
The Phylogeny of Rays and Skates (Chondrichthyes: Elasmobranchii) Based on Morphological Characters Revisited

 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
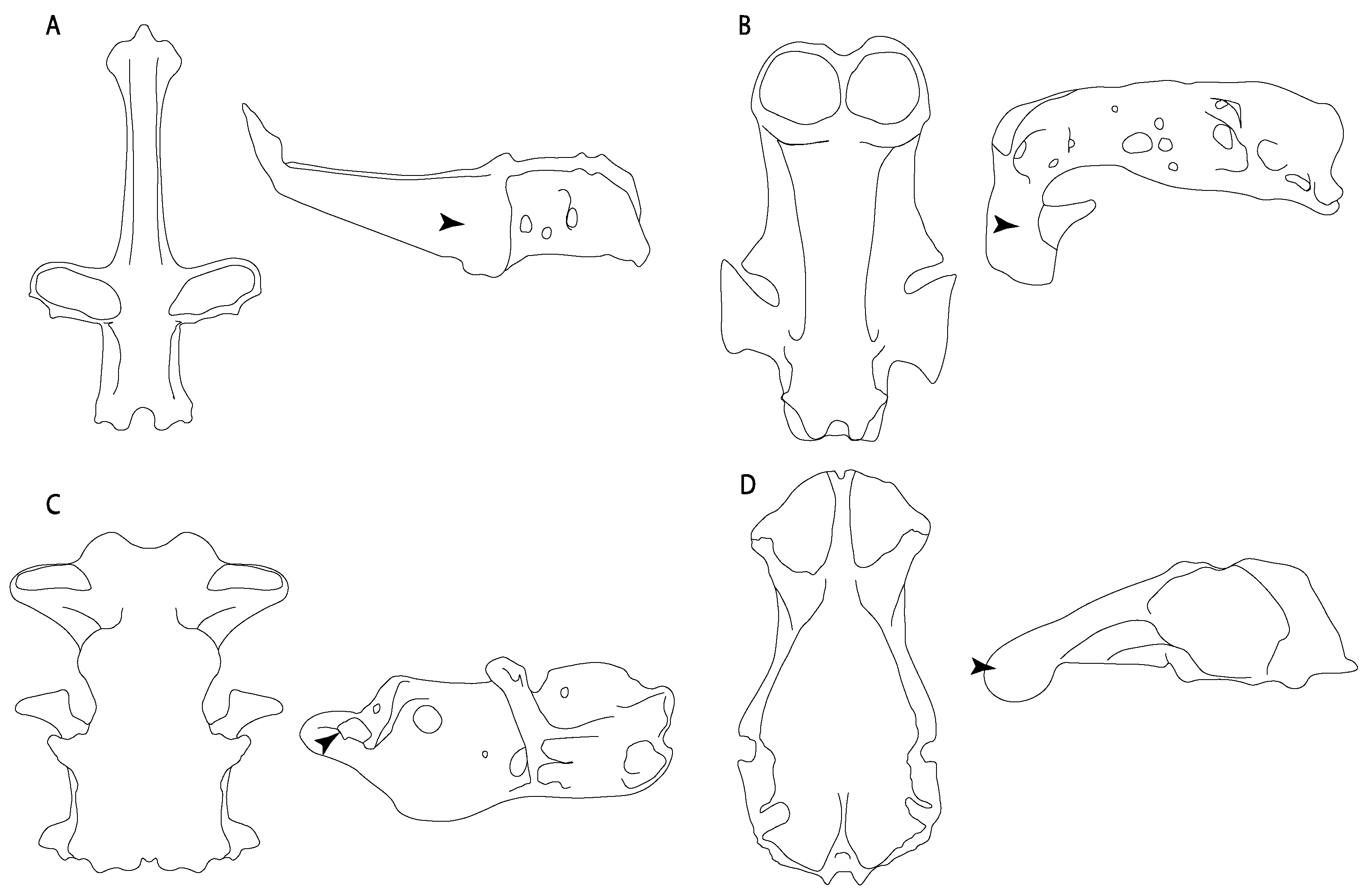{kind=link}
{kind=link}
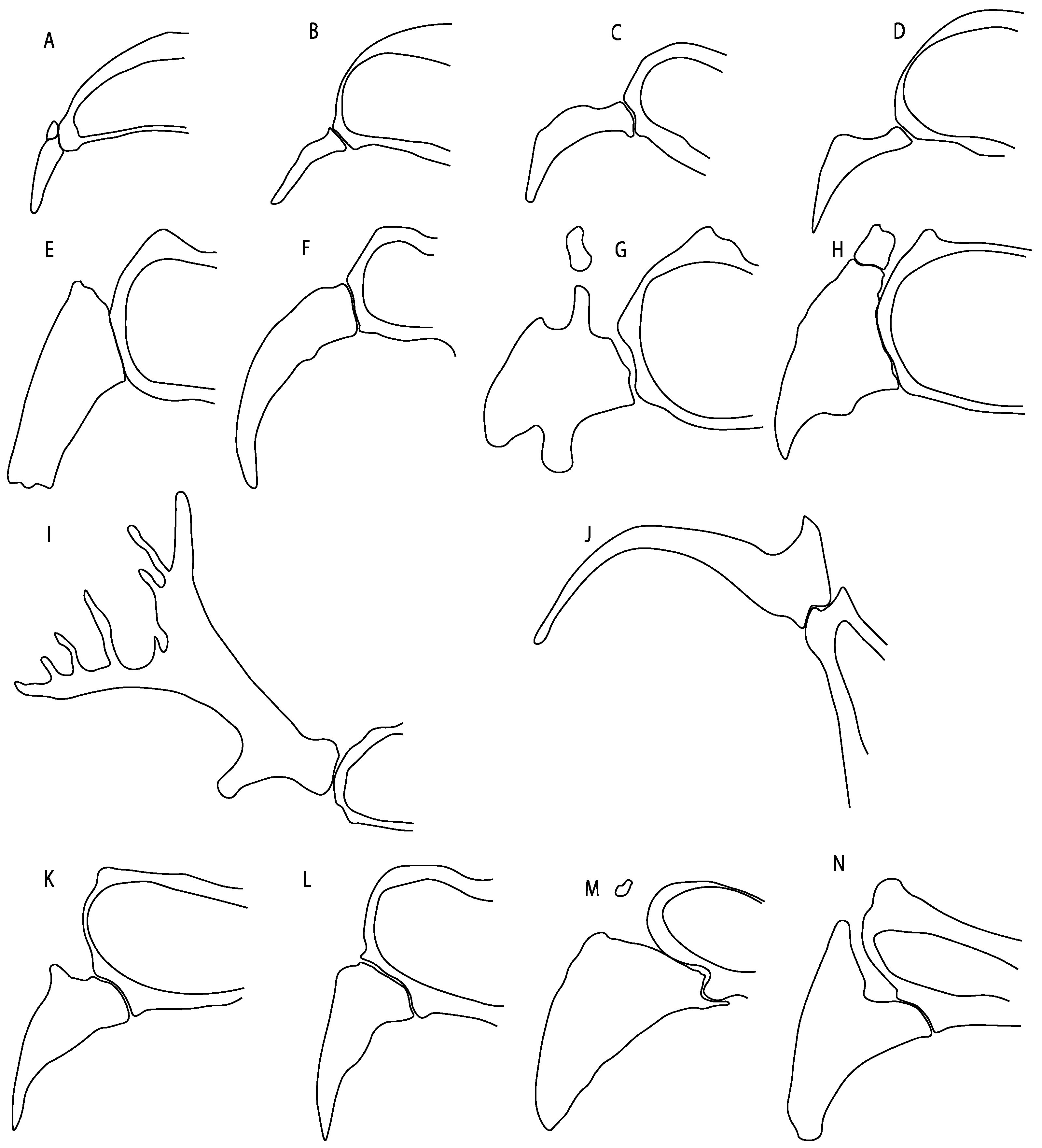{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Matrix Revision
3.1.1. External Morphological Structures
- 31.
