
One on Top of the Other: Exploring the Habitat Cascades Phenomenon in Iconic Biogenic Marine Habitats



 ,
, 

Abstract
:- (1)
- Is the ‘habitat cascades’ phenomenon present in remote areas, and if yes, what is its role in shaping local biodiversity?
- (2)
- What is the role of environmental parameters (e.g., depth, habitat type, food availability) in the presence and magnitude of habitat cascades?
- (3)
- Are there common biological traits across the organisms that facilitate habitat cascades across different types of habitats?
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thomsen, M.S.; Wernberg, T.; Altieri, A.; Tuya, F.; Gulbransen, D.; McGlathery, K.J.; Holmer, M.; Silliman, B.R. Habitat cascades: The conceptual context and global relevance of facilitation cascades via habitat formation and modification. Integr. Comp. Biol. 2010, 50, 158–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomsen, M.S.; Altieri, A.H.; Angelini, C.; Bishop, M.J.; Gribben, P.E.; Lear, G.; He, Q.; Schiel, D.R.; Silliman, B.R.; South, P.M.; et al. Secondary foundation species enhance biodiversity. Nat. Ecol. Evol. 2018, 2, 634–639. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.M.; Wheeler, A.J.; Freiwald, A. Reefs of the deep: The biology and geology of cold-water coral ecosystems. Science 2006, 312, 543–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, L.-A.; Roberts, J.M. Global biodiversity in cold-water coral reef ecosystems. In Marine Animal Forests: The Ecology of Benthic Biodiversity Hotspots; Rossi, S., Bramanti, L., Gori, A., Orejas, C., Eds.; Springer International Publishing: New York, NY, USA, 2017; pp. 1–21. [Google Scholar]
- Gerovasileiou, V.; Bianchi, C.N. Mediterranean marine caves: A synthesis of current knowledge. Oceanogr. Mar. Biol. Annu. Rev. 2021, 59, 1–88. [Google Scholar]
- Kazanidis, G.; Henry, L.-A.; Roberts, J.M. Hidden structural heterogeneity enhances marine hotspots’ biodiversity. Coral Reefs 2021, 40, 1615–1630. [Google Scholar] [CrossRef]
