
Biogeography and Diversification of Bumblebees (Hymenoptera: Apidae), with Emphasis on Neotropical Species

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Taxon and Locus Sampling
2.2. Phylogenetic Inference
2.3. Divergence Time Estimation
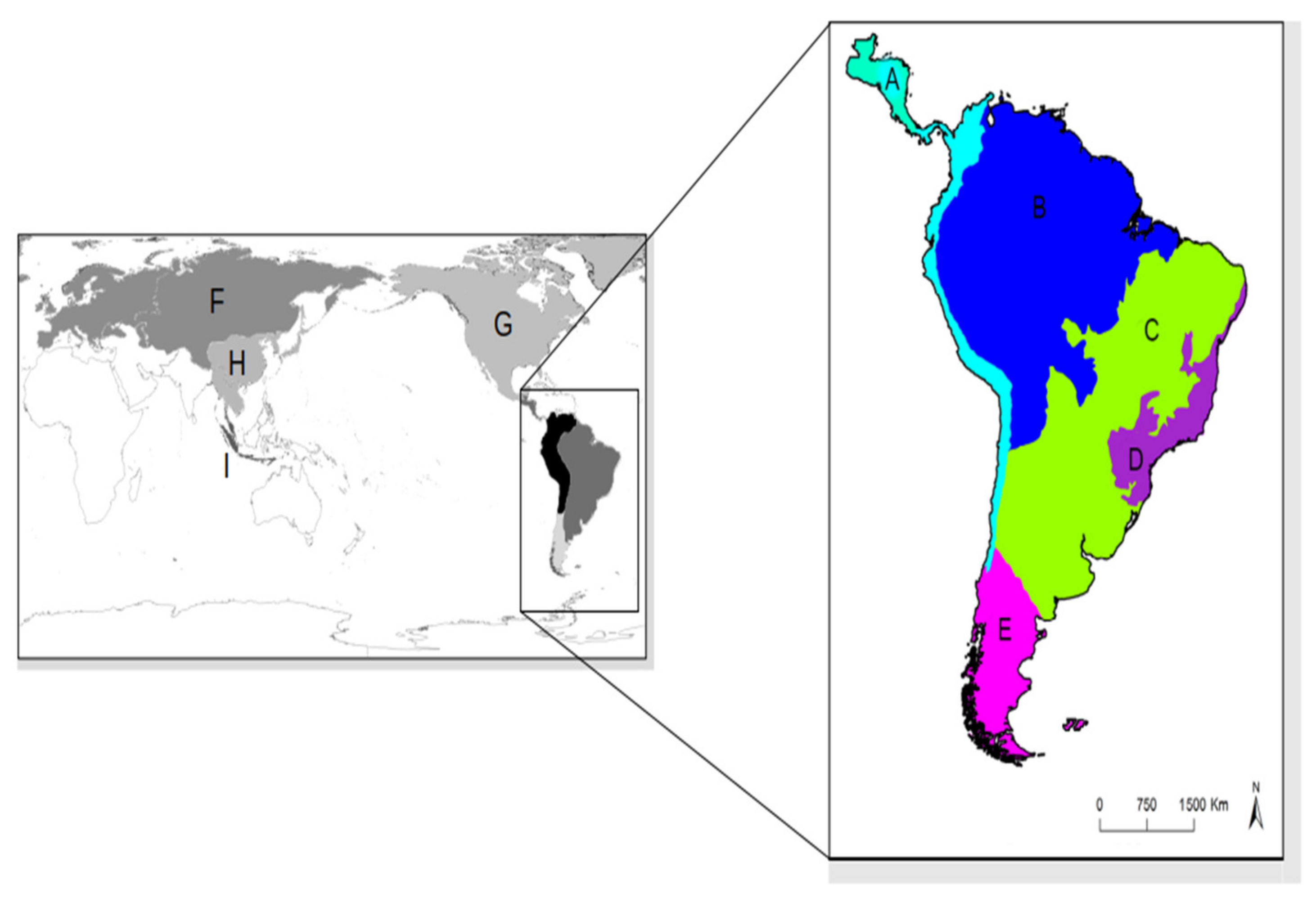
2.4. Biogeographic Analyses
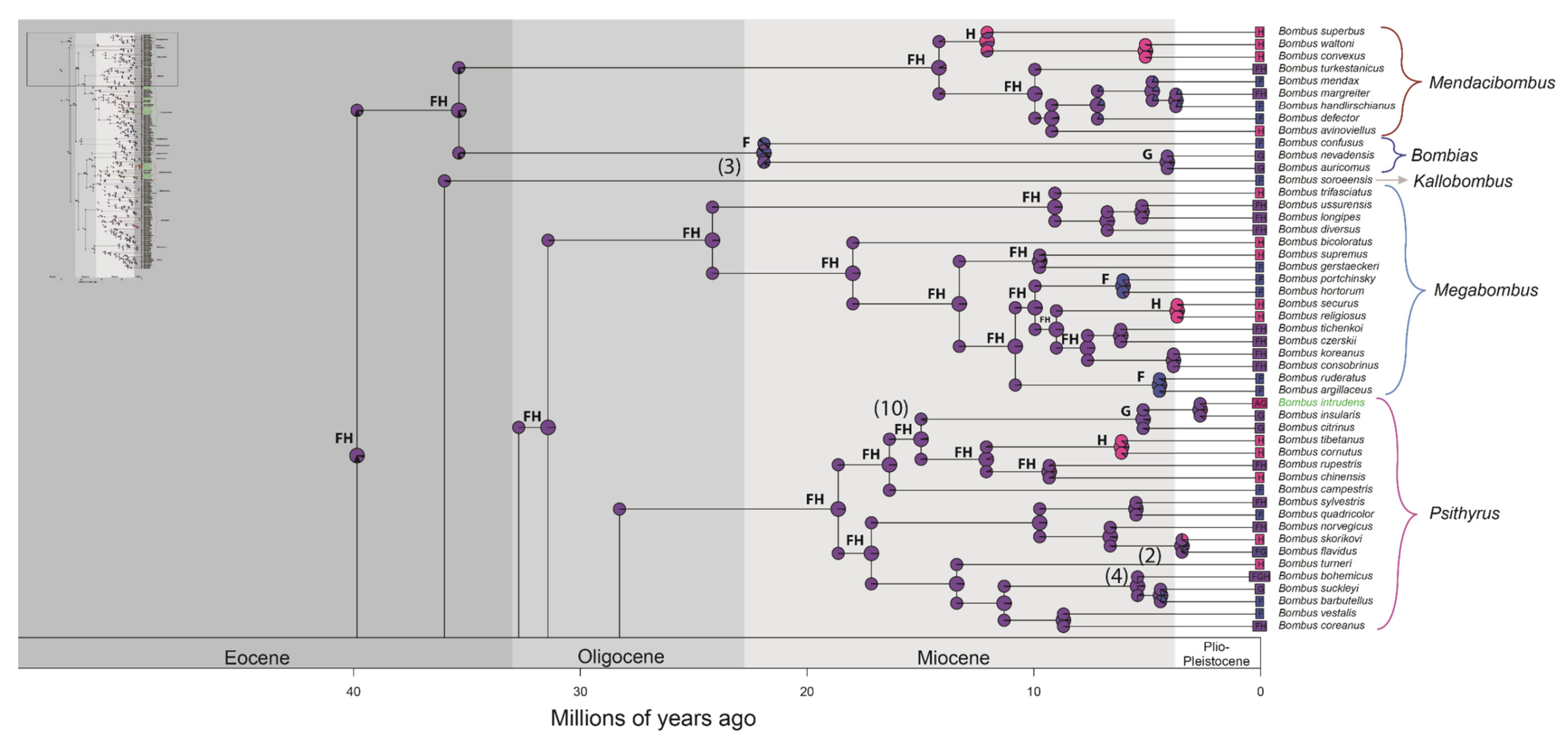
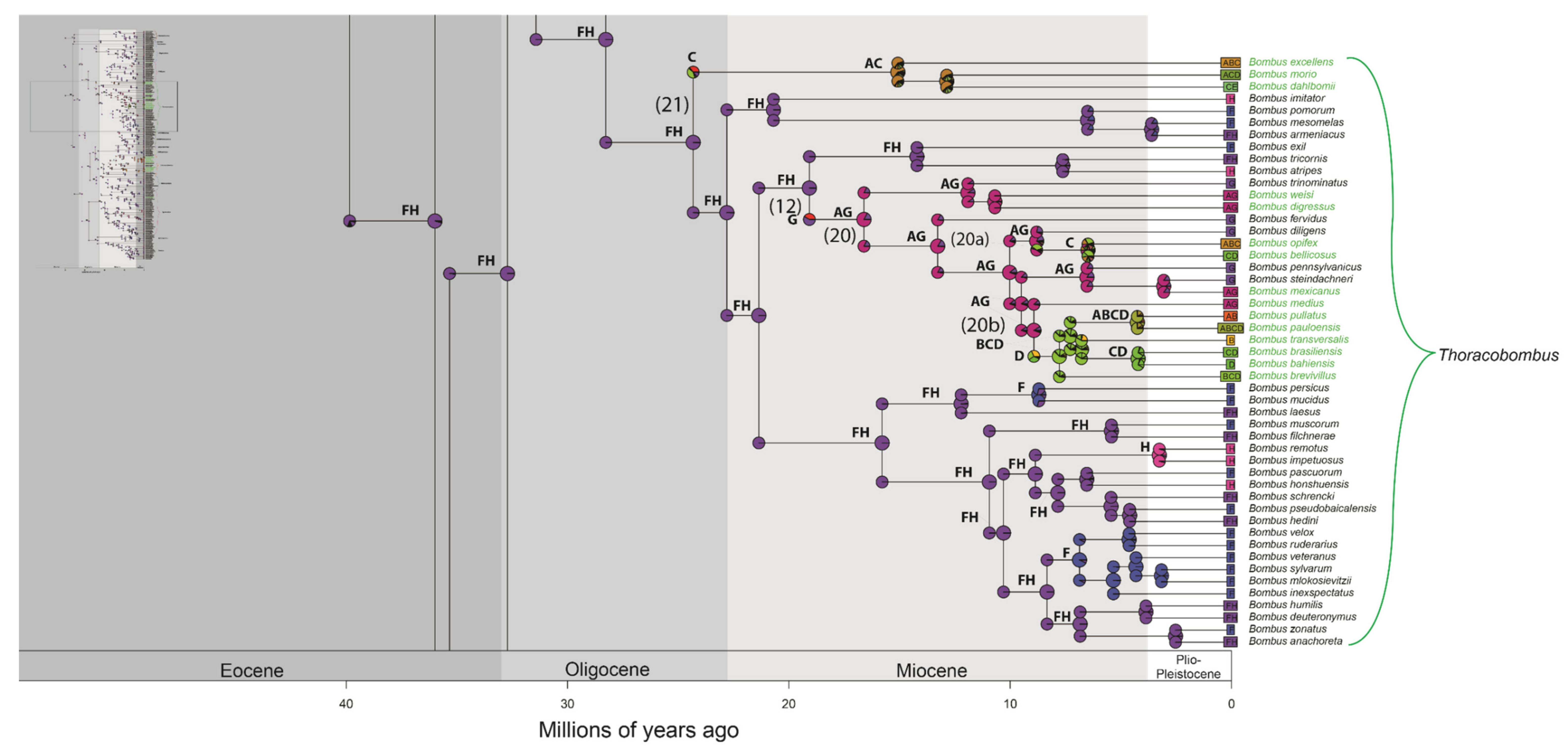
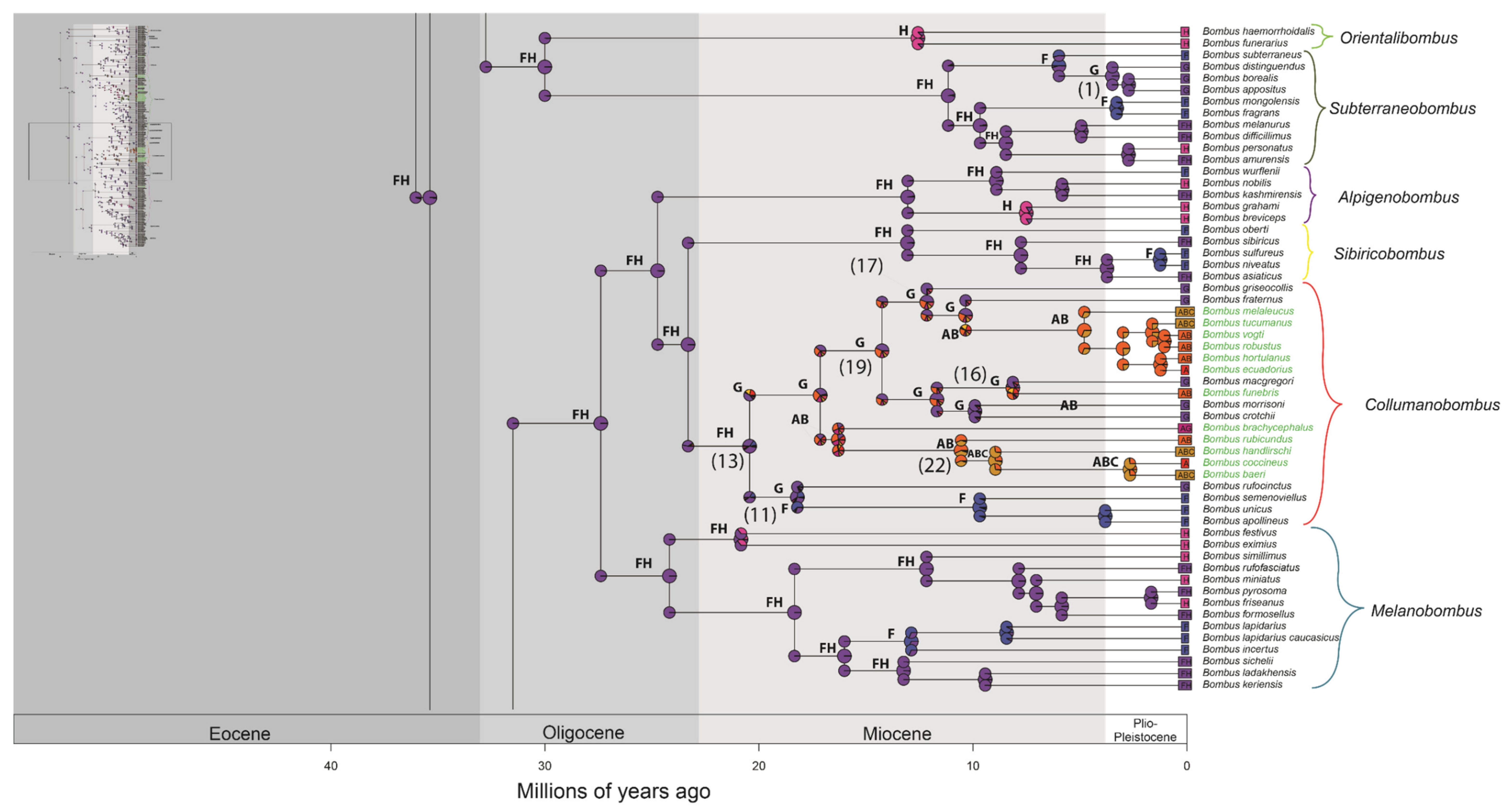
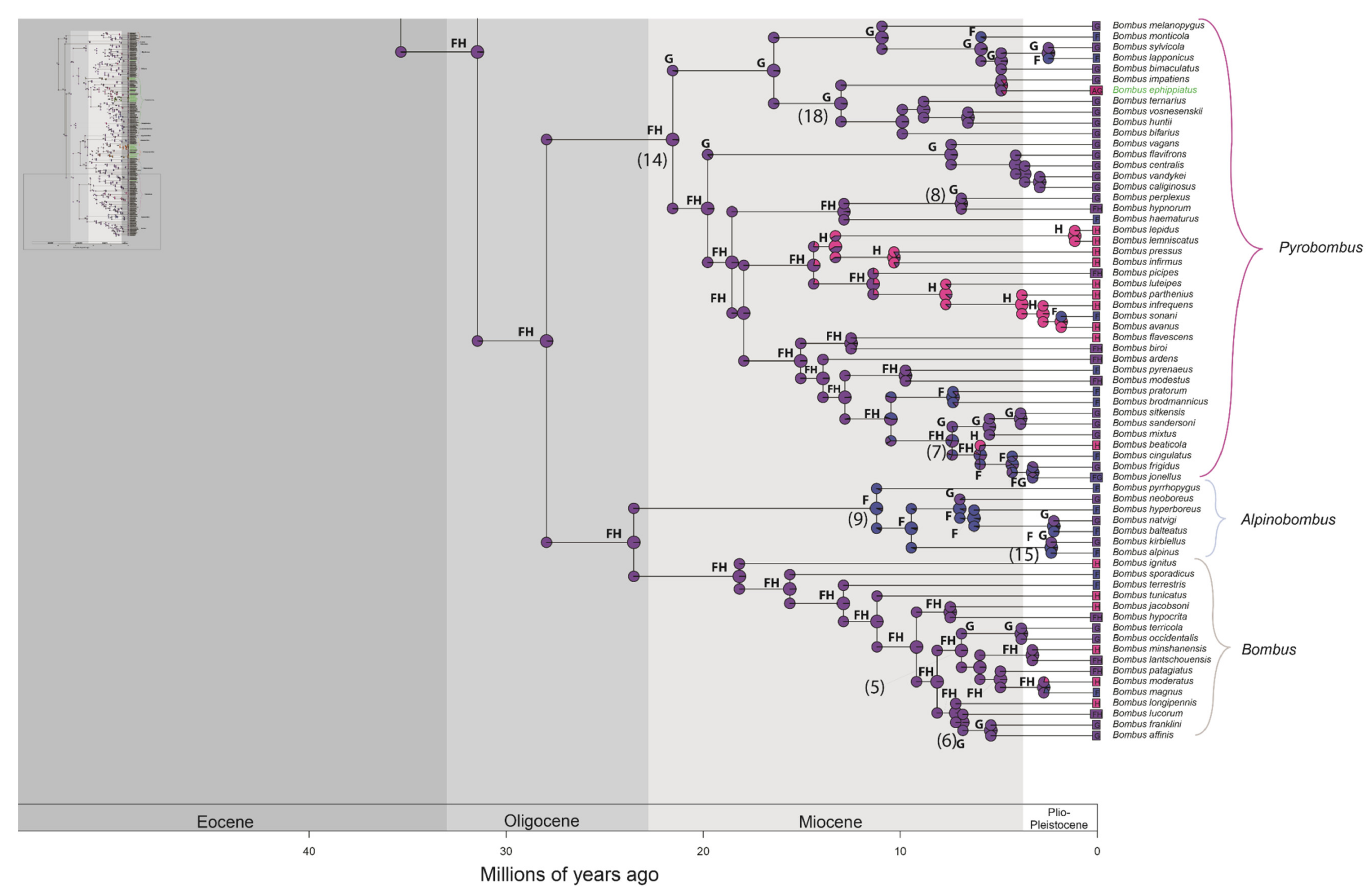
3. Results
3.1. Phylogenetic Inference
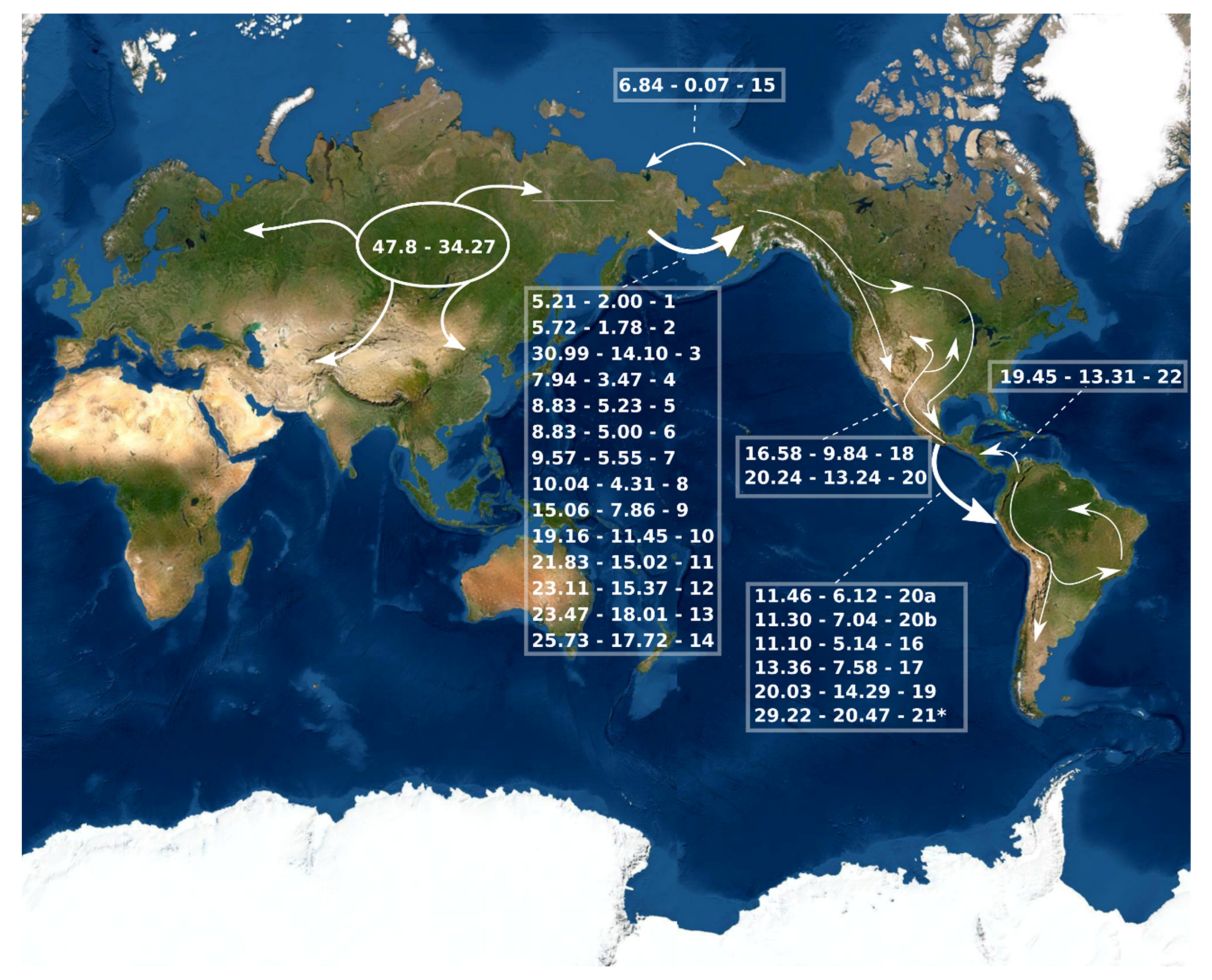
3.2. Divergence Time Estimation and Biogeographic History
4. Discussion
4.1. Phylogenetic Inference
4.2. Divergence Time Estimation and Biogeographic History
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Michener, C.D. The Bees of the World, 2nd ed.; Johns Hopkins University Press: Baltimore, MD, USA, 2007; ISBN 978-0-8018-8573-0. [Google Scholar]
- Danforth, B.N.; Cardinal, S.; Praz, C.; Almeida, E.A.B.; Michez, D. The Impact of Molecular Data on Our Understanding of Bee Phylogeny and Evolution. Annu. Rev. Entomol. 2013, 58, 57–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, R.S.; Krogmann, L.; Mayer, C.; Donath, A.; Gunkel, S.; Meusemann, K.; Kozlov, A.; Podsiadlowski, L.; Petersen, M.; Lanfear, R.; et al. Evolutionary History of the Hymenoptera. Curr. Biol. 2017, 27, 1013–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roig-Alsina, A.; Michener, C.D. Studies of the Phylogeny and Classification of Long-Tongued Bees. Univ. Kans. Sci. Bull. 1993, 55, 123–173. [Google Scholar]
- Williams, P. An Annotated Checklist of Bumble Bees with an Analysis of Patterns of Description (Hymenoptera: Apidae, Bombini). Bull. Nat. Hist. Mus. 1998, 67, 79–152. [Google Scholar]
- Santos Júnior, J.E.; Silveira, F.A.; Oliveira, U.; Dias, C.A.R.; Santos, F.R. Conservation and Historical Distribution of Two Bumblebee Species from the Atlantic Forest. Syst. Biodivers. 2019, 17, 22–38. [Google Scholar] [CrossRef]
- Williams, P.H. Mapping Variations in the Strength and Breadth of Biogeographic Transition Zones Using Species Turnover. Proc. R. Soc. Lond. B Biol. Sci. 1996, 263, 579–588. [Google Scholar]
- Williams, P.H. A Preliminary Cladistic Investigation of Relationships among the Bumble Bees (Hymenoptera, Apidae). Syst. Entomol. 1985, 10, 239–255. [Google Scholar] [CrossRef]
