
The Association of Waminoa with Reef Corals in Singapore and Its Impact on Putative Immune- and Stress-Response Genes


 ,
,  , , , ,
, , , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ecological Analysis
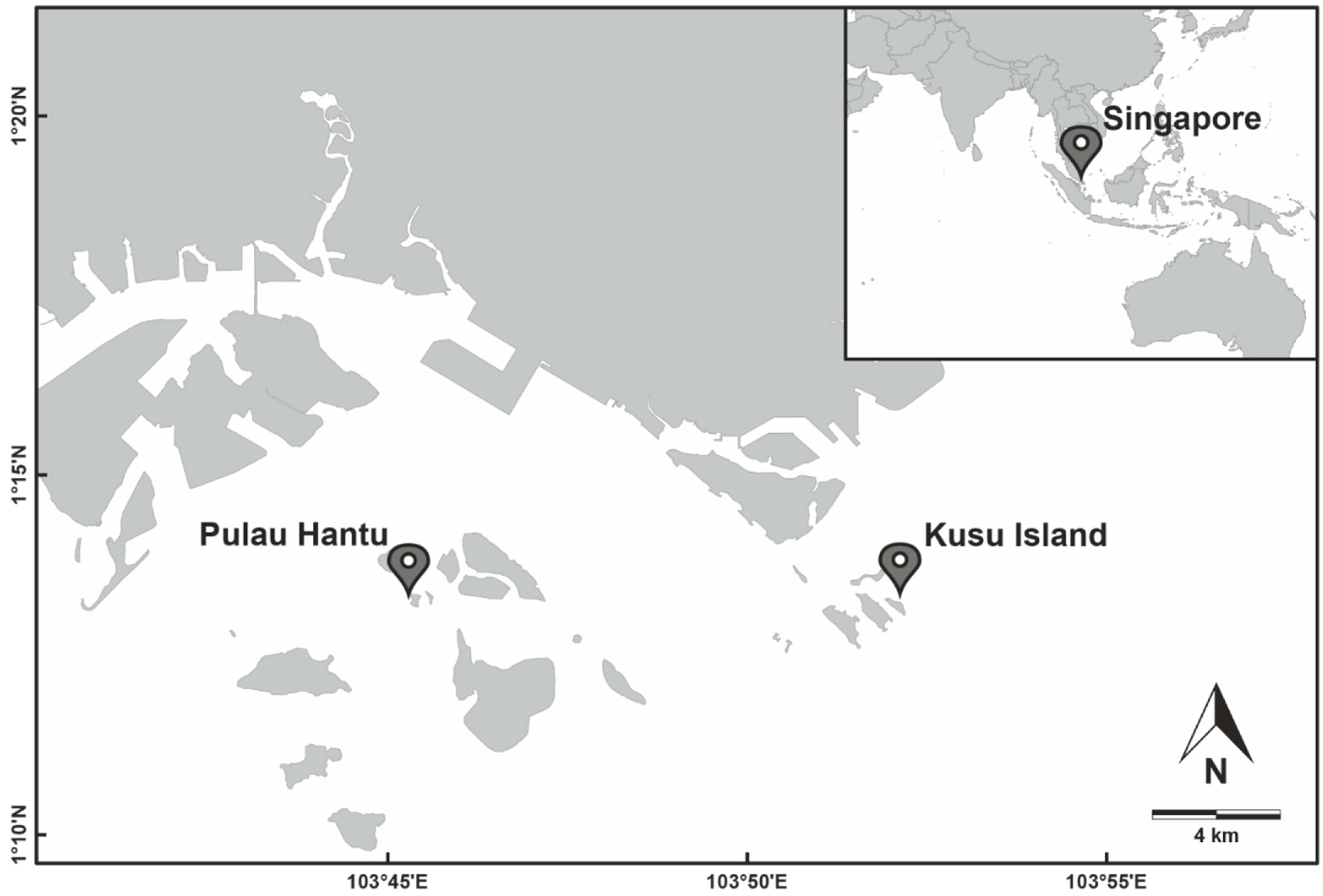
2.1.1. Study Area and Sampling Design
2.1.2. Data Analysis
2.2. Molecular Analysis
2.2.1. Coral Collection
2.2.2. RNA Extraction and REVERSE Transcription (RT)
2.2.3. Primer Design and Validation
2.2.4. Gene Expression Quantification
2.2.5. Data Analysis
3. Results
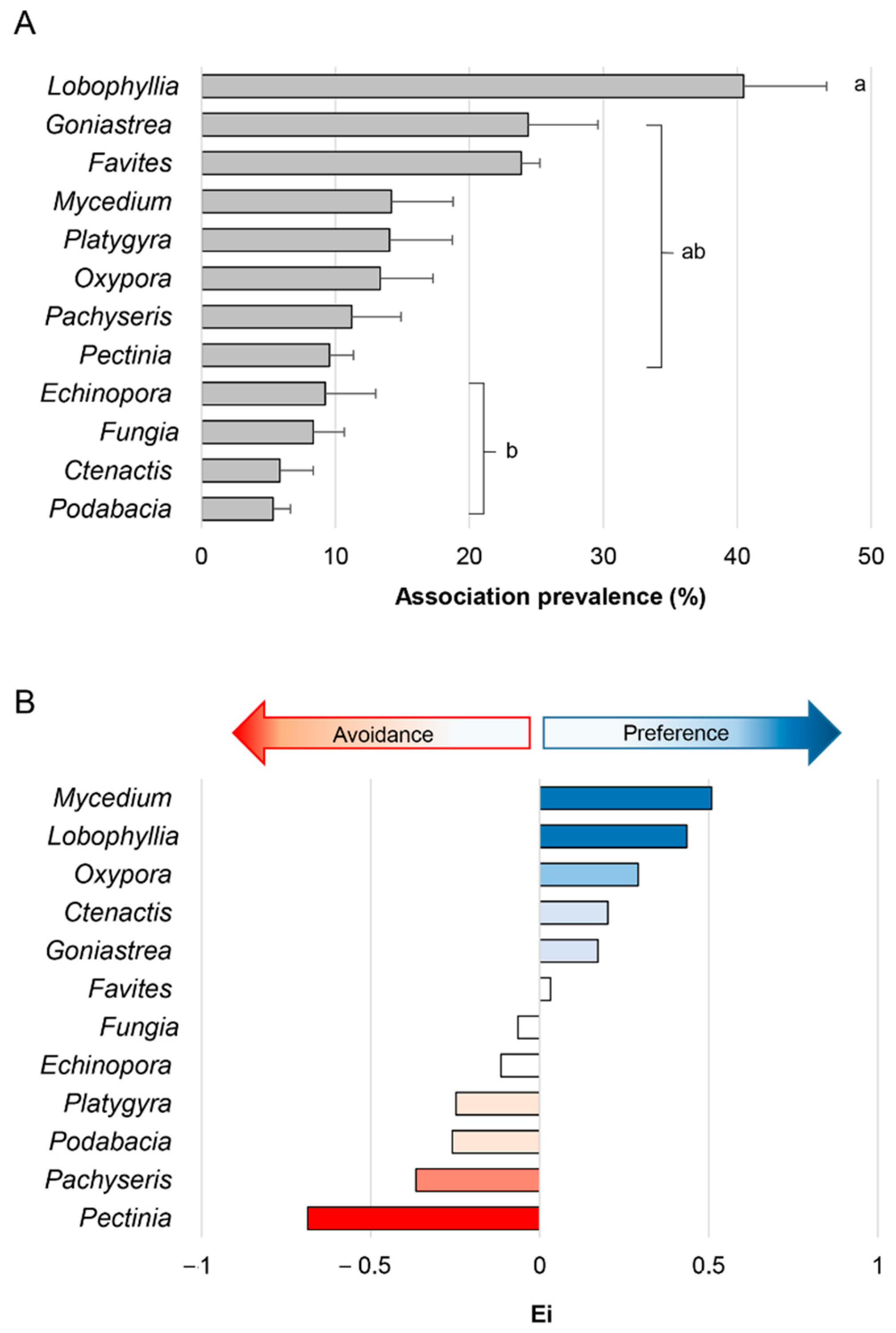
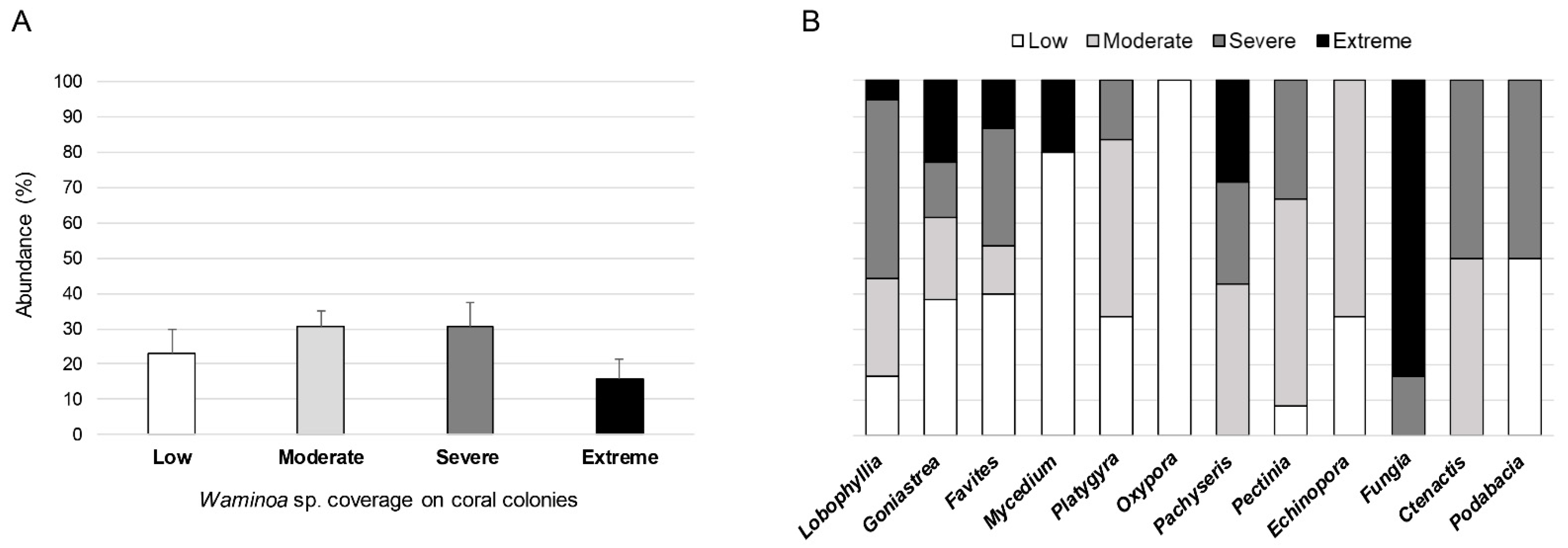
3.1. Ecological Analysis
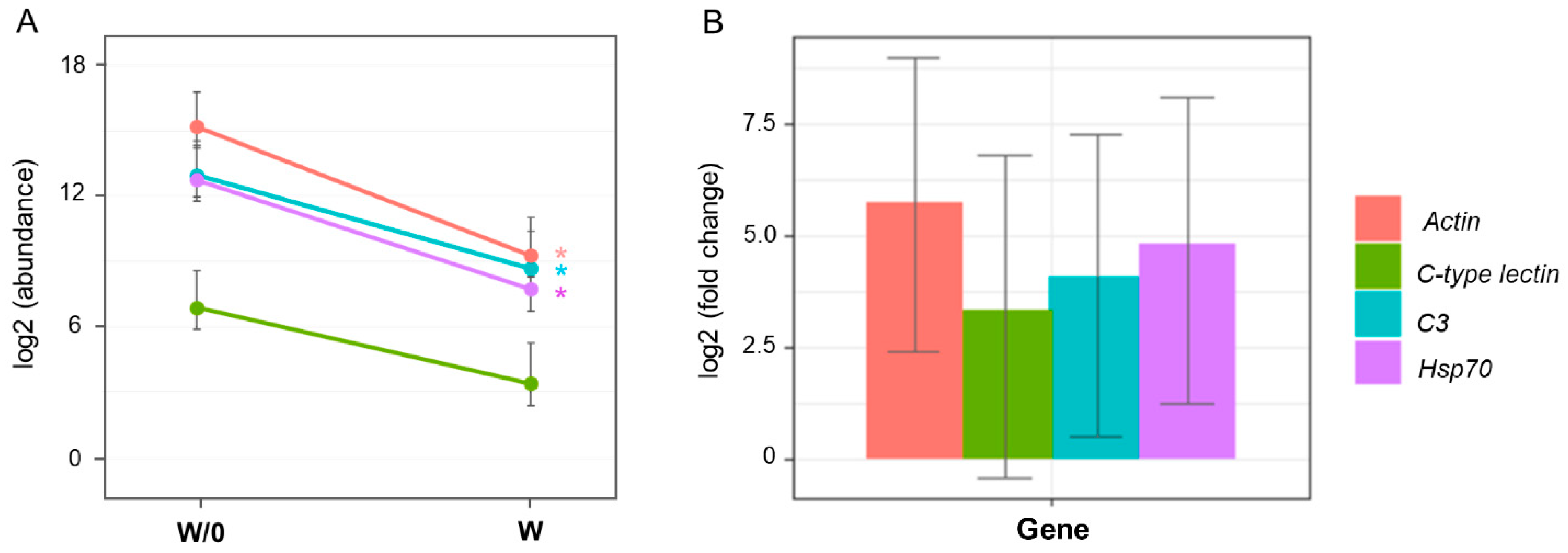
3.2. Molecular Analysis
4. Discussion
4.1. Ecology of the Waminoa-Coral Association in Singapore Reef
4.2. Effect of Waminoa sp. on Coral Putative Immune- and Stress-Response Genes
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bos, A.R. Fishes (Gobiidae and Labridae) associated with the mushroom coral Heliofungia actiniformis (Scleractinia: Fungiidae) in the Philippines. Coral Reefs 2012, 31, 133. [Google Scholar] [CrossRef] [Green Version]
- Hoeksema, B.W.; Van der Meij, S.E.T.; Fransen, C.H. The mushroom coral as a habitat. J. Mar. Biol. Assoc. 2012, 92, 647–663. [Google Scholar] [CrossRef] [Green Version]
- Bos, A.R.; Hoeksema, B.W. Cryptobenthic fishes and co-inhabiting shrimps associated with the mushroom coral Heliofungia actiniformis (Fungiidae) in the Davao Gulf, Philippines. Environ. Biol. Fish 2015, 98, 1479–1489. [Google Scholar] [CrossRef]
- Montano, S.; Seveso, D.; Galli, P.; Puce, S.; Hoeksema, B.W. Mushroom corals as newly recorded hosts of the hydrozoan symbiont Zanclea sp. Mar. Biol. Res. 2015, 11, 773–779. [Google Scholar] [CrossRef]
- Montano, S.; Fattorini, S.; Parravicini, V.; Berumen, M.L.; Galli, P.; Maggioni, D.; Arrigoni, R.; Seveso, D.; Strona, G. Corals hosting symbiotic hydrozoans are less susceptible to predation and disease. Proc. R. Soc. B Biol. Sci. 2017, 284, 20172405. [Google Scholar] [CrossRef] [Green Version]
- Maggioni., D.; Arrigoni, R.; Seveso, D.; Galli, P.; Berumen, M.L.; Denis, V.; Hoeksema, B.W.; Huang, D.; Manca, F.; Pica, D.; et al. Evolution and biogeography of the Zanclea-Scleractinia symbiosis. Coral Reefs 2020, 1–17. [Google Scholar] [CrossRef]
- Baguñà, J.; Riutort, M. Molecular phylogeny of the Platyhelminthes. Can. J. Zool. 2014, 82, 168–193. [Google Scholar] [CrossRef]
- Kunihiro, S.; Reimer, J.D. Phylogenetic analyses of Symbiodinium isolated from Waminoa and their anthozoan hosts in the Ryukyus Archipelago, southern Japan. Symbiosis 2018, 76, 253–264. [Google Scholar] [CrossRef]
- Ogunlana, M.V.; Hooge, M.D.; Tekle, Y.I.; Benayahu, Y.; Barneah, O.; Tyler, S. Waminoa brickneri n. sp. (Acoela: Acoelomorpha) associated with corals in the Red Sea. Zootaxa 2005, 14, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Hikosaka-Katayama, T.; Koike, K.; Yamashita, H.; Hikosaka, A.; Koike, K. Mechanisms of maternal inheritance of dinoflagellate symbionts in the acoelomorph worm Waminoa litus. Zool. Sci. 2012, 29, 559–567. [Google Scholar] [CrossRef] [Green Version]
- Barneah, O.; Brickner, I.; Hooge, M.; Weis, V.M.; Benayahu, Y. First evidence of maternal transmission of algal endosymbionts at an oocyte stage in a triploblastic host, with observations on reproduction in Waminoa brickneri (Acoelomorpha). Invert. Biol. 2007, 126, 113–119. [Google Scholar] [CrossRef]
