
Phytoliths from Woody Plants: A Review


Abstract
:1. Introduction
2. Morphology of Woody Plant Phytoliths
- (1)
- (2)
- (3)
- (4)
2.1. Gymnosperms
2.1.1. Pinaceae
2.1.2. Araucariaceae
2.1.3. Cupressaceae
2.2. Monocotyledonous Woody Plants
Arecaceae
2.3. Broad-Leaf Trees
2.3.1. Leaves of Broad-Leaf Trees
2.3.2. Bark, Wood, and Other Parts
3. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sangster, A.G.; Hodson, M.J.; Tubb, H.J. Chapter 5 Silicon deposition in higher plants. In Silicon in Agriculture; Datnoff, L.E., Snyder, G.H., Korndörfer, G.H., Eds.; Studies in Plant Science; Elsevier: Amsterdam, The Netherlands, 2001; Volume 8, pp. 85–113. [Google Scholar]
- Trembath-Reichert, E.; Wilson, J.P.; McGlynn, S.E.; Fischer, W.W. Four hundred million years of silica biomineralization in land plants. Proc. Natl. Acad. Sci. USA 2015, 112, 5449–5454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katz, O. Silicon content is a plant functional trait: Implications in a changing world. Flora 2019, 254, 88–94. [Google Scholar] [CrossRef]
- Stromberg, C.A.E.; Dunn, R.E.; Madden, R.H.; Kohn, M.J.; Carlini, A.A. Decoupling the spread of grasslands from the evolution of grazer-type herbivores in South America. Nat. Commun. 2013, 4, 1478. [Google Scholar] [CrossRef] [PubMed]
- Hart, T.C. Issues and directions in phytolith analysis. J. Archaeol. Sci. 2016, 68, 24–31. [Google Scholar] [CrossRef]
- Rashid, I.; Mir, S.H.; Zurro, D.; Dar, R.A.; Reshi, Z.A. Phytoliths as proxies of the past. Earth-Sci. Rev. 2019, 194, 234–250. [Google Scholar] [CrossRef]
- Hodson, M.J.; Song, Z.; Ball, T.B.; Elbaum, R.; Struyf, E. Editorial: Frontiers in Phytolith Research. Front. Plant Sci. 2020, 11, 454. [Google Scholar] [CrossRef] [Green Version]
- Alexandre, A.; Crespin, J.; Sylvestre, F.; Sonzogni, C.; Hilbert, D.W. The oxygen isotopic composition of phytolith assemblages from tropical rainforest soil tops (Queensland, Australia): Validation of a new paleoenvironmental tool. Clim. Past 2012, 8, 307–324. [Google Scholar] [CrossRef] [Green Version]
- Hodson, M.J. The development of phytoliths in plants and its influence on their chemistry and isotopic composition. Implications for palaeoecology and archaeology. J. Archaeol. Sci. 2016, 68, 62–69. [Google Scholar] [CrossRef]
- Coe, H.H.G.; Alexandre, A.; Carvalho, C.N.; Santos, G.M.; da Silva, A.S.; Sousa, L.O.F.; Lepsch, I.F. Changes in Holocene tree cover density in Cabo Frio (Rio de Janeiro, Brazil): Evidence from soil phytolith assemblages. Quat. Int. 2013, 287, 63–72. [Google Scholar] [CrossRef]
- Albert, R.M.; Bamford, M.K.; Esteban, I. Reconstruction of ancient palm vegetation landscapes using a phytolith approach. Quat. Int. 2015, 369, 51–66. [Google Scholar] [CrossRef]
- Lu, H.-Y.; Wu, N.-Q.; Yang, X.-D.; Jiang, H.; Liu, K.-b.; Liu, T.-S. Phytoliths as quantitative indicators for the reconstruction of past environmental conditions in China I: Phytolith-based transfer functions. Quat. Sci. Rev. 2006, 25, 945–959. [Google Scholar] [CrossRef]
- Piperno, D.R. Phytolith Analysis: An Archaeological and Geological Perspective; Academic Press Inc.: San Diego, CA, USA, 1988; p. 280. [Google Scholar]
- Piperno, D.R. Phytoliths: A Comprehensive Guide for Archaeologists and Paleoecologists; AltaMira Press: Lanham, MD, USA, 2006; p. 239. [Google Scholar]
- Rovner, I. Potential of Opal Phytoliths for use in Paleoecological Reconstruction. Quat. Res. 1971, 1, 343–359. [Google Scholar] [CrossRef]
- Wilding, L.P.; Drees, L.R. Biogenic Opal in Ohio Soils. Soil Sci. Soc. Am. J. 1971, 35, 1004–1010. [Google Scholar] [CrossRef]
- Geis, J.W. Biogenic Silica in Selected Species of Deciduous Angiosperms. Soil Sci. 1973, 116, 113–130. [Google Scholar] [CrossRef]
- Kondo, R. On the Opal Phytoliths of Tree Origins. Pedologist 1977, 20, 176–185. [Google Scholar]
- Klein, R.L.; Geis, J.W. Biogenic Silica in the Pinaceae. Soil Sci. 1978, 126, 145–156. [Google Scholar] [CrossRef]