- Cameron, S.A.; Hines, H.M.; Williams, P.H. A Comprehensive Phylogeny of the Bumble Bees (Bombus). Biol. J. Linn. Soc. 2007, 91, 161–188. [Google Scholar] [CrossRef] [Green Version]
- Kawakita, A.; Sota, T.; Ito, M.; Ascher, J.S.; Tanaka, H.; Kato, M.; Roubik, D.W. Phylogeny, Historical Biogeography, and Character Evolution in Bumble Bees (Bombus: Apidae) Based on Simultaneous Analysis of Three Nuclear Gene Sequences. Mol. Phylogenetics Evol. 2004, 31, 799–804. [Google Scholar] [CrossRef]
- Hines, H.M. Historical Biogeography, Divergence Times, and Diversification Patterns of Bumble Bees (Hymenoptera: Apidae: Bombus). Syst. Biol. 2008, 57, 58–75. [Google Scholar] [CrossRef]
- Williams, P.H.; Lobo, J.M.; Meseguer, A.S. Bumblebees Take the High Road: Climatically Integrative Biogeography Shows That Escape from Tibet, Not Tibetan Uplift, Is Associated with Divergences of Present-Day Mendacibombus. Ecography 2018, 41, 461–477. [Google Scholar] [CrossRef] [Green Version]
- Nee, S.; May, M.R.; Harvey, P.H. The Reconstructed Evolutionaty Process. Philos. Trans. R. Soc. B Biol. Sci. 1994, 344, 305–311. [Google Scholar]
- Jablonski, D.; Roy, K.; Valentine, J.W.; Price, R.M.; Anderson, P.S. The Impact of the Pull of the Recent on the History of Marine Diversity. Science 2003, 300, 1133–1135. [Google Scholar] [CrossRef] [Green Version]
- Nee, S. Birth-Death Models in Macroevolution. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 1–17. [Google Scholar] [CrossRef]
- Etienne, R.S.; Rosindell, J. Prolonging the Past Counteracts the Pull of the Present: Protracted Speciation Can Explain Observed Slowdowns in Diversification. Syst. Biol. 2012, 61, 204–213. [Google Scholar] [CrossRef] [Green Version]
- Engel, M.S. A Monograph of the Baltic Amber Bees and Evolution of the Apoidea (Hymenoptera). Bull. Am. Mus. Nat. Hist. 2001, 259, 1–192. [Google Scholar] [CrossRef]
- Wappler, T.; De Meulemeester, T.; Murat Aytekin, A.; Michez, D.; Engel, M.S. Geometric Morphometric Analysis of a New Miocene Bumble Bee from the Randeck Maar of Southwestern Germany (Hymenoptera: Apidae). Syst. Entomol. 2012, 37, 784–792. [Google Scholar] [CrossRef]
- Prokop, J.; Dehon, M.; Michez, D.; Engel, M.S. An Early Miocene Bumble Bee from Northern Bohemia (Hymenoptera, Apidae). ZooKeys 2017, 710, 43–63. [Google Scholar] [CrossRef]
