoud, T. The Promise of Molecular and Genomic Techniques for Biodiversity Research and DNA Barcoding of the Arabian Peninsula Flora. Front. Plant Sci. 2019, 9, 1929. [Google Scholar] [CrossRef] [PubMed]
- Shapcott, A.; Forster, P.I.; Guymer, G.P.; McDonald, W.J.F.; Faith, D.P.; Erickson, D.; Kress, W.J. Mapping Biodiversity and Setting Conservation Priorities for SE Queensland’s Rainforests Using DNA Barcoding. PLoS ONE 2015, 10, e0122164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kress, W.J.; Erickson, D.L.; Jones, F.A.; Swenson, N.G.; Perez, R.; Sanjur, O.; Bermingham, E. Plant DNA Barcodes and a Community Phylogeny of a Tropical Forest Dynamics Plot in Panama. Proc. Natl. Acad Sci. USA 2009, 106, 18621–18626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurado-Rivera, J.A.; Vogler, A.P.; Reid, C.A.M.; Petitpierre, E.; Gómez-Zurita, J. DNA Barcoding Insect-Host Plant Associations. Proc. R. Soc. B Biol. Sci. 2009, 276, 639–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollingsworth, P.M.; Graham, S.W.; Little, D.P. Choosing and Using a Plant DNA Barcode. PLoS ONE 2011, 6, e19254. [Google Scholar] [CrossRef]
- Hollingsworth, P.M.; Li, D.Z.; Van Der Bank, M.; Twyford, A.D. Telling Plant Species Apart with DNA: From Barcodes to Genomes. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371. [Google Scholar] [CrossRef] [PubMed]
- Kress, W.J.; Wurdack, K.J.; Zimmer, E.A.; Weigt, L.A.; Janzen, D.H. Use of DNA Barcodes to Identify Flowering Plants. Proc. Natl. Acad. Sci. USA 2005, 102, 8369–8374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kress, W.J.; García-Robledo, C.; Uriarte, M.; Erickson, D.L. DNA Barcodes for Ecology, Evolution, and Conservation. Trends Ecol. Evol. 2014, 30, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Valentini, A.; Miquel, C.; Nawaz, M.A.; Bellemain, E.; Coissac, E.; Pompanon, F.; Gielly, L.; Cruaud, C.; Nascetti, G.; Wincker, P.; et al. New Perspectives in Diet Analysis Based on DNA Barcoding and Parallel Pyrosequencing: The TrnL Approach. Mol. Ecol. Resour. 2009, 9, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Wilson, E.O. Half Earth: Our Planet’s Fight for Life; WW Norton and Company: New York, NY, USA, 2016. [Google Scholar]
- Hebert, P.D.N.; Penton, E.H.; Burns, J.M.; Janzen, D.H.; Hallwachs, W. Ten Species in One: DNA Barcoding Reveals Cryptic Species in the Neotropical Skipper Butterfly Astraptes Fulgerator. Proc. Natl. Acad Sci. USA 2004, 104, 41. [Google Scholar] [CrossRef] [Green Version]
- Janzen, D.H.; Hallwachs, W.; Blandin, P.; Burns, J.M.; Cadiou, J.M.; Chacon, I.; Dapkey, T.; Deans, A.R.; Epstein, M.E.; Espinoza, B.; et al. Integration of DNA Barcoding into an Ongoing Inventory of Complex Tropical Biodiversity. Mol. Ecol. Resour. 2009, 9, 1–26. [Google Scholar] [CrossRef] [Green Version]
- García-Robledo, C.; Kuprewicz, E.K.; Staines, C.L.; Kress, W.J.; Erwin, T.L. Using a Comprehensive DNA Barcode Library to Detect Novel Egg and Larval Host Plant Associations in a Cephaloleia Rolled-Leaf Beetle (Coleoptera: Chrysomelidae). Sci. Rep. 2013, 8, 13398. [Google Scholar]
- García-Robledo, C.; Kuprewicz, E.K.; Staines, C.L.; Erwin, T.L.; Kress, W.J. Limited Tolerance by Insects to High Temperatures across Tropical Elevational Gradients and the Implications of Global Warming for Extinction. Proc. Natl. Acad Sci. USA 2016, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, M.A.; Baraloto, C.; Engel, J.; Mori, S.A.; Pétronelli, P.; Riéra, B.; Roger, A.; Thébaud, C.; Chave, J. Identification of Amazonian Trees with DNA Barcodes. PLoS ONE 2009, 4, e7483. [Google Scholar] [CrossRef] [PubMed]
- Dexter, K.G.; Pennington, T.D.; Cunningham, C.W. Using DNA to Assess Errors in Tropical Tree Identifications: How Often Are Ecologists Wrong and When Does It Matter? Ecol. Monogr. 2010, 80, 267–286. [Google Scholar] [CrossRef] [Green Version]
- Kenfack, D.; Abiem, I.; Chapman, H. The Efficiency of DNA Barcoding in the Identification of Afromontane Forest Tree Species. Diversity 2022, 14. [Google Scholar]
- Costion, C.; Ford, A.; Cross, H.; Crayn, D.; Harrington, M.; Lowe, A. Plant Dna Barcodes Can Accurately Estimate Species Richness in Poorly Known Floras. PLoS ONE 2011, 6, e26841. [Google Scholar] [CrossRef] [Green Version]
- Parmentier, I.; Duminil, J.; Kuzmina, M.; Philippe, M.; Thomas, D.W.; Kenfack, D.; Chuyong, G.B.; Cruaud, C.; Hardy, O.J. How Effective Are DNA Barcodes in the Identification of African Rainforest Trees? PLoS ONE 2013, 8, e54921. [Google Scholar] [CrossRef]
- Dick, C.W.; Webb, C.O. Plant DNA Barcodes, Taxonomic Management, and Species Discovery in Tropical Forests. DNA Barcodes 2012, 858, 379–393. [Google Scholar] [CrossRef] [Green Version]
- Ter Steege, H.; Pitman, N.C.A.; Sabatier, D.; Baraloto, C.; Salomão, R.P.; Guevara, J.E.; Phillips, O.L.; Castilho, C.V.; Magnusson, W.E.; Molino, J.F.; et al. Hyperdominance in the Amazonian Tree Flora. Science 2013, 342, 1243092. [Google Scholar] [CrossRef] [Green Version]