- Kondo, R.; Sumida, T. The Study on Opal Phytoliths of Tree Leaves (Part 1): Opal Phytoliths in Gymnosperm and Monocotyledonous Angiosperm Tree Leaves. Soil Sci. Plant Nutr. 1978, 49, 138–144. [Google Scholar] [CrossRef]
- Kondo, R.; Peason, T. Opal phytoliths in tree leaves (part 2): Opal phytoliths in dicotyledonous angiosperm tree leaves. Res. Bull. Obihiro Univ. 1981, 22, 217–229. [Google Scholar]
- Kealhofer, L.; Piperno, D.R. Opal Phytoliths in Southeast Asian Flora. Smithson. Contrib. Bot. 1998, 88, 1–39. [Google Scholar] [CrossRef]
- Lentfer, C.J. Plants, People, and Landscapes in Prehistoric Papua New Guinea: A Compendium of Phytolith (and Starch) Analyses; Southern Cross University: Lismore, Australia, 2003. [Google Scholar]
- Wallis, L. An overview of leaf phytolith production patterns in selected northwest Australian flora. Rev. Palaeobot. Palynol. 2003, 125, 201–248. [Google Scholar] [CrossRef]
- Carnelli, A.L.; Theurillat, J.-P.; Madella, M. Phytolith types and type-frequencies in subalpine–alpine plant species of the European Alps. Rev. Palaeobot. Palynol. 2004, 129, 39–65. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, H. The Study of Phytolith and Its Application; China Ocean Press: Beijing, China, 1993; p. 234. [Google Scholar]
- Kondo, R. An Introduction to Phytolith by Scanning Electron Microscopy; Hokkaido University Press: Sapporo, Japan, 2010; p. 387. [Google Scholar]
- Madella, M.; Alexandre, A.; Ball, T.; Icpn Working, G. International code for phytolith nomenclature 1.0. Ann. Bot. 2005, 96, 253–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ICPN; Neumann, K.; Strömberg, C.A.E.; Ball, T.; Albert, R.M.; Vrydaghs, L.; Cummings, L.S. International Code for Phytolith Nomenclature (ICPN) 2.0. Ann. Bot. 2019, 124, 189–199. [Google Scholar] [CrossRef]
- Blinnikov, M.S. Phytoliths in plants and soils of the interior Pacific Northwest, USA. Rev. Palaeobot. Palynol. 2005, 135, 71–98. [Google Scholar] [CrossRef]
- Iriarte, J.; Paz, E.A. Phytolith analysis of selected native plants and modern soils from southeastern Uruguay and its implications for paleoenvironmental and archeological reconstruction. Quat. Int. 2009, 193, 99–123. [Google Scholar] [CrossRef]
- Mercader, J.; Bennett, T.; Esselmont, C.; Simpson, S.; Walde, D. Phytoliths in woody plants from the Miombo woodlands of Mozambique. Ann. Bot. 2009, 104, 91–113. [Google Scholar] [CrossRef] [Green Version]
- Morris, L.R.; Baker, F.A.; Morris, C.; Ryel, R.J. Phytolith types and type-frequencies in native and introduced species of the sagebrush steppe and pinyon–juniper woodlands of the Great Basin, USA. Rev. Palaeobot. Palynol. 2009, 157, 339–357. [Google Scholar] [CrossRef]
- Das, S.; Ghosh, R.; Bera, S. Application of non-grass phytoliths in reconstructing deltaic environments: A study from the Indian Sunderbans. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 376, 48–65. [Google Scholar] [CrossRef]
- McCune, J.L.; Pellatt, M.G. Phytoliths of Southeastern Vancouver Island, Canada, and their potential use to reconstruct shifting boundaries between Douglas-fir forest and oak savannah. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 383–384, 59–71. [Google Scholar] [CrossRef]
- Watling, J.; Iriarte, J. Phytoliths from the coastal savannas of French Guiana. Quat. Int. 2013, 287, 162–180. [Google Scholar] [CrossRef]
- Das, S.; Ghosh, R.; Paruya, D.K.; Yao, Y.-F.; Li, C.-S.; Bera, S. Phytolith spectra in respiratory aerial roots of some mangrove plants of the Indian Sunderbans and its efficacy in ancient deltaic environment reconstruction. Quat. Int. 2014, 325, 179–196. [Google Scholar] [CrossRef]
- Collura, L.V.; Neumann, K. Wood and bark phytoliths of West African woody plants. Quat. Int. 2017, 434, 142–159. [Google Scholar] [CrossRef]
- Esteban, I.; Vlok, J.; Kotina, E.L.; Bamford, M.K.; Cowling, R.M.; Cabanes, D.; Albert, R.M. Phytoliths in plants from the south coast of the Greater Cape Floristic Region (South Africa). Rev. Palaeobot. Palynol. 2017, 245, 69–84. [Google Scholar] [CrossRef]