- Dehon, M.; Michez, D.; Nel, A.; Engel, M.S.; De Meulemeester, T. Wing Shape of Four New Bee Fossils (Hymenoptera: Anthophila) Provides Insights to Bee Evolution. PLoS ONE 2014, 9, e108865. [Google Scholar] [CrossRef] [Green Version]
- Dehon, M.; Engel, M.S.; Gérard, M.; Aytekin, A.M.; Ghisbain, G.; Williams, P.H.; Rasmont, P.; Michez, D. Morphometric Analysis of Fossil Bumble Bees (Hymenoptera, Apidae, Bombini) Reveals Their Taxonomic Affinities. Zookeys 2019, 891, 71–118. [Google Scholar] [CrossRef] [Green Version]
- Engel, M.S.; Grimaldi, D.A.; Gonzalez, V.H.; Hinojosa-Díaz, I.A.; Michener, C.D. An Exomalopsine Bee in Early Miocene Amber from the Dominican Republic (Hymenoptera: Apidae). Am. Mus. Novit. 2012, 3758, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.; Russel, D.W. Molecular Cloning. A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Santos Júnior, J.E.; Santos, F.R.; Silveira, F.A. Hitting an Unintended Target: Phylogeography of Bombus Brasiliensis Lepeletier, 1836 and the First New Brazilian Bumblebee Species in a Century (Hymenoptera: Apidae). PLoS ONE 2015, 10, e0125847. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A Novel Method for Rapid Multiple Sequence Alignment Based on Fast Fourier Transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An Integrated and Extendable Desktop Software Platform for the Organization and Analysis of Sequence Data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Castresana, J. Selection of Conserved Blocks from Multiple Alignments for Their Use in Phylogenetic Analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Talavera, G.; Castresana, J. Improvement of Phylogenies after Removing Divergent and Ambiguously Aligned Blocks from Protein Sequence Alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [Green Version]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation Software for the Fast Assembly of Multi-Gene Datasets with Character Set and Codon Information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New Methods for Selecting Partitioned Models of Evolution for Molecular and Morphological Phylogenetic Analyses. Mol. Biol. Evol. 2016, 34, 772–773. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Pattengale, N.; Aberer, A.; Swenson, K.; Stamatakis, A.; Moret, B. Uncovering Hidden Phylogenetic Consensus in Large Data Sets. IEEE/ACM Trans. Comput. Biol. Bioinform. 2011, 8, 902–911. [Google Scholar] [CrossRef] [PubMed]