- Barneah, O.; Ben-Dov, E.; Benayahu, Y.; Brickner, I.; Kushmaro, A. Molecular diversity and specificity of acoel worms associated with corals in the Gulf of Eilat (Red Sea). Aquat. Biol. 2012, 14, 277–281. [Google Scholar] [CrossRef]
- Haapkylä, J.; Seymour, A.S.; Barneah, O.; Brickner, I.; Hennige, S.; Suggett, D.; Smith, D. Association of Waminoa sp. (Acoela) with corals in the Wakatobi Marine Park, South-East Sulawesi, Indonesia. Mar. Biol. 2009, 156, 1021–1027. [Google Scholar] [CrossRef]
- Biondi, P.; Masucci, G.D.; Kunihiro, S.; Reimer, J.D. The distribution of reef-dwelling Waminoa flatworms in bays and on capes of Okinawa Island. Mar. Biodiv. 2019, 49, 405–413. [Google Scholar] [CrossRef]
- Kunihiro, S.; Farenzena, Z.; Hoeksema, B.W.; Groenenberg, D.S.; Hermanto, B.; Reimer, J.D. Morphological and phylogenetic diversity of Waminoa and similar flatworms (Acoelomorpha) in the western Pacific Ocean. Zoology 2019, 136, 125692. [Google Scholar] [CrossRef] [PubMed]
- Barton, J.A.; Bourne, D.G.; Humphrey, C.; Hutson, K.S. Parasites and coral-associated invertebrates that impact coral health. Rev. Aquacul. 2020, 12, 2284–2303. [Google Scholar] [CrossRef]
- Naumann, M.S.; Mayr, C.; Struck, U.; Wild, C. Coral mucus stable isotope composition and labeling: Experimental evidence for mucus uptake by epizoic acoelomorph worms. Mar. Biol. 2010, 157, 2521–2531. [Google Scholar] [CrossRef]
- Wijgerde, T.; Schots, P.; Van Onselen, E.; Janse, M.; Karruppannan, E.; Verreth, J.A.; Osinga, R. Epizoic acoelomorph flatworms impair zooplankton feeding by the scleractinian coral Galaxea fascicularis. Biol. Open 2012, 2, 10–17. [Google Scholar] [CrossRef] [Green Version]
- Brown, B.E.; Bythell, J.C. Perspectives on mucus secretion in reef corals. Mar. Ecol. Prog. Ser. 2005, 296, 291–309. [Google Scholar] [CrossRef] [Green Version]
- Hoeksema, B.W.; Farenzena, Z.T. Tissue loss in corals infested by acoelomorph flatworms (Waminoa sp.). Coral Reefs 2012, 31, 869. [Google Scholar] [CrossRef] [Green Version]
- Downs, C.A. Cellular diagnostics and its application to aquatic and marine toxicology. In Techniques in Aquatic Toxicology; Ostrander, G.K., Ed.; CRC Press: Boca Raton, FL, USA, 2005; Volume 2, pp. 181–208. [Google Scholar]
- Mydlarz, L.D.; McGinty, E.S.; Harvell, C.D. What are the physiological and immunological responses of coral to climate warming and disease? J. Exp. Biol. 2010, 213, 934–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosic, N.; Kaniewska, P.; Chan, C.K.K.; Ling, E.Y.S.; Edwards, D.; Dove, S.; Hoegh-Guldberg, O. Early transcriptional changes in the reef-building coral Acropora aspera in response to thermal and nutrient stress. BMC Genom. 2014, 15, 1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seveso, D.; Montano, S.; Reggente, M.A.L.; Maggioni, D.; Orlandi, I.; Galli, P.; Vai, M. The cellular stress response of the scleractinian coral Goniopora columna during the progression of the black band disease. Cell Stress Chap. 2017, 22, 225–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, C.V.; Traylor-Knowles, N. Towards an integrated network of coral immune mechanisms. Proc. R. Soc. B Biol. Sci. 2012, 279, 4106–4114. [Google Scholar] [CrossRef] [Green Version]
