
Two New Species of Dacrymyces (Dacrymycetales, Basidiomycota) from Southwestern China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Morphological Studies
2.3. DNA Extraction, Polymerase Chain Reaction, and Sequencing
2.4. Sequence Alignment and Phylogenetic Analysis
3. Results
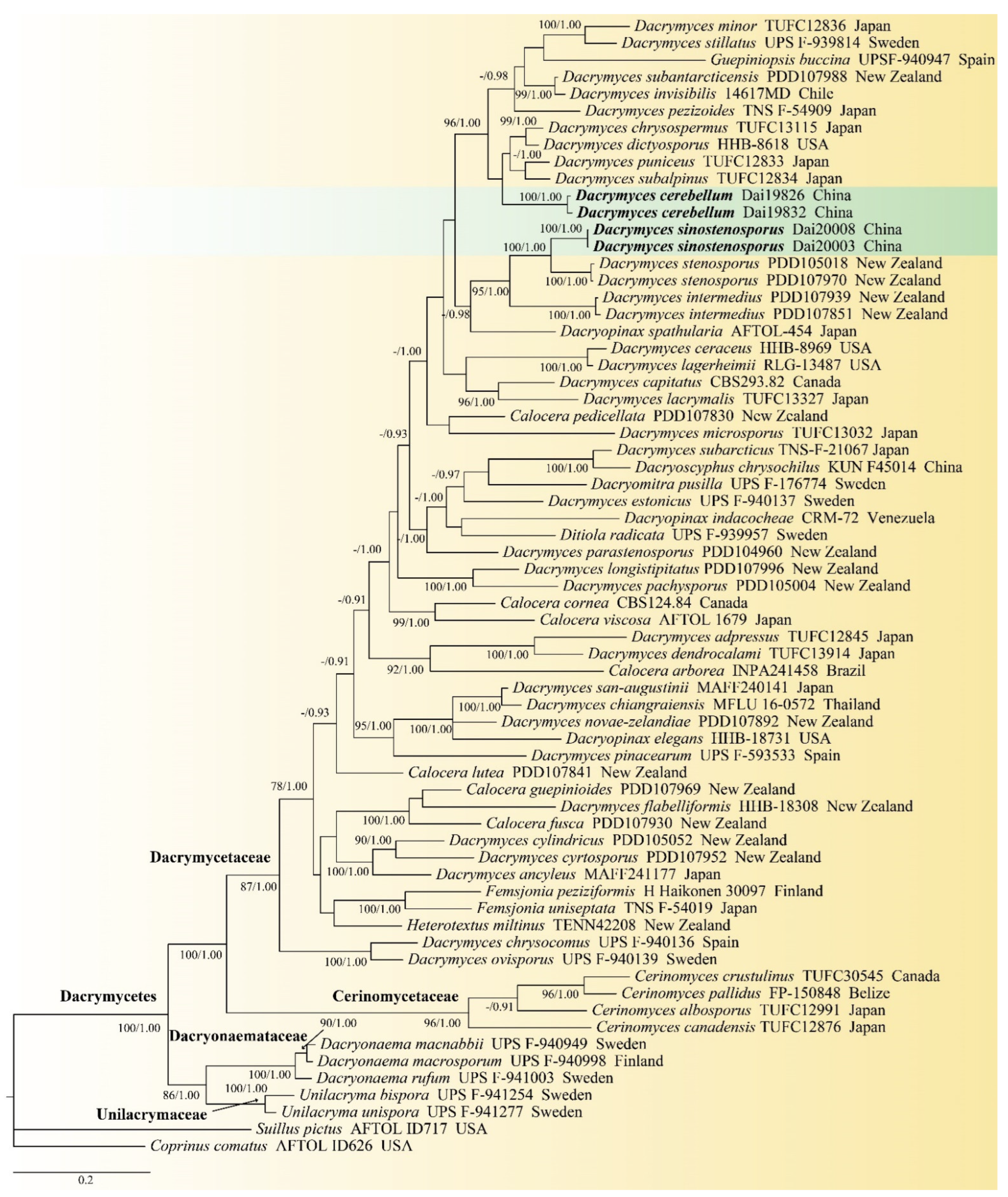
3.1. Phylogenetic Results
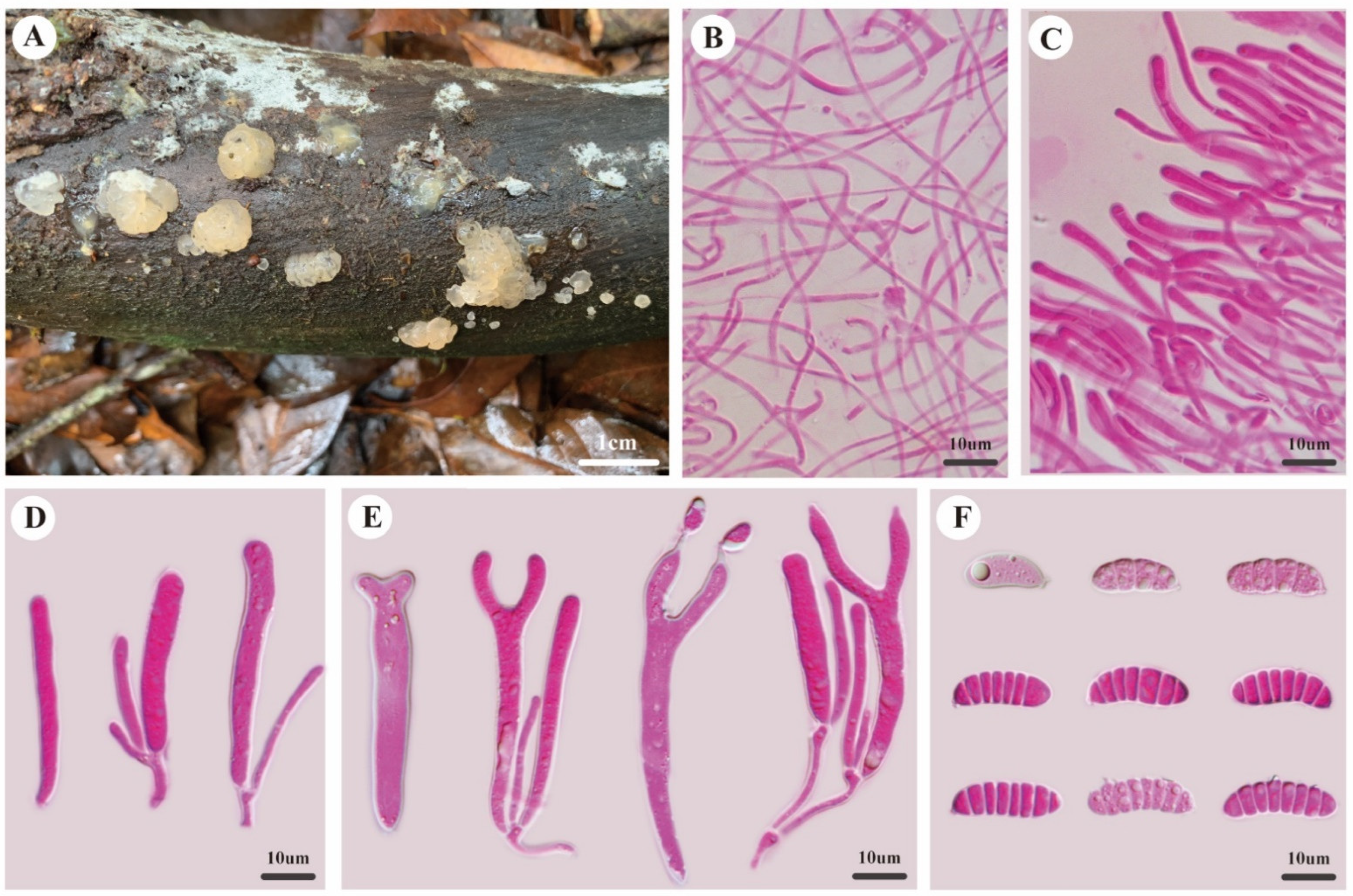
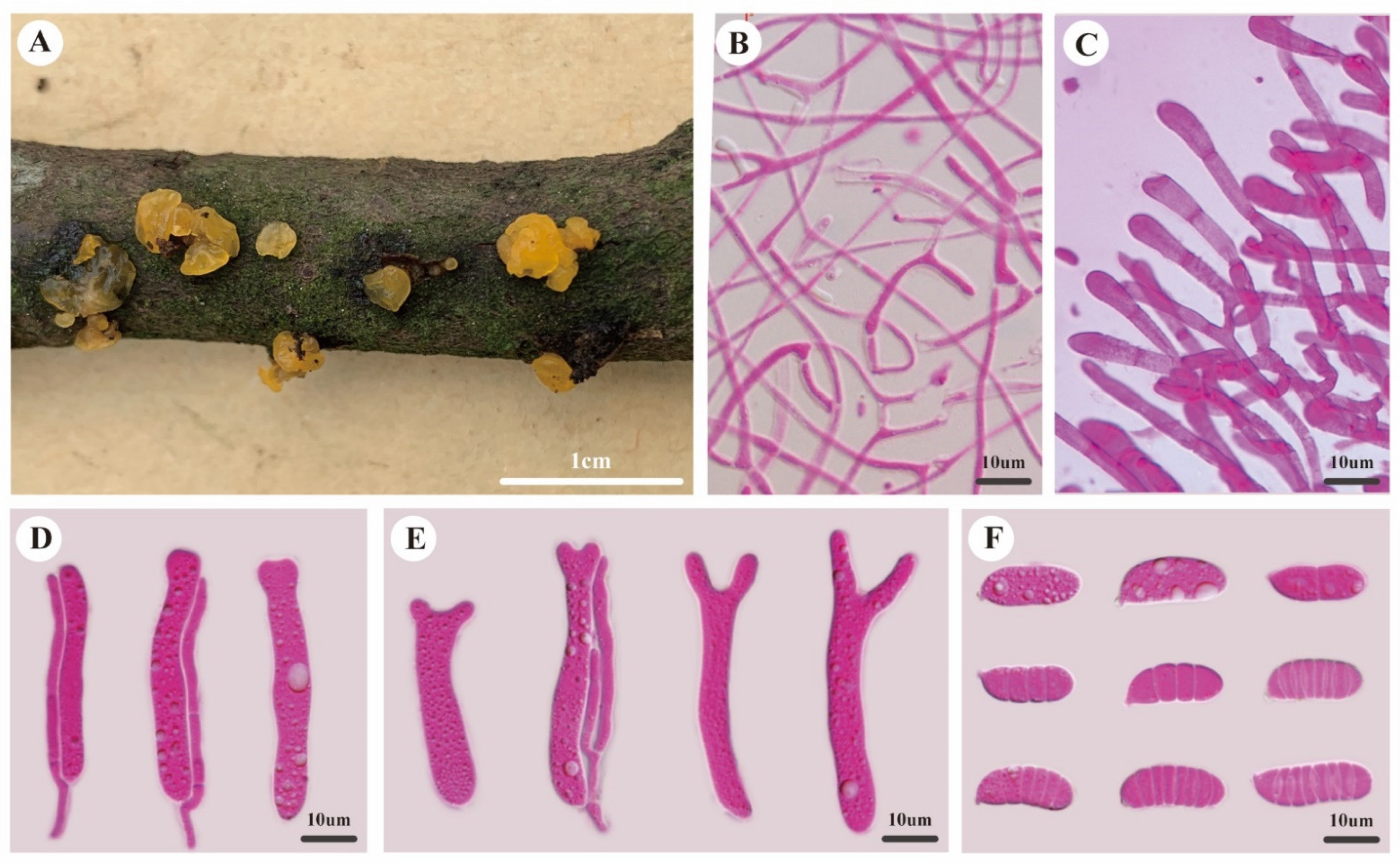
3.2. Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nees von Esenbeck, C.G. Das System der Pilze und Schwämme; Stahelschen Buchhandlung: Würtzburg, Germany, 1816; Volume 2, pp. 1–329. [Google Scholar]
- Fries, E.M. Systema Mycologicum; Officina Berlingiana: Berlin, Germany, 1822; Volume 2, pp. 1–275. [Google Scholar]
- Fries, E.M. Summa Vegetabilium Scandinaviae; Bonnier: Stockholm, Sweden, 1849; Volume 2, pp. 259–572. [Google Scholar]
- Martin, G.W. New or noteworthy tropical fungi. IV. Lloydia 1948, 11, 111–122. [Google Scholar]
- Martin, G.W. The genus Ceracea Cragin. Mycologia 1949, 41, 77–86. [Google Scholar] [CrossRef]
- McNabb, R.F.R. Taxonomic studies in the Dacrymycetaceae I. Cerinomyces Martin. New Zealand J. Bot. 1964, 2, 415–424. [Google Scholar] [CrossRef]
- McNabb, R.F.R. Taxonomic studies in the Dacrymycetaceae III. Dacryopinax Martin. New Zealand J. Bot. 1965, 3, 59–72. [Google Scholar] [CrossRef] [Green Version]
- McNabb, R.F.R. Taxonomic studies in the Dacrymycetaceae VI. Femsjonia Fries. New Zealand J. Bot. 1965, 3, 223–228. [Google Scholar] [CrossRef]
- McNabb, R.F.R. Taxonomic studies in the Dacrymycetaceae VII. Ditiola Fries. New Zealand J. Bot. 1966, 4, 546–558. [Google Scholar] [CrossRef]
- Kobayasi, Y. On the Dacrymyces-group (Fungorum ordinis Tremellalium studia monographica III). Sci. Rep. Tokyo Bunrika Daigaku Sect. B 1939, 4, 105–128. [Google Scholar]