- Westover, K.M.; Rusinko, J.P.; Hoin, J.; Neal, M. Rogue Taxa Phenomenon: A Biological Companion to Simulation Analysis. Mol. Phylogenetics Evol. 2013, 69, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian Inference of Phylogenetic Trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiens, J.J.; Morrill, M.C. Missing Data in Phylogenetic Analysis: Reconciling Results from Simulations and Empirical Data. Syst. Biol. 2011, 60, 719–731. [Google Scholar] [CrossRef]
- Guillerme, T.; Cooper, N. Effects of Missing Data on Topological Inference Using a Total Evidence Approach. Mol. Phylogenetics Evol. 2016, 94, 146–158. [Google Scholar] [CrossRef]
- Wiens, J.J.; Moen, D.S. Missing Data and the Accuracy of Bayesian Phylogenetics. J. Syst. Evol. 2008, 46, 307–314. [Google Scholar]
- Dunn, K.A.; McEachran, J.D.; Honeycutt, R.L. Molecular Phylogenetics of Myliobatiform Fishes (Chondrichthyes: Myliobatiformes), with Comments on the Effects of Missing Data on Parsimony and Likelihood. Mol. Phylogenetics Evol. 2003, 27, 259–270. [Google Scholar] [CrossRef]
- Simmons, M.P. Misleading Results of Likelihood-Based Phylogenetic Analyses in the Presence of Missing Data. Cladistics 2012, 28, 208–222. [Google Scholar] [CrossRef]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.-H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A Software Platform for Bayesian Evolutionary Analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Suchard, M.A.; Drummond, A.J. Tracer v1.6: Molecular Evolution, Phylogenetics and Epidemiology. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 1 January 2022).
- Matzke, N.J. Probabilistic Historical Biogeography: New Models for Founder-Event Speciation, Imperfect Detection, and Fossils Allow Improved Accuracy and Model-Testing. Front. Biogeogr. 2013, 5, 242–248. [Google Scholar] [CrossRef] [Green Version]
- Paradis, E.; Schliep, K. Ape 5.0: An Environment for Modern Phylogenetics and Evolutionary Analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- Morrone, J.J. Cladistic Biogeography of the Neotropical Region: Identifying the Main Events in the Diversification of the Terrestrial Biota. Cladistics 2013, 30, 1–13. [Google Scholar] [CrossRef]
