
Salamander Demography at Isolated Wetlands within Mature and Regenerating Forests

Abstract
:1. Introduction
2. Materials and Methods
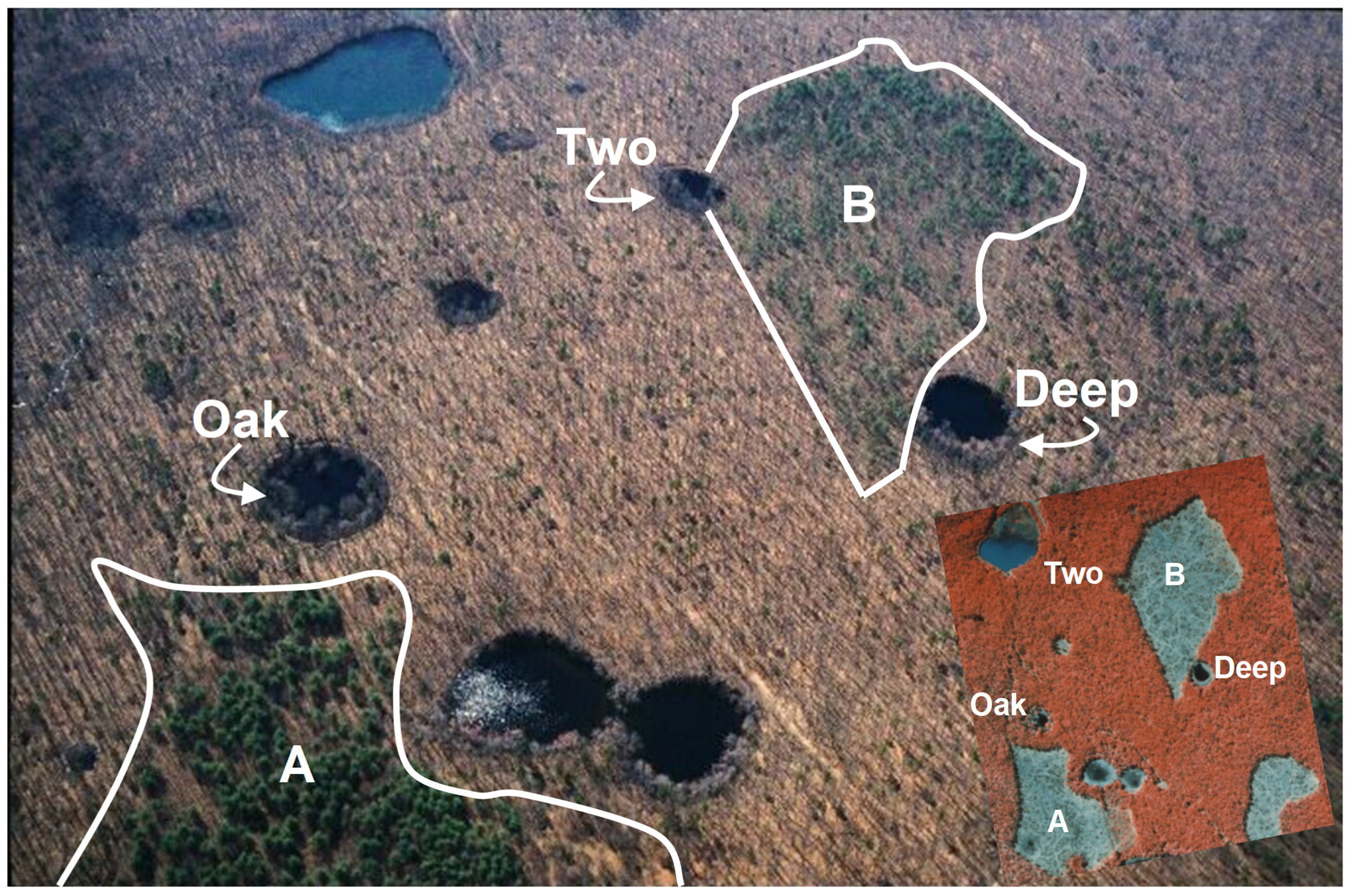
2.1. Study System and Field Methods
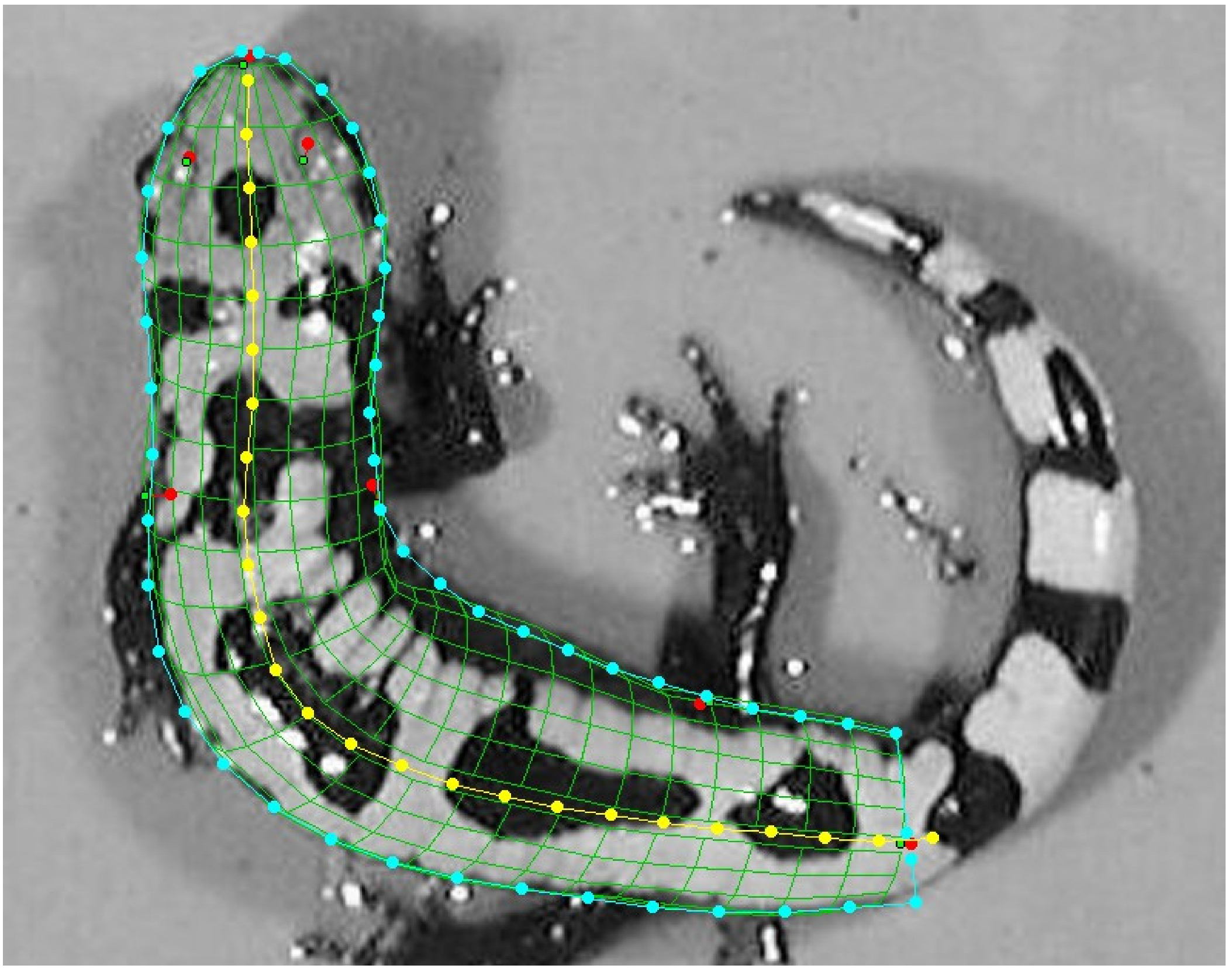
2.2. Reconstructing Individual Capture Histories
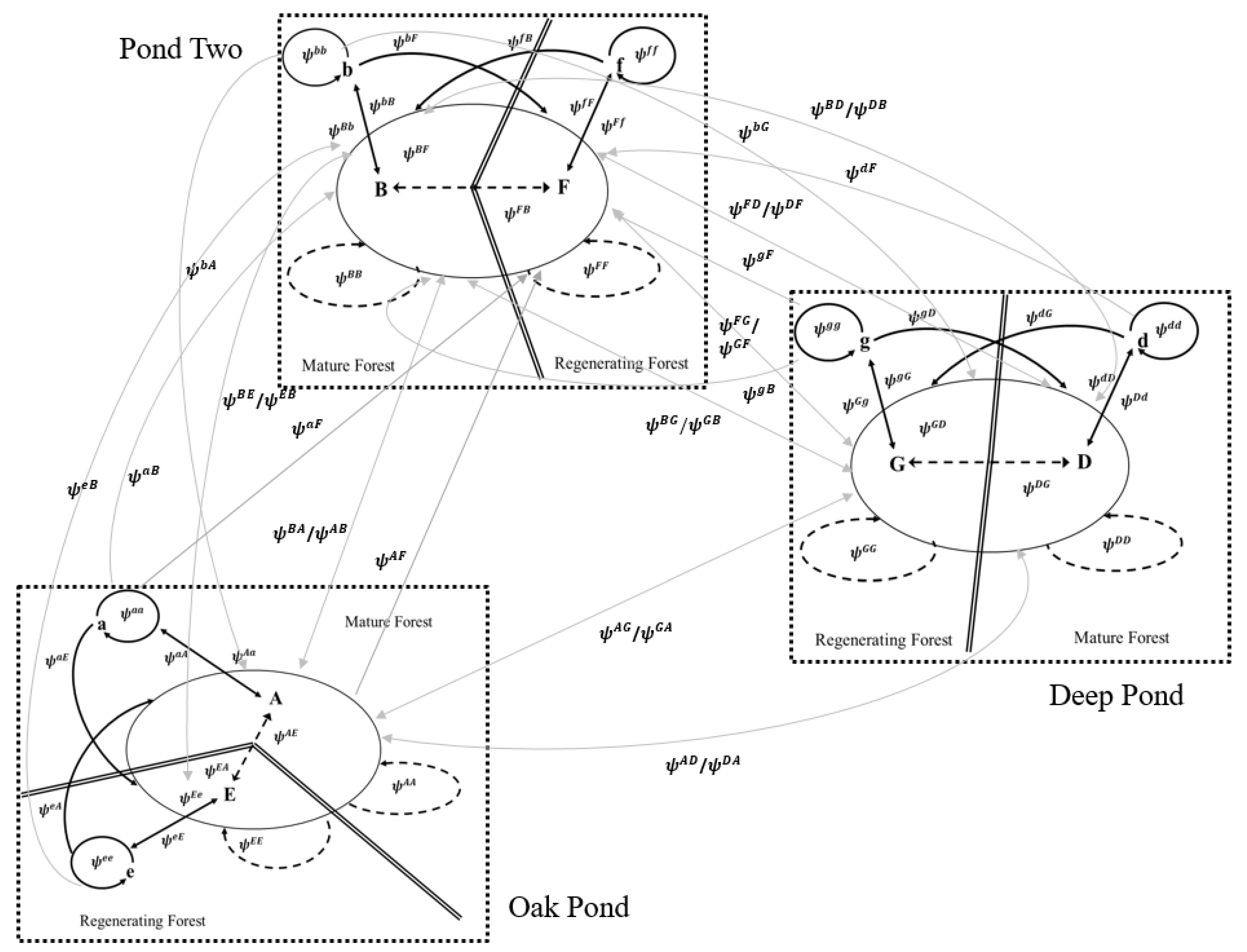
2.3. Multistate Mark-Recapture Analysis and Multimodel Inference
2.4. Candidate Model Set and Model Selection
3. Results
3.1. Matching Individual Capture Histories
3.2. Overall Inference and Model Selection Results
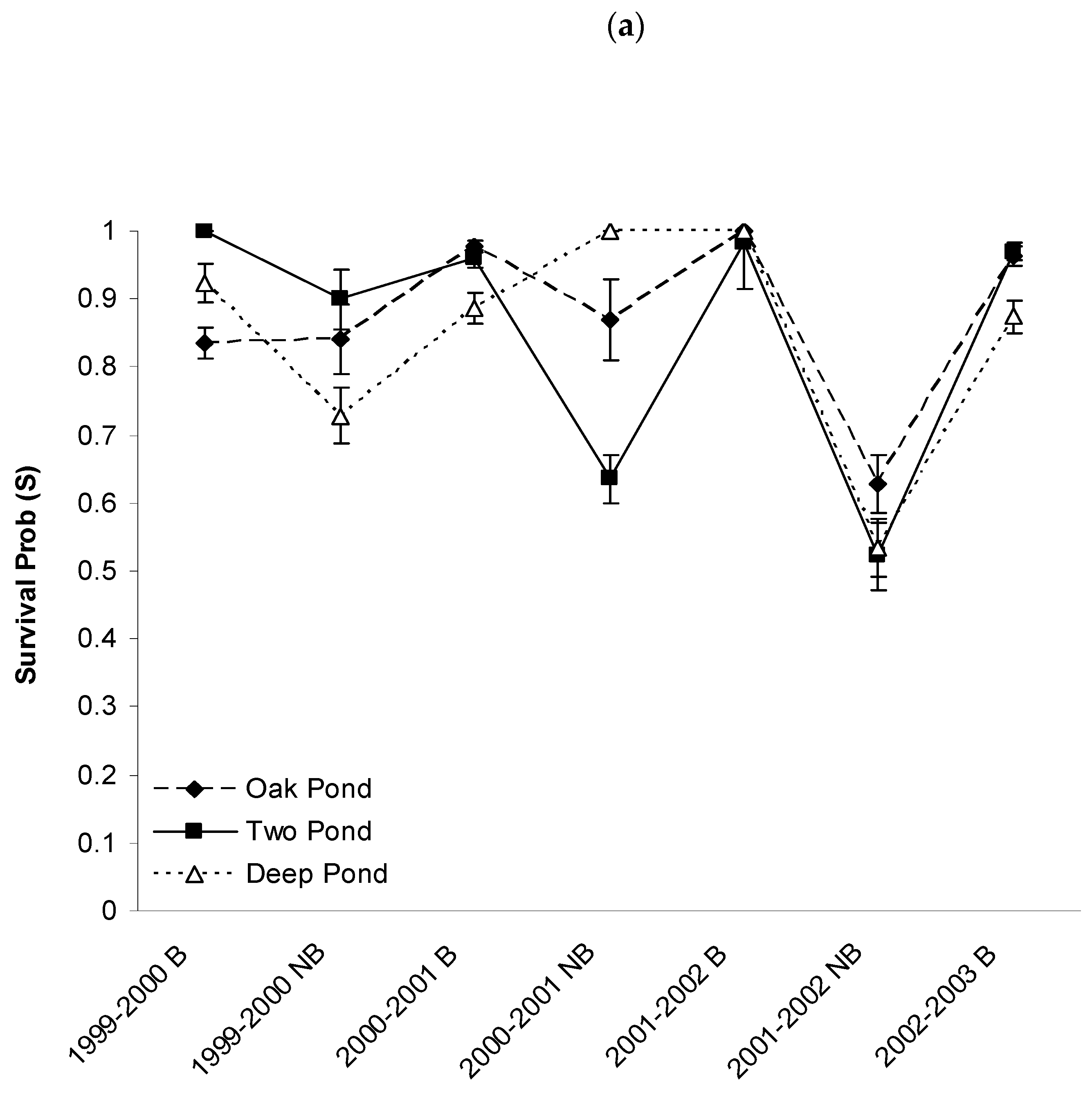
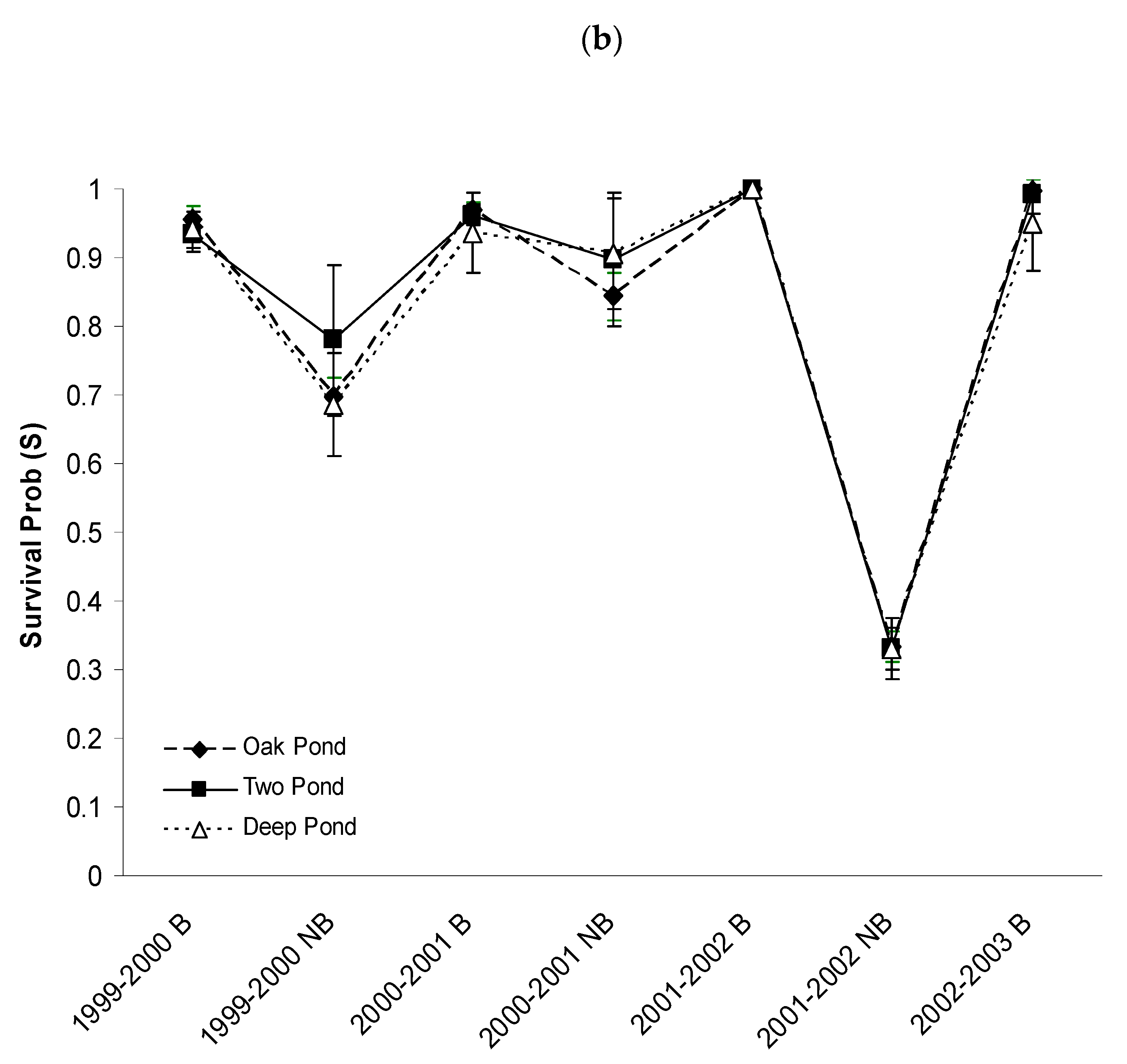
3.3. Apparent Survival Probabilities
3.4. Breeding Probabilities
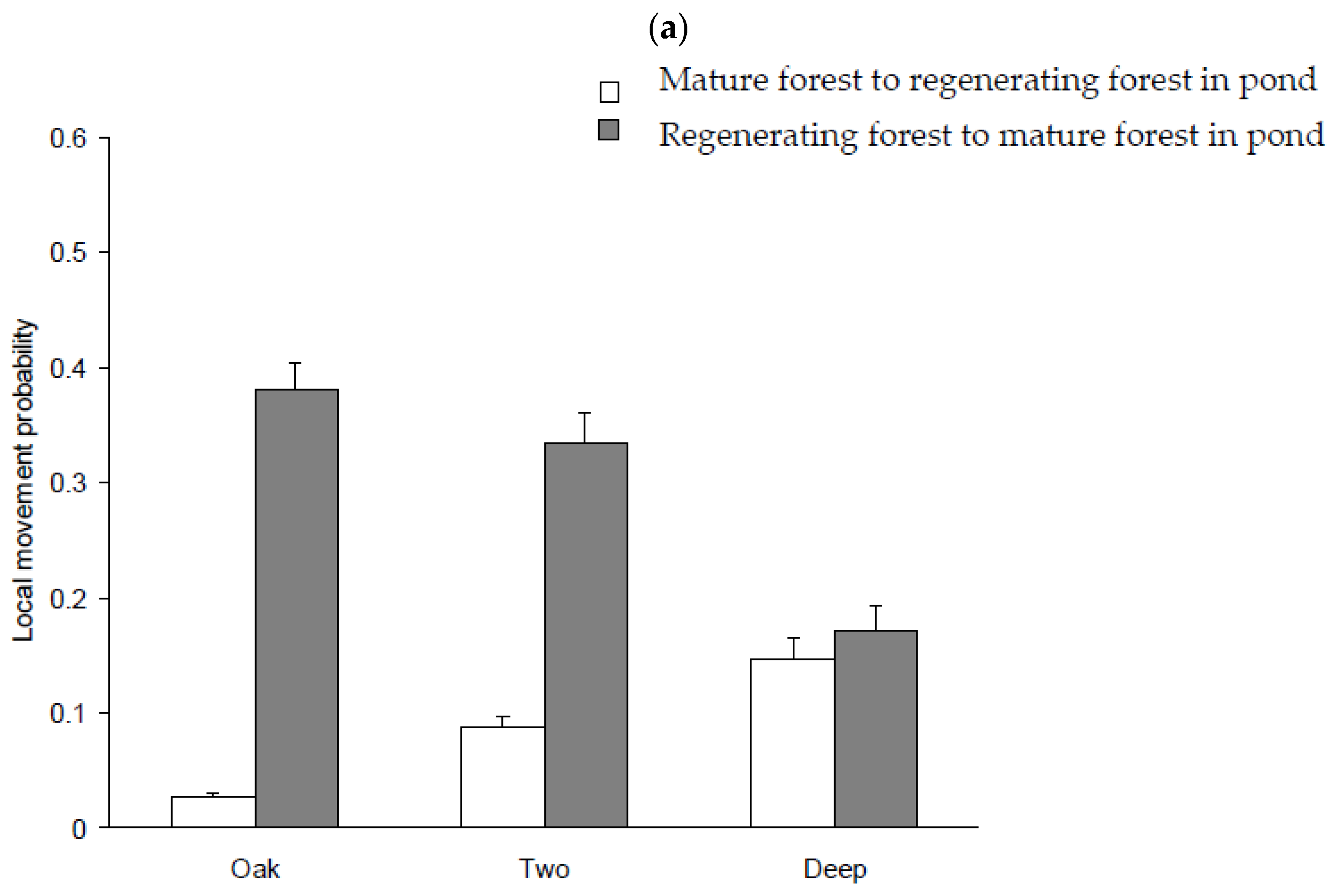
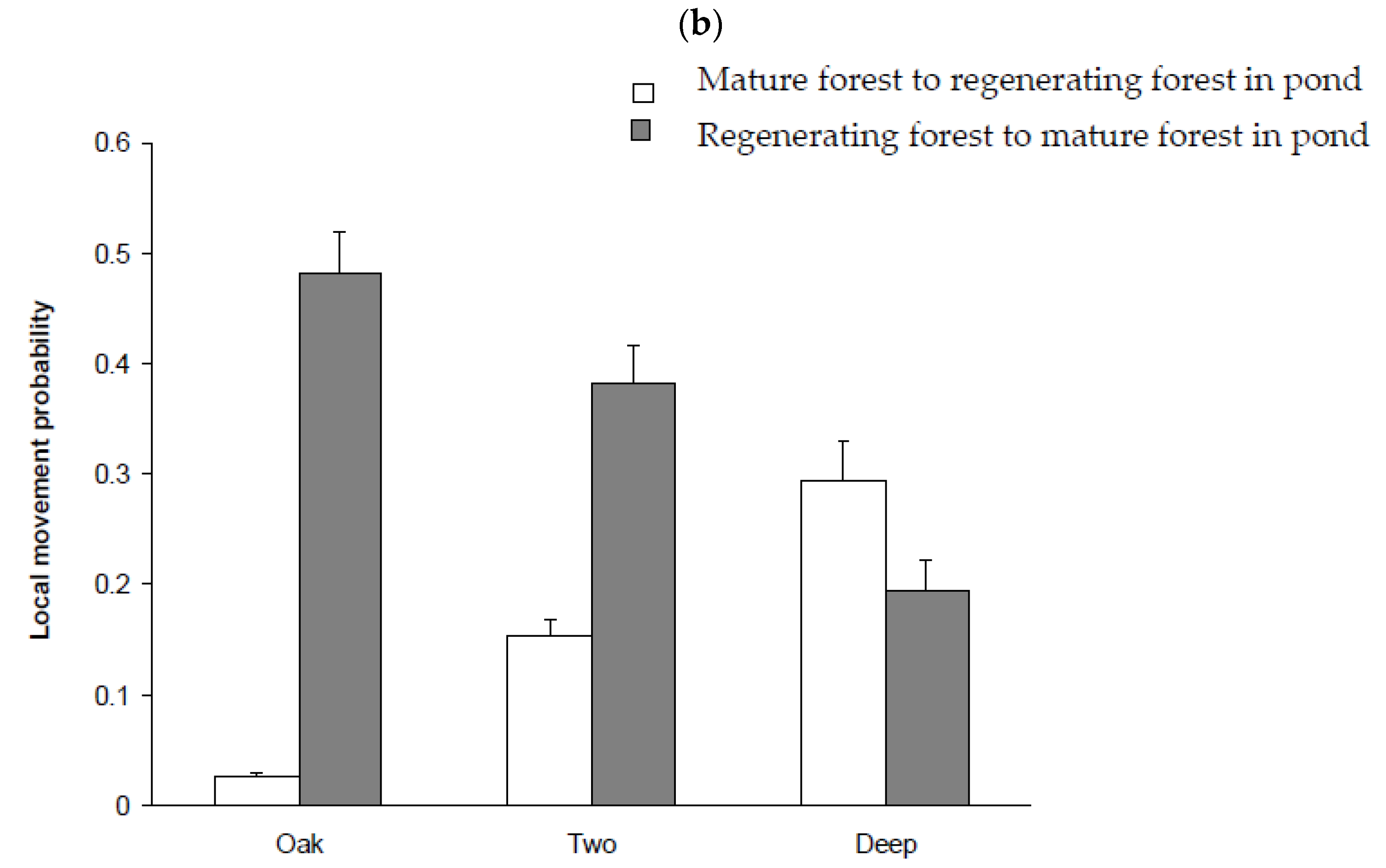
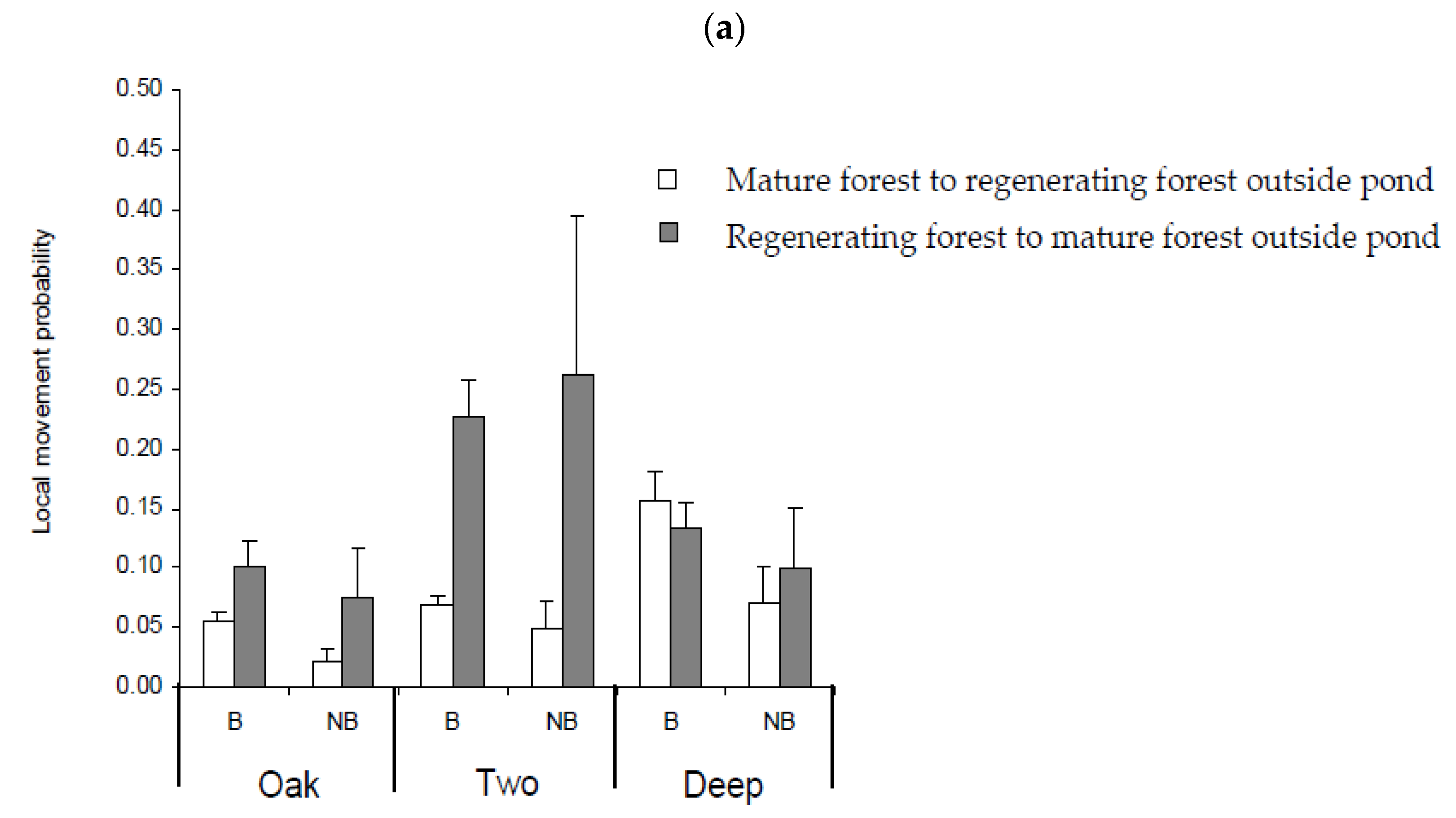
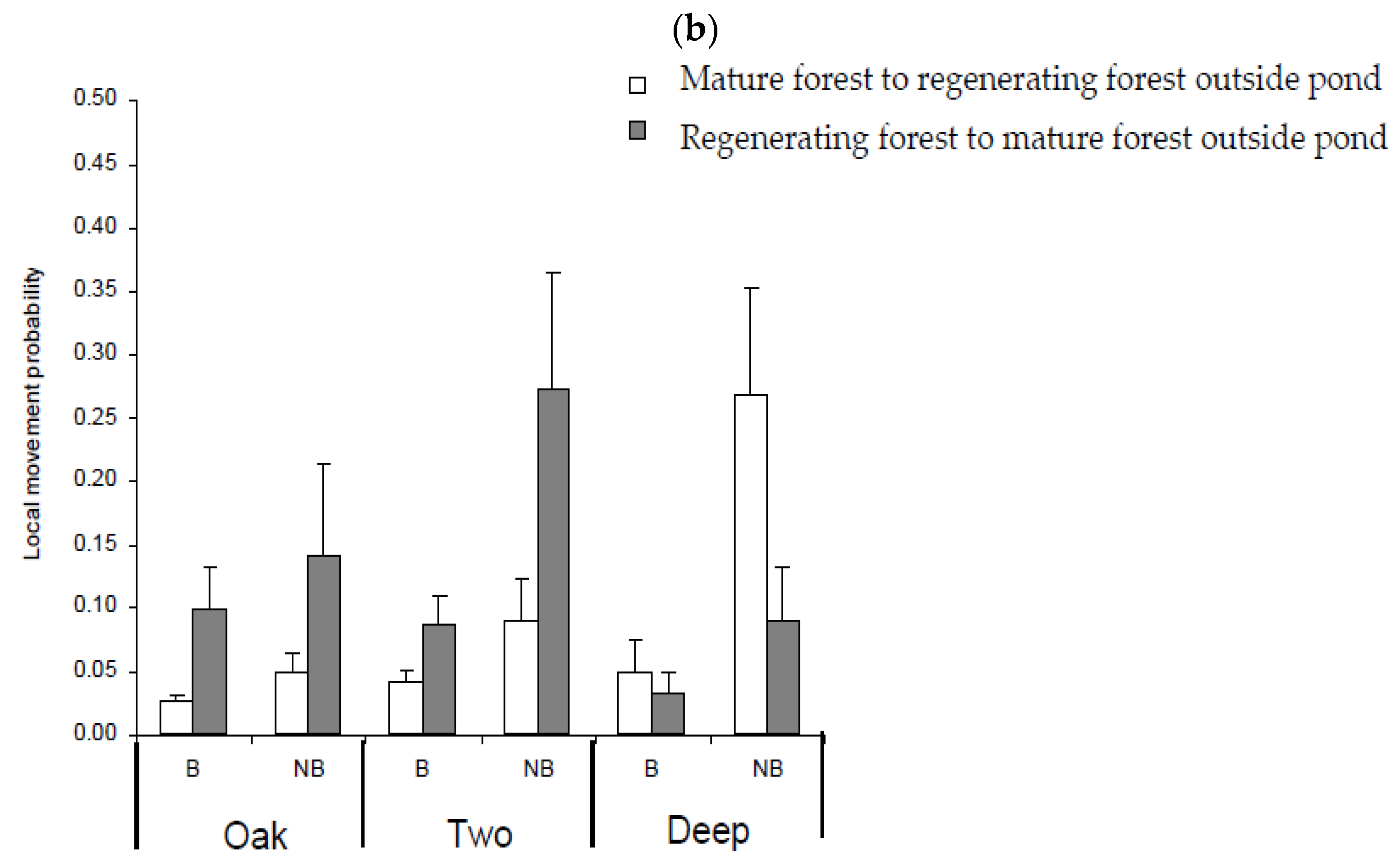
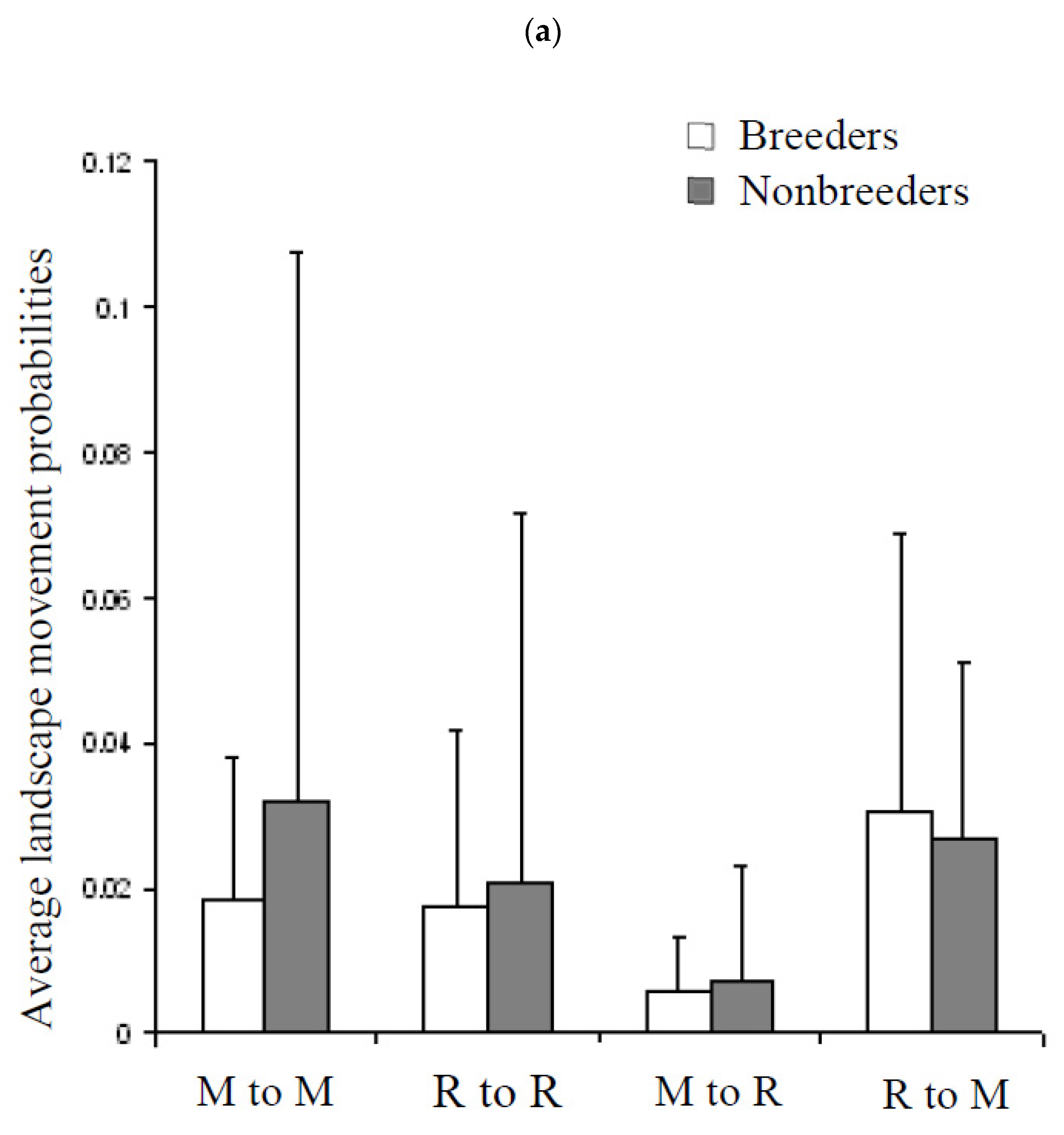
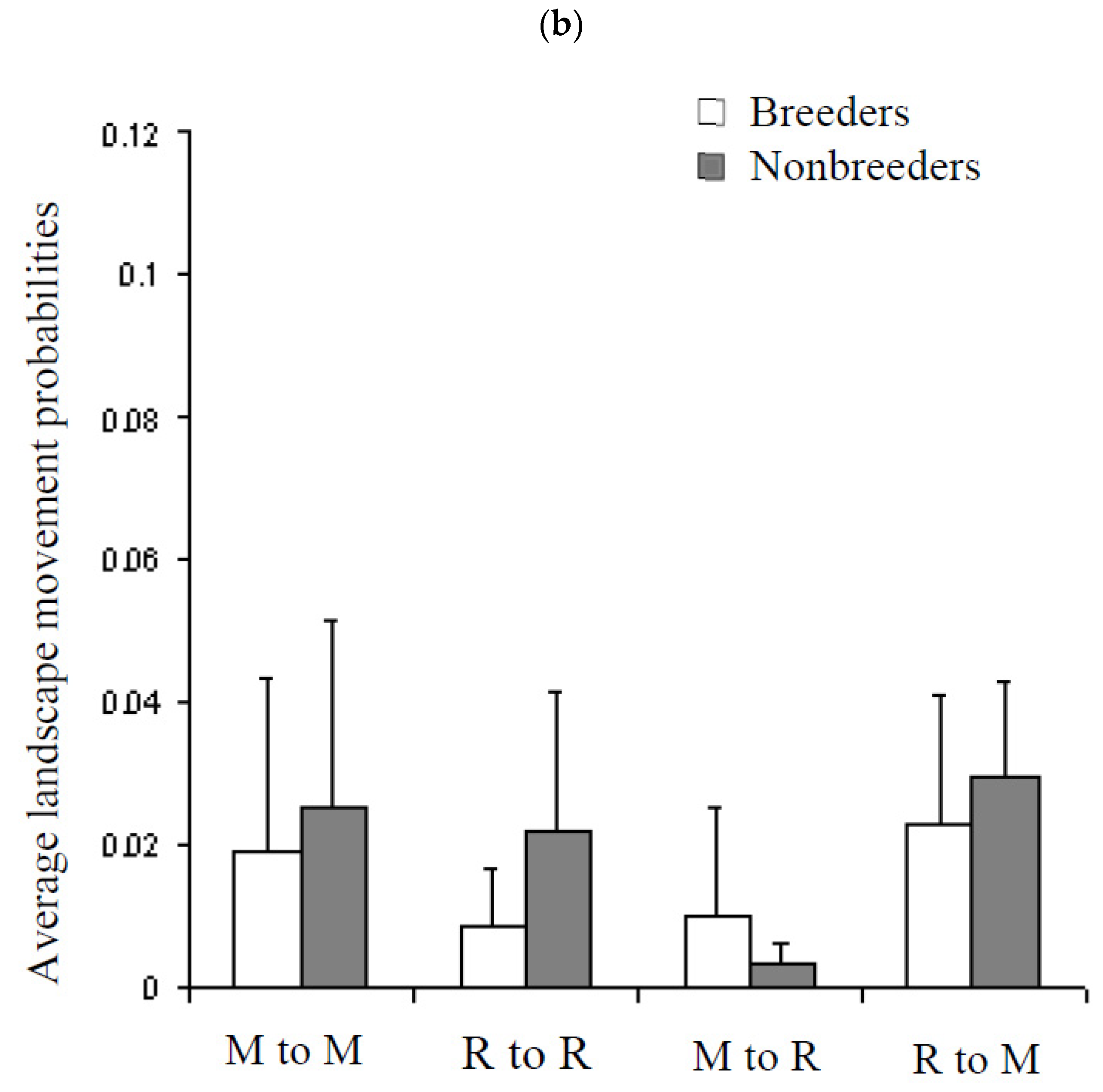
3.5. Movement Probabilities: Local and Landscape
3.6. Capture Probabilities
4. Discussion
4.1. Factors Influencing Survival Probabilities
4.2. Factors Influencing Breeding Probabilities
4.3. Factors Influencing Local and Landscape Movements
4.4. Caveats
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
| Pond (Habitat Ratio) | Sex | Year | Count Mature Forest | Count Regen. Forest | Count Ratio Mature: Regen | Residence Time |
| Oak Pond (3.1:1) | Females | 1999–2000 | 678 | 113 | 6.0:1 | 61.6 ± 6.0 |
| 2000–2001 | 704 | 131 | 5.4:1 | 63.2 ± 8.1 | ||
| 2001–2002 | 994 | 141 | 7.0:1 | 56.9 ± 5.5 | ||
| 2002–2003 | 528 | 63 | 8.4:1 | 45.5 ± 2.6 | ||
| Males | 1999–2000 | 436 | 113 | 3.9:1 | 23.3 ± 7.2 | |
| 2000–2001 | 1160 | 287 | 4.0:1 | 32.3 ± 8.3 | ||
| 2001–2002 | 1273 | 243 | 5.2:1 | 35.7 ± 1.2 | ||
| 2002–2003 | 1453 | 195 | 7.5:1 | 25.3 ± 4.6 | ||
| Pond Two (2.0:1) | Females | 1999–2000 | 359 | 105 | 3.4:1 | 36.3 ± 3.3 * |
| 2000–2001 | 552 | 181 | 3.1:1 | 54.5 ± 1.7 | ||
| 2001–2002 | 426 | 131 | 3.3:1 | 56.0 ± 1.5 | ||
| 2002–2003 | 129 | 48 | 2.7:1 | 50.9 ± 4.7 | ||
| Males | 1999–2000 | 143 | 78 | 1.8:1 | 16.1 ± 4.0 | |
| 2000–2001 | 1123 | 288 | 3.9:1 | 27.0 ± 1.6 | ||
| 2001–2002 | 592 | 190 | 3.1:1 | 34.6 ± 1.8 | ||
| 2002–2003 | 307 | 86 | 3.6:1 | 27.1 ± 2.6 | ||
| Deep Pond (1.1:1) | Females | 1999–2000 | 123 | 133 | 0.9:1 | 28.4 ± 3.7 * |
| 2000–2001 | 41 | 40 | 1.0:1 | 63.2 ± 17.0 | ||
| 2001–2002 | 130 | 121 | 1.1:1 | 68.8 ± 7.9 | ||
| 2002–2003 | 156 | 116 | 1.3:1 | 45.3 ± 2.7 | ||
| Males | 1999–2000 | 107 | 71 | 1.5:1 | 20.9 ± 3.8 | |
| 2000–2001 | 215 | 137 | 1.6:1 | 28.3 ± 3.3 | ||
| 2001–2002 | 283 | 228 | 1.2:1 | 51.9 ± 6.2 | ||
| 2002–2003 | 312 | 295 | 1.1:1 | 25.1 ± 3.6 | ||
| * Note: A hurricane filled the ponds overnight during the 1999–2000 breeding season forcing early evacuation of females in Two and Deep Ponds. | ||||||
Appendix B
| Females | |||||
| Model | QDev. | K | QAIC | ΔQAIC | w |
| S(t) ψ (full) p(t*) | 1863.88 | 121 | 13,456.36 | 0.00 | 0.53 |
| S(t×pond) ψ (br_hab×pond, loc_hab×pond, land_hab) p(t*) | 1901.35 | 103 | 13,457.01 | 0.65 | 0.38 |
| S(t×hab) ψ (br_hab×pond, loc_hab×pond, land_hab) p(t*) | 1918.81 | 96 | 13,460.18 | 3.83 | 0.08 |
| S(t×pond) ψ (full) p(t*) | 1843.02 | 135 | 13,464.22 | 7.86 | 0.01 |
| S(t×hab) ψ (full) p(t*) | 1859.81 | 128 | 13,466.64 | 10.28 | 0.00 |
| S(t×pond) ψ (br_hab×pond, loc_hab×pond, land_hab) p(ct) | 1958.01 | 85 | 13,476.98 | 20.63 | 0.00 |
| S(t) ψ (br_hab×pond, loc_hab×pond, land_hab) p(ct) | 1986.56 | 71 | 13,477.09 | 20.73 | 0.00 |
| S(t) ψ (full) p(ct) | 1927.29 | 103 | 13,482.94 | 26.59 | 0.00 |
| S(t×pond) ψ (full) p(ct) | 1899.04 | 117 | 13,483.32 | 26.96 | 0.00 |
| S(t×hab) ψ (br_hab×pond, loc_hab×pond, land_hab) p(ct) | 1983.30 | 78 | 13,488.04 | 31.69 | 0.00 |
| S(t×hab) ψ (full) p(ct) | 1923.89 | 110 | 13,493.85 | 37.5 | 0.00 |
| S(t×hab×pond) ψ (br_hab×pond, loc_hab×pond, land_hab) p(ct) | 1932.75 | 106 | 13,494.54 | 38.18 | 0.00 |
| S(t×hab×pond) ψ (full) p(ct) | 1881.78 | 138 | 13,509.15 | 52.79 | 0.00 |
| S(t) ψ (br_hab×pond, loc_hab×pond) p(t*) | 2022.90 | 81 | 13,533.73 | 77.38 | 0.00 |