- Traylor-Knowles, N.; Connelly, M.T. What is currently known about the effects of climate change on the coral immune response. Cur. Clim. Change Rep. 2017, 3, 252–260. [Google Scholar] [CrossRef]
- Carroll, M.C. The role of complement and complement receptors in induction and regulation of immunity. Annu. Rev. Immun. 1998, 16, 545–568. [Google Scholar] [CrossRef]
- Fujita, T. Evolution of the lectin–complement pathway and its role in innate immunity. Nat. Rev. Immun. 2002, 2, 346–353. [Google Scholar] [CrossRef]
- Christophides, G.K.; Zdobnov, E.; Barillas-Mury, C.; Birney, E.; Blandin, S.; Blass, C.; Brey, P.T.; Collins, F.H.; Danielli, A.; Dimopoulos, G.; et al. Immunity-related genes and gene families in Anopheles gambiae. Science 2002, 298, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Ling, E.; Yu, X.Q. Cellular encapsulation and melanization are enhanced by immulectins, pattern recognition receptors from the tobacco hornworm Manduca sexta. Dev. Compar. Immun. 2006, 30, 289–299. [Google Scholar] [CrossRef]
- Endo, Y.; Nakazawa, N.; Iwaki, D.; Takahashi, M.; Matsushita, M.; Fujita, T. Interactions of ficolin and mannose-binding lectin with fibrinogen/fibrin augment the lectin complement pathway. J. Innate Immun. 2010, 2, 33–42. [Google Scholar] [CrossRef]
- Dishaw, L.J.; Smith, S.L.; Bigger, C.H. Characterization of a C3-like cDNA in a coral: Phylogenetic implications. Immunogenetics 2005, 57, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.J.; Hemmrich, G.; Ball, E.E.; Hayward, D.C.; Khalturin, K.; Funayama, N.; Agata, K.; Bosch, T.C. The innate immune repertoire in Cnidaria-ancestral complexity and stochastic gene loss. Genome Biol. 2007, 8, R59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kvennefors, E.C.E.; Leggat, W.; Kerr, C.C.; Ainsworth, T.D.; Hoegh-Guldberg, O.; Barnes, A.C. Analysis of evolutionarily conserved innate immune components in coral links immunity and symbiosis. Dev. Comp. Immun. 2010, 34, 1219–1229. [Google Scholar] [CrossRef] [PubMed]
- Shinzato, C.; Shoguchi, E.; Kawashima, T.; Hamada, M.; Hisata, K.; Tanaka, M.; Fujie, M.; Fujiwara, M.; Koyanagi, R.; Ikuta, T.; et al. Using the Acropora digitifera genome to understand coral responses to environmental change. Nature 2011, 476, 320–323. [Google Scholar] [CrossRef]
- Ocampo, I.D.; Zárate-Potes, A.; Pizarro, V.; Rojas, C.A.; Vera, N.E.; Cadavid, L.F. The immunotranscriptome of the Caribbean reef-building coral Pseudodiploria strigosa. Immunogenetics 2015, 67, 515–530. [Google Scholar] [CrossRef]
- Perrin, B.J.; Ervasti, J.M. The actin gene family: Function follows isoform. Cytoskeleton 2010, 67, 630–634. [Google Scholar] [CrossRef]
- Kenkel, C.D.; Aglyamova, G.; Alamaru, A.; Bhagooli, R.; Capper, R.; Cunning, R.; deVillers, A.; Haslun, J.A.; Hédouin, L.; Keshavmurthy, S.; et al. Development of gene expression markers of acute heat-light stress in reef-building corals of the genus Porites. PLoS ONE 2011, 6, e26914. [Google Scholar] [CrossRef] [Green Version]
- Balchin, D.; Hayer-Hartl, M.; Hartl, F.U. In vivo aspects of protein folding and quality control. Science 2016, 353, aac4354. [Google Scholar] [CrossRef]