- Ree, R.H.; Smith, S.A. Maximum Likelihood Inference of Geographic Range Evolution by Dispersal, Local Extinction, and Cladogenesis. Syst. Biol. 2008, 57, 4–14. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F. Dispersal-Vicariance Analysis: A New Approach to the Quantification of Historical Biogeography. Syst. Biol. 1997, 46, 195–203. [Google Scholar] [CrossRef]
- Landis, M.J.; Matzke, N.J.; Moore, B.R.; Huelsenbeck, J.P. Bayesian Analysis of Biogeography When the Number of Areas Is Large. Syst. Biol. 2013, 62, 789–804. [Google Scholar] [CrossRef] [Green Version]
- Matzke, N.J. Model Selection in Historical Biogeography Reveals That Founder-Event Speciation Is a Crucial Process in Island Clades. Syst. Biol. 2014, 63, 951–970. [Google Scholar] [CrossRef]
- Williams, P.H.; Cameron, S.A.; Hines, H.M.; Cederberg, B.; Rasmont, P. A Simplified Subgeneric Classification of the Bumblebees (Genus Bombus). Apidologie 2008, 39, 46–74. [Google Scholar] [CrossRef] [Green Version]
- Françoso, E.; de Oliveira, F.F.; Arias, M.C. An Integrative Approach Identifies a New Species of Bumblebee (Hymenoptera: Apidae: Bombini) from Northeastern Brazil. Apidologie 2016, 47, 171–185. [Google Scholar] [CrossRef] [Green Version]
- Moure, J.S.; Sakagami, S. As Mamamgabas Sociais Do Brasil (Bombus Latreille) (Hymenoptera, Apoidea). Stud. Entomol. 1962, 51, 65–194. [Google Scholar]
- Milliron, H.E. A Monograph of the Western Hemisphere Bumblebees (Hymenoptera: Apidae; Bombinae. II. The Genus Megabombus Subgenus Megabombus. Mem. Entomol. Soc. Can. 1973, 89, 81–237. [Google Scholar] [CrossRef]
- Sanmartín, I.; Enghoff, H.; Ronquist, F. Patterns of Animal Dispersal, Vicariance and Diversification in the Holarctic. Biol. J. Linn. Soc. 2001, 73, 345–390. [Google Scholar] [CrossRef]
- Uba, C.E.; Strecker, M.R.; Schmitt, A.K. Increased Sediment Accumulation Rates and Climatic Forcing in the Central Andes during the Late Miocene. Geology 2007, 35, 979. [Google Scholar] [CrossRef]
- Retallack, G.J.; Kirby, M.X. Middle Miocene Global Change and Paleogeography of Panama. PALAIOS 2007, 22, 667–679. [Google Scholar] [CrossRef]
- Goulson, D. Bumblebees: Behaviour, Ecology, and Conservation, 2nd ed.; Oxford Biology; Oxford University Press: Oxford, UK; New York, NY, USA, 2010; ISBN 978-0-19-955306-8. [Google Scholar]