- Mcnabb, R.F.R. Taxonomic studies in the Dacrymycetaceae VIII. Dacrymyces Nees ex Fries. N. Z. J. Bot. 1973, 11, 461–564. [Google Scholar] [CrossRef]
- Liu, B.; Fan, L. Two new species of Dacrymycetaceae from China. Acta Microbiol. Sin. 1989, 8, 22–24. [Google Scholar]
- Liu, B.; Fan, L. Four new species and one new variety of Dacrymycetaceae from China. Acta Microbiol. Sin. 1990, 9, 12–19. [Google Scholar]
- Shirouzu, T.; Hirose, D.; Tokumasu, S. Taxonomic study of the Japanese Dacrymycetes. Persoonia 2009, 23, 16–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirouzu, T.; Hirose, D.; Oberwinkler, F.; Shimomura, N.; Maekawa, N.; Tokumasu, S. Combined molecular and morphological data for improving phylogenetic hypothesis in Dacrymycetes. Mycologia 2013, 105, 1110–1125. [Google Scholar] [CrossRef]
- Shirouzu, T.; Hosaka, K.; Nam, K.O.; Weir, B.S.; Johnston, P.R.; Hosoya, T. Phylogenetic relationships of eight new Dacrymycetes collected from New Zealand. Persoonia 2017, 38, 156–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamora, J.C.; Ekman, S. Phylogeny and character evolution in the Dacrymycetes, and systematics of Unilacrymaceae and Dacryonaemataceae fam. nov. Persoonia 2020, 44, 161–205. [Google Scholar] [CrossRef] [PubMed]
- Savchenko, A.; Zamora, J.C.; Shirouzu, T.; Spirin, V.; Malysheva, V.; Kõljalg, U.; Miettinen, O. Revision of Cerinomyces (Dacrymycetes, Basidiomycota) with notes on morphologically and historically related taxa. Stud. Mycol. 2021, 99, 100117. [Google Scholar] [CrossRef] [PubMed]
- Shirouzu, T.; Hirose, D.; Tokumasu, S. Sequence analyses of the 28S rRNA gene D1/D2 region suggest Dacrymyces (Heterobasidiomycetes, Dacrymycetales) is polyphyletic. Mycoscience 2007, 48, 388–394. [Google Scholar] [CrossRef]