- Gao, G.; Jie, D.; Liu, L.; Liu, H.; Gao, Z.; Li, D.; Li, N. Phytolith characteristics and preservation in trees from coniferous and broad-leaved mixed forest in an eastern mountainous area of Northeast China. Rev. Palaeobot. Palynol. 2018, 255, 43–56. [Google Scholar] [CrossRef]
- Novello, A.; Bamford, M.K.; van Wijk, Y.; Wurz, S. Phytoliths in modern plants and soils from Klasies River, Cape Region (South Africa). Quat. Int. 2018, 464, 440–459. [Google Scholar] [CrossRef]
- Ricardo, S.D.F.; Coe, H.H.G.; Dias, R.R.; de Sousa, L.d.O.F.; Gomes, E. Reference collection of plant phytoliths from the Caatinga biome, Northeast Brazil. Flora 2018, 249, 1–8. [Google Scholar] [CrossRef]
- Levin, M.J. Prehistoric agricultural transformations in tropical Remote Oceania: Towards comprehensive phytolith analysis. Quat. Int. 2019, 529, 75–90. [Google Scholar] [CrossRef]
- Ge, Y. Studies on the Morphology and Application of Phytolith in Common Modern Plants in China. Ph.D. Thesis, University of Chinese Academy of Sciences, Beijing, China, 2017. [Google Scholar]
- Piperno, D.R.; McMichael, C. Phytoliths in modern plants from amazonia and the neotropics at large: Implications for vegetation history reconstruction. Quat. Int. 2020, 565, 54–74. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, H.; Jie, D.; Gao, G.; Meng, M.; Zhang, G. Phytolith morphotypes of woody plants and their preservation in soil in the warm temperate humid zones of China. Quat. Int. 2021, 599–600, 158–169. [Google Scholar] [CrossRef]
- Xu, D.; Li, Q.; Lü, H. Morphological Analysis of Phytoliths on Palmae and Its Environmental Significance. Quat. Sci. 2005, 25, 785–792. [Google Scholar]
- Parr, J.; Watson, L. Morphological characteristics observed in the leaf phytoliths of selected Gymnosperms of eastern Australia. In Plants, People and Places: Recent Studies in Phytolithic Analysis; Oxbow Books: Oxword, UK, 2007; pp. 73–78. [Google Scholar]
- Koeppen, R.C. Silica Booies in Wood of Arborescent Leguminosae. IAWA J. 1980, 1, 180–184. [Google Scholar] [CrossRef]
- Kondo, R.; Oosawa, S.; Tsutsuki, K.; Tani, M.; Shibano, S. Characteristics of the opal phytoliths derived from Piaceae leaves. Pedologist 2003, 47, 90–103. [Google Scholar] [CrossRef]
- An, X.-H. Morphological characteristics of phytoliths from representative conifers in China. Palaeoworld 2016, 25, 116–127. [Google Scholar] [CrossRef]
- Kondo, R.; Tsutsuki, K.; Tani, M.; Maruyama, R. Differentiation of Genera Pinus, Picea, and Abies by the Transfusion Tracheid Phytoliths of Pinaceae Leaves. Pedologist 2002, 46, 32–35. [Google Scholar] [CrossRef]
- Fenwick, R.S.H.; Lentfer, C.J.; Weisler, M.I. Palm reading: A pilot study to discriminate phytoliths of four Arecaceae (Palmae) taxa. J. Archaeol. Sci. 2011, 38, 2190–2199. [Google Scholar] [CrossRef]
- Benvenuto, M.L.; Fernandez Honaine, M.; Osterrieth, M.L.; Morel, E. Differentiation of globular phytoliths in Arecaceae and other monocotyledons: Morphological description for paleobotanical application. Turk. J. Bot. 2015, 39, 341–353. [Google Scholar] [CrossRef]
- Hu, M.; Zuo, L.; Jie, D.; Gao, G.; Meng, X.; Zhang, T.; Chen, N.; Bahayila, H.; Hu, H. The identification of phytolith characteristics of the herbaceous and woody plants in Northeast China. Acta Micropalaeontol. Sin. 2018, 35, 122–139. [Google Scholar] [CrossRef]
- Watling, J.; Castro, M.T.; Simon, M.F.; Rodrigues, F.O.; Brilhante de Medeiros, M.; De Oliveira, P.E.; Neves, E.G. Phytoliths from native plants and surface soils from the Upper Madeira river, SW Amazonia, and their potential for paleoecological reconstruction. Quat. Int. 2020, 550, 85–110. [Google Scholar] [CrossRef]
- Lins, U.; Barros, C.F.; da Cunha, M.; Miguens, F.C. Structure, morphology, and composition of silicon biocomposites in the palm tree Syagrus coronata (Mart.) Becc. Protoplasma 2002, 220, 89–96. [Google Scholar] [CrossRef]
- Tsutsui, O.; Sakamoto, R.; Obayashi, M.; Yamakawa, S.; Handa, T.; Nishio-Hamane, D.; Matsuda, I. Light and SEM observation of opal phytoliths in the mulberry leaf. Flora Morphol. Distrib. Funct. Ecol. Plants 2016, 218, 44–50. [Google Scholar] [CrossRef] [Green Version]