- Lepais, O.; Darvill, B.; O’Connor, S.; Osborne, J.L.; Sanderson, R.A.; Cussans, J.; Goffe, L.; Goulson, D. Estimation of Bumblebee Queen Dispersal Distances Using Sibship Reconstruction Method. Mol. Ecol. 2010, 19, 819–831. [Google Scholar] [CrossRef]
- Owen, R.E.; Otterstatter, M.C.; Cartar, R.V.; Farmer, A.; Colla, S.R.; O’Toole, N. Significant Expansion of the Distribution of the Bumble Bee Bombus Moderatus (Hymenoptera: Apidae) in Alberta over 20 Years 1 This Paper Is Dedicated to the Memory of Adolf Scholl. Can. J. Zool. 2012, 90, 133–138. [Google Scholar] [CrossRef]
- Makinson, J.C.; Woodgate, J.L.; Reynolds, A.; Capaldi, E.A.; Perry, C.J.; Chittka, L. Harmonic Radar Tracking Reveals Random Dispersal Pattern of Bumblebee (Bombus Terrestris) Queens after Hibernation. Sci. Rep. 2019, 9, 4651. [Google Scholar] [CrossRef] [Green Version]
- Futuyma, D.J.; Kirkpatrick, M. Evolution, 4th ed.; Sinauer Associates, Inc.: Massachusetts, UK, 2017; ISBN 978-1-60535-696-9. [Google Scholar]
- Hoorn, C.; Wesselingh, F.P.; ter Steege, H.; Bermudez, M.A.; Mora, A.; Sevink, J.; Sanmartin, I.; Sanchez-Meseguer, A.; Anderson, C.L.; Figueiredo, J.P.; et al. Amazonia Through Time: Andean Uplift, Climate Change, Landscape Evolution, and Biodiversity. Science 2010, 330, 927–931. [Google Scholar] [CrossRef] [Green Version]
- Condamine, F.L.; Hines, H.M. Historical Species Losses in Bumblebee Evolution. Biol. Lett. 2015, 11, 20141049. [Google Scholar] [CrossRef] [PubMed]
- Raup, D.M.; Gould, S.J.; Schopf, T.J.; Simberloff, D.S. Stochastic Models of Phylogeny and the Evolution of Diversity. J. Geol. 1973, 81, 525–542. [Google Scholar] [CrossRef]
- Purvis, A.; Orme, C.D.L.; Toomey, N.H.; Pearson, P.N. Temporal Patterns in Diversification Rates. In Speciation and Patterns of Diversity; Butlin, R., Schluter, D., Bridle, J., Eds.; Cambridge University Press: Cambridge, UK, 2009; Chapter 15. [Google Scholar]
- Phillimore, A.B.; Price, T.D. Density-Dependent Cladogenesis in Birds. PLoS Biol. 2008, 6, e71. [Google Scholar] [CrossRef] [Green Version]
- Ricklefs, R.E. Evolutionary Diversification, Coevolution between Populations and Their Antagonists, and the Filling of Niche Space. Proc. Natl. Acad. Sci. USA 2010, 107, 1265–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosindell, J.; Harmon, L.J.; Etienne, R.S. Unifying Ecology and Macroevolution with Individual-Based Theory. Ecol. Lett. 2015, 18, 472–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartenberger, J.-L. An Asian Grande Coupure. Nature 1998, 394, 321. [Google Scholar] [CrossRef]