- Downs, C.A.; Fauth, J.E.; Halas, J.C.; Dustan, P.; Bemiss, J.; Woodley, C.M. Oxidative stress and seasonal coral bleaching. Free Rad. Biol. Med. 2002, 33, 533–543. [Google Scholar] [CrossRef]
- Downs, C.A.; Woodley, C.M.; Fauth, J.E.; Knutson, S.; Burtscher, M.M.; May, L.A.; Ostrander, G.K. A survey of environmental pollutants and cellular-stress markers of Porites astreoides at six sites in St. John, US Virgin Islands. Ecotoxicology 2011, 20, 1914–1931. [Google Scholar]
- Louis, Y.D.; Bhagooli, R.; Seveso, D.; Maggioni, D.; Galli, P.; Vai, M.; Dyall, S.D. Local acclimatisation-driven differential gene and protein expression patterns of Hsp70 in Acropora muricata: Implications for coral tolerance to bleaching. Mol. Ecol. 2020, 29, 4382–4394. [Google Scholar] [CrossRef] [PubMed]
- Seveso, D.; Montano, S.; Strona, G.; Orlandi, I.; Galli, P.; Vai, M. Exploring the effect of salinity changes on the levels of Hsp60 in the tropical coral Seriatopora caliendrum. Mar. Environ. Res. 2013, 90, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Seveso, D.; Montano, S.; Strona, G.; Orlandi, I.; Galli, P.; Vai, M. Hsp60 expression profiles in the reef-building coral Seriatopora caliendrum subjected to heat and cold shock regimes. Mar. Environ. Res. 2016, 119, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Montalbetti, E.; Biscéré, T.; Ferrier-Pagès, C.; Houlbrèque, F.; Orlandi, I.; Forcella, M.; Galli, P.; Vai, M.; Seveso, D. Manganese benefits heat-stressed corals at the cellular level. Front. Mar. Sci. 2021, 8, 803. [Google Scholar] [CrossRef]
- Seveso, D.; Montano, S.; Strona, G.; Orlandi, I.; Vai, M.; Galli, P. Up-regulation of Hsp60 in response to skeleton eroding band disease but not by algal overgrowth in the scleractinian coral Acropora muricata. Mar. Environ. Res. 2012, 78, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.; Bourne, D.; Rodriguez-Lanetty, M. Transcriptional activation of c3 and hsp70 as part of the immune response of Acropora millepora to bacterial challenges. PLoS ONE 2013, 8, e67246. [Google Scholar] [CrossRef]
- Libro, S.; Kaluziak, S.T.; Vollmer, S.V. RNA-seq profiles of immune related genes in the staghorn coral Acropora cervicornis infected with white band disease. PLoS ONE 2013, 8, e81821. [Google Scholar] [CrossRef] [Green Version]
- Fuess, L.E.; Weil, E.; Mydlarz, L.D. Associations between transcriptional changes and protein phenotypes provide insights into immune regulation in corals. Dev. Compar. Immun. 2016, 62, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Guest, J.R.; Tun, K.; Low, J.; Vergés, A.; Marzinelli, E.M.; Campbell, A.H.; Bauman, A.G.; Feary, D.A.; Chou, L.M.; Steinberg, P.D. 27 years of benthic and coral community dynamics on turbid, highly urbanised reefs off Singapore. Sci. Rep. 2016, 6, 36260. [Google Scholar] [CrossRef] [Green Version]
- Chou, L.M.; Huang, D.; Tan, K.S.; Toh, T.C.; Goh, B.P.L.; Tun, K. World Seas: An Environmental Evaluation. In The Indian Ocean to the Pacific. World Seas: An Environmental Evaluation; Sheppard, C.R.C., Ed.; Academic Press: London, UK, 2019; Volume 2, pp. 539–558. [Google Scholar]
- Todd, P.A.; Ong, X.; Chou, L.M. Impacts of pollution on marine life in Southeast Asia. Biodiv. Conserv. 2010, 19, 1063–1082. [Google Scholar] [CrossRef]