- Dai, Y.C.; Yang, Z.L.; Cui, B.K.; Wu, G.; Yuan, H.S.; Zhou, L.W.; He, S.H.; Ge, Z.W.; Wu, F.; Wei, Y.L.; et al. Diversity and systematics of the important macrofungi in Chinese forests. Mycosystema 2021, 40, 770–805. [Google Scholar] [CrossRef]
- Fan, L.F.; Wu, Y.D.; Wu, F.; Dai, Y.C. Calocera tibetica sp. nov. (Dacrymycetaceae, Dacrymycetales) from southwestern China. Phytotaxa 2021, 500, 133–141. [Google Scholar] [CrossRef]
- Petersen, J.H. Farvekort. In The Danish Mycological Society’s Colour-Chart; Foreningen til Svampekundskabens Fremme: Greve, Denmark, 1996. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols, a Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analysis with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Silvestro, D.; Michalak, I. Raxmlgui: A graphical front-end for raxml. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burdsall, H.; Laursen, G.A.; Cripps, C.L. Fungi of New Zealand’s subantarctic islands. I: Two new species of Dacrymyces (Basidiomycotina). In Fungi in Forest Ecosystems: Systematics, Diversity and Ecology; Miller, O.K., Cripps, C.L., Eds.; New York Botanical Garden: New York, NY, USA, 2004; pp. 107–111. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Species | Locality | Sample No. | GenBank Accession No. | |
|---|---|---|---|---|
| ITS | nrLSU | |||
| Calocera arborea | Brazil | INPA 241458 | AB744230 | AB723514 |
| Calocera cornea | Canada | CBS124.84 | AB712437 | AB472738 |
| Calocera fusca | New Zealand | PDD 107930 | LC131405 | LC131364 |
| Calocera guepinioides | New Zealand | PDD 107969 | LC131411 | LC131370 |
| Calocera lutea | New Zealand | PDD 107841 | LC131413 | LC131372 |
| Calocera pedicellata | New Zealand | PDD 107830 | LC131415 | LC131374 |