- Meng, J.; McKenna, M.C. Faunal Turnovers of Palaeogene Mammals from the Mongolian Plateau. Nature 1998, 394, 364–367. [Google Scholar] [CrossRef]
- Zachos, J.; Pagani, M.; Sloan, L.; Thomas, E.; Billups, K. Trends, Rhythms, and Aberrations in Global Climate 65 Ma to Present. Science 2001, 292, 686–693. [Google Scholar] [CrossRef]
- Montes, C.; Cardona, A.; Jaramillo, C.; Pardo, A.; Silva, J.C.; Valencia, V.; Ayala, C.; Pérez-Angel, L.C.; Rodriguez-Parra, L.A.; Ramirez, V.; et al. Middle Miocene Closure of the Central American Seaway. Science 2015, 348, 226–229. [Google Scholar] [CrossRef] [Green Version]
- Cozzuol, M.A. The Acre Vertebrate Fauna: Age, Diversity, and Geography. J. South Am. Earth Sci. 2006, 21, 185–203. [Google Scholar] [CrossRef]
- O’Dea, A.; Lessios, H.A.; Coates, A.G.; Eytan, R.I.; Restrepo-Moreno, S.A.; Cione, A.L.; Collins, L.S.; de Queiroz, A.; Farris, D.W.; Norris, R.D.; et al. Formation of the Isthmus of Panama. Sci. Adv. 2016, 2, e1600883. [Google Scholar] [CrossRef] [Green Version]
- Williams, P.; Colla, S.; Xie, Z. Bumblebee Vulnerability: Common Correlates of Winners and Losers across Three Continents. Conserv. Biol. 2009, 23, 931–940. [Google Scholar] [CrossRef]
- Francisco, F.O.; Santiago, L.R.; Mizusawa, Y.M.; Oldroyd, B.P.; Arias, M.C. Genetic Structure of Island and Mainland Populations of a Neotropical Bumble Bee Species. J. Insect Conserv. 2016, 20, 383–394. [Google Scholar] [CrossRef]
- Ehlers, T.A.; Poulsen, C.J. Influence of Andean Uplift on Climate and Paleoaltimetry Estimates. Earth Planet. Sci. Lett. 2009, 281, 238–248. [Google Scholar] [CrossRef]
- Ortiz-Jaureguizar, E.; Cladera, G.A. Paleoenvironmental Evolution of Southern South America during the Cenozoic. J. Arid Environ. 2006, 66, 498–532. [Google Scholar] [CrossRef]
- Abrahamovich, A.H.; Díaz, N.B.; Morrone, J.J. Distributional Patterns of the Neotropical and Andean Species of the Genus Bombus (Hymenoptera: Apidae). Acta Zoológica Mex. 2004, 20, 99–117. [Google Scholar]
- Moure, J.S.; Urban, D.; Melo, G.A.R. Catalogue of Bees (Hymenoptera, Apoidea) in the Neotropical Region—Online Version. Available online: http://www.moure.cria.org.br/catalogue (accessed on 2 November 2019).