- Kazanidis, G.; Henry, L.-A.; Vad, J.; Johnson, C.; De Clippele, L.H.; Roberts, J.M. ATLAS Mingulay Reef Complex Macrobenthos and Environmental Data; Pangaea: Bremen, Germany, 2020. [Google Scholar] [CrossRef]
- Nakamura, A.; Kitching, R.L.; Cao, M.; Creedy, T.J.; Fayle, T.M.; Freiberg, M.; Hewitt, C.N.; Itioka, T.; Pin Koh, L.; Ma, K.; et al. Forests and their canopies: Achievements and horizons in canopy science. Trends Ecol. Evol. 2017, 32, 438–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belmonte, G.; Ingrosso, G.; Poto, M.; Quarta, G.; D’Elia, M.; Onorato, R.; Calcagnile, L. Biogenic stalactites in submarine caves at the Cape of Otranto (SE Italy): Dating and hypothesis on their formation. Mar. Ecol. 2009, 30, 376–382. [Google Scholar] [CrossRef]
- Belmonte, G.; Guido, A.; Mastandrea, A.; Onorato, R.; Rosso, A.; Sanfilippo, R. Animal forests in submarine caves. In Perspectives on the Marine Animal Forests of the World; Rossi, S., Bramanti, L., Eds.; Springer: Cham, Switzerland, 2020; pp. 129–145. [Google Scholar]
- Guido, A.; Jimenez, C.; Achilleos, K.; Rosso, A.; Sanfilippo, R.; Hadjioannou, L.; Petrou, A.; Russo, F.; Mastandrea, A. Cryptic serpulid-microbialite bioconstructions in the Kakoskali submarine cave (Cyprus, Eastern Mediterranean). Facies 2017, 63, 21. [Google Scholar] [CrossRef]
- Guido, A.; Gerovasileiou, V.; Russo, F.; Rosso, A.; Sanfilippo, R.; Voultsiadou, E.; Mastandrea, A. Composition and biostratinomy of sponge-rich biogenic crusts in submarine caves (Aegean Sea, Eastern Mediterranean). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 534, 109338. [Google Scholar] [CrossRef]
- Sanfilippo, R.; Rosso, A.; Guido, A.; Gerovasileiou, V. Serpulid communities from two marine caves in the Aegean Sea, eastern Mediterranean. J. Mar. Biol. Assoc. UK 2017, 97, 1059–1068. [Google Scholar] [CrossRef] [Green Version]