- Pante, E.; Schoelinck, C.; Puillandre, N. From Integrative Taxonomy to Species Description: One Step Beyond. Syst. Biol. 2015, 64, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Meierotto, S.; Sharkey, M.J.; Janzen, D.H.; Hallwachs, W.; Hebert, P.D.N.; Chapman, E.G.; Smith, M.A. A Revolutionary Protocol to Describe Understudied Hyperdiverse Taxa and Overcome the Taxonomic Impediment. Dtsch. Entomol. Zeitschrift 2019, 66, 119–145. [Google Scholar] [CrossRef] [Green Version]
- Funk, V.A.; Gostel, M.; Devine, A.; Kelloff, C.L.; Wurdack, K.; Tuccinardi, C.; Radosavljevic, A.; Peters, M.; Coddington, J. Guidelines for Collecting Vouchers and Tissues Intended for Genomic Work (Smithsonian Institution): Botany Best Practices. Biodivers. Data J. 2017, 5, e11625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weigand, H.; Beermann, A.J.; Čiampor, F.; Costa, F.O.; Csabai, Z.; Duarte, S.; Geiger, M.F.; Grabowski, M.; Rimet, F.; Rulik, B.; et al. DNA Barcode Reference Libraries for the Monitoring of Aquatic Biota in Europe: Gap-Analysis and Recommendations for Future Work. Sci. Total Environ. 2019, 678, 499–524. [Google Scholar] [CrossRef] [PubMed]
- Litman, J.; Chittaro, Y.; Birrer, S.; Praz, C.; Wermeille, E.; Fluri, M.; Stalling, T.; Schmid, S.; Wyler, S.; Gonseth, Y. A DNA Barcode Reference Library for Swiss Butterflies and Forester Moths as a Tool for Species Identification, Systematics and Conservation. PLoS ONE 2018, 13, e0208639. [Google Scholar] [CrossRef] [PubMed]
- Bezeng, B.S.; Van Der Bank, H.F. DNA Barcoding of Southern African Crustaceans Reveals a Mix of Invasive Species and Potential Cryptic Diversity. PLoS ONE 2019, 14, e0222047. [Google Scholar] [CrossRef] [Green Version]
- Del-Prado, R.; Buaruang, K.; Lumbsch, H.T.; Crespo, A.; Divakar, P.K. DNA Sequence-Based Identification and Barcoding of a Morphologically Highly Plastic Lichen Forming Fungal Genus (Parmotrema, Parmeliaceae) from the Tropics. Bryologist 2019, 122, 281–291. [Google Scholar] [CrossRef]
- Faith, D.P. Conservation Evaluation and Phylogenetic Diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Faith, D.P. Phylogenetic Pattern and the Quantification of Organismal Biodiversity. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 1994, 345, 45–58. [Google Scholar] [CrossRef]
- Miller, J.T.; Jolley-Rogers, G.; Mishler, B.D.; Thornhill, A. Phylogenetic Diversity Is a Better Measure of Biodiversity than Taxon Counting. J. Syst. Evol. 2018, 56, 663–667. [Google Scholar] [CrossRef] [Green Version]
- Steinke, D.; de Waard, S.L.; Sones, J.E.; Ivanova, N.V.; Prosser, S.W.J.; Perez, K.; Braukmann, T.W.A.; Milton, M.; Zakharov, E.V.; de Waard, J.R.; et al. Message in a Bottle–Metabarcoding Enables Biodiversity Comparisons Across Ecoregions. bioRxiv 2021. [Google Scholar] [CrossRef]
- Winter, M.; Devictor, V.; Schweiger, O. Phylogenetic Diversity and Nature Conservation: Where Are We? Trends Ecol. Evol. 2013, 28, 199–204. [Google Scholar] [CrossRef]
- Flynn, D.F.B.; Mirotchnick, N.; Jain, M.; Palmer, M.I.; Naeem, S. Functional and Phylogenetic Diversity as Predictors of Biodiversity- Ecosystem-Function Relationships. Ecology 2011, 92, 1573–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazel, F.; Mooers, A.; Riva, G.V.D.; Pennell, M.W. Conserving Phylogenetic Diversity Can Be a Poor Strategy for Conserving Functional Diversity. Biol. Sci. 2017, 66, 1019–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazel, F.; Pennell, M.W.; Cadotte, M.W.; Diaz, S.; Riva, V.D.; Grenyer, R.; Leprieur, F.; Mooers, A.O.; Mouillot, D.; Tucker, C.M.; et al. Prioritizing Phylogenetic Diversity Captures Functional Diversity Unreliably. Nat. Commun. 2018, 9, 2888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, M.A.; Roger, A.; Courtois, E.A.; Jabot, F.; Norden, N.; Paine, C.E.T.; Baraloto, C.; Thébaud, C.; Chave, J. Shifts in Species and Phylogenetic Diversity between Sapling and Tree Communities Indicate Negative Density Dependence in a Lowland Rain Forest. J. Ecol. 2010, 98, 137–146. [Google Scholar] [CrossRef]
- Kress, W.J.; Erickson, D.L.; Swenson, N.G.; Thompson, J.; Uriarte, M.; Zimmerman, J.K. Advances in the Use of DNA Barcodes to Build a Community Phylogeny for Tropical Trees in a Puerto Rican Forest Dynamics Plot. PLoS ONE 2010, 5, e15409. [Google Scholar] [CrossRef] [Green Version]
- Pei, N.; Erickson, D.; Chen, B.; Ge, X.; Mi, X.; Swenson, M.G.; Zhang, J.-L.; Jones, F.A.; Huang, C.L.; Ye, W.; et al. Closely-Related Taxa Influence Woody Species Discrimination via DNA Barcoding: Evidence from Global Forest Dynamics Plots. Sci. Rep. 2015, 5, 15127. [Google Scholar] [CrossRef] [Green Version]
- Whitfeld, T.J.S.; Kress, W.J.; Erickson, D.L.; Weiblen, G.D. Change in Community Phylogenetic Structure during Tropical Forest Succession: Evidence from New Guinea. Ecography 2012, 35, 821–830. [Google Scholar] [CrossRef]
- Swenson, N.G. Phylogenetic Analyses of Ecological Communities Using DNA Barcode Data. Methods Mol. Biol. 2012, 858, 409–419. [Google Scholar] [CrossRef]
- Schreeg, L.A.; Kress, W.J.; Erickson, D.L.; Swenson, N.G. Phylogenetic Analysis of Local-Scale Tree Soil Associations in a Lowland Moist Tropical Forest. PLoS ONE 2010, 5, e13685. [Google Scholar] [CrossRef] [Green Version]