- Browne, N.K.; Tay, J.K.; Low, J.; Larson, O.; Todd, P.A. Fluctuations in coral health of four common inshore reef corals in response to seasonal and anthropogenic changes in water quality. Mar. Environ. Res. 2015, 105, 39–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heery, E.C.; Hoeksema, B.W.; Browne, N.K.; Reimer, J.D.; Ang, P.O.; Huang, D.; Friess, D.A.; Chou, L.M.; Loke, L.; Saksena-Taylor, P.; et al. Urban coral reefs: Degradation and resilience of hard coral assemblages in coastal cities of East and Southeast Asia. Mar. Poll. Bull. 2018, 135, 654–681. [Google Scholar] [CrossRef] [PubMed]
- Chow, G.S.E.; Chan, Y.K.S.; Jain, S.S.; Huang, D. Light limitation selects for depth generalists in urbanised reef coral communities. Mar. Environ. Res. 2019, 147, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Guest, J.R.; Baird, A.H.; Maynard, J.A.; Muttaqin, E.; Edwards, A.J.; Campbell, S.J.; Chou, L.M. Contrasting patterns of coral bleaching susceptibility in 2010 suggest an adaptive response to thermal stress. PLoS ONE 2012, 7, e33353. [Google Scholar] [CrossRef] [PubMed]
- Chou, L.M.; Toh, T.C.; Toh, K.B.; Ng, C.S.L.; Cabaitan, P.; Tun, K.; Goh, E.; Afiq-Rosli, L.; Taira, D.; Du, R.C.; et al. Differential response of coral assemblages to thermal stress underscores the complexity in predicting bleaching susceptibility. PLoS ONE 2016, 11, e0159755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, C.S.L.; Huang, D.; Toh, K.B.; Sam, S.Q.; Kikuzawa, Y.P.; Toh, T.C.; Taira, D.; Chan, Y.K.S.; Hung, L.Z.T.; Sim, W.T.; et al. Responses of urban reef corals during the 2016 mass bleaching event. Mar. Poll. Bull. 2020, 154, 111111. [Google Scholar] [CrossRef] [PubMed]
- Januchowski-Hartley, F.A.; Bauman, A.G.; Morgan, K.M.; Seah, J.C.; Huang, D.; Todd, P.A. Accreting coral reefs in a highly urbanized environment. Coral Reefs 2020, 39, 717–731. [Google Scholar] [CrossRef]
- Huang, D.; Benzoni, F.; Arrigoni, R.; Baird, A.H.; Berumen, M.L.; Bouwmeester, J.; Chou, L.M.; Fukami, H.; Licuanan, W.Y.; Lovell, E.R.; et al. Towards a phylogenetic classification of reef corals: The Indo-Pacific genera Merulina, Goniastrea and Scapophyllia (Scleractinia, Merulinidae). Zool. Scrip. 2014, 43, 531–548. [Google Scholar] [CrossRef]
- Wong, J.S.Y.; Chan, Y.K.S.; Ng, C.S.L.; Tun, K.P.P.; Darling, E.S.; Huang, D. Comparing patterns of taxonomic, functional and phylogenetic diversity in reef coral communities. Coral Reefs 2018, 37, 737–750. [Google Scholar] [CrossRef]
- Raymundo, L.J.; Bruckner, A.W.; Work, T.M.; Willis, B. Coral Disease Handbook Guidelines For Assessment, Monitoring and Management; Couch, C.A., Harvell, C.D., Raymundo, L., Eds.; Coral Reef Targeted Research and Capacity Building for Management Program: St. Lucia, QLD, Australia; Coral Gables, FL, USA, 2008. [Google Scholar]
- Hill, J.; Wilkinson, C. Methods for Ecological Monitoring of Coral Reefs; Australian Institute of Marine Science: Townsville, Australia, 2004. [Google Scholar]
- Lechowicz, M.J. The sampling characteristics of electivity indices. Oecologia 1982, 52, 22–30. [Google Scholar] [CrossRef]
- Chesson, J. Measuring preference in selective predation. Ecology 1978, 59, 211–215. [Google Scholar] [CrossRef]
- Van der Ploeg, H.A.; Scavia, D. Calculation and use of selectivity coefficients of feeding: Zooplankton grazing. Ecol. Model. 1979, 7, 135–149. [Google Scholar] [CrossRef]