| Calocera viscosa (Type species) | Japan | AFTOL-ID 1679 | DQ520102 | DQ520102 |
| Cerinomyces albosporus | Japan | TUFC12991 | AB712440 | AB299050 |
| Cerinomyces canadensis | Japan | TUFC12876 | AB712441 | AB472696 |
| Cerinomyces crustulinus | Canada | TUFC 30545 | AB712443 | AB712423 |
| Cerinomyces pallidus (Type species) | Belize | FP-150848 | AB712446 | AB712426 |
| Dacrymyces adpressus | Japan | TUFC12845 | AB712447 | AB472707 |
| Dacrymyces ancyleus | Japan | MAFF241177 | AB712448 | AB472713 |
| Dacrymyces capitatus | Canada | CBS293.82 | AB712450 | AB472741 |
| Dacrymyces ceraceus | USA | HHB-8969 | AB712442 | AB712422 |
| Dacrymyces cerebriformis | China | Dai 19826 | OM955201 | OM955196 |
| Dacrymyces cerebriformis | China | Dai 19832 | OM955202 | OM955197 |
| Dacrymyces chiangraiensis | Thailand | MFLU:16-0572 | KY498587 | – |
| Dacrymyces chrysocomus | Spain | UPS F-940136 | MN595629 | MN595629 |
| Dacrymyces chrysospermus | Japan | TUFC 13115 | AB712452 | AB299073 |
| Dacrymyces cylindricus | New Zealand | PDD 105052 | LC131419 | LC131378 |
| Dacrymyces cyrtosporus | New Zealand | PDD 107952 | LC131421 | LC131380 |
| Dacrymyces dendrocalami | Japan | TUFC13914 | AB712453 | AB712428 |
| Dacrymyces dictyosporus | USA | HHB-8618 | AB712454 | AB712429 |
| Dacrymyces estonicus | Sweden | UPS F-940137 | MN595632 | MN595632 |
| Dacrymyces flabelliformis | New Zealand | HHB-18308 | AB712455 | AB712430 |
| Dacrymyces intermedius | New Zealand | PDD 107939 | – | LC131385 |
| Dacrymyces intermedius | New Zealand | PDD 107851 | – | LC131384 |
| Dacrymyces invisibilis | Chile | 14617MD | MH230101 | MH230103 |
| Dacrymyces lacrymalis | Japan | TUFC13327 | AB712456 | AB299069 |