| S(t×pond) ψ (br_hab×pond, loc_hab×pond) p(t*) | 2002.00 | 95 | 13,541.33 | 84.97 | 0.00 |
| S(t×hab) ψ (br_hab×pond, loc_hab×pond) p(t*) | 2018.34 | 88 | 13,543.42 | 87.06 | 0.00 |
| S(t×hab×pond) ψ (br_hab×pond, loc_hab×pond) p(t*) | 1970.83 | 116 | 13,553.06 | 96.7 | 0.00 |
| S(t) ψ(br_hab×pond, loc_hab×pond) p(ct) | 2087.72 | 63 | 13,562.03 | 105.68 | 0.00 |
| S(t×pond) ψ (br_hab×pond, loc_hab×pond) p(ct) | 2060.20 | 77 | 13,562.91 | 106.55 | 0.00 |
| S(t×hab) ψ (br_hab×pond, loc_hab×pond) p(ct) | 2083.85 | 70 | 13,572.35 | 115.99 | 0.00 |
| S(t×hab×pond) ψ (br_hab×pond, loc_hab×pond) p(ct) | 2029.39 | 98 | 13,574.84 | 118.48 | 0.00 |
| S(t×pond)ψ (br_hab, loc_hab, land_hab) p(t*) | 2186.68 | 83 | 13,701.58 | 245.22 | 0.00 |
| S(t) ψ (br_hab, loc_hab, land_hab) p(t*) | 2223.17 | 69 | 13,709.64 | 253.29 | 0.00 |
| S(t×hab) ψ (br_hab, loc_hab, land_hab) p(t*) | 2217.70 | 76 | 13,718.37 | 262.02 | 0.00 |
| S(t×pond) ψ (br_hab, loc_hab, land_hab) p(ct) | 2249.03 | 65 | 13,727.39 | 271.03 | 0.00 |
| S(t×hab×pond) ψ (br_hab, loc_hab, land_hab) p(ct) | 2219.11 | 86 | 13,740.12 | 283.76 | 0.00 |
| S(t) ψ (br_hab, loc_hab, land_hab) p(ct) | 2297.69 | 51 | 13,747.73 | 291.37 | 0.00 |
| S(t×hab) ψ (br_hab, loc_hab, land_hab) p(ct) | 2294.54 | 58 | 13,758.73 | 302.37 | 0.00 |
| S(t×pond) ψ (br_hab, loc_hab) p(t*) | 2272.59 | 75 | 13,771.24 | 314.88 | 0.00 |
| S(t) ψ (br_hab, loc_hab) p(t*) | 2308.11 | 61 | 13,778.37 | 322.01 | 0.00 |
| S(t×hab×pond) ψ (br_hab, loc_hab) p(t*) | 2245.64 | 96 | 13,787.01 | 330.66 | 0.00 |
| S(t×hab) ψ (br_hab, loc_hab) p(t*) | 2302.80 | 68 | 13,787.25 | 330.89 | 0.00 |
| S(t×pond) ψ (br_hab, loc_hab) p(ct) | 2339.46 | 57 | 13,801.62 | 345.27 | 0.00 |
| S(t×hab×pond) ψ (br_hab, loc_hab) p(ct) | 2309.37 | 78 | 13,814.11 | 357.75 | 0.00 |
| S(t) ψ (br_hab, loc_hab) p(ct) | 2388.09 | 43 | 13,821.97 | 365.62 | 0.00 |
| S(t×hab) ψ (br_hab, loc_hab) p(ct) | 2385.50 | 50 | 13,833.52 | 377.16 | 0.00 |
| S(t) ψ (br_pond, loc_pond) p(t*) | 2472.32 | 60 | 13,940.55 | 484.20 | 0.00 |
| S(t×pond) ψ (br_pond, loc_pond) p(t*) | 2447.43 | 74 | 13,944.04 | 487.68 | 0.00 |
| S(t×hab) ψ (br_pond, loc_pond) p(t*) | 2461.89 | 67 | 13,944.31 | 487.95 | 0.00 |
| S(t×hab×pond) ψ (br_pond, loc_pond) p(t*) | 2415.67 | 95 | 13,955.00 | 498.64 | 0.00 |
| S(t×pond) ψ (br_pond, loc_pond) p(ct) | 2526.72 | 56 | 13,986.86 | 530.50 | 0.00 |
| S(t×hab×pond) ψ (br_pond, loc_pond) p(ct) | 2489.20 | 77 | 13,991.91 | 535.56 | 0.00 |
| S(t×hab) ψ (br_pond, loc_pond) p(ct) | 2596.43 | 46 | 14,036.36 | 580.01 | 0.00 |
| S(t) ψ (br_pond, loc_pond) p(ct) | 2607.31 | 44 | 14,043.21 | 586.86 | 0.00 |
| S(t×pond) ψ (.) p(t*) | 2579.46 | 64 | 14,055.79 | 599.44 | 0.00 |
| S(t×pond) ψ (.) p(ct) | 2659.69 | 46 | 14,099.63 | 643.27 | 0.00 |
| S(t×hab) ψ (.) p(t*) | 2634.55 | 62 | 14,106.83 | 650.48 | 0.00 |
| S(t) ψ (.) p(t*) | 2655.09 | 55 | 14,113.21 | 656.85 | 0.00 |
| S(t×hab×pond) ψ (.) p(ct) | 2632.84 | 67 | 14,115.26 | 658.90 | 0.00 |
| S(t×hab) ψ (.) p(ct) | 2731.66 | 44 | 14,167.56 | 711.20 | 0.00 |
| S(t) ψ (.) p(ct) | 2752.04 | 37 | 14,173.83 | 717.47 | 0.00 |
| S(t×hab×pond) ψ (br_hab×pond, loc_hab×pond, land_hab) p(t*) | DNC | 132 | |||
| S(t×hab×pond) ψ (br_hab, loc_hab, land_hab) p(t*) | DNC | 104 | |||
| S(t×hab) ψ (br_pond, loc_pond) p(ct) | SING | 52 | |||
| S(t×hab×pond) ψ (.) p(t*) | SING | 85 | |||
| Males | |||||
| Model | QDev. | K | QAIC | ΔQAIC | w |
| S(t×pond) ψ (full) p(t*) | 3496.27 | 135 | 22,670.49 | 0.00 | 1.00 |
| S(t×pond) ψ (full) p(ct) | 3561.10 | 117 | 22,698.71 | 28.23 | 0.00 |
| S(t×hab×pond) ψ (full) p(ct) | 3525.85 | 138 | 22,706.17 | 35.69 | 0.00 |
| S(t×hab×pond) ψ (br_hab×pond, loc_hab×pond, land_hab) p(t*) | 3577.68 | 132 | 22,745.79 | 75.30 | 0.00 |
| S(t) ψ (full) p(t*) | 3636.18 | 121 | 22,781.92 | 111.43 | 0.00 |
| S(t×pond) ψ (br_hab×pond, loc_hab×pond, land_hab) p(t*) | 3675.87 | 103 | 22,785.07 | 114.59 | 0.00 |
| S(t×hab) ψ (full) p(t*) | 3626.03 | 128 | 22,786.00 | 115.51 | 0.00 |
| S(t×pond) ψ (br_hab×pond, loc_hab×pond, land_hab) p(ct) | 3740.90 | 85 | 22,813.65 | 143.17 | 0.00 |