- Seveso, D.; Montano, S.; Reggente, M.A.L.; Orlandi, I.; Galli, P.; Vai, M. Modulation of Hsp60 in response to coral brown band disease. Dis. Aquat. Org. 2015, 115, 15–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poquita-Du, R.C.; Goh, Y.L.; Huang, D.; Chou, L.M.; Todd, P.A. Gene expression and photophysiological changes in Pocillopora acuta coral holobiont following heat stress and recovery. Microorganisms 2020, 8, 1227. [Google Scholar] [CrossRef] [PubMed]
- Quek, Z.B.R.; Huang, D. Effects of missing data and data type on phylotranscriptomic analysis of stony corals (Cnidaria: Anthozoa: Scleractinia). Mol. Phylogen. Evol. 2019, 134, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Voolstra, C.R.; Miller, D.J.; Ragan, M.A.; Hoffmann, A.; Hoegh-Guldberg, O.; Bourne, D.; Ball, E.E.; Ying, H.; Foret, S.; Takahashi, S.; et al. The ReFuGe 2020 Consortium—Using “omics” approaches to explore the adaptability and resilience of coral holobionts to environmental change. Front. Mar. Sci. 2015, 2, 68. [Google Scholar]
- Liew, Y.J.; Aranda, M.; Voolstra, C.R. Reefgenomics.Org–a repository for marine genomics data. Database 2016, 2016, baw152. [Google Scholar] [CrossRef]
- Matz, M.V.; Wright, R.M.; Scott, J.G. No control genes required: Bayesian analysis of qRT-PCR data. PLoS ONE 2013, 8, e71448. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Getting Started with PRIMER v7; PRIMER-E Ltd.: Plymouth, UK, 2015. [Google Scholar]
- Anderson, M.; Gorley, R.; Clarke, K. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E Ltd: Plymouth, UK, 2008. [Google Scholar]
- Ponti, M.; Fratangeli, F.; Dondi, N.; Reinach, M.S.; Serra, C.; Sweet, M.J. Baseline reef health surveys at Bangka Island (North Sulawesi, Indonesia) reveal new threats. Peer J. 2016, 4, e2614. [Google Scholar] [CrossRef] [Green Version]
- Trench, R.K.; Winsor, H. Symbiosis with Dinoflagellates in Two Pelagic Flatworms, Amphiscolops sp. and Haplodiscus sp.; Balaban Publishers: Philadelphia, PA, USA, 1987. [Google Scholar]
- Winsor, L. Marine Turbellaria (Acoela) from North Queensland. Mem. Queensl. Mus. 1990, 28, 785–800. [Google Scholar]
- Cooper, C.; Clode, P.L.; Thomson, D.P.; Stat, M. A flatworm from the genus Waminoa (Acoela: Convolutidae) associated with bleached corals in Western Australia. Zool. Sci. 2015, 32, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.; Hwang, J.S.; Yamashiro, H.; Tang, S.L. Spatial and cross-seasonal patterns of coral diseases in reefs of Taiwan: High prevalence and regional variation. Dis. Aquat. Organ. 2021, 146, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.Y.; Chen, C.C.; Chu, L.; McCarl, B. Evaluating the economic damage of climate change on global coral reefs. Glob. Environ. Change 2015, 30, 12–20. [Google Scholar] [CrossRef]
- Morgan, K.M.; Moynihan, M.A.; Sanwlani, N.; Switzer, A.D. Light limitation and depth-variable sedimentation drives vertical reef compression on turbid coral reefs. Front. Mar. Sci. 2020, 7, 931. [Google Scholar] [CrossRef]
- Stafford-Smith, M.G.; Ormond, R.F.G. Sediment-rejection mechanisms of 42 species of Australian scleractinian corals. Mar. Fresh Res. 1992, 43, 683–705. [Google Scholar] [CrossRef]