| Dacrymyces lagerheimii | USA | RLG-13487 | AB712445 | AB712425 |
| Dacrymyces longistipitatus | New Zealand | PDD 107996 | LC131425 | LC131386 |
| Dacrymyces microsporus | Japan | TUFC 13032 | AB712457 | AB472712 |
| Dacrymyces minor | Japan | TUFC 12836 | AB712458 | AB299063 |
| Dacrymyces novae-zelandiae | New Zealand | PDD 107892 | LC131427 | LC131390 |
| Dacrymyces ovisporus | Sweden | UPS F-940139 | MN595635 | MN595635 |
| Dacrymyces pachysporus | New Zealand | PDD 105004 | LC131429 | LC131392 |
| Dacrymyces parastenosporus | New Zealand | PDD 104960 | LC131431 | LC131394 |
| Dacrymyces pezizoides | Japan | TNS-F-54909 | LC386890 | LC386894 |
| Dacrymyces pinacearum | Japan | UPS F-593533 | MN595637 | MN595637 |
| Dacrymyces puniceus | Japan | TUFC 12833 | AB712449 | AB299057 |
| Dacrymyces san-augustinii | Japan | MAFF240141 | AB712463 | AB299081 |
| Dacrymyces sinostenosporus | China | Dai 20003 | MW540888 | MW540890 |
| Dacrymyces sinostenosporus | China | Dai 20008 | MW540889 | MW540891 |
| Dacrymyces stenosporus | New Zealand | PDD 105018 | LC131433 | LC131396 |
| Dacrymyces stenosporus | New Zealand | PDD 107970 | LC131434 | LC131397 |
| Dacrymyces stillatus (Type species) | Sweden | UPS F-939814 | MN595677 | MN595677 |
| Dacrymyces subalpinus | Japan | TUFC 12834 | AB712465 | AB299060 |
| Dacrymyces subantarcticensis | New Zealand | PDD 107988 | LC131436 | LC131400 |
| Dacrymyces subarcticus | Japan | TNS-F-21067 | AB712467 | AB472727 |
| Dacryomitra pusilla (Type species) | Sweden | UPS F-176774 | MN595639 | MN595639 |
| Dacryonaema macnabbii | Sweden | UPS F-940949 | MN595650 | MN595650 |
| Dacryonaema macrosporum | Finland | UPS F-940998 | MN595660 | MN595660 |
| Dacryonaema rufum (Type species) | Sweden | UPS F-941003 | MN595645 | MN595645 |