| S(t×hab×pond) ψ (br_hab×pond, loc_hab×pond, land_hab) p(ct) | 3709.28 | 106 | 22,824.56 | 154.08 | 0.00 |
| S(t) ψ (full) p(ct) | 3719.76 | 103 | 22,828.96 | 158.47 | 0.00 |
| S(t×hab) ψ (full) p(ct) | 3710.51 | 110 | 22,833.91 | 163.43 | 0.00 |
| S(t×hab) ψ (br_hab×pond, loc_hab×pond, land_hab) p(t*) | 3809.46 | 96 | 22,904.48 | 234.00 | 0.00 |
| S(t×pond) ψ (br_hab×pond, loc_hab×pond) p(t*) | 3833.59 | 95 | 22,926.58 | 256.10 | 0.00 |
| S(t×hab×pond) ψ (br_hab×pond, loc_hab×pond) p(t*) | 3792.41 | 116 | 22,927.99 | 257.50 | 0.00 |
| S(t) ψ (br_hab×pond, loc_hab×pond, land_hab) p(ct) | 3902.35 | 71 | 22,946.82 | 276.33 | 0.00 |
| S(t×hab) ψ (br_hab×pond, loc_hab×pond, land_hab) p(ct) | 3893.56 | 78 | 22,952.16 | 281.68 | 0.00 |
| S(t×pond) ψ (br_hab×pond, loc_hab×pond) p(ct) | 3899.58 | 77 | 22,956.16 | 285.68 | 0.00 |
| S(t×hab×pond) ψ (br_hab×pond, loc_hab×pond) p(ct) | 3863.20 | 98 | 22,962.27 | 291.78 | 0.00 |
| S(t×hab×pond)ψ (br_hab, loc_hab, land_hab) p(t*) | 3887.86 | 104 | 22,999.09 | 328.61 | 0.00 |
| S(t×pond) ψ (br_hab, loc_hab, land_hab) p(t*) | 3944.37 | 83 | 23,013.08 | 342.59 | 0.00 |
| S(t) ψ (br_hab×pond, loc_hab×pond) p(t*) | 3975.31 | 81 | 23,039.97 | 369.49 | 0.00 |
| S(t×hab×pond) ψ (br_hab, loc_hab, land_hab) p(ct) | 3969.93 | 86 | 23,044.71 | 374.22 | 0.00 |
| S(t×hab) ψ (br_hab×pond, loc_hab×pond) p(t*) | 3966.55 | 88 | 23,045.37 | 374.89 | 0.00 |
| S(t×pond) ψ (br_hab, loc_hab, land_hab) p(ct) | 4017.38 | 65 | 23,049.74 | 379.25 | 0.00 |
| S(t) ψ (br_hab×pond, loc_hab×pond) p(ct) | 4059.68 | 63 | 23,088.00 | 417.52 | 0.00 |
| S(t×hab) ψ (br_hab×pond, loc_hab×pond) p(ct) | 4051.53 | 70 | 23,093.98 | 423.49 | 0.00 |
| S(t×hab×pond) ψ (br_hab, loc_hab) p(t*) | 4036.86 | 96 | 23,131.88 | 461.39 | 0.00 |
| S(t) ψ (br_hab, loc_hab, land_hab) p(t*) | 4091.76 | 69 | 23,132.19 | 461.70 | 0.00 |
| S(t×hab) ψ (br_hab, loc_hab, land_hab) p(t*) | 4081.84 | 76 | 23,136.41 | 465.92 | 0.00 |
| S(t×pond) ψ (br_hab, loc_hab) p(t*) | 4096.88 | 75 | 23,149.43 | 478.94 | 0.00 |
| S(t×hab×pond) ψ (br_hab, loc_hab) p(ct) | 4120.71 | 78 | 23,179.32 | 508.83 | 0.00 |
| S(t) ψ (br_hab, loc_hab, land_hab) p(ct) | 4177.04 | 51 | 23,181.18 | 510.69 | 0.00 |
| S(t×hab) ψ (br_hab, loc_hab, land_hab) p(ct) | 4167.59 | 58 | 23,185.83 | 515.35 | 0.00 |
| S(t×pond) ψ (br_hab, loc_hab) p(ct) | 4171.42 | 57 | 23,187.65 | 517.17 | 0.00 |
| S(t) ψ (br_hab, loc_hab) p(t*) | 4244.41 | 61 | 23,268.70 | 598.21 | 0.00 |
| S(t×hab) ψ (br_hab, loc_hab) p(t*) | 4235.05 | 68 | 23,273.46 | 602.98 | 0.00 |
| S(t) ψ (br_hab, loc_hab) p(ct) | 4330.29 | 43 | 23,318.33 | 647.85 | 0.00 |
| S(t×hab) ψ (br_hab, loc_hab) p(ct) | 4321.36 | 50 | 23,323.49 | 653.00 | 0.00 |
| S(t×hab×pond) ψ (br_pond, loc_pond) p(t*) | 4445.26 | 95 | 23,538.26 | 867.77 | 0.00 |
| S(t×pond) ψ (br_pond, loc_pond) p(t*) | 4516.38 | 74 | 23,566.90 | 896.42 | 0.00 |
| S(t×hab×pond) ψ (br_pond, loc_pond) p(ct) | 4527.47 | 77 | 23,584.06 | 913.57 | 0.00 |
| S(t×pond) ψ (br_pond, loc_pond) p(ct) | 4593.59 | 56 | 23,607.81 | 937.32 | 0.00 |
| S(t×hab×pond) ψ (.) p(t*) | 4575.75 | 85 | 23,648.50 | 978.02 | 0.00 |
| S(t×pond) ψ (.) p(t*) | 4642.40 | 64 | 23,672.74 | 1002.26 | 0.00 |
| S(t×hab) ψ (br_pond, loc_pond) p(t*) | 4639.29 | 67 | 23,675.68 | 1005.20 | 0.00 |
| S(t) ψ (br_pond, loc_pond) p(t*) | 4660.21 | 60 | 23,682.49 | 1012.00 | 0.00 |
| S(t×hab×pond) ψ (.) p(ct) | 4669.04 | 67 | 23,705.43 | 1034.95 | 0.00 |
| S(t×pond)ψ (.) p(ct) | 4743.10 | 46 | 23,737.18 | 1066.70 | 0.00 |
| S(t×hab) ψ (br_pond, loc_pond) p(ct) | 4785.40 | 52 | 23,791.56 | 1121.07 | 0.00 |
| S(t) ψ (br_pond, loc_pond) p(ct) | 4800.41 | 44 | 23,792.48 | 1121.99 | 0.00 |
| S(t×hab) ψ (.) p(t*) | 4824.65 | 62 | 23,850.96 | 1180.47 | 0.00 |
| S(t) ψ (.) p(t*) | 4841.00 | 55 | 23,853.20 | 1182.72 | 0.00 |
| S(t×hab) ψ (.) p(ct) | 4926.95 | 44 | 23,917.00 | 1246.52 | 0.00 |
| S(t) ψ (.) p(ct) | 4942.01 | 37 | 23,917.99 | 1247.50 | 0.00 |
| S(t×hab) ψ (br_pond, loc_pond) p(ct) | DNC | 46 | |||
Appendix C

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Males | ||||||||||||
| Oak Pond | Pond Two | Deep Pond | ||||||||||