- Lisztes-Szabó, Z.; Braun, M.; Csík, A.; Pető, Á. Phytoliths of six woody species important in the Carpathians: Characteristic phytoliths in Norway spruce needles. Veg. Hist. Archaeobotany 2019, 28, 649–662. [Google Scholar] [CrossRef] [Green Version]
- Lisztes-Szabó, Z.; Filep, A.F.; Csík, A.; Pető, Á.; Kertész, T.G.; Braun, M. pH-dependent silicon release from phytoliths of Norway spruce (Picea abies). J. Paleolimnol. 2019, 63, 65–81. [Google Scholar] [CrossRef] [Green Version]
- Yapuchura, E.R.; Tartaglia, R.S.; Cunha, A.G.; Freitas, J.C.C.; Emmerich, F.G. Observation of the transformation of silica phytoliths into SiC and SiO2 particles in biomass-derived carbons by using SEM/EDS, Raman spectroscopy, and XRD. J. Mater. Sci. 2018, 54, 3761–3777. [Google Scholar] [CrossRef]
- Bokor, B.; Soukup, M.; Vaculík, M.; Vd’ačný, P.; Weidinger, M.; Lichtscheidl, I.; Vavrova, S.; Soltys, K.; Sonah, H.; Deshmukh, R.; et al. Silicon Uptake and Localisation in Date Palm (Phoenix dactylifera)—A Unique Association With Sclerenchyma. Front. Plant Sci. 2019, 10, 988. [Google Scholar] [CrossRef] [Green Version]
- George, N.; Antony, A.; Ramachandran, T.; Hamed, F.; Kamal-Eldin, A. Microscopic Investigationsof Silicification and Lignification Suggest Their Coexistence in Tracheary Phytoliths in Date Fruits (Phoenix dactylifera L.). Front. Plant Sci. 2020, 11, 977. [Google Scholar] [CrossRef]
- Christenhusz, M.J.M.; Reveal, J.L.; Farjon, A.; Gardner, M.F.; Mill, R.R.; Chase, M.W. A new classification and linear sequence of extant gymnosperms. Phytotaxa 2011, 19, 55–70. [Google Scholar] [CrossRef]
- Stromberg, C.A.E. The Origin and Spread of Grass-Dominated Ecosystems during the Tertiary of North America and How It Relates to the Evolution of Hypsodonty in Equids. Ph.D. Thesis, University of California, Berkeley, CA, USA, 2003. [Google Scholar]
- Biswas, O.; Mukherjee, B.; Bera, S. Phytolith production in gymnosperms: Its pattern and application. J. Bot. Soc. Bengal 2016, 70, 75–83. [Google Scholar]
- Sangster, A.G.; Hodson, M.J. Silicification of conifers and its significance to the environment. In Plants, People and Places Recent Studies in Phytolith Analysis; Madella, M., Ed.; Oxbow Books: Oxford, UK, 2007. [Google Scholar]
- Sangster, A.G.; Hodson, M.J.; Ling, L.E.C. Biomineralisation/environment interactions in conifers: Illustrated by hemlock, Tsuga canadensis (L.) Carr. Quat. Int. 2009, 193, 3–10. [Google Scholar] [CrossRef]
- Bozarth, S.R. Biosilicate assemblages of boreal forests and aspen parklands. In Current Research in Phytoliths Analysis: Applications in Archeology and Paleoecology; Pearsall, D., Piperno, D., Eds.; University of Pennsylvania: Philadelphia, PA, USA, 1993; Volumes 95–105. [Google Scholar]
- Hodson, M.J.; Sangster, A.G. Aluminium/silicon interactions in conifers. J. Inorg. Biochem. 1999, 76, 89–98. [Google Scholar] [CrossRef]
- Chen, I.; Li, K.-t.; Tsang, C.-h. Silicified bulliform cells of Poaceae: Morphological characteristics that distinguish subfamilies. Bot. Stud. 2020, 61, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.-S.; Yao, B.-J. Transfusion tissue in gymnosperm leaves. Bot. J. Linn. Soc. 1981, 83, 263–272. [Google Scholar] [CrossRef]
- Boddi, S.; Bonzi, L.M.; Calamassi, R. Structure and ultrastructure of Pinus halepensis primary needles. Flora Morphol. Distrib. Funct. Ecol. Plants 2002, 197, 10–23. [Google Scholar] [CrossRef]
- Kerns, B.K. Diagnostic Phytoliths for a Ponderosa Pine-Bunchgrass Community near Flagstaff, Arizona. Southwest. Nat. 2001, 46, 282–294. [Google Scholar] [CrossRef]
- Sangster, A.G.; Ling, L.; Gérard, F.; Hodson, M.J. X-ray Microanalysis of Needles from Douglas Fir Growing in Environments of Contrasting Acidity. In Acid Rain—Deposition to Recovery; Brimblecombe, P., Hara, H., Houle, D., Novak, M., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 143–149. [Google Scholar]