- Erftemeijer, P.L.; Riegl, B.; Hoeksema, B.W.; Todd, P.A. Environmental impacts of dredging and other sediment disturbances on corals: A review. Mar. Poll. Bull. 2012, 64, 1737–1765. [Google Scholar] [CrossRef]
- Bessell-Browne, P.; Negri, A.P.; Fisher, R.; Clode, P.L.; Duckworth, A.; Jones, R. Impacts of turbidity on corals: The relative importance of light limitation and suspended sediments. Mar. Poll. Bull. 2017, 117, 161–170. [Google Scholar] [CrossRef]
- Browne, N.; Braoun, C.; McIlwain, J.; Nagarajan, R.; Zinke, J. Borneo coral reefs subject to high sediment loads show evidence of resilience to various environmental stressors. Peer J. 2019, 7, e7382. [Google Scholar] [CrossRef] [Green Version]
- Houlbrèque, F.; Ferrier-Pagès, C. Heterotrophy in tropical scleractinian corals. Biol. Rev. 2009, 84, 1–17. [Google Scholar] [CrossRef]
- Louis, Y.D.; Bhagooli, R.; Kenkel, C.D.; Baker, A.C.; Dyall, S.D. Gene expression biomarkers of heat stress in scleractinian corals: Promises and limitations. Compar. Biochem. Physiol. Toxic Pharmacol. 2017, 191, 63–77. [Google Scholar] [CrossRef] [Green Version]
- Cziesielski, M.J.; Schmidt-Roach, S.; Aranda, M. The past, present, and future of coral heat stress studies. Ecol. Evol. 2019, 9, 10055–10066. [Google Scholar] [CrossRef] [PubMed]
- Mydlarz, L.D.; Fuess, L.; Mann, W.; Pinzón, J.H.; Gochfeld, D.J. Cnidarian immunity: From genomes to phenomes. In The Cnidaria, Past, Present and Future; Springer: Cham, Switzerland, 2016; pp. 441–446. [Google Scholar]
- Wright, R.M.; Aglyamova, G.V.; Meyer, E.; Matz, M.V. Gene expression associated with white syndromes in a reef building coral Acropora hyacinthus. BMC Genom. 2015, 16, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seneca, F.O.; Davtian, D.; Boyer, L.; Czerucka, D. Gene expression kinetics of Exaiptasia pallida innate immune response to Vibrio parahaemolyticus infection. BMC Genom. 2020, 21, 768. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.R.; Cumbo, V.R.; Harii, S.; Shinzato, C.; Chan, C.X.; Ragan, M.A.; Satoh, N.; Ball, E.E.; Miller, D.J. Deciphering the nature of the coral–Chromera association. ISME J. 2018, 12, 776–790. [Google Scholar] [CrossRef]
- Rodriguez-Lanetty, M.; Harii, S.; Hoegh-Guldberg, O. Early molecular responses of coral larvae to hyperthermal stress. Mol. Ecol. 2009, 18, 5101–5114. [Google Scholar] [CrossRef]
- Vidal-Dupiol, J.; Adjeroud, M.; Roger, E.; Foure, L.; Duval, D.; Mone, Y.; Mitta, G. Coral bleaching under thermal stress: Putative involvement of host/symbiont recognition mechanisms. BMC Physiol. 2009, 9, 14. [Google Scholar] [CrossRef] [Green Version]
- Pinzón, J.H.; Kamel, B.; Burge, C.A.; Harvell, C.D.; Medina, M.; Weil, E.; Mydlarz, L.D. Whole transcriptome analysis reveals changes in expression of immune-related genes during and after bleaching in a reef-building coral. R. Soc. Open Sci. 2015, 2, 140214. [Google Scholar] [CrossRef] [Green Version]
- Rosenzweig, R.; Nillegoda, N.B.; Mayer, M.P.; Bukau, B. The Hsp70 chaperone network. Nat. Rev. Mol. Cell. Biol. 2019, 20, 665–680. [Google Scholar] [CrossRef]
- Rosic, N.N.; Pernice, M.; Dove, S.; Dunn, S.; Hoegh-Guldberg, O. Gene expression profiles of cytosolic heat shock proteins Hsp70 and Hsp90 from symbiotic dinoflagellates in response to thermal stress: Possible implications for coral bleaching. Cell Stress Chap. 2011, 16, 69–80. [Google Scholar] [CrossRef] [Green Version]