| Breeding Season | Non-breeding Season | Breeding Season | Non-breeding Season | Breeding Season | Non-breeding Season | |||||||
| Year | ||||||||||||
| 1999–2000 | 0.88 | 0.02 | 0.84 | 0.05 | 1.00 | - | 0.90 | 0.04 | 0.92 | 0.03 | 0.73 | 0.04 |
| 2000–2001 | 0.98 | 0.01 | 0.87 | 0.06 | 0.96 | 0.01 | 0.64 | 0.04 | 0.89 | 0.02 | 1.00 | - |
| 2001–2002 | 1.00 | - | 0.63 | 0.04 | 0.98 | 0.07 | 0.52 | 0.05 | 1.00 | - | 0.54 | 0.04 |
| 2002–2003 | 0.96 | 0.01 | 0.97 | 0.01 | 0.87 | 0.02 | ||||||
| Females | ||||||||||||
| Oak Pond | Pond Two | Deep Pond | ||||||||||
| Seasons: | Breeding Season | Non-breeding Season | Breeding Season | Non-breeding Season | Breeding Season | Non-breeding Season | ||||||
| Year | ||||||||||||
| 1999–2000 | 0.96 | 0.02 | 0.70 | 0.03 | 0.93 | 0.02 | 0.78 | 0.11 | 0.94 | 0.03 | 0.69 | 0.08 |
| 2000–2001 | 0.97 | 0.01 | 0.84 | 0.04 | 0.96 | 0.02 | 0.90 | 0.10 | 0.94 | 0.06 | 0.91 | 0.08 |
| 2001–2002 | 1.00 | - | 0.33 | 0.02 | 1.00 | - | 0.33 | 0.03 | 1.00 | - | 0.33 | 0.04 |
| 2002–2003 | 0.99 | 0.02 | 0.99 | 0.03 | 0.95 | 0.07 | ||||||
| Males: Monthly Survival | ||||||||||||
| Oak Pond | Pond Two | Deep Pond | ||||||||||
| Breeding Season | Non-breeding Season | Breeding Season | Non-breeding Season | Breeding Season | Non-breeding Season | |||||||
| Year | ||||||||||||
| 1999–2000 | 0.85 | 0.027 | 0.98 | 0.004 | 1.00 | - | 0.99 | 0.003 | 0.89 | 0.057 | 0.97 | 0.003 |
| 2000–2001 | 0.98 | 0.009 | 0.99 | 0.005 | 0.96 | 0.011 | 0.96 | 0.002 | 0.88 | 0.021 | 1.00 | - |
| 2001–2002 | 1.00 | - | 0.96 | 0.002 | 0.98 | 0.060 | 0.94 | 0.003 | 1.00 | - | 0.94 | 0.004 |
| 2002–2003 | 0.95 | 0.012 | 0.97 | 0.011 | 0.85 | 0.025 | ||||||
| Females: Monthly Survival | ||||||||||||
| Oak Pond | Pond Two | Deep Pond | ||||||||||
| Breeding Season | Non-breeding Season | Breeding Season | Non-breeding Season | Breeding Season | Non-breeding Season | |||||||
| Year | ||||||||||||
| 1999–2000 | 0.98 | 0.009 | 0.97 | 0.002 | 0.94 | 0.016 | 0.98 | 0.008 | 0.94 | 0.032 | 0.97 | 0.005 |
| 2000–2001 | 0.99 | 0.005 | 0.98 | 0.003 | 0.98 | 0.011 | 0.99 | 0.009 | 0.97 | 0.0628 | 0.99 | 0.007 |
| 2001–2002 | 1.00 | - | 0.90 | 0.001 | 1.00 | - | 0.90 | 0.001 | 1.00 | - | 0.89 | 0.001 |
| 2002–2003 | 0.99 | 0.013 | 0.99 | 0.018 | 0.97 | 0.046 | ||||||
Appendix D
| Male: LocalLandscape Movement Probabilities, (SE()) | |||||||
| Breeders | |||||||
| Oak Mature | Oak Reg. | Two Mature | Two Reg. | Deep Mature | Deep Reg. | ||
| Oak Mature | 0.055 (0.007) | 0.015 (0.003) | 0.000 (-) | 0.002 (0.001) | 0.006 (0.002) | ||
| Oak Reg. | 0.101 (0.021) | 0.003 (0.003) | 0.000 (-) | 0.000 (-) | 0.000 (-) | ||
| Two Mature | 0.054 (0.008) | 0.004 (0.002) | 0.067 (0.009) | 0.004 (0.002) | 0.020 (0.005) | ||
| Two Reg. | 0.006 (0.006) | 0.000 (-) | 0.226 (0.031) | 0.018 (0.009) | 0.057 (0.016) | ||
| Deep Mature | 0.006 (0.004) | 0.000 (-) | 0.028 (0.009) | 0.005 (0.004) | 0.156 (0.024) | ||
| Deep Reg. | 0.064 (0.014) | 0.008 (0.005) | 0.092 (0.017) | 0.038 (0.011) | 0.133 (0.020) | ||
| Non-Breeders | |||||||
| Oak Mature | Oak Reg. | Two Mature | Two Reg. | Deep Mature | Deep Reg. | ||
| Oak Mature | 0.021 (0.011) | 0.001 (0.003) | 0.000 (-) | 0.000 (-) | 0.000 (-) | ||
| Oak Reg. | 0.075 (0.040) | 0.000 (-) | 0.000 (-) | 0.000 (-) | 0.000 (-) | ||
| Two Mature | 0.186 (0.053) | 0.004 (0.007) | 0.048 (0.023) | 0.000 (-) | 0.039 (0.020) | ||
| Two Reg. | 0.026 (0.033) | 0.000 (-) | 0.262 (0.132) | 0.048 (0.046) | 0.125 (0.079) | ||
| Deep Mature | 0.006 (0.010) | 0.000 (-) | 0.000 (-) | 0.000 (-) | 0.069 (0.031) | ||
| Deep Reg. | 0.059 (0.037) | 0.000 (-) | 0.029 (0.026) | 0.000 (-) | 0.099 (0.050) | ||
| Female: Local and Landscape Movement Probabilities, (SE()) | |||||||
| Breeders | |||||||
| Oak Mature | Oak Reg. | Two Mature | Two Reg. | Deep Mature | Deep Reg. | ||
| Oak Mature | 0.026 (0.005) | 0.036 (0.009) | 0.002 (0.002) | 0.003 (0.002) | 0.004 (0.002) | ||
| Oak Reg. | 0.099 (0.033) | 0.002 (0.004) | 0.000 (-) | 0.022 (0.014) | 0.010 (0.008) | ||
| Two Mature | 0.061 (0.019) | 0.005 (0.003) | 0.041 (0.010) | 0.002 (0.002) | 0.006 (0.004) | ||
