
Importance of Mangroves for Bat Research and Conservation: A Case Study from Vietnam with Notes on Echolocation of Myotis hasselti

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bat Capture and Morphological Measurements
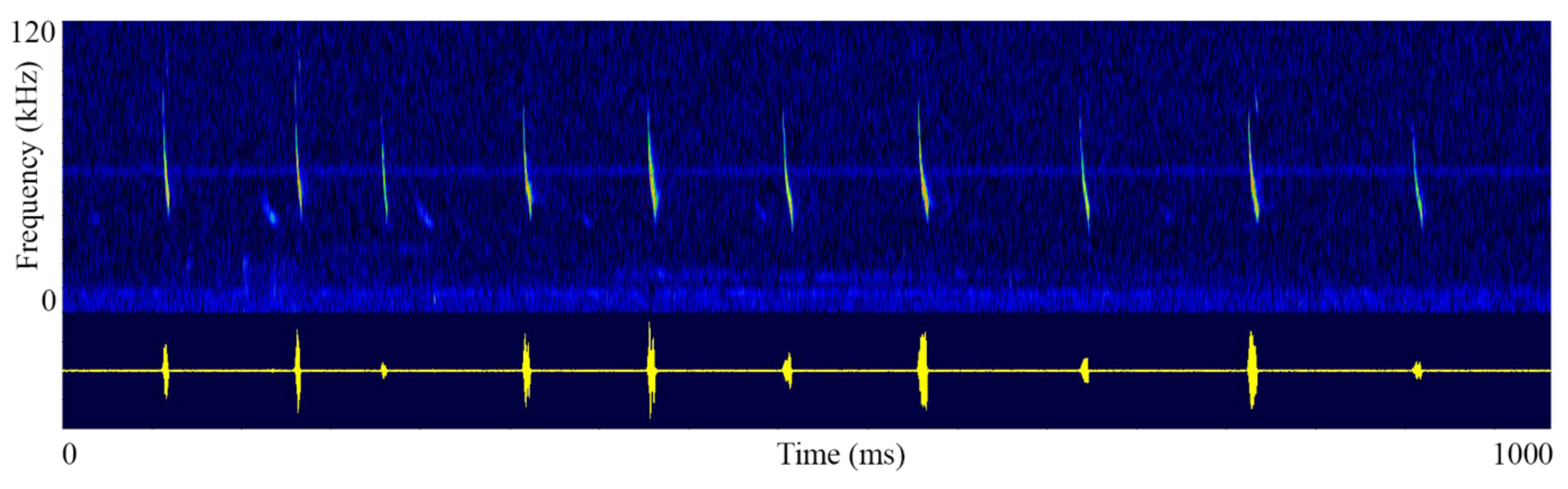
2.2. Echolocation Recording and Analyses
2.3. Tissue Sampling and Genetic Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nguyen, H.T.T.; Hardy, G.E.S.; Le, T.V.; Nguyen, H.Q.; Nguyen, H.H.; Nguyen, T.V.; Dell, B. Mangrove Forest Landcover Changes in Coastal Vietnam: A Case Study from 1973 to 2020 in Thanh Hoa and Nghe An Provinces. Forests 2021, 12, 637. [Google Scholar] [CrossRef]
- Veettil, B.K.; Ward, R.D.; Quang, N.X.; Trang, N.T.T.; Giang, T.H. Mangroves of Vietnam: Historical development, current state of research and future threats. Estuar. Coast. Shelf Sci. 2019, 218, 212–236. [Google Scholar] [CrossRef]
- Thuy, P.T.; Phuong, V.T.; Chien, P.D.; Trang, D.L.H.; Truong, N.V.; Hoa, H.N.V.; Long, H.T.; Chi, D.T.L.; Tien, N.D. Opportunities and challenges for mangrove management in Vietnam: Lessons learned from Thai Binh, Quang Ninh and Thanh Hoa provinces. Occas. Pap. 2019, 197, 1–55. [Google Scholar]
- Tuan, L.A.; Du, L.V.; Skinner, T. Rapid integrated and ecosystem-based assessment of climate change vulnerability and adaption for Ben Tre province, Vietnam. J. Sci. Technol. 2014, 52, 287–293. [Google Scholar]
- Veettil, B.K.; Quang, N.X.; Trang, N.T.T. Changes in mangrove vegetation, aquaculture and paddy cultivation in the Mekong Delta: A study from Ben Tre Province, southern Vietnam. Estuar. Coast. Shelf Sci. 2019, 226, 106273. [Google Scholar] [CrossRef]
- Carugati, L.; Gatto, B.; Rastelli, E.; Martire, M.L.; Coral, C.; Greco, S.; Danovaro, R. Impact of mangrove forests degradation on biodiversity and ecosystem functioning. Sci. Rep. 2018, 8, 13298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sikes, R.S.; Gannon, W.L. The Animal Care and Use Committee of the American Society of Mammalogists. Guidelines of the American Society of Mammalogists for the use of wild mammals in research. J. Mammal. 2011, 92, 235–253. [Google Scholar] [CrossRef]
- Sikes, R.S.; Animal Care and Use Committee of the American Society of Mammalogists. 2016 Guidelines of the American Society of Mammalogists for the use of wild mammals in research and education. J. Mammal. 2016, 97, 663–688. [Google Scholar] [CrossRef] [PubMed]
- Bates, P.J.J.; Harrison, D.L. Bats of the Indian Subcontinent; Harrison Zoological Museum: Sevenoaks, UK, 1997; pp. 1–258. [Google Scholar]
- Borissenko, A.V.; Kruskop, S.V. Bats of Vietnam and Adjacent Territories: An Identification Manual; Joint Russian-Vietnamese Science and Technological Tropical Centre: Moscow, Russia; Hanoi, Vietnam, 2003; pp. 1–212. [Google Scholar]