- Westbrook, J.W.; Kitajima, K.; Burleigh, J.G.; Kress, W.J.; Erickson, D.L.; Wright, S.J. What Makes a Leaf Tough? Patterns of Correlated Evolutionbetween Leaf Toughness Traits and Demographic Rates among 197 Shade-Tolerant Woody Species in a Neotropical Forest. Am. Nat. 2011, 177, 800–811. [Google Scholar] [CrossRef]
- Martin, A.R.; Erickson, D.L.; Kress, W.J.; Thomas, S.C. Wood Nitrogen Concentrations in Tropical Trees: Phylogenetic Patterns and Ecological Correlates. New Phytol. 2014, 204, 484–495. [Google Scholar] [CrossRef]
- Mcmanus, K.M.; Asner, G.P.; Martin, R.E.; Dexter, K.G.; Kress, W.J.; Field, C.B.; Ustin, S.L.; Wynne, R.H.; Thenkabail, P.S. Phylogenetic Structure of Foliar Spectral Traits in Tropical Forest Canopies. Remote Sens. 2016, 8, 196. [Google Scholar] [CrossRef] [Green Version]
- Chauvin, K.M.M.; Asner, G.P.; Martin, R.E.; Kress, W.J.; Wright, S.J.; Field, C.B. Decoupled Dimensions of Leaf Economic and Anti-Herbivore Defense Strategies in a Tropical Canopy Tree Community. Oecologia 2018, 186, 765–782. [Google Scholar] [CrossRef]
- Swenson, N.G. The Assembly of Tropical Tree Communities—The Advances and Shortcomings of Phylogenetic and Functional Trait Analyses. Ecography 2013, 36, 264–276. [Google Scholar] [CrossRef]
- Jones, F.A.; Erickson, D.L.; Bernal, M.A.; Bermingham, E.; Kress, W.J.; Herre, E.A.; Muller-Landau, H.C.; Turner, B.L. The Roots of Diversity: Below Ground Species Richness and Rooting Distributions in a Tropical Forest Revealed by DNA Barcodes and Inverse Modeling. PLoS ONE 2011, 6, e24506. [Google Scholar] [CrossRef] [PubMed]
- Basset, Y.; Donoso, D.A.; Hajibabaei, M.; Wright, M.T.G.; Perez, K.H.J.; Lamarre, G.P.A.; De León, L.F.; Palacios-Vargas, J.G.; Castaño-Meneses, G.; Rivera, M.; et al. Methodological Considerations for Monitoring Soil/Litter Arthropods in Tropical Rainforests Using DNA Metabarcoding, with a Special Emphasis on Ants, Springtails. Metabarcoding Metagenomics 2020, 4, 151–163. [Google Scholar] [CrossRef]
- Baker, T.R.; Pennington, R.T.; Dexter, K.G.; Fine, P.V.A.; Fortune-Hopkins, H.; Honorio, E.N.; Huamantupa-Chuquimaco, I.; Klitgård, B.B.; Lewis, G.P.; de Lima, H.C.; et al. Maximising Synergy among Tropical Plant Systematists, Ecologists, and Evolutionary Biologists. Trends Ecol. Evol. 2017, 32, 258–267. [Google Scholar] [CrossRef] [Green Version]
- Pei, N.; Lian, J.Y.; Erickson, D.L.; Swenson, N.G.; Kress, W.J.; Ye, W.H.; Ge, X.J. Exploring Tree-Habitat Associations in a Chinese Subtropical Forest Plot Using a Molecular Phylogeny Generated from DNA Barcode Loci. PLoS ONE 2011, 6, e21273. [Google Scholar] [CrossRef]
- de Oliveira, A.A.; Vicentini, A.; Chave, J.; Castanho, C.D.T.; Davies, S.J.; Martini, A.M.Z.; Lima, R.A.F.; Ribeiro, R.R.; Iribar, A.; Souza, V.C. Habitat Specialization and Phylogenetic Structure of Tree Species in a Coastal Brazilian White-Sand Forest. J. Plant Ecol. 2017, 7, 134–144. [Google Scholar] [CrossRef] [Green Version]
- Mi, X.; Swenson, N.G.; Valencia, R.; John Kress, W.; Erickson, D.L.; Pérez, Á.J.; Ren, H.; Su, S.H.; Gunatilleke, N.; Gunatilleke, S.; et al. The Contribution of Rare Species to Community Phylogenetic Diversity across a Global Network of Forest Plots. Am. Nat. 2012, 180, E17–E30. [Google Scholar] [CrossRef] [Green Version]
- Muscarella, R.; Uriarte, M.; Erickson, D.L.; Swenson, N.G.; Zimmerman, J.K.; Kress, W.J. A Well-Resolved Phylogeny of the Trees of Puerto Rico Based on DNA Barcode Sequence Data. PLoS ONE 2014, 9, e112843. [Google Scholar] [CrossRef]
- Muscarella, R.; Uriarte, M.; Erickson, D.; Swenson, N.; Kress, W.; Zimmerman, J. Variation of Tropical Forest Assembly Processes across Regional Environmental Gradients; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar] [CrossRef] [Green Version]
- Wills, C.; Harms, K.E.; Wiegand, T.; Punchi-Manage, R.; Gilbert, G.S.; Erickson, D.; Kress, W.J.; Hubbell, S.P.; Gunatilleke, C.V.S.; Gunatilleke, I.A.U.N. Persistence of Neighborhood Demographic Influences over Long Phylogenetic Distances May Help Drive Post-Speciation Adaptation in Tropical Forests. PLoS ONE 2016, 11, e0156913. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.A.; Eveleigh, E.S.; McCann, K.S.; Merilo, M.T.; McCarthy, P.C.; van Rooyen, K.I. Barcoding a Quantified Food Web: Crypsis, Concepts, Ecology and Hypotheses. PLoS ONE 2011, 6, e14424. [Google Scholar] [CrossRef] [PubMed]
- Navarro, S.P.; Jurado-Rivera, J.A.; GãMez-Zurita, J.; Lyal, C.H.C.; Vogler, A.P.; Jurado-Rivera, J.A.; Gómez-Zurita, J. DNA Profiling of Host–Herbivore Interactions in Tropical Forests. Ecol. Èntomol. 2010, 35, 18–32. [Google Scholar] [CrossRef]
- Kishimoto-Yamada, K.; Kamiya, K.; Meleng, P.; Diway, B.; Kaliang, H.; Chong, L.; Itioka, T.; Sakai, S.; Ito, M. Wide Host Ranges of Herbivorous Beetles? Insights from DNA Bar Coding. PLoS ONE 2013, 8, e74426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Robledo, C.; Erickson, D.L.; Staines, C.L.; Erwin, T.L.; Kress, W.J. Tropical Plant-Herbivore Networks: Reconstructing Species Interactions Using DNA Barcodes. PLoS ONE 2013, 8, e52967. [Google Scholar] [CrossRef]