| Two Reg. | 0.007 (0.006) | 0.000 (-) | 0.086 (0.024) | 0.019 (0.014) | 0.016 (0.009) | ||
| Deep Mature | 0.010 (0.011) | 0.005 (0.006) | 0.005 (0.006) | 0.002 (0.002) | 0.048 (0.025) | ||
| Deep Reg. | 0.039 (0.017) | 0.006 (0.008) | 0.048 (0.023) | 0.020 (0.012) | 0.032 (0.016) | ||
| Non-Breeders | |||||||
| Oak Mature | Oak Reg. | Two Mature | Two Reg. | Deep Mature | Deep Reg. | ||
| Oak Mature | 0.049 (0.015) | 0.031 (0.016) | 0.000 (-) | 0.010 (0.006) | 0.003 (0.003) | ||
| Oak Reg. | 0.142 (0.071) | 0.017 (0.023) | 0.000 (-) | 0.017 (0.021) | 0.015 (0.019) | ||
| Two Mature | 0.076 (0.039) | 0.000 (-) | 0.089 (0.035) | 0.010 (0.008) | 0.004 (0.005) | ||
| Two Reg. | 0.021 (0.019) | 0.000 (-) | 0.272 (0.094) | 0.031 (0.021) | 0.044 (0.028) | ||
| Deep Mature | 0.019 (0.023) | 0.002 (0.003) | 0.004 (0.006) | 0.009 (0.014) | 0.268 (0.084) | ||
| Deep Reg. | 0.048 (0.032) | 0.030 (0.020) | 0.043 (0.026) | 0.042 (0.027) | 0.089 (0.043) | ||
Appendix E
| Males | ||||||||||||
| Oak Mature | Oak Reg. | Two Mature | Two Reg. | Deep Mature | Deep Reg. | |||||||
| Occasion | ||||||||||||
| t = 2 | 0.83 | 0.03 | 0.72 | 0.07 | 0.89 | 0.03 | 0.80 | 0.07 | 0.97 | 0.03 | 0.93 | 0.05 |
| t = 3 | 0.98 | 0.01 | 0.98 | 0.02 | 0.98 | 0.01 | 0.99 | 0.02 | 0.98 | 0.02 | 0.93 | 0.06 |
| t = 4 | 0.91 | 0.01 | 0.96 | 0.03 | 0.87 | 0.02 | 0.92 | 0.03 | 0.94 | 0.03 | 0.95 | 0.04 |
| t = 4 | 0.94 | 0.02 | 0.96 | 0.03 | 0.96 | 0.02 | 0.94 | 0.04 | 0.97 | 0.03 | 0.96 | 0.04 |
| t = 6 | 0.27 | 0.02 | 0.47 | 0.05 | 0.34 | 0.04 | 0.59 | 0.06 | 0.28 | 0.03 | 0.20 | 0.03 |
| t = 7&8 | 0.95 | 0.01 | 0.95 | 0.03 | 0.99 | 0.01 | 0.97 | 0.02 | 0.94 | 0.03 | 0.95 | 0.02 |
| Females | ||||||||||||
| Oak Mature | Oak Reg. | Two Mature | Two Reg. | Deep Mature | Deep Reg. | |||||||
| Occasion | ||||||||||||
| t = 2 | 0.89 | 0.02 | 0.71 | 0.09 | 0.91 | 0.03 | 0.88 | 0.08 | 0.90 | 0.07 | 0.88 | 0.06 |
| t = 3 | 0.98 | 0.01 | 1.00 | - | 0.94 | 0.03 | 0.89 | 0.08 | 0.42 | 0.28 | 0.33 | 0.23 |
| t = 4 | 0.96 | 0.01 | 1.00 | - | 0.91 | 0.02 | 0.92 | 0.05 | 0.98 | 0.09 | 0.83 | 0.10 |
| t = 4 | 0.98 | 0.01 | 1.00 | - | 0.98 | 0.03 | 0.94 | 0.10 | 1.00 | - | 0.96 | 0.04 |
| t = 6 | 0.28 | 0.02 | 0.34 | 0.07 | 0.29 | 0.03 | 0.33 | 0.06 | 0.68 | 0.07 | 0.78 | 0.06 |
| t = 7&8 | 0.87 | 0.02 | 0.88 | 0.07 | 0.97 | 0.03 | 0.93 | 0.06 | 0.90 | 0.08 | 0.80 | 0.08 |
References
- Barnes, B.V.; Zak, D.R.; Denton, S.R.; Spurr, S.H. Forest Ecology; Wiley Publishing: Indianapolis, IN, USA, 1998. [Google Scholar]
- Matlack, G. Plant Demography, Land-Use History, and the Commercial Use of Forests. Conserv. Biol. 1994, 8, 298–299. [Google Scholar] [CrossRef]
- Skelly, D.K.; Werner, E.E.; Cortwright, S.A. Long-term distributional dynamics of a Michigan amphibian assemblage. Ecology 1999, 80, 2326–2337. [Google Scholar] [CrossRef]
- Cayuela, H.; Valenzuela-Sánchez, A.; Teulier, L.; Martínez-Solano, Í.; Léna, J.P.; Merilä, J.; Muths, E.; Shine, R.; Quay, L.; Denoël, M.; et al. Determinants and consequences of dispersal in vertebrates with complex life cycles: A review of pond-breeding amphibians. Q. Rev. Biol. 2020, 95, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Stuart, S.N.; Chanson, J.S.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.L.; Fischman, D.L.; Waller, R.W. Status and trends of amphibian declines and extinctions worldwide. Science 2004, 306, 1783–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, M.J.; Miller, D.A.W.; Muths, E.; Corn, P.S.; Grant, E.H.C.; Bailey, L.L.; Fellers, G.M.; Fisher, R.N.; Sadinski, W.J.; Waddle, H.; et al. Trends in Amphibian Occupancy in the United States. PLoS ONE 2013, 8, e64347. [Google Scholar] [CrossRef]
- Grant, E.H.C.; Miller, D.A.; Muths, E. A Synthesis of Evidence of Drivers of Amphibian Declines. Herpetologica 2020, 76, 101–107. [Google Scholar] [CrossRef]
- Hoffmann, M.; Hilton-Taylor, C.; Angulo, A.; Böhm, M.; Brooks, T.M.; Butchart, S.H.M.; Carpenter, K.E.; Chanson, J.; Collen, B.; Cox, N.A.; et al. The Impact of Conservation on the Status of the World’s Vertebrates. Science 2010, 330, 1503–1509. [Google Scholar] [CrossRef] [Green Version]