| Dacryopinax elegans (Type species) | USA | HHB-18731 | AB712471 | AB712433 |
| Dacryopinax indacocheae | Venezuela | CRM-72 | AB712472 | AB712434 |
| Dacryopinax spathularia | Japan | AFTOL-ID 454 | AY854070 | AY701525 |
| Dacryoscyphus chrysochilus (Type species) | China | KUN F45014 | – | AY604567 |
| Ditiola radicata (Type species) | Sweden | UPS F-939957 | MN595641 | MN595641 |
| Femsjonia peziziformis | Finland | H Haikonen 30097 | MN595642 | MN595642 |
| Femsjonia uniseptata | Japan | TNS-F-54019 | LC222844 | LC222843 |
| Guepiniopsis buccina | Spain | UPS F-940947 | MN595643 | MN595643 |
| Heterotextus miltinus | New Zealand | TENN 42208 | MN595644 | MN595644 |
| Unilacryma bispora | Sweden | UPS F-941254 | MN595673 | MN595673 |
| Unilacryma unispora (Type species) | Sweden | UPS F-941277 | MN595665 | MN593500 |
| Coprinus comatus | USA | AFTOL-ID 626 | AY854066 | AY635772 |
| Suillus pictus | USA | AFTOL-ID 717 | AY854069 | AY684154 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lian, Y.-P.; Tohtirjap, A.; Wu, F. Two New Species of Dacrymyces (Dacrymycetales, Basidiomycota) from Southwestern China. Diversity 2022, 14, 379. https://doi.org/10.3390/d14050379
Lian Y-P, Tohtirjap A, Wu F. Two New Species of Dacrymyces (Dacrymycetales, Basidiomycota) from Southwestern China. Diversity. 2022; 14(5):379. https://doi.org/10.3390/d14050379
Chicago/Turabian StyleLian, Ya-Ping, Ablat Tohtirjap, and Fang Wu. 2022. "Two New Species of Dacrymyces (Dacrymycetales, Basidiomycota) from Southwestern China" Diversity 14, no. 5: 379. https://doi.org/10.3390/d14050379
APA StyleLian, Y. -P., Tohtirjap, A., & Wu, F. (2022). Two New Species of Dacrymyces (Dacrymycetales, Basidiomycota) from Southwestern China. Diversity, 14(5), 379. https://doi.org/10.3390/d14050379