- 32.
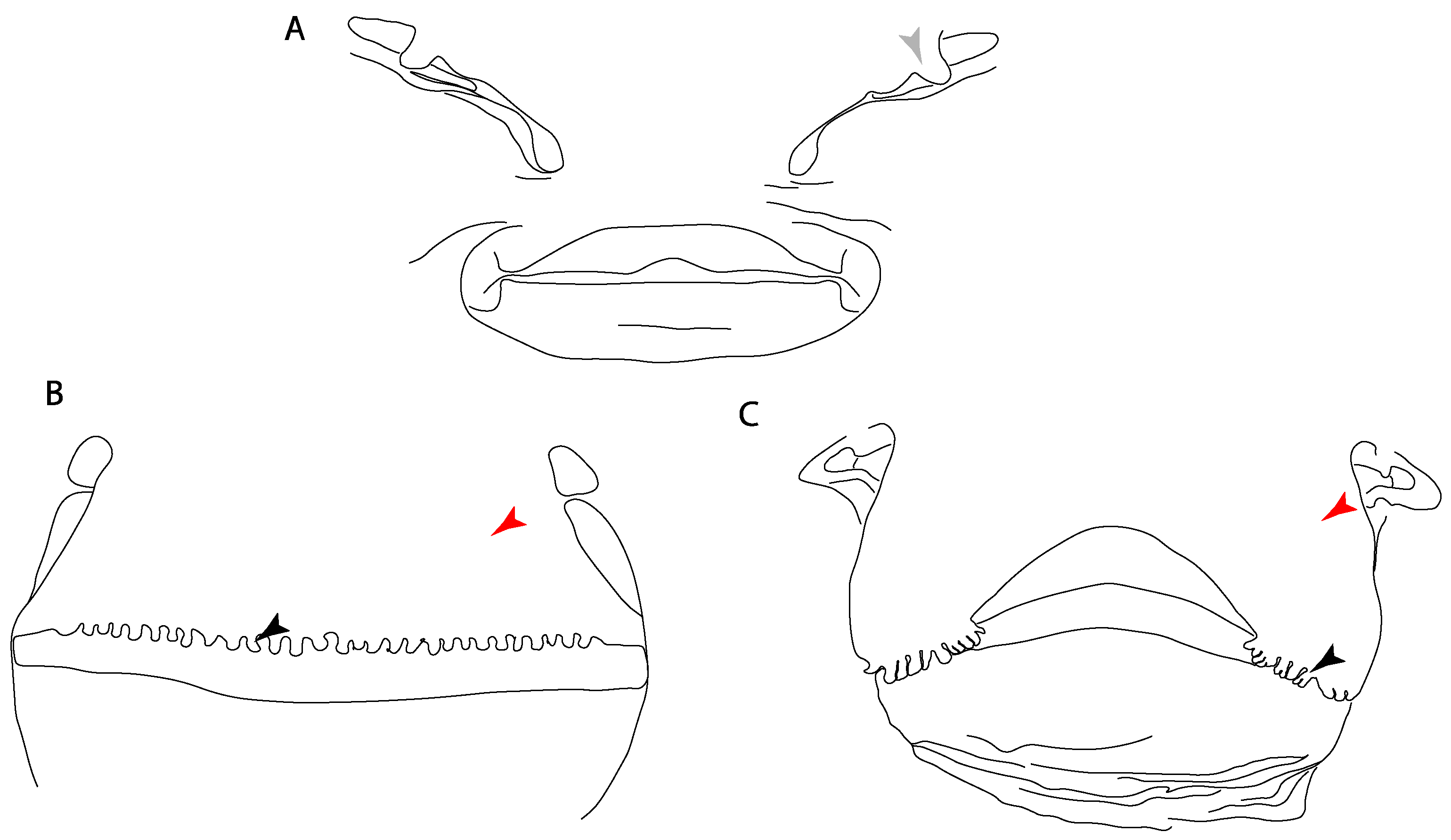
- Anterior nasal lobe: (0) Fails to cover most of the medial half of the naris; (1) covers more than the medial half of the naris.
- 33.
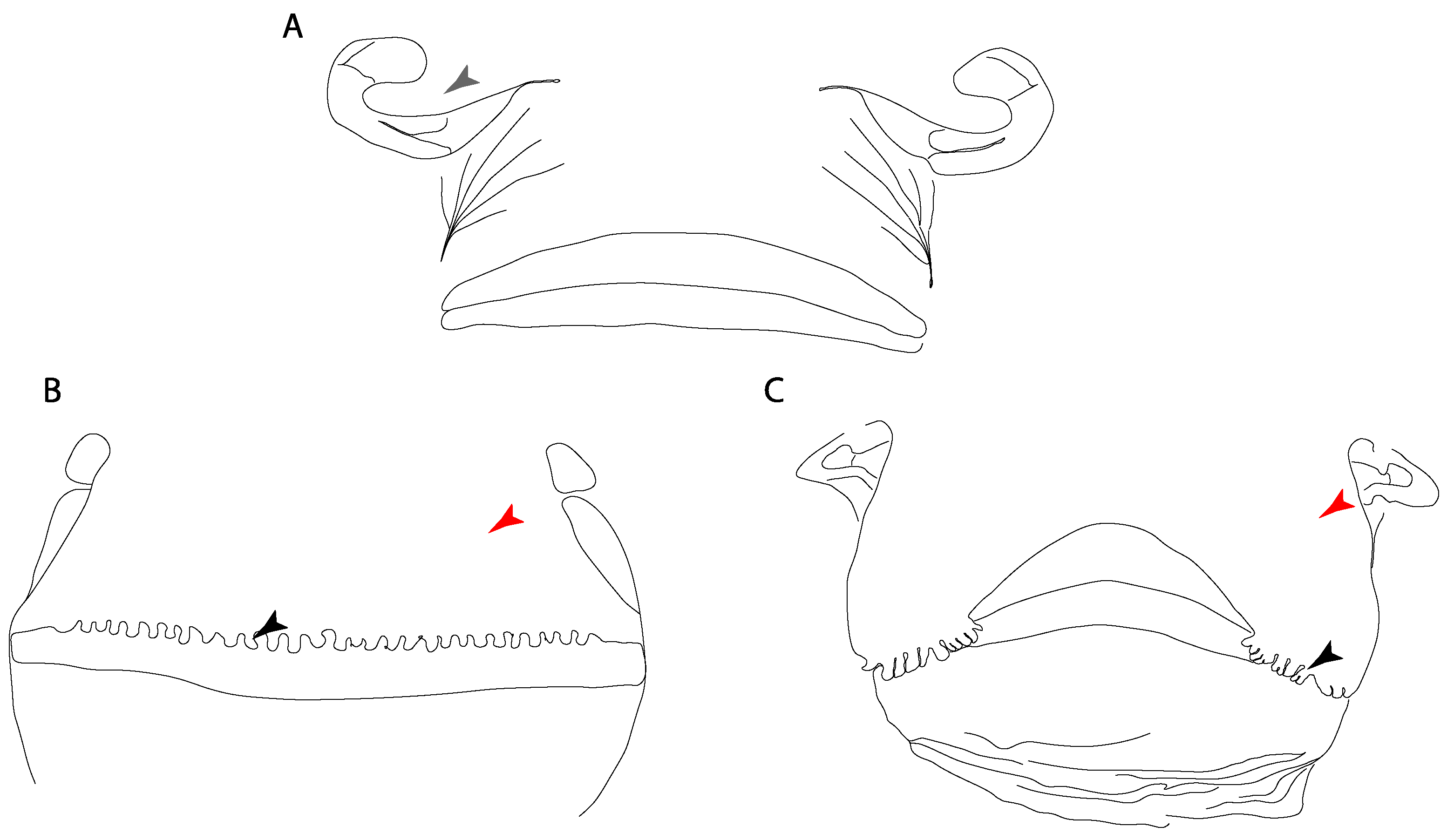
- Nasal curtain fringes: (0) Absent; (1) present (new).