- McNamee, C. Soil Phytolith Assemblages of the American Southwest: The Use of Historical Ecology in Taphonomic Studies. Ph.D. Thesis, University of Calgary, Calgary, AB, Canada, 2013. [Google Scholar]
- Mazumdar, J.; Mukhopadhyay, R. Phytoliths of gymnosperms. Bionature 2012, 32, 21–26. [Google Scholar]
- Hodson, M.J.; Wiliams, S.E.; Sangster, A.G. Silica deposition in the needles of the gymnosperms. 1. Chemical analysis and light microscopy. In The State of the Art of Phytoliths in Soils and Plants; Pinilla, A., Juan-Tresserras, J., Machado, M.J., Eds.; Monograph 4; Centro de Ciencias Medioambientales: Madrid, Spain, 1997. [Google Scholar]
- Burleigh, J.G.; Barbazuk, W.B.; Davis, J.M.; Morse, A.M.; Soltis, P.S. Exploring Diversification and Genome Size Evolution in Extant Gymnosperms through Phylogenetic Synthesis. J. Bot. 2012, 2012, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Canny, M.J. Transfusion tissue of pine needles as a site of retrieval of solutes from the transpiration stream. New Phytol. 1993, 123, 227–232. [Google Scholar] [CrossRef]
- Liesche, J.; Martens, H.J.; Schulz, A. Symplasmic transport and phloem loading in gymnosperm leaves. Protoplasma 2011, 248, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Laur, J.; Hacke, U.G. Exploring Picea glauca aquaporins in the context of needle water uptake and xylem refilling. New Phytol. 2014, 203, 388–400. [Google Scholar] [CrossRef]
- The Angiosperm Phylogeny, G.; Chase, M.W.; Christenhusz, M.J.M.; Fay, M.F.; Byng, J.W.; Judd, W.S.; Soltis, D.E.; Mabberley, D.J.; Sennikov, A.N.; Soltis, P.S.; et al. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Prychid, C.J.; Rudall, P.J.; Gregory, M. Systematics and Biology of Silica Bodies in Monocotyledons. Bot. Rev. 2003, 69, 377–440. [Google Scholar] [CrossRef]
- Dransfield, J.; Uhl, N.W.; Asmussen, C.B.; Baker, W.J.; Harley, M.M.; Lewis, C.E. A New Phylogenetic Classification of the Palm Family, Arecaceae. Kew Bull. 2005, 60, 559–569. [Google Scholar]
- Morcote-Ríos, G.; Bernal, R.; Raz, L. Phytoliths as a tool for archaeobotanical, palaeobotanical and palaeoecological studies in Amazonian palms. Bot. J. Linn. Soc. 2016, 182, 348–360. [Google Scholar] [CrossRef] [Green Version]
- Huisman, S.N.; Raczka, M.F.; McMichael, C.N.H. Palm Phytoliths of Mid-Elevation Andean Forests. Front. Ecol. Evol. 2018, 6, 193. [Google Scholar] [CrossRef] [Green Version]
- Piperno, D.R. The occurrence of phytoliths in the reproductive structures of selected tropical angiosperms and their significance in tropical paleoecology, paleoethnobotany and systematics. Rev. Palaeobot. Palynol. 1989, 61, 147–173. [Google Scholar] [CrossRef]
- Patterer, N.I. Análisis fitolíticos de las principales especies de palmeras (Arecaceae) presentes en regiones subtropicales de América del Sur. Boletín Soc. Argent. Botánica 2014, 49, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Tomlinson, P.B. Anatomy of the Monocotyledons. II. Palmae; Oxford University Press: Oxford, UK, 1961. [Google Scholar]
- Thomas, R.; De Franceschi, D. Palm stem anatomy and computer-aided identification: The Coryphoideae (Arecaceae). Am. J. Bot. 2013, 100, 289–313. [Google Scholar] [CrossRef]
- Schmitt, U.; Weiner, G.; Liese, W. The Fine Structure of the Stegmata in Calamus Axillaris during Maturation. IAWA J. 1995, 16, 61–68. [Google Scholar] [CrossRef]
- Delhon, C.; Orliac, C. The vanished palm trees of Easter Island: New radiocarbon and phytolith data. In Proceedings of the VII International Conference on Easter Island and the Pacific, Visby, Sweden, 20–25 August 2007. [Google Scholar]
- Bowdery, D. An enigma revisited: Identification of palm phytoliths extracted from the 1983 Rapa Nui, Rano Kao2 core. Veg. Hist. Archaeobotany 2015, 24, 455–466. [Google Scholar] [CrossRef]
- Chen, S.T.; Smith, S.Y. Phytolith variability in Zingiberales: A tool for the reconstruction of past tropical vegetation. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 370, 1–12. [Google Scholar] [CrossRef]