- Petrovan, S.O.; Schmidt, B.R. Volunteer conservation action data reveals large-scale and long-term negative population trends of a widespread amphibian, the common toad (Bufo bufo). PLoS ONE 2016, 11, e0161943. [Google Scholar] [CrossRef] [Green Version]
- Lawler, J.J.; Aukema, J.E.; Grant, J.B.; Halpern, B.S.; Kareiva, P.; Nelson, C.R.; Ohleth, K.; Olden, J.D.; Schlaepfer, M.A.; Silliman, B.R.; et al. Conservation science: A 20-year report card. Front. Ecol. Environ. 2006, 4, 473–480. [Google Scholar] [CrossRef] [Green Version]
- Todd, B.D.; Rothermel, B.B. Assessing quality of clearcut habitats for amphibians: Effects on abundances versus vital rates in the southern toad (Bufo terrestris). Biol. Conserv. 2006, 133, 178–185. [Google Scholar] [CrossRef]
- Cayuela, H.; Besnard, A.; Quay, L.; Helder, R.; Léna, J.P.; Joly, P.; Pichenot, J. Demographic response to patch destruction in a spatially structured amphibian population. J. Appl. Ecol. 2018, 55, 2204–2215. [Google Scholar] [CrossRef]
- Muths, E.; Chambert, T.; Schmidt, B.R.; Miller, D.A.W.; Hossack, B.R.; Joly, P.; Grolet, O.; Green, D.M.; Pilliod, D.S.; Cheylan, M.; et al. Heterogeneous responses of temperate-zone amphibian populations to climate change complicates conservation planning. Sci. Rep. 2017, 7, 17102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biek, R.; Funk, W.C.; Maxell, B.A.; Mills, L.S. What is missing inamphibian decline research: Insights from ecological sensitivity analysis. Conserv. Biol. 2002, 16, 728–734. [Google Scholar] [CrossRef]
- Baldwin, R.; Calhoun, A.; Demaynadier, P. The significance of hydroperiod and stand maturity for pool-breeding amphibians in forested landscapes. Can. J. Zool. 2006, 84, 1604–1615. [Google Scholar] [CrossRef] [Green Version]
- Patrick, D.A.; Hunter, M.L., Jr.; Calhoun, A.J.K. Effects of experimental forestry treatments on a Maine amphibian community. For. Ecol. Manag. 2006, 234, 323–332. [Google Scholar] [CrossRef]
- Semlitsch, R.D.; Conner, C.A.; Hocking, D.J.; Rittenhouse, T.A.G.; Harper, E.B. Effects of timber harvesting on pond-breeding amphibian persistence: Testing the evacuation hypothesis. Ecol. Appl. 2008, 18, 283–289. [Google Scholar] [CrossRef]
- Morris, K.M.; Maret, T.J. Effects of Timber Management on Pond-Breeding Salamanders. J. Wildl. Manag. 2007, 71, 1034–1041. [Google Scholar] [CrossRef]
- Rothermel, B.; Luhring, T. Burrow Availability and Desiccation Risk of Mole Salamanders (Ambystoma talpoideum) in Harvested versus Unharvested Forest Stands. S. Am. J. Herpetol. 2005, 39, 619–626. [Google Scholar] [CrossRef]
- Clobert, J.; Le Galliard, J.-F.; Cote, J.; Meylan, S.; Massot, M. Informed dispersal, heterogeneity in animal dispersal syndromes and the dynamics of spatially structured populations. Ecol. Lett. 2009, 12, 197–209. [Google Scholar] [CrossRef]
- Semlitsch, R.D. Differentiating Migration and Dispersal Processes for Pond-Breeding Amphibians. J. Wildl. Manag. 2008, 72, 260–267. [Google Scholar] [CrossRef]
- Buhlmann, K.A.; Mitchell, J.C.; Smith, L.R. Descriptive ecology of the Shenandoah Valley sinkhole pond system in Virginia. Banisteria 1999, 13, 23–51. [Google Scholar]
- Scott, D. The Effect of Larval Density on Adult Demographic Traits in Ambystoma opacum. Ecology 1994, 75, 1383–1396. [Google Scholar] [CrossRef]
- Gamble, L.R.; McGarigal, K.; Compton, B.W. Fidelity and dispersal in the pond-breeding amphibian, Ambystoma opacum: Implications for spatio-temporal population dynamics and conservation. Biol. Conserv. 2007, 139, 247–257. [Google Scholar] [CrossRef]
- Gamble, L.R.; McGarigal, K.; Sigourney, D.B.; Timm, B.C. Survival and Breeding Frequency in Marbled Salamanders (Ambystoma opacum): Implications for Spatio-temporal Population Dynamics. Copeia 2009, 2009, 394–407. [Google Scholar] [CrossRef]
- Church, D.R. Role of current versus historical hydrology in amphibian species turnover within local pond communities. Copeia 2008, 2008, 115–125. [Google Scholar] [CrossRef]
- Bailey, L.L.; Kendall, W.L.; Church, D.R.; Wilbur, H.M. Estimating survival and breeding probabilities for pond-breeding amphibians: A modified robust design. Ecology 2004, 85, 2456–2466. [Google Scholar] [CrossRef]