- Kruskop, S.V. Bats of Vietnam: Checklist and an Identification Manual, 2nd ed.; KMK Ltd.: Moscow, Russia, 2013; pp. 1–300. [Google Scholar]
- Racey, P.A. Reproductive assessment in bats. In Ecological and Behavioral Methods for the Study of Bats; Kunz, T.H., Parsons, S., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2009; pp. 249–264. [Google Scholar]
- Brunet-Rossinni, A.K.; Wilkinson, G.S. Methods for age estimation and the study of senescence in bats. In Ecological and Behavioral Methods for the Study of Bats; Kunz, T.H., Parsons, S., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2009; pp. 315–325. [Google Scholar]
- Ivanova, N.V.; Dewaard, J.R.; Hebert, P.D.N. An inexpensive, automation-friendly protocol for recovering high-quality DNA. Mol. Ecol. Notes 2006, 6, 998–1002. [Google Scholar] [CrossRef]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.E.; Mittermerier, R.A.; Martinez-Vilalta, A.; Leslie, D.M.; Olive, M.; Elliott, A.; Velikov, I.; Mascarell, A.; Sogorb, L.; Marti, B.; et al. Handbook of the Mammals of the World; Lynx Edicions: Barcelona, Spain, 2019; pp. 1–1008. [Google Scholar]
- Francis, C. A Guide to the Mammals of South-East Asia, 2nd ed.; Bloomsbury Publishing Plc: London, UK, 2019; pp. 1–416. [Google Scholar]
- Simmons, N.B.; Cirranello, A.L. Bat Species of the World: A Taxonomic and Geographic Database 2020. Available online: https://batnames.org (accessed on 19 May 2021).
- Jiang, T.L.; Feng, J.; Csorba, G.; Bates, P. Myotis pilosus. The IUCN Red List of Threatened Species 2019: E.T14193A22062554. Available online: https://www.iucnredlist.org/species/14193/22062554 (accessed on 19 May 2021).
- Thong, V.D.; Denzinger, A.; Sang, N.V.; Huyen, N.T.T.; Thanh, H.T.; Loi, D.N.; Nha, P.V.; Viet, N.V.; Tien, P.D.; Tuanmu, M.-N.; et al. Bat Diversity in Cat Ba Biosphere Reserve, Northeastern Vietnam: A Review with New Records from Mangrove Ecosystem. Diversity 2021, 13, 376. [Google Scholar] [CrossRef]
- Arnett, E.; Baerwald, E.; Mathews, F.; Rodrigues, L.; Rodríguez-Durán, A.; Rydell, J.; Villegas-Patraca, R.; Voigt, C. Impacts of Wind Energy Development on Bats: A Global Perspective. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Springer International Publishing AG: Cham, Switzerland, 2016; pp. 295–323. [Google Scholar]
- Tesfahunegny, W.; Datiko, D.; Wale, M.; Hailay, G.E.; Hunduma, T. Impact of wind energy development on birds and bats: The case of Adama wind farm, Central Ethiopia. J. Basic Appl. Zool. 2020, 81, 1–9. [Google Scholar] [CrossRef]
- Peterson, T.S.; Pelletier, S.K.; Boyden, S.A.; Watrous, K.S. Offshore Acoustic Monitoring of Bats in the Gulf of Maine. Northeast. Nat. 2014, 21, 86–107. [Google Scholar] [CrossRef]
- Thompson, R.H.; Thompson, A.R.; Brigham, R.M. A Flock of Myotis Bats at Sea. Northeast. Nat. 2015, 22, 27–30. [Google Scholar] [CrossRef]