- Strong, D.R., Jr. Rolled-Leaf Hispine Beetles (Chrysomelidae) and Their Zingiberales Host Plants in Middle America. JSTOR 1977, 9, 156–169. [Google Scholar] [CrossRef]
- Palmer, C.M.; Wershoven, N.L.; Martinson, S.J.; ter Hofstede, H.M.; Kress, W.J.; Symes, L.B. Patterns of Herbivory in Neotropical Forest Katydids as Revealed by DNA Barcoding of Digestive Tract Contents. Diversity 2022, 14, 120. [Google Scholar] [CrossRef]
- Colwell, R.K. Competition and Coexistence in a Simple Tropical Community. Am. Nat. 1973, 107, 737–760. [Google Scholar] [CrossRef]
- Futuyma, D.J.; Moreno, G. The Evolution of Ecological Specialization. Annu. Rev. Ecol. Syst. 1988, 19, 207–233. [Google Scholar] [CrossRef]
- Bizzarri, L.; Baer, C.S.; García-Robledo, C. DNA Barcoding Reveals Generalization and Host Overlap in Hummingbird Flower Mites: Implications for the Mating Rendezvous Hypothesis. Am. Nat. 2022. ahead of print. [Google Scholar] [CrossRef]
- Kartzinel, T.; Chen, P.; Coverdale, T. DNA Metabarcoding Illuminates Dietary Niche Partitioning by African Large Herbivores. Proc. Natl. Acad. Sci. USA 2015, 112, 8019–8024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, P.T.; Ang’ila, R.O.; Kimuyu, D.; Musili, P.M.; Ken-fack, D.; Lokeny Etelej, P.; Magid, M.; Gill, B.A.; Kartzinel, T.R. Opposing Gradients in the Abundance and Diversity of Plants and Large Herbivores Revealed with DNA Barcoding in a Semi-Arid African Savanna. Diversity 2022, 14. [Google Scholar]
- González-Varo, J.P.; Arroyo, J.; Jordano, P. Who Dispersed the Seeds? The Use of DNA Barcoding in Frugivory and Seed Dispersal Studies. Methods Ecol. Evol. 2014, 5, 806–814. [Google Scholar] [CrossRef]
- Baker, C.C.M.; Bittleston, L.S.; Sanders, J.G.; Pierce, N.E. Dissecting Host-Associated Communities with DNA Barcodes. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150328. [Google Scholar] [CrossRef] [Green Version]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Fungal Barcoding Consortium, F.B.; Fungal Barcoding Consortium Author List, F.B.C.A.; Bolchacova, E.; et al. Nuclear Ribosomal Internal Transcribed Spacer (ITS) Region as a Universal DNA Barcode Marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [Green Version]
- Lücking, R.; Aime, M.C.; Robbertse, B.; Miller, A.N.; Ariyawansa, H.A.; Aoki, T.; Cardinali, G.; Crous, P.W.; Druzhinina, I.S.; Geiser, D.M.; et al. Unambiguous Identification of Fungi: Where Do We Stand and How Accurate and Precise Is Fungal DNA Barcoding? IMA Fungus 2020, 11, 14. [Google Scholar] [CrossRef]
- Young, M.R.; Proctor, H.C.; Dewaard, J.R.; Hebert, P.D.N. DNA Barcodes Expose Unexpected Diversity in Canadian Mites. Mol. Ecol. 2019, 28, 5347–5359. [Google Scholar] [CrossRef]
- Bonato, K.O.; Silva, P.C.; Malabarba, L.R. Unrevealing Parasitic Trophic Interactions-A Molecular Approach for Fluid-Feeding Fishes. Front. Ecol. Evol. 2018, 6, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Keet, J.-H.; Ellis, A.G.; Hui, C.; Roux, J.J.L. Legume–Rhizobium Symbiotic Promiscuity and Effectiveness Do Not Affect Plant Invasiveness. Ann. Bot. 2017, 119, 1319–1331. [Google Scholar] [CrossRef]
- Smith, H.B.; Dal Grande, F.; Muggia, L.; Keuler, R.; Divakar, P.K.; Grewe, F.; Schmitt, I.; Lumbsch, H.T.; Leavitt, S.D. Metagenomic Data Reveal Diverse Fungal and Algal Communities Associated with the Lichen Symbiosis. Symbiosis 2020, 82, 133–147. [Google Scholar] [CrossRef]
- Taberlet, P.; Coissac, E.; Pompanon, F. Towards Next-generation Biodiversity Assessment Using DNA Metabarcoding. Molecular 2012, 21, 2045–2050. [Google Scholar] [CrossRef] [PubMed]
- Widmer, A.; Cozzolino, S.; Pellegrino, G.; Soliva, M.; Dafni, A. Molecular Analysis of Orchid Pollinaria and Pollinaria-Remains Found on Insects. Mol. Ecol. 2000, 9, 1911–1914. [Google Scholar] [CrossRef] [PubMed]
- Sickel, W.; Ankenbrand, M.J.; Grimmer, G.; Holzschuh, A.; Härtel, S.; Lanzen, J.; Steffan-Dewenter, I.; Keller, A. Increased Efficiency in Identifying Mixed Pollen Samples by Meta-Barcoding with a Dual-Indexing Approach. BMC Ecol. 2015, 15, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowe, A.; Jones, L.; Witter, L.; Creer, S.; de Vere, N. Using DNA Metabarcoding to Identify Floral Visitation by Pollinators. Diversity 2022, 14. [Google Scholar]
- Clare, E.L.; Schiestl, F.P.; Leitch, A.R.; Chittka, L. The Promise of Genomics in the Study of Plant-Pollinator Interactions. Genome Biol. 2013, 14, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Soininen, E.M.; Valentini, A.; Coissac, E.; Miquel, C.; Gielly, L.; Brochmann, C.; Brysting, A.K.; Sønstebø, J.H.; Ims, R.A.; Yoccoz, N.G.; et al. Analysing Diet of Small Herbivores: The Efficiency of DNA Barcoding Coupled with High-Throughput Pyrosequencing for Deciphering the Composition of Complex Plant Mixtures. Front. Zool. 2009, 6, 16. [Google Scholar] [CrossRef] [Green Version]