- Bailey, L.L.; Kendall, W.L.; Church, D.R. Exploring extensions to multi-state models with multiple unobservable states. In Modeling Demographic Processes in Marked Populations; Environmental and Ecological Statistics Series; Thomson, D.L., Cooch, E.G., Conroy, M.J., Eds.; Springer Science & Business Media: New York, NY, USA, 2009; Volume 3, pp. 693–710. [Google Scholar]
- Bailey, L.L.; Converse, S.J.; Kendall, W.L. Bias, precision, and parameter redundancy in complex multi-state mod-els with unobservable states. Ecology 2010, 91, 1598–1604. [Google Scholar] [CrossRef]
- Kendall, W.L. Coping with unobservable and mis-classified states in capture-recapture studies. Anim. Biodivers. Conserv. 2004, 27, 97–101. [Google Scholar]
- White, G.C.; Burnham, K.P. Program MARK: Survival estimation from populations of marked animals. Bird Study 1999, 46, S120–S139. [Google Scholar] [CrossRef]
- Todd, B.; Blomquist, S.M.; Harper, E.B.; Osbourn, M.S. Effects of timber harvesting on terrestrial survival of pond-breeding amphibians. For. Ecol. Manag. 2014, 313, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Rittenhouse, T.A.; Harper, E.B.; Rehard, L.R.; Semlitsch, R.D. The role of microhabitats in the desiccation and survival of anurans in recently harvested oak–hickory forest. Copeia 2008, 2008, 807–814. [Google Scholar] [CrossRef]
- Connette, G.M.; Semlitsch, R.D. A multistate mark–recapture approach to estimating survival of PIT-tagged salamanders following timber harvest. J. Appl. Ecol. 2015, 52, 1316–1324. [Google Scholar] [CrossRef]
- Chazal, A.C.; Niewiarowski, P.H. Responses of mole salamanders to clearcutting: Using field experiments in forest management. Ecol. Appl. 1998, 8, 1133–1143. [Google Scholar] [CrossRef]
- Daszak, P.; Scott, D.; Kilpatrick, A.M.; Faggioni, C.; Gibbons, J.W.; Porter, D. Amphibian population declines at Savannah River Site are linked to climate, not chytridiomycosis. Ecology 2005, 86, 3232–3237. [Google Scholar] [CrossRef]
- Church, D.R.; Bailey, L.L.; Wilbur, H.M.; Kendall, W.L.; Hines, J.E. Iteroparity in the variable environment of the salamander Ambystoma tigrinum. Ecology 2007, 88, 891–903. [Google Scholar] [CrossRef]
- Vonesh, J.R.; De la Cruz, O. Complex life cycles and density dependence: Assessing the contribution of egg mortality to amphibian declines. Oecologia 2002, 133, 325–333. [Google Scholar] [CrossRef]
- Church, D.R. Population Ecology of Ambystoma tigrinum (Caudata, Ambystomatidae) and Occupancy Dynamics in an Appalachian Pond-Breeding Amphibian Assemblage. Ph.D. Thesis, University of Virginia, Charlottesville, VA, USA, 2004. [Google Scholar]
- Taylor, B.E.; Scott, D.E.; Gibbons, J.W. Catastrophic reproductive failure, terrestrial survival, and persistence of the marbled salamander. Conserv. Biol. 2006, 20, 792–801. [Google Scholar] [CrossRef]
- Petrovan, S.O.; Schmidt, B.R. Neglected juveniles; a call for integrating all amphibian life stages in assessments of mitigation success (and how to do it). Biol. Conserv. 2019, 236, 252–260. [Google Scholar] [CrossRef]
- Cayuela, H.; Arsovski, D.; Thirion, J.M.; Bonnaire, E.; Pichenot, J.; Boitaud, S.; Miaud, C.; Joly, P.; Besnard, A. Demographic responses to weather fluctuations are context dependent in a long-lived amphibian. Glob. Change. Biol. 2016, 22, 2676–2687. [Google Scholar] [CrossRef]
- Schmidt, B.R.; Feldmann, R.; Schaub, M. Demographic Processes Underlying Population Growth and Decline in Salamandra salamandra. Conserv. Biol. 2005, 19, 1149–1156. [Google Scholar] [CrossRef]
- Muths, E.; Scherer, R.D.; Lambert, B.A. Unbiased survival estimates and evidence for skipped breeding opportunities in females. Methods Ecol. Evol. 2010, 1, 123–130. [Google Scholar] [CrossRef]
- Cifuentes-Croquevielle, C.; Stanton, D.E.; Armesto, J.J. Soil invertebrate diversity loss and functional changes in temperate forest soils replaced by exotic pine plantations. Sci. Rep. 2020, 10, 7762. [Google Scholar] [CrossRef] [PubMed]