- Sanfilippo, R.; Rosso, A.; Guido, A.; Mastandrea, A.; Russo, F.; Riding, R.; Taddei Ruggiero, E. Metazoan/microbial biostalactites from present-day submarine caves in the Mediterranean Sea. Mar. Ecol. 2015, 36, 1277–1293. [Google Scholar] [CrossRef]
- Rosso, A.; Sanfilippo, R.; Guido, A.; Gerovasileiou, V.; Taddei Ruggiero, E.; Belmonte, G. Colonisers of the dark: Biostalactite-associated metazoans from “lu Lampiùne” submarine cave (Apulia, Mediterranean Sea). Mar. Ecol. 2021, 42, e12634. [Google Scholar] [CrossRef]
- Guido, A.; Heindel, K.; Birgel, D.; Rosso, A.; Mastandrea, A.; Sanfilippo, R.; Russo, F.; Peckmann, J. Pendant bioconstructions cemented by microbial carbonate in submerged marine caves (Holocene, SE Sicily). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 388, 166–180. [Google Scholar] [CrossRef]
- Guido, A.; Rosso, A.; Sanfilippo, R.; Russo, F.; Mastandrea, A. Frutexites from microbial/metazoan bioconstructions of recent and Pleistocene marine caves (Sicily, Italy). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 453, 127–138. [Google Scholar] [CrossRef]
- Gischler, E.; Heindel, K.; Birgel, D.; Brunner, B.; Reitner, J.; Peckmann, J. Cryptic biostalactites in a submerged karst cave of the Belize Barrier Reef revisited: Pendant bioconstructions cemented by microbial micrite. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 278, 34–51. [Google Scholar] [CrossRef]
- Gischler, E.; Birgel, D.; Brunner, B.; Eisenhauer, A.; Meyer, G.; Buhre, S.; Peckmann, J. A giant underwater stalactite from the Blue Hole, Belize, revisited: A complex history of massive carbonate accretion under changing meteoric and marine conditions. J. Sediment. Res. 2017, 87, 1260–1284. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Altieri, A.H.; Angelini, C.; Bishop, M.J.; Bulleri, F.; Farhan, R.; Frühling, V.M.M.; Gribben, P.E.; Harrison, S.B.; He, Q.; et al. Heterogeneity within and among co-occurring foundation species increases biodiversity. Nat. Commun. 2022, 13, 581. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Habitat (Region) | Habitat Cascades: Basal Habitat Formers → Intermediate Habitat Formers → Focal Organisms |