- de Vere, N.; Rich, T.C.G.; Ford, C.R.; Trinder, S.A.; Long, C.; Moore, C.W.; Satterthwaite, D.; Davies, H.; Allainguillaume, J.; Ronca, S.; et al. DNA Barcoding the Native Flowering Plants and Conifers of Wales. PLoS ONE 2012, 7, e37945. [Google Scholar] [CrossRef] [Green Version]
- Bell, K.L.; Batchelor, K.L.; Bradford, M.; McKeown, A.; MacDonald, S.L.; Westcott, D. Optimisation of a Pollen DNA Metabarcoding Method for Diet Analysis of Flying-Foxes (Pteropus Spp.). Aust. J. Zool. 2021, 68, 273–284. [Google Scholar] [CrossRef]
- de Vere, N.; Jones, L.E.; Gilmore, T.; Moscrop, J.; Lowe, A.; Smith, D.; Hegarty, M.J.; Creer, S.; Ford, C.R. Using DNA Metabarcoding to Investigate Honey Bee Foraging Reveals Limited Flower Use despite High Floral Availability. Sci. Rep. 2017, 7, 42838. [Google Scholar] [CrossRef] [Green Version]
- Jones, L.; Brennan, G.L.; Lowe, A.; Creer, S.; Ford, C.R.; de Vere, N. Shifts in Honeybee Foraging Reveal Historical Changes in Floral Resources. Commun. Biol. 2021, 4, 37. [Google Scholar] [CrossRef] [PubMed]
- Richardson, R.T.; Lin, C.-H.; Sponsler, D.B.; Quijia, J.O.; Goodell, K.; Johnson, R.M. Application of ITS2 Metabarcoding to Determine the Provenance of Pollen Collected by Honey Bees in an Agroecosystem. Appl. Plant Sci. 2015, 3, 1400066. [Google Scholar] [CrossRef] [PubMed]
- Bell, K.L.; Burgess, K.S.; Botsch, J.C.; Dobbs, E.K.; Read, T.D.; Brosi, B.J. Quantitative and Qualitative Assessment of Pollen DNA Metabarcoding Using Constructed Species Mixtures. Mol. Ecol. 2019, 28, 431–455. [Google Scholar] [CrossRef] [PubMed]
- Harper, L.R.; Niemiller, M.L.; Benito, J.B.; Paddock, L.E. BeeDNA: Microfluidic Environmental DNA Metabarcoding as a Tool for Connecting Plant and Pollinator Communities. bioRxiv 2021. [Google Scholar] [CrossRef]
- Madden, M.J.L.; Young, R.G.; Brown, J.W.; Miller, S.E.; Frewin, A.J.; Hanner, R.H. Using DNA Barcoding to Improve Invasive Pest Identification at U.S. Ports-of-Entry. PLoS ONE 2019, 14, e0222291. [Google Scholar] [CrossRef]
- Whitehurst, L.E.; Cunard, C.E.; Reed, J.N.; Worthy, S.J.; Marsico, T.D.; Lucardi, R.D.; Burgess, K.S. Preliminary Application of DNA Barcoding toward the Detection of Viable Plant Propagules at an Initial, International Point-of-Entry in Georgia, USA. Biol. Invasions 2020, 22, 1585–1606. [Google Scholar] [CrossRef]
- Krishnamurthy, P.K.; Francis, R.A. A Critical Review on the Utility of DNA Barcoding in Biodiversity Conservation. Springer 2012, 21, 1901–1919. [Google Scholar] [CrossRef]
- Liu, M.; Zhao, Y.; Sun, Y.; Li, Y.; Wu, P.; Zhou, S.; Ren, L. Comparative Study on Diatom Morphology and Molecular Identification in Drowning Cases. Forensic Sci. Int. 2020, 317, 110552. [Google Scholar] [CrossRef]
- Bell, K.L.; Burgess, K.S.; Okamoto, K.C.; Aranda, R.; Brosi, B.J. Review and Future Prospects for DNA Barcoding Methods in Forensic Palynology. Forensic Sci. Int. Genet. 2016, 21, 110–116. [Google Scholar] [CrossRef]
- Liu, J.; Milne, R.I.; Möller, M.; Zhu, G.F.; Ye, L.J.; Luo, Y.H.; Yang, J.B.; Wambulwa, M.C.; Wang, C.N.; Li, D.Z.; et al. Integrating a Comprehensive DNA Barcode Reference Library with a Global Map of Yews (Taxus L.) for Forensic Identification. Mol. Ecol. Resour. 2018, 18, 1115–1131. [Google Scholar] [CrossRef]
- Lees, D.C.; Lack, H.W.; Rougerie, R.; Hernandez-Lopez, A.; Raus, T.; Avtzis, N.D.; Augustin, S.; Lopez-Vaamonde, C. Tracking Origins of Invasive Herbivores through Herbaria and Archival DNA: The Case of the Horse-Chestnut Leaf Miner. Front. Ecol. Environ. 2011, 9, 322–328. [Google Scholar] [CrossRef]
- Veneza, I.; Silva, R.; Freitas, L.; Silva, S.; Martins, K.; Sampaio, I.; Schneider, H.; Gomes, G. Molecular Authentication of Pargo Fillets Lutjanus Purpureus (Perciformes: Lutjanidae) by DNA Barcoding Reveals Commercial Fraud. Neotrop. Ichthyol. 2018, 16, 1–6. [Google Scholar] [CrossRef]
- Muellner, A.N.; Schaefer, H.; Lahaye, R. Evaluation of Candidate DNA Barcoding Loci for Economically Important Timber Species of the Mahogany Family (Meliaceae). Mol. Ecol. Resour. 2011, 11, 450–460. [Google Scholar] [CrossRef]
- Nithaniyal, S.; Newmaster, S.G.; Ragupathy, S.; Krishnamoorthy, D.; Vassou, S.L.; Parani, M. DNA Barcode Authentication of Wood Samples of Threatened and Commercial Timber Trees within the Tropical Dry Evergreen Forest of India. PLoS ONE 2014, 9, e107669. [Google Scholar] [CrossRef] [PubMed]
- Bolson, M.; De Camargo Smidt, E.; Brotto, M.L.; Silva-Pereira, V. ITS and TrnH-PsbA as Efficient DNA Barcodes to Identify Threatened Commercial Woody Angiosperms from Southern Brazilian Atlantic Rainforests. PLoS ONE 2015, 10, e0143049. [Google Scholar] [CrossRef] [PubMed]
- Hassold, S.; Lowry, P.P.; Bauert, M.R.; Razafintsalama, A.; Ramamonjisoa, L.; Widmer, A. DNA Barcoding of Malagasy Rosewoods: Towards a Molecular Identification of CITES-Listed Dalbergia Species. PLoS ONE 2016, 11, e0157881. [Google Scholar] [CrossRef] [Green Version]
- Lahaye, R.; Van Der Bank, M.; Bogarin, D.; Warner, J.; Pupulin, F.; Gigot, G.; Maurin, O.; Duthoit, S.; Barraclough, T.G.; Savolainen, V. DNA Barcoding the Floras of Biodiversity Hotspots. Proc. Natl. Acad Sci. USA 2008, 105, 2923–2928. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, A.; Rothbart, A.; Frankham, G.; Johnson, R.N.; Neaves, L.E. Could Do Better! A High School Market Survey of Fish Labelling in Sydney, Australia, Using DNA Barcodes. PeerJ 2019, 7, e7138. [Google Scholar] [CrossRef]