3.1.2. Lateral Line
- 83.
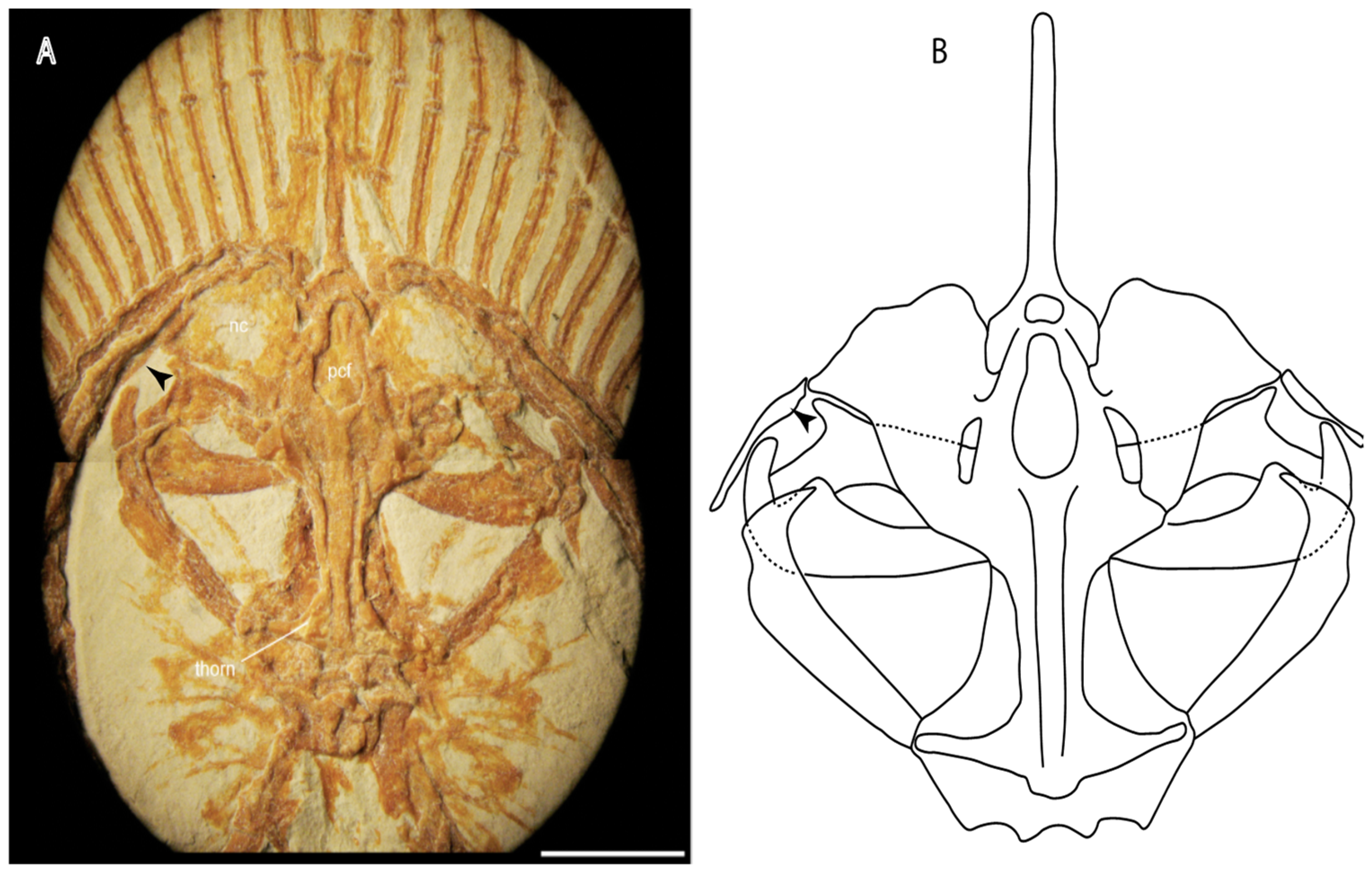
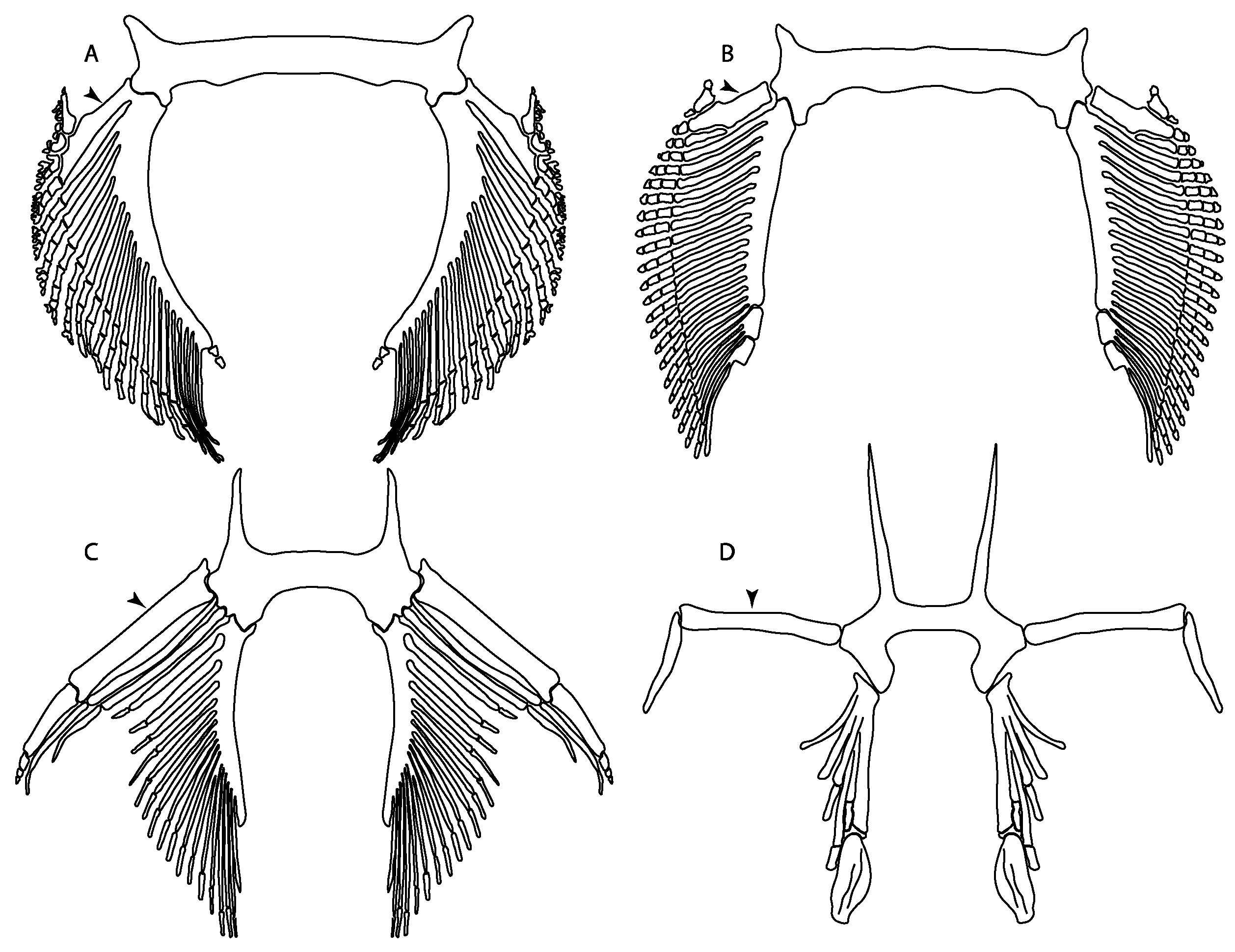
- Abdominal canal on coracoid bar: (0) Absent; (1) present. Modified from Aschliman et al. [7] (char. 24). The original character was split into two different ones (83–84) to increase the grouping information regarding the variation on the canals on the coracoid bar. The coding for Pristis was changed based on Wueringer et al. Text-Figure 1 in [54].