- Olesen, J.M. Behaviour and Nest Structure of the Amazonian Bombus Transversalis in Ecuador. J. Trop. Ecol. 1989, 5, 243–246. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lineage | Date of Invasion (Ma) | Invasion Direction | Extant Included Species |
|---|---|---|---|
| B. morio group | 29.22–20.47 | Palearctic region to South America | B. morio, B. excellens, and B. dahlbomii |
| B. transversalis group | 20.24–13.24 | Nearctic to Neotropical region | B. trinominatus, B. weisi, B. digressus, B. fervidus, B. diligens, B. opifex, B. bellicosus, B. pennsylvanicus, B. steindachneri, B. mexicanus, B. medius, B. pullatus, B. pauloensis, B. transversalis, B. brasiliensis, B. bahiensis, B. brevivillus |
| B. bellicosus subgroup | 11.46–6.12 | Central America to South America | B. opifex, B. bellicosus |
| B. pullatus subgroup | 11.3–7.04 | Central America to South America | B. pullatus, B. pauloensis, B. transversalis, B. brasiliensis, B. bahiensis, B. brevivillus |
| B. brachycephalus group | 22.03–14.29 | Nearctic region to South America | B. brachycephalus, B. rubicundus1, B. handlirschi, B. coccineus, B. baeri |
| B. ephippiatus | 16.58–9.84 | Nearctic to Neotropical region | B. ephippiatus |
| B. intrudens | 4.8–0.97 | Nearctic to Neotropical border | B. intrudens |
| B. robustus group 2 | 13.36–7.58 | Nearctic region to South America | B. melaleucus, B. tucumanus, B. vogti, B. robustus, B. hortulanus, B. ecuadorius |
| B. funebris | 11.1–5.14 | Nearctic region to South America | B. funebris |
| B. brachycephalus | 19.45–13.31 | South America to Neotropical and Nearctic regions | B. brachycephalus |
| Subgenera | Species | Estimated Age (95% HPD) | Locality |
|---|---|---|---|
| Cullumanobombus | B. brachycephalus | 19.45–13.31 | North and Central America |
| Cullumanobombus | B. rubicundus | 14.11–7.39 | South America |
| Cullumanobombus | B. handlirschi | 12.33–5.85 | South America |
| Cullumanobombus | B. funebris/B. macgregori | 11.1–5.14 | South America |
| Cullumanobombus | B. melaleucus | 6.78–3.05 | South America |
| Cullumanobombus | B. baeri/B. coccineus | 4.76–1.02 | South America |
| Cullumanobombus | B. tucumanus | 2.66–0.68 | South America |
| Cullumanobombus | B. ecuadorius/B. hortulanus | 2.31–0.39 | South America |
| Cullumanobombus | B. robustus/B. vogti | 1.91–0.32 | South America |
| Psithyrus | B. intrudens | 4.8–0.97 | North and Central America |
| Pyrobombus | B. ephippiatus | 7.69–2.63 | North and Central America |
| Thoracobombus | B. excellens | 20.02–10.24 | South America |
| Thoracobombus | B. dahlbomii/B. morio | 17.92–8.23 | South America |
| Thoracobombus | B. weisi/B. digressus | 15.02–7.41 | North and Central America |
| Thoracobombus | B. medius | 11.3–7.04 | North and Central America |
| Thoracobombus | B. brevivillus | 9.81–5.94 | South America |
| Thoracobombus | B. bellicosus/B. opifex | 9.46–3.8 | South America |
| Thoracobombus | B. transversalis | 8.77–5.04 | South America |
| Thoracobombus | B. pauloensis/B. pullatus * | 6.21–2.53 | South America |
| Thoracobombus | B. bahiensis/B. brasiliensis | 6–2.75 | South America |
| Thoracobombus | B. mexicanus/B.steindachneri | 5.32–1.33 | North and Central America |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos Júnior, J.E.; Williams, P.H.; Dias, C.A.R.; Silveira, F.A.; Faux, P.; Coimbra, R.T.F.; Campos, D.P.; Santos, F.R. Biogeography and Diversification of Bumblebees (Hymenoptera: Apidae), with Emphasis on Neotropical Species. Diversity 2022, 14, 238. https://doi.org/10.3390/d14040238
Santos Júnior JE, Williams PH, Dias CAR, Silveira FA, Faux P, Coimbra RTF, Campos DP, Santos FR. Biogeography and Diversification of Bumblebees (Hymenoptera: Apidae), with Emphasis on Neotropical Species. Diversity. 2022; 14(4):238. https://doi.org/10.3390/d14040238
Chicago/Turabian StyleSantos Júnior, José Eustáquio, Paul H. Williams, Cayo A. Rocha Dias, Fernando A. Silveira, Pierre Faux, Raphael T. F. Coimbra, Davidson P. Campos, and Fabrício Rodrigues Santos. 2022. "Biogeography and Diversification of Bumblebees (Hymenoptera: Apidae), with Emphasis on Neotropical Species" Diversity 14, no. 4: 238. https://doi.org/10.3390/d14040238
APA StyleSantos Júnior, J. E., Williams, P. H., Dias, C. A. R., Silveira, F. A., Faux, P., Coimbra, R. T. F., Campos, D. P., & Santos, F. R. (2022). Biogeography and Diversification of Bumblebees (Hymenoptera: Apidae), with Emphasis on Neotropical Species. Diversity, 14(4), 238. https://doi.org/10.3390/d14040238