- Siozios, S.; Massa, A.; Parr, C.L.; Verspoor, R.L.; Hurst, G.D.D. DNA Barcoding Reveals Incorrect Labelling of Insects Sold as Food in the UK. PeerJ 2020, 8, e8496. [Google Scholar] [CrossRef]
- Thongkhao, K.; Pongkittiphan, V.; Phadungcharoen, T.; Tungphatthong, C.; Urumarudappa, S.K.J.; Pengsuparp, T.; Sutanthavibul, N.; Wiwatcharakornkul, W.; Kengtong, S.; Sukrong, S. Differentiation of Cyanthillium Cinereum, a Smoking Cessation Herb, from Its Adulterant Emilia Sonchifolia Using Macroscopic and Microscopic Examination, HPTLC Profiles and DNA Barcodes. Sci. Rep. 2020, 10, 14753. [Google Scholar] [CrossRef]
- Staats, M.; Arulandhu, A.J.; Gravendeel, B.; Holst-Jensen, A.; Scholtens, I.; Peelen, T.; Prins, T.W.; Kok, E. Advances in DNA Metabarcoding for Food and Wildlife Forensic Species Identification. Anal. Bioanal. Chem. 2016, 408, 4615–4630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, L.; Yu, M.; Wiedenhoeft, A.C.; He, T.; Li, J.; Liu, B.; Jiang, X.; Yin, Y. DNA Barcode Authentication and Library Development for the Wood of Six Commercial Pterocarpus Species: The Critical Role of Xylarium Specimens. Sci. Rep. 2018, 8, 1945. [Google Scholar] [CrossRef] [PubMed]
- Newmaster, S.G.; Grguric, M.; Shanmughanandhan, D.; Ramalingam, S.; Ragupathy, S. DNA Barcoding Detects Contamination and Substitution in North American Herbal Products. BMC Med. 2013, 11, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Wu, X.; Liu, C.; Newmaster, S.; Ragupathy, S.; Kress, W.J. Progress in the Use of DNA Barcodes in the Identification and Classification of Medicinal Plants. Ecotoxicol. Environ. Saf. 2021, 208, 111691. [Google Scholar] [CrossRef] [PubMed]
- Stoeckle, M.; Gamble, C.; Kirpekar, R.; Young, G.; Ahmed, S.; Little, D.P. Commercial Teas Highlight Plant DNA Barcode Identification Successes and Obstacles. Sci. Rep. 2011, 1, 42. [Google Scholar] [CrossRef] [PubMed]
- Shapcott, A.; Liu, Y.; Howard, M.; Forster, P.; Kress, W.J.; Erickson, D.; Shimizu, Y.; McDonald, W. Comparing floristic diversity and conservation priorities across south east Queensland regional rain forest ecosystems using phylodiversity indexes. Int. J. Plant Sci. 2017, 178, 211–229. [Google Scholar] [CrossRef]
- De Boer, H.J.; Ouarghidi, A.; Martin, G.; Abbad, A.; Kool, A. DNA Barcoding Reveals Limited Accuracy of Identifications Based on Folk Taxonomy. PLoS ONE 2014, 9, e84291. [Google Scholar] [CrossRef]
- Pimentel, D.; Zuniga, R.; Morrison, D. Update on the Environmental and Economic Costs Associated with Alien-Invasive Species in the United States. Ecol. Econ. 2004, 52, 273–288. [Google Scholar] [CrossRef]
- Comtet, T.; Sandionigi, A.; Viard, F.; Casiraghi, M. DNA (Meta)Barcoding of Biological Invasions: A Powerful Tool to Elucidate Invasion Processes and Help Managing Aliens. Biol. Invasions 2015, 17, 905–922. [Google Scholar] [CrossRef]
- Ardura, A.; Zaiko, A.; Borrell, Y.J.; Samuiloviene, A.; Garcia-Vazquez, E. Novel Tools for Early Detection of a Global Aquatic Invasive, the Zebra Mussel Dreissena Polymorpha. Aquat. Conserv. Mar. Freshw. Ecosyst. 2017, 27, 165–176. [Google Scholar] [CrossRef]
- Smith, M.A.; Fisher, B.L. Invasions, DNA Barcodes, and Rapid Biodiversity Assessment Using Ants of Mauritius. Front. Zool. 2009, 6, 31. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.; Wu, H.; Zhu, X.; Lin, J. Species Identification of Conyza Bonariensis Assisted by Chloroplast Genome Sequencing. Front. Genet. 2018, 9, 374. [Google Scholar] [CrossRef] [Green Version]
- Agnarsson, I.; Biology, M.K.-S. Taxonomy in a Changing World: Seeking Solutions for a Science in Crisis. Syst. Biol. 2007, 56, 531–539. [Google Scholar] [CrossRef] [Green Version]
- Gomes, A.C.; Andrade, A.; Barreto-Silva, J.S.; Brenes-Arguedas, T.; López, D.C.; Cde Freitas, C.; Lang, C.; de Oliveira, A.A.; Pérez, A.J.; Perez, R.; et al. Local Plant Species Delimitation in a Highly Diverse A Mazonian Forest: Do We All See the Same Species? J. Veg. Sci. 2013, 24, 70–79. [Google Scholar] [CrossRef]
- Owen, N.R.; Gumbs, R.; Gray, C.L.; Faith, D.P. Phylogenetic Diversity and Conservation. Nat. Commun. 2019, 10, 859. [Google Scholar] [CrossRef] [Green Version]
- Howard, M.G.; McDonald, W.J.F.; Forster, P.I.; Kress, W.J.; Erickson, D.; Faith, D.P.; Shapcott, A. Patterns of Phylogenetic Diversity of Subtropical Rainforest of the Great Sandy Region, Australia Indicate Long Term Climatic Refugia. PLoS ONE 2016, 11, e0153565. [Google Scholar] [CrossRef] [Green Version]
- Humphreys, A.M.; Vorontsova, M.; Govaerts, R.; Ficinski, S.Z.; Lughadha, N.; Vorontsova, M.S. Global Dataset Shows Geography and Life Form Predict Modern Plant Extinction and Rediscovery. Nat. Ecol. Evol. 2019, 3, 1043–1047. [Google Scholar] [CrossRef]
- Costion, C.M.; Kress, W.J.; Crayn, D.M. DNA Barcodes Confirm the Taxonomic and Conservation Status of a Species of Tree on the Brink of Extinction in the Pacific. PLoS ONE 2016, 11, e0155118. [Google Scholar] [CrossRef]