- Pelletier, S.K.; Peterson, T.S. Bats at sea: A final 6-year summary of bat monitoring on islands, offshore structures, and coastal sites in the Gulf of Maine, mid-Atlantic States, and Great Lakes Regions. In Proceedings of the Wind Wildlife Research Meeting XI, Broomfield, CO, USA, 29 November–2 December 2016. [Google Scholar]
- Mas, M.; Flaquer, C.; Rebelo, H.; López-Baucells, A. Bats and wetlands: Synthesising gaps in current knowledge and future opportunities for conservation. Mammal Rev. 2021, 51, 369–384. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Site | Locality Name | Coordinates | Captured Species (Individuals) |
|---|---|---|---|
| 1 | Thua My 2 (Cay Bang), Binh Dai District | 10°10′37″ N; 106°44′38″ E | M. hasselti (6♂, 8♀); Myotis pilosus (1♂, 1♀) |
| 2 | Binh Thuan (30–4), Binh Dai District | 10°09′39″ N; 106°42′23″ E | M. hasselti (3♂, 9♀); T. melanopogon (3♂, 3♀) |
| 3 | Thua Tien 1, Binh Dai District | 10°09′36″ N; 106°46′39″ E | C. brachyotis (9♂, 12♀); M. minimus (7♂, 15♀); M. hasselti (6♂, 9♀); T. melanopogon (1♂, 2♀) |
| 4 | Thua Tien 5, Binh Dai District | 10°08′56″ N; 106°47′15″ E | M. minimus (2♂, 2♀) |
| 5 | Thua Tien 11, Binh Dai District | 10°08′53″ N; 106°46′49″ E | C. brachyotis (1♂, 1♀); M. minimus (2♂, 3♀) |
| 6 | Thua Loi 8, Binh Dai District | 10°07′34″ N; 106°47′10″ E | C. brachyotis (3♂, 3♀); M. minimus (1♂, 3♀) |
| 7 | Thua Loi 2, Binh Dai District | 10°06′59″ N; 106°47′17″ E | C. brachyotis (3♂, 6♀); M. minimus (3♂, 3♀) |
| 8 | Thua Loi 11, Binh Dai District | 10°06′32″ N; 106°47′13″ E | C. brachyotis (1♂, 1♀) |
| 9 | Thanh Hai 8, Thanh Phu District | 09°50′46″ N; 106°39′47″ E | C. brachyotis (3♂, 9♀); M. minimus (2♂, 6♀) |
| 10 | Thanh Hai 1, Thanh Phu District | 09°50′27″ N; 106°39′32″ E | M. hasselti (6♂, 6♀); Myotis pilosus (1♀) |
| 11 | Thanh Hai 2, Thanh Phu District | 09°50′25″ N; 106°39′37″ E | C. brachyotis (2♂, 5♀); M. minimus (3♂, 3♀) |
| 12 | Thanh Hai 5, Thanh Phu District | 09°49′10″ N; 106°38′39″ E | C. brachyotis (3♂, 3♀); Myotis pilosus (1♂) |
| 13 | Dau Go Area, Ha Long Bay | 20°54′37″ N; 107°01′13″ E | M. hasselti (4♂, 6♀); Myotis pilosus (3♂, 3♀), T. melanopogon (7♀) |
| Species | n | Sex | FA | EH | TIB | HF | Tail |
|---|---|---|---|---|---|---|---|
| Cynopterus brachyotis | 40 | ♀♀ | 65.8 ± 1.0 63.8–66.9 | 16.0 ± 0.2 15.8–16.2 (3) | 23.9 ± 0.3 23.6–24.1 (3) | 12.5 ± 0.2 12.3–12.6 (3) | 11.1 ± 0.5 10.6–11.5 (3) |
| 25 | ♂♂ | 65.4 ± 1.0 63.5–66.8 | 15.7 ± 0.1 15.6–15.8 (3) | 25.1 ± 0.5 24.6–25.6 (3) | 13.0 ± 0.9 12.2–13.9 (3) | 13.3 ± 1.6 11.5–14.7 (3) | |
| Macroglossus minimus | 35 | ♀♀ | 40.6 ± 1.0 39.1–44.5 | 16.3 (1) | 16.1 (1) | 10.3 (1) | 2.5 (1) |
| 20 | ♂♂ | 40.3 ± 0.5 39.5–41.5 | 15.8; 16.2 (2) | 15.9; 16.4 (2) | 10.0; 10.5 (2) | 2.3; 2.5 (1) | |