- Stromberg, C.A.E. Using phytolith assemblages to reconstruct the origin and spread of grass-dominated habitats in the great plains of North America during the late Eocene to early Miocene. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2004, 207, 239–275. [Google Scholar] [CrossRef]
- Sharma, R.; Kumar, V.; Kumar, R. Distribution of phytoliths in plants: A review. Geol. Ecol. Landsc. 2018, 3, 123–148. [Google Scholar] [CrossRef] [Green Version]
- Bozarth, S.R. Classification of Opal Phytoliths Formed in Selected Dicotyledons Native to the Great Plains. In Phytolith Systematics: Emerging Issues; Rapp, G., Mulholland, S.C., Eds.; Springer: Boston, MA, USA, 1992; pp. 193–214. [Google Scholar]
- Tsartsidou, G.; Lev-Yadun, S.; Albert, R.-M.; Miller-Rosen, A.; Efstratiou, N.; Weiner, S. The phytolith archaeological record: Strengths and weaknesses evaluated based on a quantitative modern reference collection from Greece. J. Archaeol. Sci. 2007, 34, 1262–1275. [Google Scholar] [CrossRef]
- Honaine, M.F.; Zucol, A.F.; Osterrieth, M.L. Phytolith analysis of Cyperaceae from the Pampean region, Argentina. Aust. J. Bot. 2009, 57, 512–523. [Google Scholar] [CrossRef]
- Mukherjee, B.; Biswas, O.; Ghosh, R.; Paruya, D.K.; Bera, S. Phytolith spectra of some eastern Himalayan pteridophytes: Assessing potential for distinguishing different vegetation–climate zones along the elevation gradient. Flora 2019, 254, 95–112. [Google Scholar] [CrossRef]
- Ge, Y.; Lu, H.; Wang, C.; Gao, X. Phytoliths in selected broad-leaved trees in China. Sci. Rep. 2020, 10, 15577. [Google Scholar] [CrossRef] [PubMed]
- Lepsch, I.; Oliveira, L.; Pinilla-Navarro, A.; Coe, H. Phytoliths from the Rough Leaves of Two Dicotyledonous Species from the Brazilian Cerrado Biome and Their Occurrence in Soils. In Synthesis of Some Phytolith Studies in South America (Brazil and Argentina); Nova Publishers: New York, NY, USA, 2014; pp. 75–89. [Google Scholar]
- Feng, Y.; Jie, D.; Guo, M.; Dong, S.; Chen, X.; Liu, H.; Liu, L.; Li, N. Phytolith loss and enrichment in soil phytolith assemblages revealed by comparisons of phytoliths in vegetation and surface soils of altitudinal belts in the Changbai Mountains, Northeast China. Flora 2017, 236–237, 84–93. [Google Scholar] [CrossRef]
- Zucol, A.F.; Brea, M.; Bellosi, E.S. Phytolith studies in Gran Barranca (central Patagonia, Argentina): The middle–late Eocene. In The Paleontology of Gran Barranca: Evolution and Environmental Change through the Middle Cenozoic of Patagonia; Cambridge University Press: Cambridge, UK, 2010; pp. 317–340. [Google Scholar]
- Beck, C.B. An Introduction to Plant Structure and Development: Plant Anatomy for the Twenty-First Century; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Schweingruber, F.H.; Börner, A. The Plant Stem: A Microscopic Aspect; Springer: Cham, Switzerland, 2018. [Google Scholar]
- Crang, R.; Lyons-Sobaski, S.; Wise, R. Plant Anatomy: A Concept-Based Approach to the Structure of Seed Plants; Springer: New York, NY, USA, 2018. [Google Scholar]
- Testé, M.; Garnier, A.; Limondin-Lozouet, N.; Oxlaj, E.; Castanet, C.; Purdue, L.; Lemonnier, E.; Dussol, L.; Nondédéo, P. The phytoliths of Naachtun (Petén, Guatemala): Development of a modern reference for the characterization of plant communities in the Maya Tropical Lowlands. Rev. Palaeobot. Palynol. 2020, 272, 104130. [Google Scholar] [CrossRef]
- Postek, M.T. The Occurrence of Silica in the Leaves of Magnolia grandiflora L. Bot. Gaz. 1981, 142, 124–134. [Google Scholar] [CrossRef]
- Welle, B.J.H.t. Silica grains in woody plants of the neotropics, especially Surinam. Leiden Bot. Ser. 1976, 3, 107–142. [Google Scholar]
- Torelli, N.; Čufar, K. Mexican tropical hardwoods. Comparative study of ash and silica content. Holz Roh-Werkst. 1995, 53, 61–62. [Google Scholar] [CrossRef]
- Pearsall, D.M.B.A.; Duncan, N.A.; Chandler-Ezell, C.; Grimm, B.; Hart, T.; Logan, A.; O’Brien, M.; Stewart, S.; Veintimilla, C.; Zhao, Z. Phytoliths in the Flora of Ecuador: The University of Missouri Online Phytolith Database. Available online: http://phytolith.missouri.edu (accessed on 22 January 2022).