- 84.
- Abdominal canal on coracoid bar (if present): (0) Groove, cephalic lateral line forms abdominal canal on coracoid bar; (1) pores.
3.1.3. Neurocranium
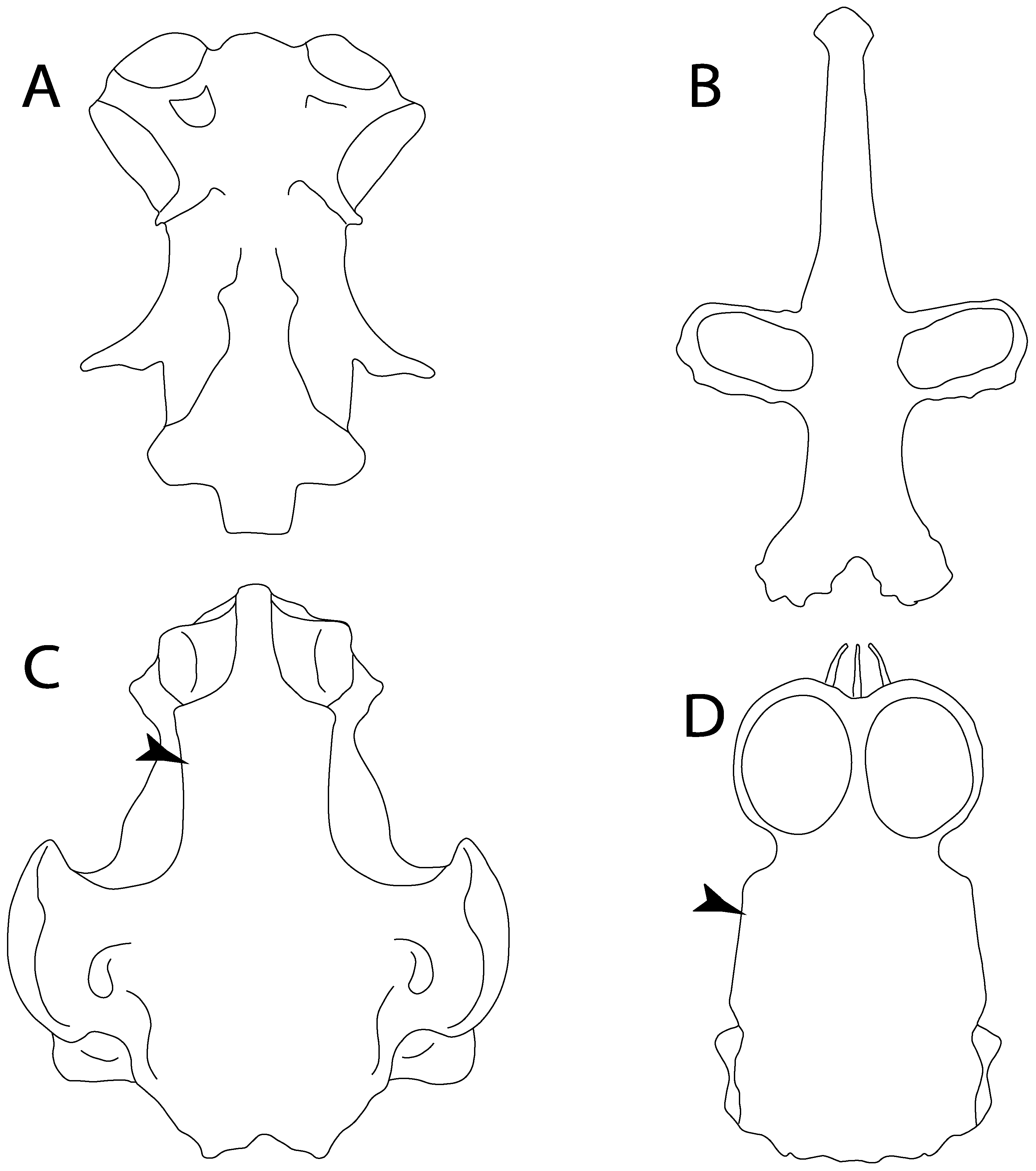
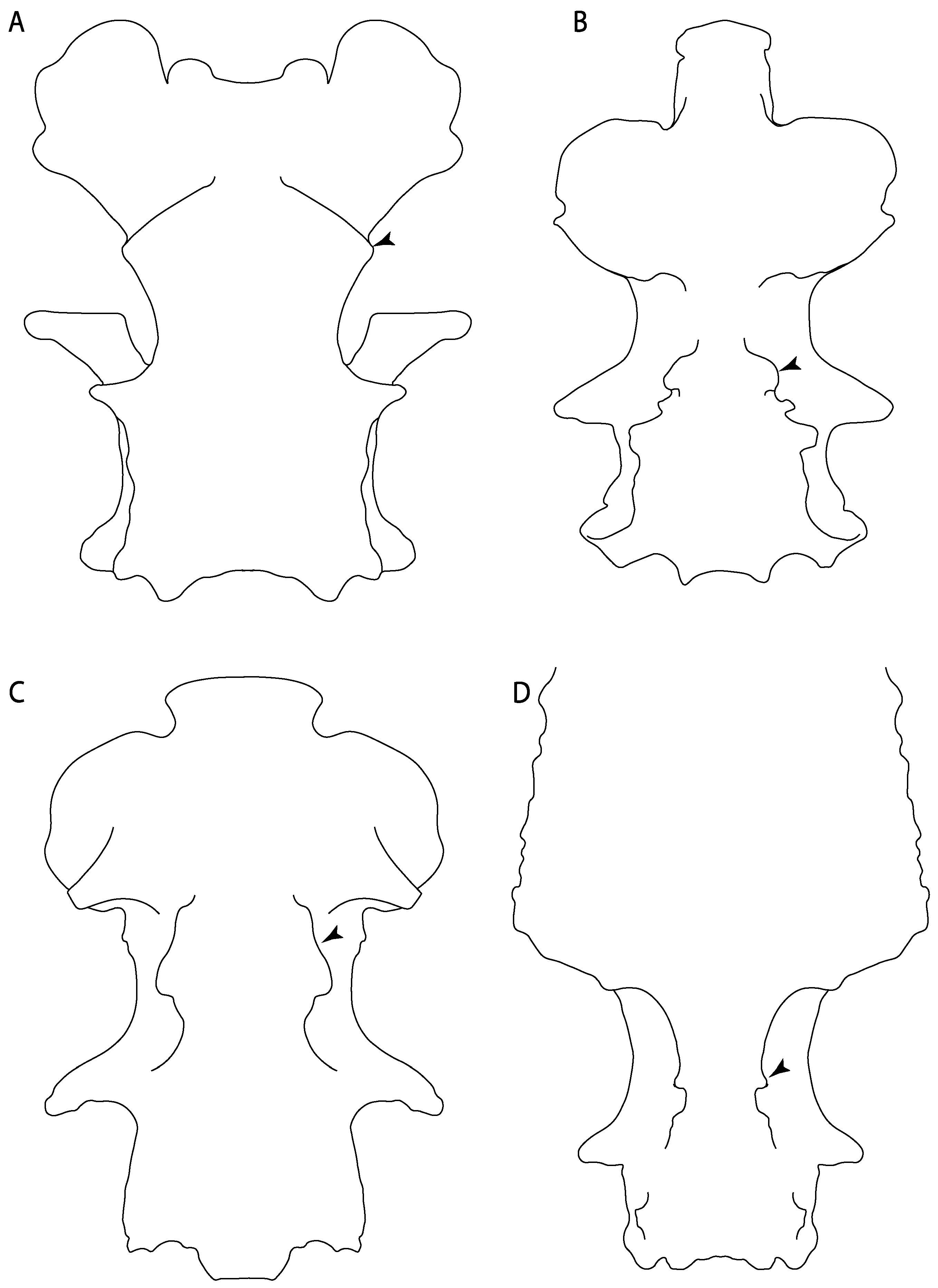
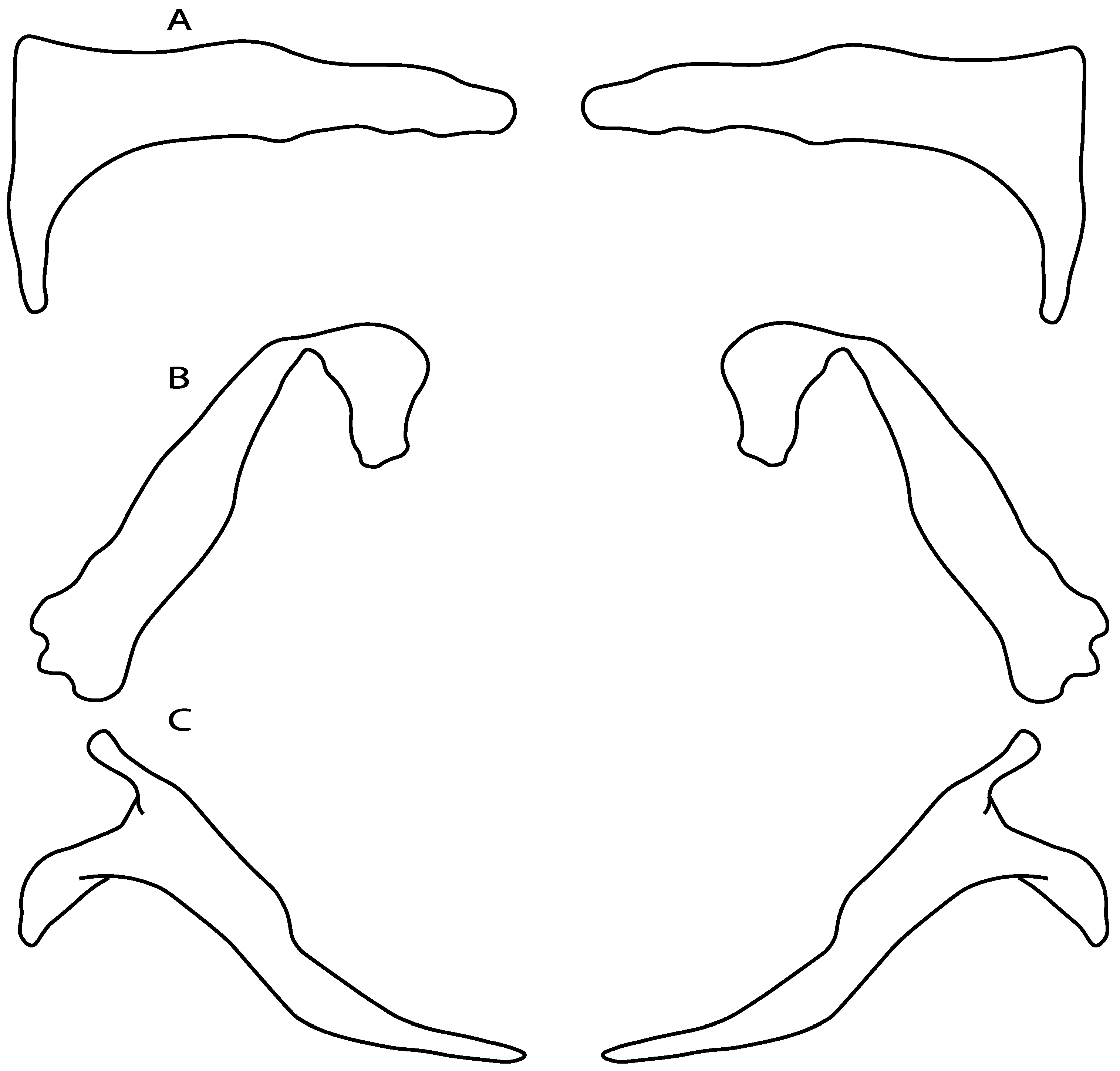
- 3.