| Taphozous melanopogon | 12 | ♀♀ | 65.0 ± 1.3 63.3–66.8 | 18.8 ± 0.8 17.5–19.8 | 24.7 ± 1.1 23.5–26.8 | 16.8 ± 0.8 15.7–18.0 | 27.8 ± 0.6 26.8–28.5 |
| 4 | ♂♂ | 64.4 ± 0.9 63.8–65.8 | 19.1 ± 0.4 18.6–19.6 | 25.1 ± 0.8 24.3–25.8 | 17.1 ± 0.5 16.5–17.6 | 27.8 ± 1.5 26.5–29.9 | |
| Myotis hasseltii | 38 | ♀♀ | 38.7 ± 0.4 37.5–39.8 | 13.9 ± 0.4 13.3–14.5 (8) | 16.3 ± 0.4 15.8–16.8 (8) | 9.6 ± 0.2 9.4–9.8 (8) | 39.8 ± 0.8 37.5–40.0 (8) |
| 25 | ♂♂ | 38.0 ± 0.4 37.3–38.9 | 12.2 ± 0.5 11.5–12.6 (5) | 16.6 ± 0.4 15.9–16.9 (5) | 9.2 ± 0.4 8.8–9.6 (5) | 38.3 ± 0.5 37.5–38.7 (5) | |
| Myotis pilosus | 9 | ♀♀ | 56.8 ± 0.7 55.6–57.9 | 16.6 ± 0.7 15.5–17.5 | 20.0 ± 0.9 18.6–21.5 | 16.3 ± 0.7 15.3–17.5 | 41.2 ± 3.1 38.6–46.8 |
| 6 | ♂♂ | 55.7 ± 1.3 53.8–56.8 | 16.9 ± 0.4 16.3–17.5 | 19.1 ± 0.5 18.5–19.8 | 15.9 ± 0.6 15.3–16.8 | 42.3 ± 1.5 39.8–43.8 |
| Species | n | iFM | tFM | BW | PD |
|---|---|---|---|---|---|
| Myotis hasseltii | 49 (first harmonic) | 76.4 ± 9.6 60.1–95.8 | 33.9 ± 2.4 24.9–38.0 | 42.3 ± 9.4 25.8–66.2 | 5.1 ± 1.1 2.6–6.6 |
| Myotis pilosus | 14 (first harmonic) | 62.1 ± 4.3 56.4–70.0 | 24.2 ± 1.7 21.6–26.8 | 37.9 ± 4.6 31.0–46.5 | 8.4 ± 0.5 7.6–9.3 |
| Taphozous melanopogon | 10 (second harmonic) | 32.5 ± 1.7 30.1–35.2 | 20.6 ± 0.6 19.7–21.6 | 11.9 ± 2.0 9.4–15.5 | 9.3 ± 2.4 4.8–12.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thong, V.D.; Denzinger, A.; Long, V.; Sang, N.V.; Huyen, N.T.T.; Thien, N.H.; Luong, N.K.; Tuan, L.Q.; Ha, N.M.; Luong, N.T.; et al. Importance of Mangroves for Bat Research and Conservation: A Case Study from Vietnam with Notes on Echolocation of Myotis hasselti. Diversity 2022, 14, 258. https://doi.org/10.3390/d14040258
Thong VD, Denzinger A, Long V, Sang NV, Huyen NTT, Thien NH, Luong NK, Tuan LQ, Ha NM, Luong NT, et al. Importance of Mangroves for Bat Research and Conservation: A Case Study from Vietnam with Notes on Echolocation of Myotis hasselti. Diversity. 2022; 14(4):258. https://doi.org/10.3390/d14040258
Chicago/Turabian StyleThong, Vu Dinh, Annette Denzinger, Vu Long, Nguyen Van Sang, Nguyen Thi Thu Huyen, Nguyen Hoang Thien, Nguyen Khanh Luong, Le Quang Tuan, Nguyen Manh Ha, Nguyen Thanh Luong, and et al. 2022. "Importance of Mangroves for Bat Research and Conservation: A Case Study from Vietnam with Notes on Echolocation of Myotis hasselti" Diversity 14, no. 4: 258. https://doi.org/10.3390/d14040258
APA StyleThong, V. D., Denzinger, A., Long, V., Sang, N. V., Huyen, N. T. T., Thien, N. H., Luong, N. K., Tuan, L. Q., Ha, N. M., Luong, N. T., & Schnitzler, H. -U. (2022). Importance of Mangroves for Bat Research and Conservation: A Case Study from Vietnam with Notes on Echolocation of Myotis hasselti. Diversity, 14(4), 258. https://doi.org/10.3390/d14040258