- Sangster, A.G.; Hodson, M.J. Silica Deposition in Subterranean Organs. In Phytolith Systematics: Emerging Issues; Rapp, G., Mulholland, S.C., Eds.; Springer: Boston, MA, USA, 1992; pp. 239–251. [Google Scholar]


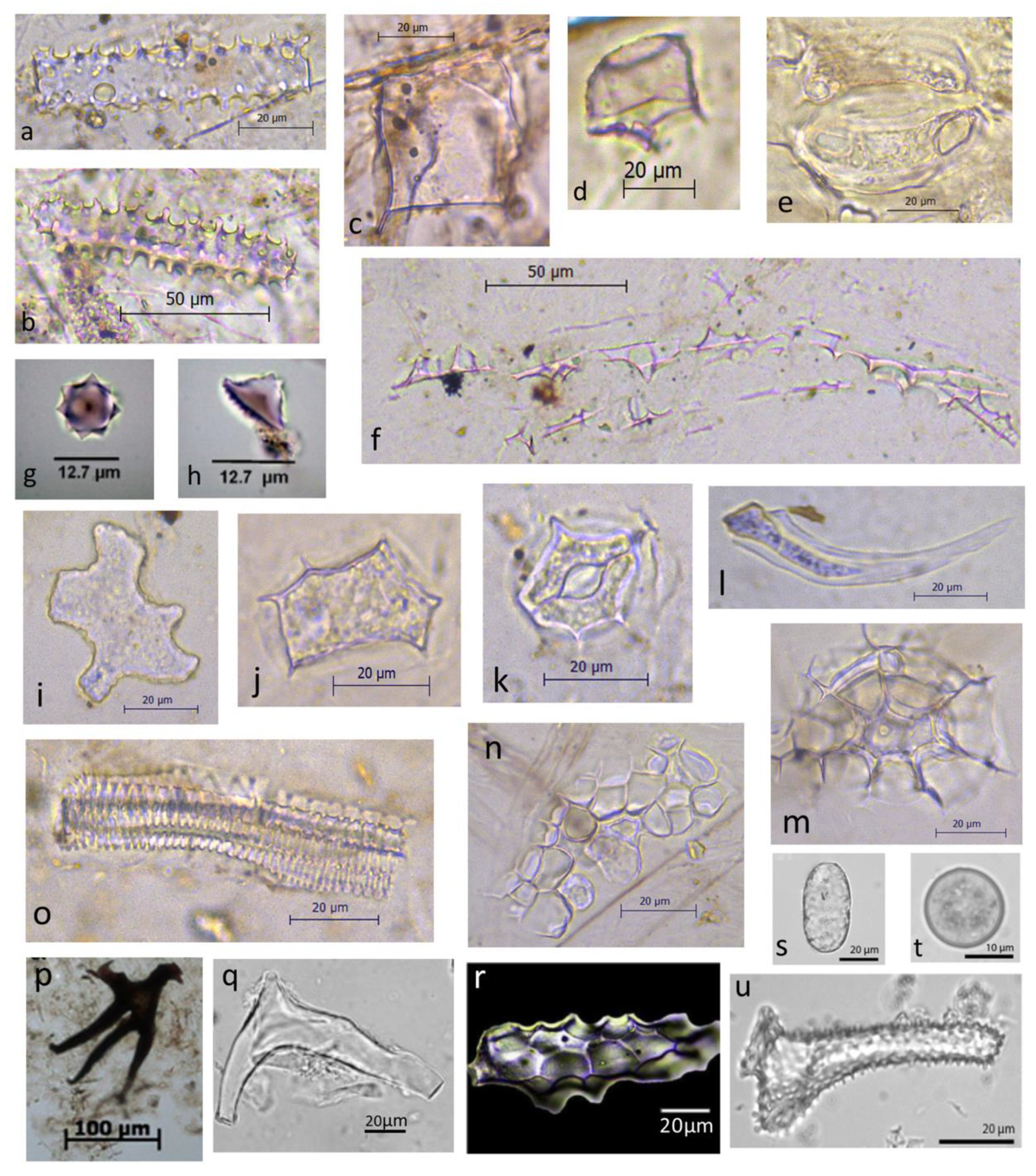{kind=link}
| Morphotype | Description | Family (Genus) | Reference |
|---|---|---|---|
| Specific epidermal phytoliths | Large (>50 μm) Attached to the periphery of hair bases | Dichapetalaceae | (Piperno and McMichael 2020) |
| Tabular to blocky and half-decorated with a papillate texture | Chrysobalanaceae | (Watling et al., 2020) | |
| Shape square to rectangular epidermal cell) with round/spherical bulb or projection in centre of one flattened side. All surfaces smooth, but surface of rectangle and edges are upturned and undulating. | Fabaceae | http://phytolith.missouri.edu (accessed on 22 January 2022) | |
| Y-shaped or brachiate Sclereids | Usually irregular elongate, branched, and may have psilate or fluted surfaces. Sizes are 50 to >90 μm in length. | Magnoliaceae Hamamelidaceae | (Kondo and Peason 1981) (Kondo 2010) |
| Fagaceae | (Kondo and Peason 1981) (Piperno and McMichael 2020) | ||
| Annonaceae | (Kealhofer and Piperno 1998) (Watling and Iriarte 2013) | ||
| Thymelaeaceae | (Iriarte and Paz 2009) | ||
| Sterculiaceae Dipterocarpaceae Sapindaceae Lauraceae | (Kondo 2010) | ||
| Anacardiaceae Erythroxylaceae Myrtaceae | (Watling and Iriarte 2013) | ||
| Picrodendraceae | (Piperno and McMichael 2020) | ||
| Papilionaceae | (Lentfer 2003) | ||
| Bombacaceae Ericaceae Ebenaceae Dichapetalaceae | http://phytolith.missouri.edu (accessed on 22 January 2022) | ||
| Acute Bulbosus (Specifichairs) | Elongated cones with tuberculate or echinate ornamentation | Cannabaceae Loganiaceae Orobanchaceae | (Wallis 2003) |
| Moraceae | (Wallis 2003) (Levin 2019) (Ge et al., 2020) | ||
| Boraginaceae Urticaceae (Pipturus) | (Levin 2019) | ||
| Squat cones with tuberculate or echinate ornamentation | Ulmaceae | (Wallis 2003) (Morris et al., 2009) | |
| Urticaceae (Pipturus) Moraceae | (Levin 2019) | ||
| Cannabaceae | (Watling and Iriarte 2013) | ||
| Segmented acicular hair | Asteraceae | (Bozarth 1992) (Lentfer 2003) (Iriarte and Paz 2009) | |
| Euphorbiaceae Piperaceae | http://phytolith.missouri.edu (accessed on 22 January 2022) | ||
| Facetate phytoliths | Spheroidal, elongate, or irregular body Large (usually >50 μm) The facetate surface may be formed by the squeeze of surrounding cells | Annonaceae | (Kealhofer and Piperno 1998) (Watling and Iriarte 2013) (Piperno and McMichael 2020) |
| Flacourtiaceae | (Kealhofer and Piperno 1998) | ||
| Fagaceae | (Kondo and Peason 1981) (Kealhofer and Piperno 1998) (Kondo 2010) (Watling and Iriarte 2013) | ||
| Lauraceae | (Kondo and Peason 1981) (Kondo 2010) (Ge et al., 2020) | ||
| Cunoniaceae Magnoliaceae Sapindaceae | (Kondo and Peason 1981) (Kondo 2010) | ||