- Ji, Y.; Ashton, L.; Pedley, S.M.; Edwards, D.P.; Tang, Y.; Nakamura, A.; Kitching, R.; Dolman, P.M.; Woodcock, P.; Edwards, F.A.; et al. Reliable, Verifiable and Efficient Monitoring of Biodiversity via Metabarcoding. Ecol. Lett. 2013, 16, 1245–1257. [Google Scholar] [CrossRef]
- Procopio, N.; Ghignone, S.; Voyron, S.; Chiapello, M.; Williams, A.; Chamberlain, A.; Mello, A.; Buckley, M. Soil Fungal Communities Investigated by Metabarcoding Within Simulated Forensic Burial Contexts. Front. Microbiol. 2020, 11, 1686. [Google Scholar] [CrossRef]
- Rivera, S.F.; Vasselon, V.; Jacquet, S.; Bouchez, A.; Ariztegui, D.; Rimet, F. Metabarcoding of Lake Benthic Diatoms: From Structure Assemblages to Ecological Assessment. Hydrobiologia 2018, 807, 37–51. [Google Scholar] [CrossRef]
- Leray, M.; Knowlton, N. DNA Barcoding and Metabarcoding of Standardized Samples Reveal Patterns of Marine Benthic Diversity. Proc. Natl. Acad. Sci. USA 2015, 112, 2076–2081. [Google Scholar] [CrossRef] [Green Version]
- Aguayo, J.; Fourrier-Jeandel, C.; Husson, C.; Loos, R. Assessment of Passive Traps Combined with High-Throughput. Appl. Environ. Microbiol. 2018, 84, e02637-17. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.D.; Fokar, M.; Cox, R.D.; Barnes, M.A. Airborne Environmental DNA Metabarcoding Detects More Diversity, with Less Sampling Effort, than a Traditional Plant Community Survey. BMC Ecol. Evol. 2021, 21, 218. [Google Scholar] [CrossRef]
- Cowart, D.A.; Pinheiro, M.; Mouchel, O.; Maguer, M.; Grall, J.; Miné, J.; Arnaud-Haond, S. Metabarcoding Is Powerful yet Still Blind: A Comparative Analysis of Morphological and Molecular Surveys of Seagrass Communities. PLoS ONE 2015, 10, e0117562. [Google Scholar] [CrossRef] [Green Version]
- Pompanon, F.; Deagle, B.E.; Symondson, W.O.C.; Brown, D.S.; Jarman, S.N.; Taberlet, P. Who Is Eating What: Diet Assessment Using next Generation Sequencing. Mol. Ecol. 2012, 21, 1931–1950. [Google Scholar] [CrossRef] [Green Version]
- Goldberg, A.R.; Conway, C.J.; Tank, D.C.; Andrews, K.R.; Gour, D.S.; Waits, L.P. Diet of a Rare Herbivore Based on DNA Metabarcoding of Feces: Selection, Seasonality, and Survival. Ecol. Evol. 2020, 10, 7627–7643. [Google Scholar] [CrossRef]
- Chua, P.Y.S.; Crampton-Platt, A.; Lammers, Y.; Alsos, I.G.; Boessenkool, S.; Bohmann, K. Metagenomics: A Viable Tool for Reconstructing Herbivore Diet. Mol. Ecol. Resour. 2021, 21, 2249–2263. [Google Scholar] [CrossRef]
- Vasquez, A.A.; Mohiddin, O.; Li, Z.; Bonnici, B.L.; Gurdziel, K.; Ram, J.L. Molecular Diet Studies of Water Mites Reveal Prey Biodiversity. PLoS ONE 2021, 16, e0254598. [Google Scholar] [CrossRef]
- Leoro-Garzon, P.; Gonedes, A.J.; Olivera, I.E.; Tartar, A. Oomycete Metabarcoding Reveals the Presence of Lagenidium Spp. In Phytotelmata. PeerJ 2019, 7, e7903. [Google Scholar] [CrossRef]
- Trzebny, A.; Slodkowicz-Kowalska, A.; Becnel, J.J.; Sanscrainte, N.; Dabert, M. A New Method of Metabarcoding Microsporidia and Their Hosts Reveals High Levels of Microsporidian Infections in Mosquitoes (Culicidae). Mol. Ecol. Resour. 2020, 20, 1486–1504. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Braukmann, T.W.A.; Prosser, S.W.J.; Ratnasingham, S.; de Waard, J.R.; Ivanova, N.V.; Janzen, D.H.; Hallwachs, W.; Naik, S.; Sones, J.E.; et al. A Sequel to Sanger: Amplicon Sequencing That Scales. BMC Genomics 2018, 19, 219. [Google Scholar] [CrossRef] [Green Version]
- Parks, M.; Cronn, R.; Liston, A. Increasing Phylogenetic Resolution at Low Taxonomic Levels Using Massively Parallel Sequencing of Chloroplast Genomes. BMC Biol. 2009, 7, 84. [Google Scholar] [CrossRef] [Green Version]
- Kuo, L.-Y.; Li, F.-W.; Chiou, W.-L.; Wang, C.-N. First Insights into Fern MatK Phylogeny. Mol. Phylogenetics Evol. 2011, 59, 556–566. [Google Scholar] [CrossRef]
- McKain, M.R.; Johnson, M.G.; Uribe-Convers, S.; Eaton, D.; Yang, Y. Practical Considerations for Plant Phylogenomics. Appl. Plant Sci. 2018, 6, e1038. [Google Scholar] [CrossRef]
- Blanchet, S.; Prunier, J.G.; De Kort, H. Time to Go Bigger: Emerging Patterns in Macrogenetics. Trends Genet. 2017, 33, 579–580. [Google Scholar] [CrossRef]
- Leigh, D.M.; van Rees, C.B.; Millette, K.L.; Breed, M.F.; Schmidt, C.; Bertola, L.D.; Hand, B.K.; Hunter, M.E.; Jensen, E.L.; Kershaw, F.; et al. Opportunities and Challenges of Macrogenetic Studies. Nat. Rev. Genet. 2021, 22, 791–807. [Google Scholar] [CrossRef]
- Edwards, J.L. Research and Societal Benefits of the Global Biodiversity Information Facility. BioScience 2004, 54, 485–486. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very High Resolution Interpolated Climate Surfaces for Global Land Areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Vision, T. The Dryad Digital Repository: Published Evolutionary Data as Part of the Greater Data Ecosystem. Nat. Preced. 2010. [Google Scholar] [CrossRef]
- The International Nucleotide Sequence Database Collaboration. Available online: https://www.insdc.org (accessed on 26 January 2022).