- Seveso, D.; Montano, S.; Strona, G.; Orlandi, I.; Galli, P.; Vai, M. The susceptibility of corals to thermal stress by analyzing Hsp60 expression. Mar. Environ. Res. 2014, 99, 69–75. [Google Scholar] [CrossRef]
- Seveso, D.; Arrigoni, R.; Montano, S.; Maggioni, D.; Orlandi, I.; Berumen, M.L.; Galli, P.; Vai, M. Investigating the heat shock protein response involved in coral bleaching across scleractinian species in the central Red Sea. Coral Reefs 2020, 39, 85–98. [Google Scholar] [CrossRef]
- Zheng, B.; Han, M.; Bernier, M.; Wen, J.K. Nuclear actin and actin-binding proteins in the regulation of transcription and gene expression. FEBS J. 2009, 276, 2669–2685. [Google Scholar] [CrossRef] [PubMed]
- DeSalvo, M.K.; Voolstra, C.R.; Sunagawa, S.; Schwarz, J.A.; Stillman, J.H.; Coffroth, M.A.; Szmant, A.M.; Medina, M. Differential gene expression during thermal stress and bleaching in the Caribbean coral Montastraea faveolata. Mol. Ecol. 2008, 17, 3952–3971. [Google Scholar] [CrossRef] [PubMed]
- Kaniewska, P.; Campbell, P.R.; Kline, D.I.; Rodriguez-Lanetty, M.; Miller, D.J.; Dove, S.; Hoegh-Guldberg, O. Major cellular and physiological impacts of ocean acidification on a reef building coral. PLoS ONE 2012, 7, e34659. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence | Tm (°C) | GC % | PCR Product |
|---|---|---|---|---|
| C- type lectin | F: 5′–GTT CTA CTG GGT AGA CGA CA–3′ | 53.2 | 50.00 | 155 bp |
| R: 5′–GAA CAT CAT TCC ATG GTC CC–3′ | 53.4 | 50.00 | ||
| C3 | F: 5′–GTT GAG TTC CCT GAT GCA AT–3′ | 50.9 | 40.00 | 159 bp |
| R: 5′–CAA CAG GTA AAC GCT TTG G–3′ | 52.0 | 47.37 | ||
| Hsp70 | F: 5′–ACA ACT CCC AGC TAT GTC GC–3′ | 57.3 | 55.00 | 226 bp |
| R: 5′–TCC ACT CTC CCT TGG TCT GT–3′ | 57.6 | 55.00 | ||
| Actin | F: 5′–ATG GTT GGT ATG GGT CAG AAA G–3′ | 54.8 | 45.45 | 219 bp |
| R: 5′–TCT GTT AGC TTT TGG GTT GAG T–3′ | 54.3 | 40.91 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maggioni, G.; Huang, D.; Maggioni, D.; Jain, S.S.; Quek, R.Z.B.; Poquita-Du, R.C.; Montano, S.; Montalbetti, E.; Seveso, D. The Association of Waminoa with Reef Corals in Singapore and Its Impact on Putative Immune- and Stress-Response Genes. Diversity 2022, 14, 300. https://doi.org/10.3390/d14040300
Maggioni G, Huang D, Maggioni D, Jain SS, Quek RZB, Poquita-Du RC, Montano S, Montalbetti E, Seveso D. The Association of Waminoa with Reef Corals in Singapore and Its Impact on Putative Immune- and Stress-Response Genes. Diversity. 2022; 14(4):300. https://doi.org/10.3390/d14040300
Chicago/Turabian StyleMaggioni, Giorgia, Danwei Huang, Davide Maggioni, Sudhanshi S. Jain, Randolph Z. B. Quek, Rosa Celia Poquita-Du, Simone Montano, Enrico Montalbetti, and Davide Seveso. 2022. "The Association of Waminoa with Reef Corals in Singapore and Its Impact on Putative Immune- and Stress-Response Genes" Diversity 14, no. 4: 300. https://doi.org/10.3390/d14040300
APA StyleMaggioni, G., Huang, D., Maggioni, D., Jain, S. S., Quek, R. Z. B., Poquita-Du, R. C., Montano, S., Montalbetti, E., & Seveso, D. (2022). The Association of Waminoa with Reef Corals in Singapore and Its Impact on Putative Immune- and Stress-Response Genes. Diversity, 14(4), 300. https://doi.org/10.3390/d14040300