| Chrysobalanaceae Dilleniaceae Elaeocarpaceae | (Watling and Iriarte 2013) | ||
| Pittosporaceae | (Ge et al., 2020) | ||
| Ebenaceae | http://phytolith.missouri.edu (accessed on 22 January 2022) | ||
| Piperaceae Sapotaceae | (Lentfer 2003) | ||
| Annulate/helical bodies | Usually, irregular elongate solid body Large (>50 μm) Originate from silicified tracheid tissues | Adoxaceae (Viburnum) | (Kondo and Peason 1981) |
| Lauraceae | (Kondo and Peason 1981) (Kondo 2010) (Ge et al., 2020) | ||
| Cunoniaceae Fagaceae Sapindaceae | (Kondo 2010) | ||
| Malvaceae | (McNamee 2013) | ||
| Magnoliaceae Pittosporaceae | (Ge et al., 2020) | ||
| Flacourtiaceae | http://phytolith.missouri.edu (accessed on 22 January 2022) | ||
| Specific Spheroids | Spheroid folded | Anacardiaceae | (Kealhofer and Piperno 1998) |
| Spheroid bisected | Solanaceae (Solanum) | (Mercader et al., 2009) (Ge et al., 2020) | |
| Single rounded projection or “hat” on one side Fracture marks on the other side | Lauraceae | http://phytolith.missouri.edu (accessed on 22 January 2022) | |
| Cystoliths | spheroidal phytoliths with contiguous verrucate sculpturing and stalk-like projections | Ulmaceae | (Kealhofer and Piperno 1998) |
| Moraceae | (Bozarth 1992) (Kealhofer and Piperno 1998) (Watling and Iriarte 2013) | ||
| Cannabaceae | (Bozarth 1992) (Watling and Iriarte 2013) | ||
| Urticaceae | (Bozarth 1992) | ||
| Boraginaceae (Cordia) | (Ricardo et al., 2018) | ||
| Bombacaceae | http://phytolith.missouri.edu (accessed on 22 January 2022) | ||
| Campanulaceae | (Lentfer 2003) | ||
| Papillae | Similar to sedge, but non-ornamented silicified papillae-like cells | Proteaceae | (Novello et al., 2018) |
| Baculate Elongates | baculate irregular bodies (named as boney bodies) The phytoliths have elaborated terminals that consist of a bunch of irregular protrusions on one end and one flatter end that is greater in diameter than the cylindrical stem | Burseraceae | (Watling and Iriarte 2013) (Piperno and McMichael 2020) |
| baculate irregular bodies without elaborated terminals | Achariaceae Burseraceae Chrysobalanaceae Dichapetalaceae Moraceae Sapindaceae | (Piperno and McMichael 2020) | |
| baculate rectangular | Achariaceae Chysobalanaceae Dichapetalaceae Euphorbiaceae Moraceae Phyllanthaceae Urticaceae | (Piperno and McMichael 2020) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, X.; Xie, B. Phytoliths from Woody Plants: A Review. Diversity 2022, 14, 339. https://doi.org/10.3390/d14050339
An X, Xie B. Phytoliths from Woody Plants: A Review. Diversity. 2022; 14(5):339. https://doi.org/10.3390/d14050339
Chicago/Turabian StyleAn, Xiaohong, and Binrong Xie. 2022. "Phytoliths from Woody Plants: A Review" Diversity 14, no. 5: 339. https://doi.org/10.3390/d14050339
APA StyleAn, X., & Xie, B. (2022). Phytoliths from Woody Plants: A Review. Diversity, 14(5), 339. https://doi.org/10.3390/d14050339