- Kress, W.J.; Soltis, D.E.; Kersey, P.J.; Wegrzyn, J.L.; Leebens-Mack, J.H.; Gostel, M.R.; Liu, X.; Soltis, P.S. Green Plant Genomes: What We Know in an Era of Rapidly Expanding Opportunities. Proc. Natl. Acad Sci. USA 2022, 119, 2115640118. [Google Scholar] [CrossRef]



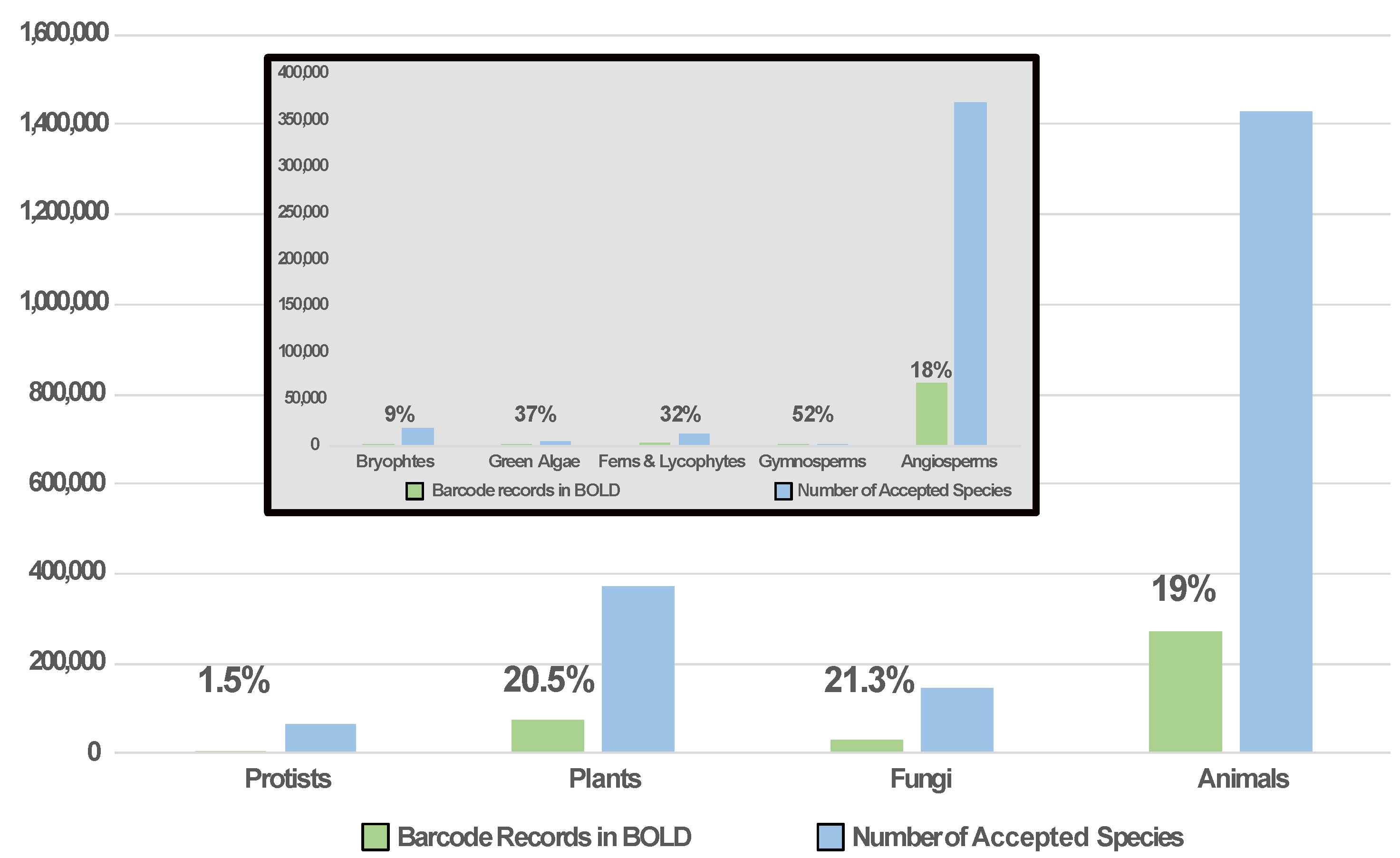{kind=link}
{kind=link}
| Taxon | Barcode Sequences Available 1 | |
|---|---|---|
| Animals | 11,607,692 | |
| Acanthocephala | 2302 | |
| Acoelomorpha | 20 | |
| Annelida | 112,010 | |
| Arthropoda | 11,486,730 | |
| Brachiopoda | 326 | |
| Bryozoa | 4529 | |
| Chaetognatha | 1775 | |
| Chordata | 877,866 | |
| Cnidaria | 32,680 | |
| Ctenophora | 649 | |
| Echinodermata | 326 | |
| Entoprocta | 76 | |
| Gastrotricha | 1351 | |
| Gnathostomulida | 24 | |
| Hemichordata | 263 | |
| Kinorhyncha | 720 | |
| Mollusca | 258,885 | |
| Nematoda | 36,513 | |
| Nematomorpha | 408 | |
| Nemertea | 6443 | |
| Onychophora | 1394 | |
| Phoronida | 172 | |
| Placozoa | 20 | |
| Platyhelminthes | 41,262 | |
| Porifera | 9668 | |
| Priapulida | 151 | |
| Rhombozoa | 48 | |
| Rotifera | 13,758 | |
| Sipuncula | 1367 | |
| Tardigrada | 3175 | |
| Xenacoelomorpha | 18 | |
| Fungi | 178,231 | |
| Ascomycota | 99,779 | |
| Basidiomycota | 71,120 | |
| Chytridiomycota | 293 | |
| Glomeromycota | 3529 | |
| Myxomycota | 235 | |
| Zygomycota | 3275 | |
| Plants | 572,154 | |
| Bryophyta | 22,675 | |
| Chlorophyta | 18,286 | |
| Lycopodiophyta | 1338 | |
| Magnoliophyta | 454,329 | |
| Pinophyta | 7661 | |
| Pteridophyta | 11,671 | |
| Rhodophyta | 56,194 | |
| Protists | 10,463 | |
| Chlorarachniophyta | 67 | |
| Ciliophora | 819 | |
| Heterokontophyta | 7238 | |
| Pyrrophycophyta | 2339 | |
| Total | 12,368,540 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gostel, M.R.; Kress, W.J. The Expanding Role of DNA Barcodes: Indispensable Tools for Ecology, Evolution, and Conservation. Diversity 2022, 14, 213. https://doi.org/10.3390/d14030213
Gostel MR, Kress WJ. The Expanding Role of DNA Barcodes: Indispensable Tools for Ecology, Evolution, and Conservation. Diversity. 2022; 14(3):213. https://doi.org/10.3390/d14030213
Chicago/Turabian StyleGostel, Morgan R., and W. John Kress. 2022. "The Expanding Role of DNA Barcodes: Indispensable Tools for Ecology, Evolution, and Conservation" Diversity 14, no. 3: 213. https://doi.org/10.3390/d14030213
APA StyleGostel, M. R., & Kress, W. J. (2022). The Expanding Role of DNA Barcodes: Indispensable Tools for Ecology, Evolution, and Conservation. Diversity, 14(3), 213. https://doi.org/10.3390/d14030213