- Semlitsch, R.D. Terrestrial activity and summer home range of the mole salamander (Ambystoma talpoideum). Can. J. Zool. 1981, 59, 315–322. [Google Scholar] [CrossRef]
- Madison, D.M. The Emigration of Radio-Implanted Spotted Salamanders, Ambystoma maculatum. S. Am. J. Herpetol. 1997, 31, 542. [Google Scholar] [CrossRef]
- Rittenhouse, T.A.G.; Semlitsch, R.D. Distribution of amphibians in terrestrial habitat surrounding wetlands. Wetlands 2007, 27, 153–161. [Google Scholar] [CrossRef]
- Rothermel, B.B. Migratory success of juveniles: A potential constraint on connectivity for pond-breeding amphibians. Ecol. Appl. 2004, 14, 1535–1546. [Google Scholar] [CrossRef] [Green Version]
- Mazerolle, M.; Vos, C.C. Choosing the Safest Route: Frog Orientation in an Agricultural Landscape. S. Am. J. Herpetol. 2006, 40, 435–441. [Google Scholar] [CrossRef]
- Graeter, G.J.; Rothermel, B.B.; Gibbons, J.W. Habitat selection and movement of pond-breeding amphibians in experimentally fragmented pine forests. J. Wildl. Manag. 2008, 72, 473–482. [Google Scholar] [CrossRef]
- Todd, B.D.; Luhring, T.M.; Rothermel, B.; Gibbons, J.W. Effects of forest removal on amphibian migrations: Implications for habitat and landscape connectivity. J. Appl. Ecol. 2009, 46, 554–561. [Google Scholar] [CrossRef]
- Cayuela, H.; Grolet, O.; Joly, P. Context-dependent dispersal, public information, and heterospecific attraction in newts. Oecologia 2018, 188, 1069–1080. [Google Scholar] [CrossRef]
- Unglaub, B.; Cayuela, H.; Schmidt, B.R.; Preißler, K.; Glos, J.; Steinfartz, S. Context-dependent dispersal determines relatedness and genetic structure in a patchy amphibian population. Mol. Ecol. 2021, 30, 5009–5028. [Google Scholar] [CrossRef] [PubMed]
- Rittenhouse, T.A.; Semlitsch, R.D. Grasslands as movement barriers for a forest-associated salamander: Migration behavior of adult and juvenile salamanders at a distinct habitat edge. Biol. Conserv. 2006, 131, 14–22. [Google Scholar] [CrossRef]
- Brown, J.H.; Kodric-Brown, A. Turnover rates in insular biogeography: Effect of immigration on extinction. Ecology 1977, 58, 445–449. [Google Scholar] [CrossRef] [Green Version]
- Semlitsch, R.D.; Todd, B.D.; Blomquist, S.M.; Calhoun, A.J.; Gibbons, J.W.; Gibbs, J.P.; Graeter, G.J.; Harper, E.B.; Hocking, D.L.; Hunter, M.L., Jr.; et al. Effects of timber harvest on amphibian populations: Understanding mechanisms from forest experiments. BioScience 2009, 59, 853–862. [Google Scholar] [CrossRef] [Green Version]
- Bailey, L.L.; Muths, E. Integrating amphibian movement studies across scales better informs conservation deci-sions. Biol. Conserv. 2019, 236, 261–268. [Google Scholar] [CrossRef]












| Females | |||||
| Model | QDev. | K | QAIC | ΔQAIC | w |
| S(t) ψ (br_hab×pond, loc_hab×pond, land_hab×pond) p(t*) | 1863.9 | 121 | 13456.4 | 0.0 | 0.53 |
| S(t×pond) ψ (br_hab×pond, loc_hab×pond, land_hab) p(t*) | 1901.4 | 103 | 13457.0 | 0.7 | 0.38 |
| S(t×hab) ψ (br_hab×pond, loc_hab×pond, land_hab) p(t*) | 1918.8 | 96 | 13460.2 | 3.8 | 0.08 |
| Males | |||||
| Model | QDev. | K | QAIC | ΔQAIC | w |
| S(t×pond) ψ (br_hab×pond, loc_hab×pond, land_hab×pond) p(t*) | 3496.3 | 135 | 22670.5 | 0.0 | 1.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Church, D.R.; Bailey, L.L.; Wilbur, H.M.; Green, J.H.; Hiby, L. Salamander Demography at Isolated Wetlands within Mature and Regenerating Forests. Diversity 2022, 14, 309. https://doi.org/10.3390/d14050309
Church DR, Bailey LL, Wilbur HM, Green JH, Hiby L. Salamander Demography at Isolated Wetlands within Mature and Regenerating Forests. Diversity. 2022; 14(5):309. https://doi.org/10.3390/d14050309
Chicago/Turabian StyleChurch, Don R., Larissa L. Bailey, Henry M. Wilbur, James H. Green, and Lex Hiby. 2022. "Salamander Demography at Isolated Wetlands within Mature and Regenerating Forests" Diversity 14, no. 5: 309. https://doi.org/10.3390/d14050309
APA StyleChurch, D. R., Bailey, L. L., Wilbur, H. M., Green, J. H., & Hiby, L. (2022). Salamander Demography at Isolated Wetlands within Mature and Regenerating Forests. Diversity, 14(5), 309. https://doi.org/10.3390/d14050309