|---|---|
| Cold-water coral reefs (Mingulay Reef Complex, NE Atlantic) | Cold-water corals → Molluscs → Bryozoans, brachiopods Cold-water corals → Serpulid polychaetes → Barnacles, bryozoans Cold-water corals → Eunicid polychaetes → Anthozoans, tunicates |
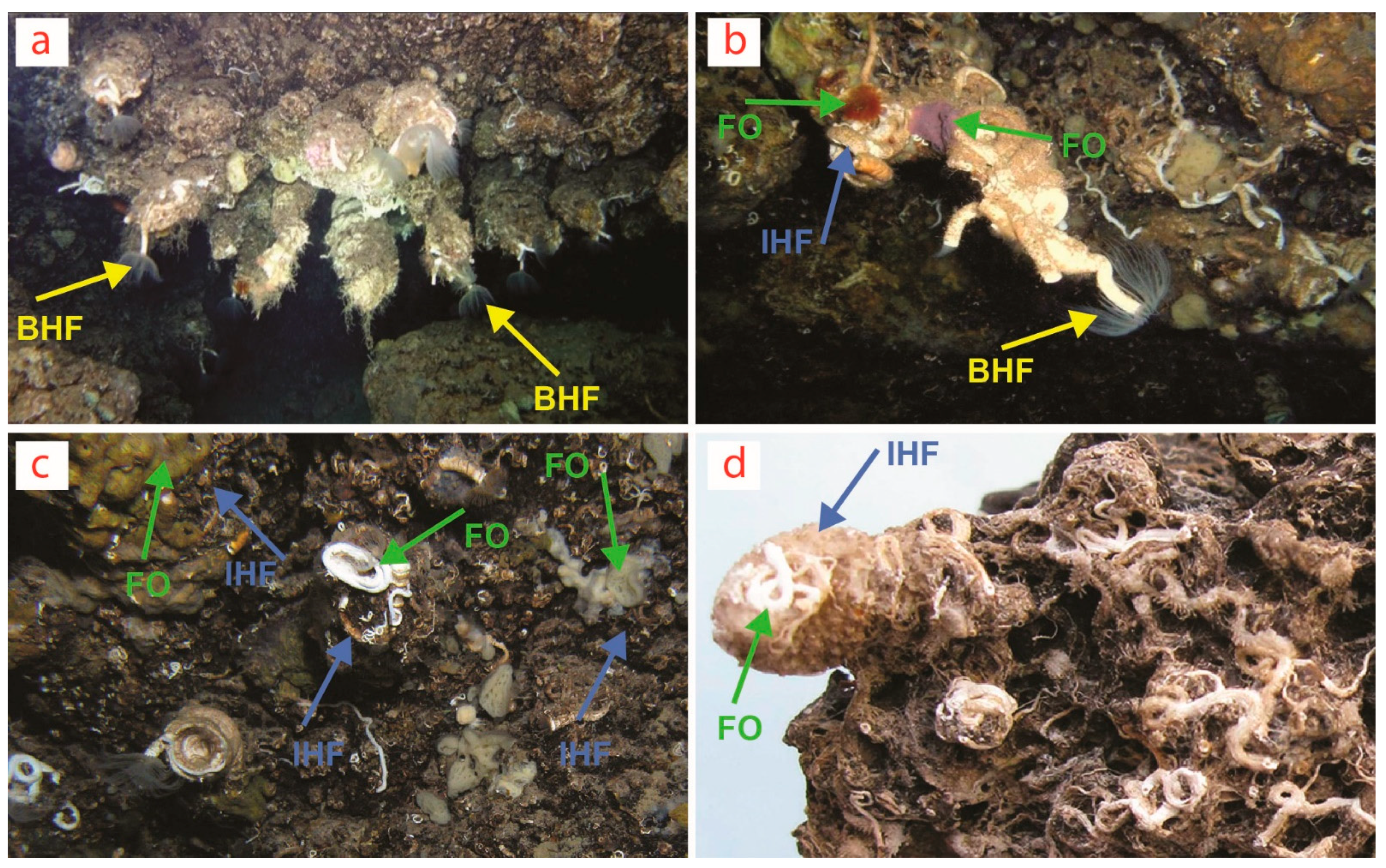
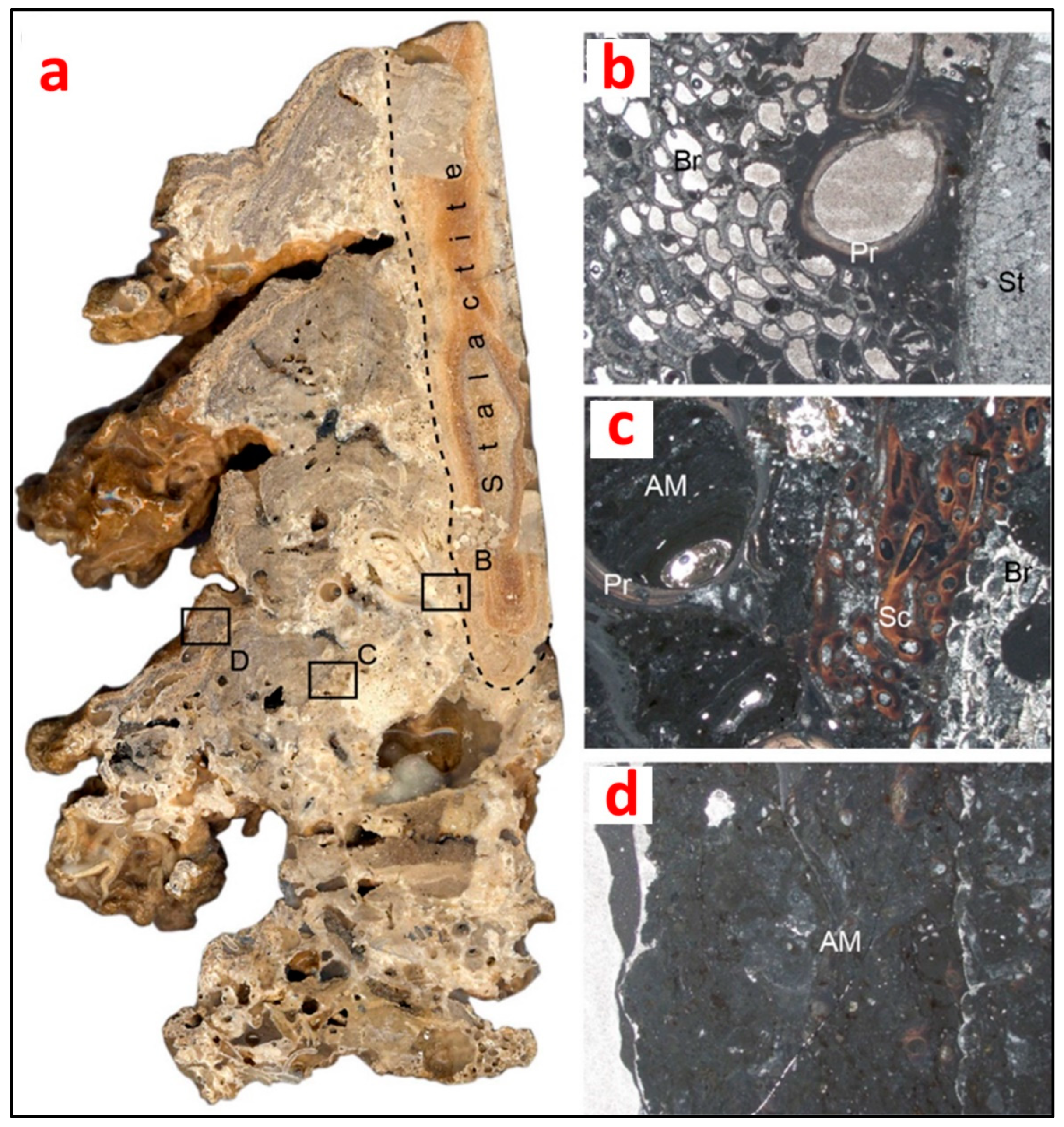
| Biogenic stalactites in marine caves (Mediterranean Sea) | Serpulid polychaetes → Other serpulids and encrusting bryozoans → Epibiota (e.g., sponges, brachiopods, foraminiferans) |
| Serpulid polychaetes → Other serpulids and encrusting bryozoans → Cavity-dwelling taxa (e.g., insinuating/boring sponges, boring bivalves, heterotrophic bacterial communities) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kazanidis, G.; Guido, A.; Rosso, A.; Sanfilippo, R.; Roberts, J.M.; Gerovasileiou, V. One on Top of the Other: Exploring the Habitat Cascades Phenomenon in Iconic Biogenic Marine Habitats. Diversity 2022, 14, 290. https://doi.org/10.3390/d14040290
Kazanidis G, Guido A, Rosso A, Sanfilippo R, Roberts JM, Gerovasileiou V. One on Top of the Other: Exploring the Habitat Cascades Phenomenon in Iconic Biogenic Marine Habitats. Diversity. 2022; 14(4):290. https://doi.org/10.3390/d14040290
Chicago/Turabian StyleKazanidis, Georgios, Adriano Guido, Antonietta Rosso, Rossana Sanfilippo, J. Murray Roberts, and Vasilis Gerovasileiou. 2022. "One on Top of the Other: Exploring the Habitat Cascades Phenomenon in Iconic Biogenic Marine Habitats" Diversity 14, no. 4: 290. https://doi.org/10.3390/d14040290
APA StyleKazanidis, G., Guido, A., Rosso, A., Sanfilippo, R., Roberts, J. M., & Gerovasileiou, V. (2022). One on Top of the Other: Exploring the Habitat Cascades Phenomenon in Iconic Biogenic Marine Habitats. Diversity, 14(4), 290. https://doi.org/10.3390/d14040290