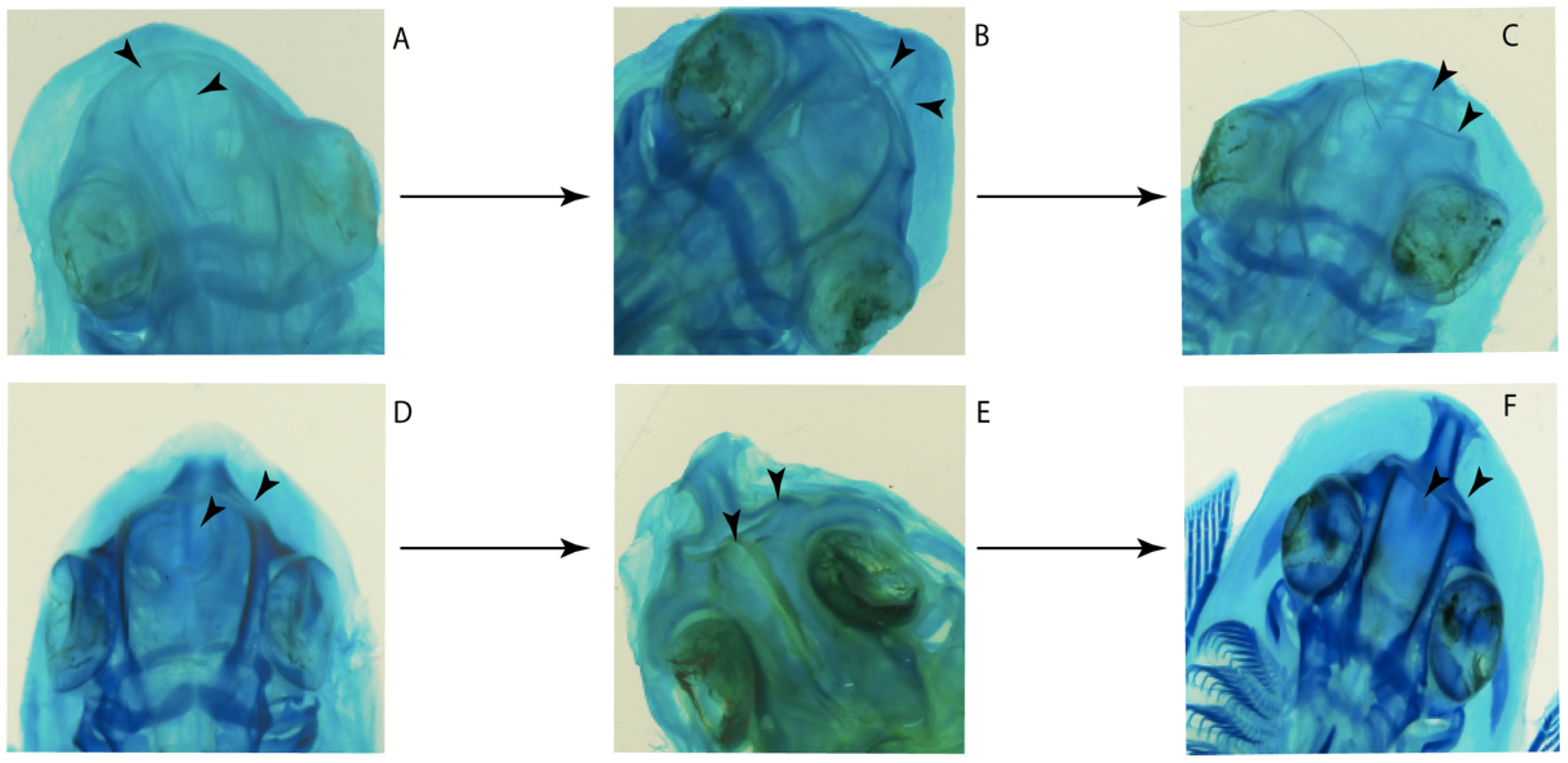
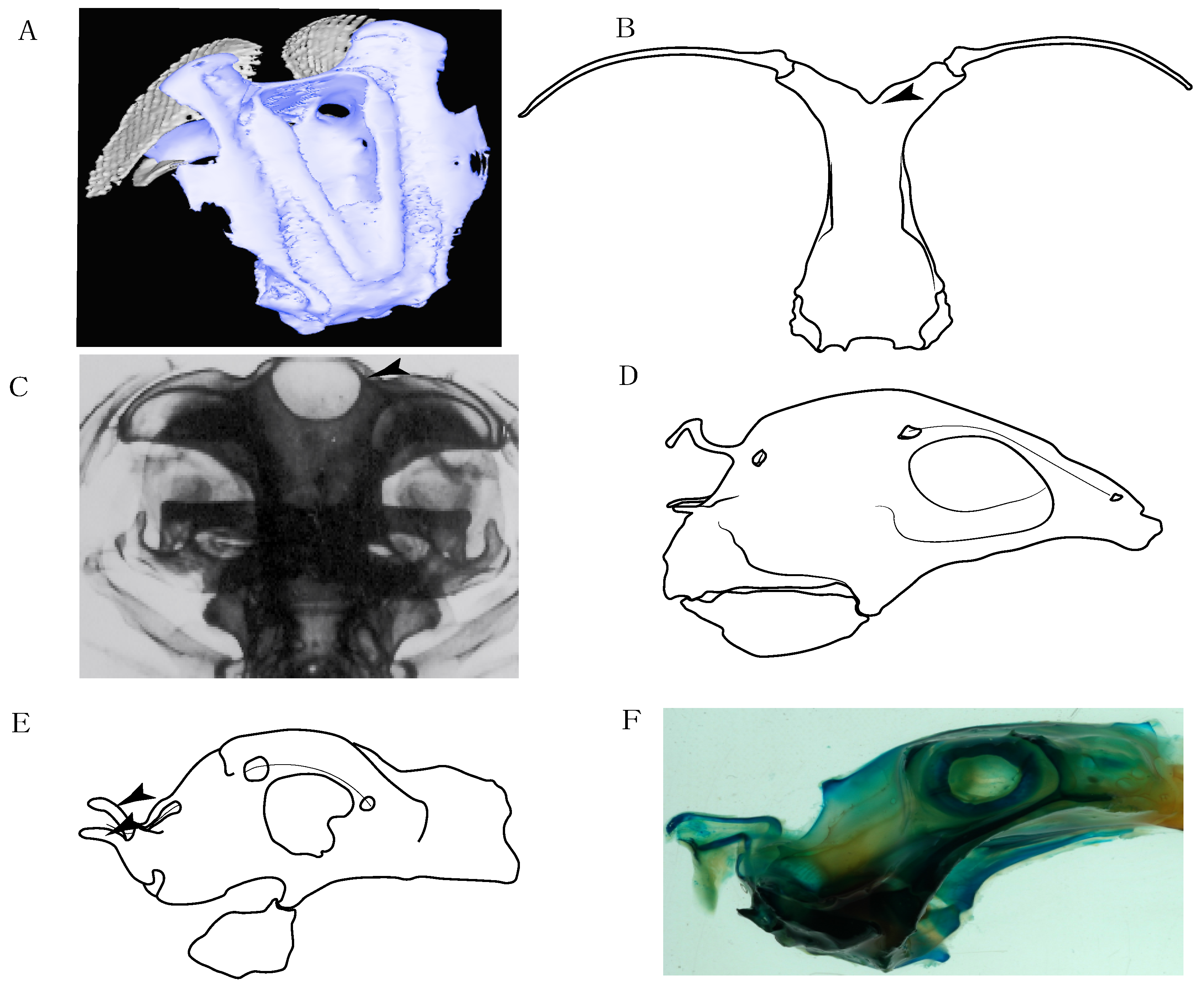
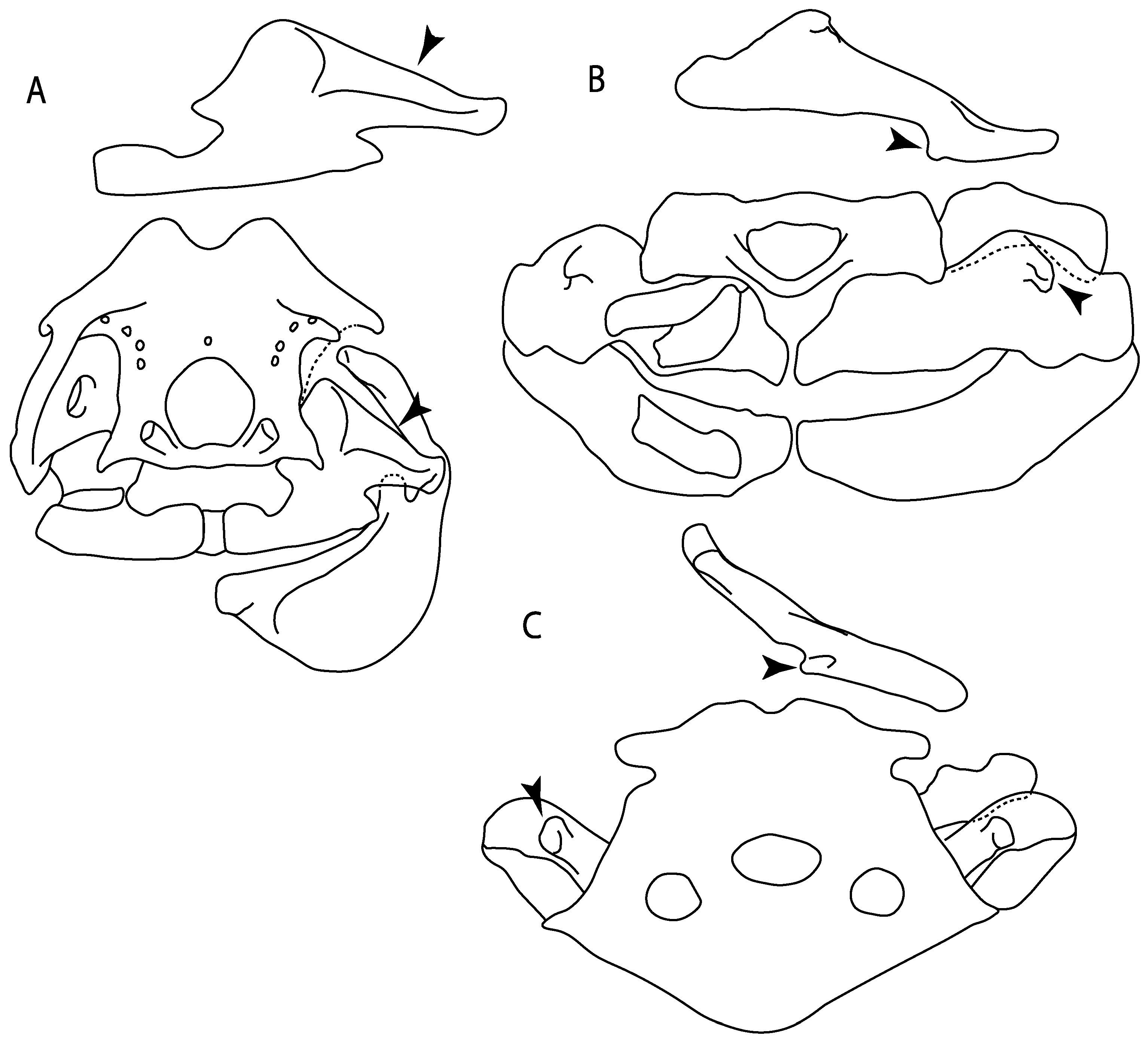
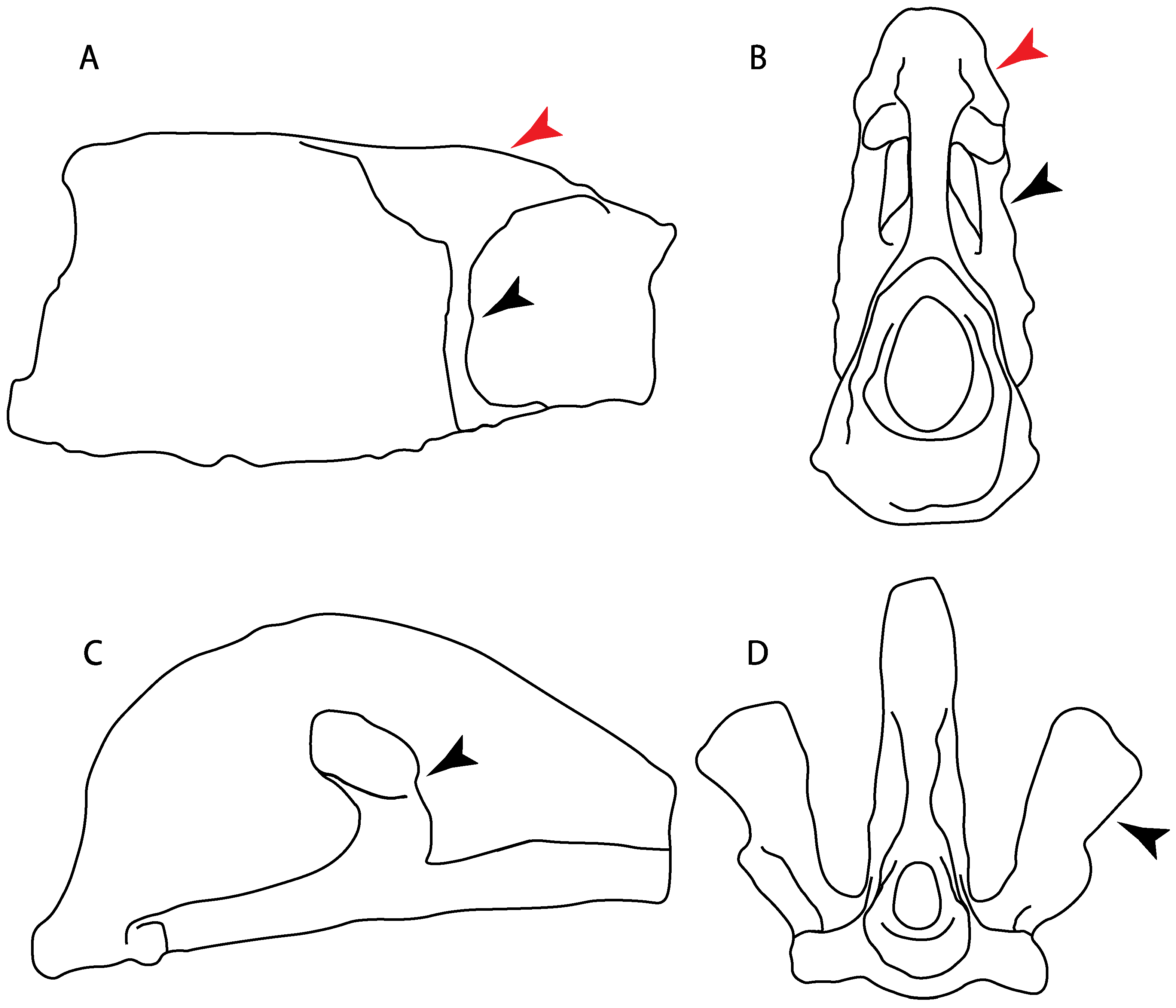
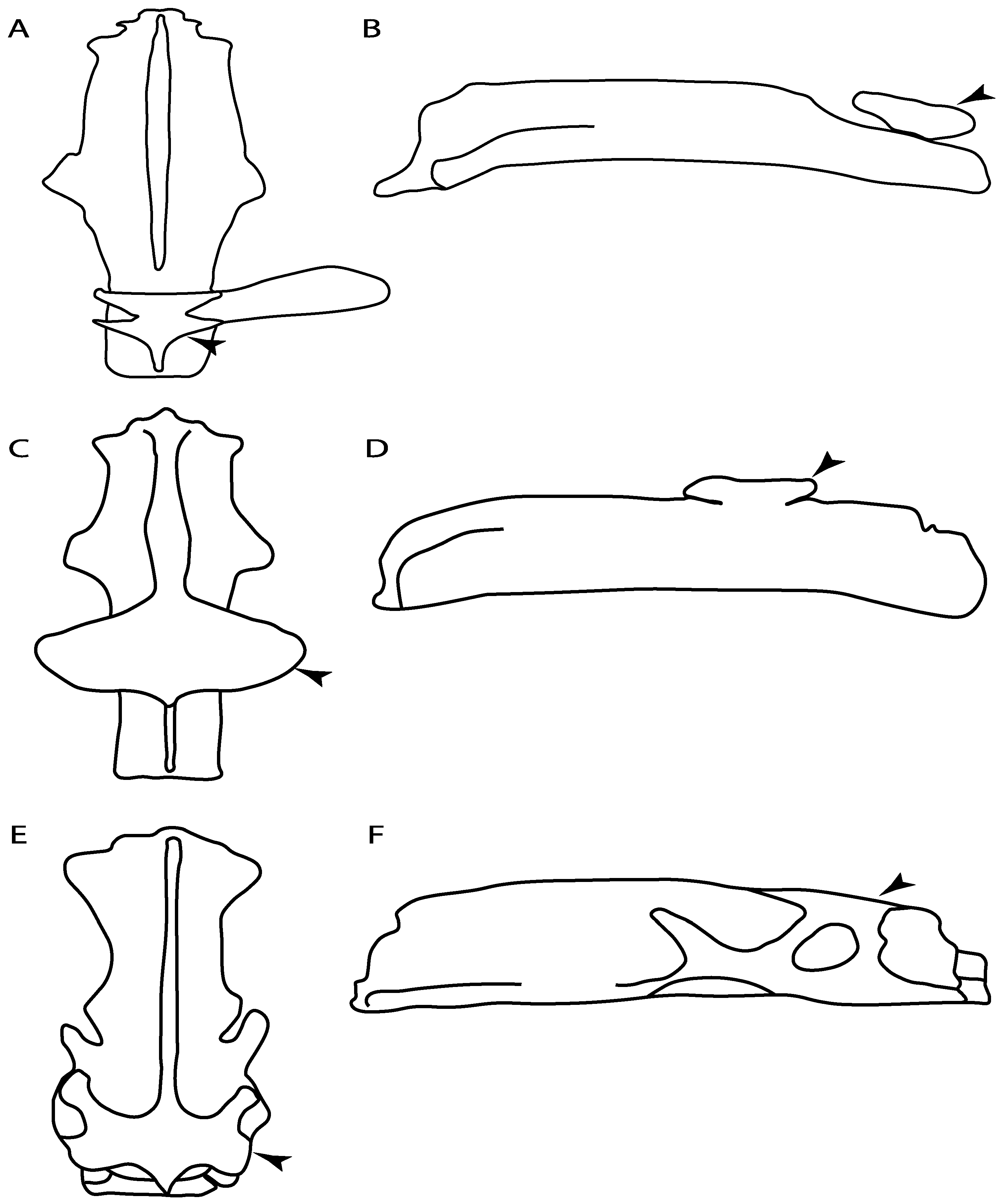
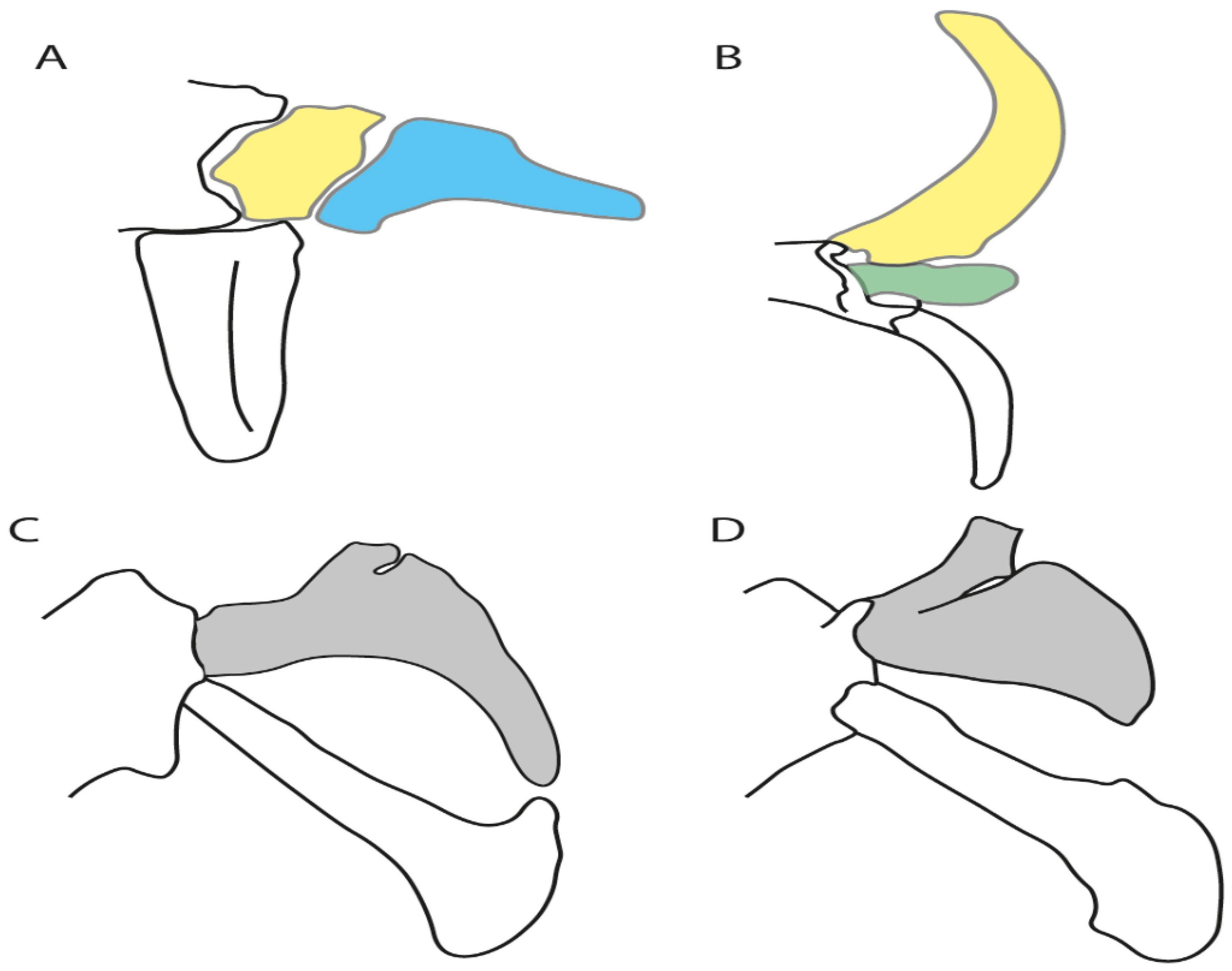
- Rostral cartilages: (0) Arise from the medial area of the trabecula only; (1) medial area of the trabecula + lamina orbitonasalis. According to De Beer and Moy-Thomas [55] and Miyake et al. [56], no evidence suggests the homology between the rostral cartilages in elasmobranchs and holocephalians, as the rostral cartilages of holocephalians arise from the medial area of the trabecula possibly without any contribution from the lamina orbitonasalis. Conversely, in most modern elasmobranchs, these two embryological cartilages (medial trabecula and the lamina orbitonasalis) contribute to the formation of the rostral cartilages (Figure 4).
- 4.
- Rostral cartilage: (0) Well-developed rostral plate with various degrees of contribution from the lamina orbitonasalis; (1) reaches the tip of the snout (carried by the growth of the pectoral fin); (2) reaches the tip of the snout (growth of lamina orbitonasalis to support the cephalic fins). Modified from Aschliman et al. [7] (char. 26). This character aims to include the variation observed in Neoselachii (sensu [12]), following observations made by Miyake et al. [56], Maisey [57] and Lane [58]. Many taxa remain uncharacterized as the present coding provides grouping information for those taxa whose rostral cartilages arise from the interaction between the medial area of the trabecula and lamina orbitonasalis.
- 5.
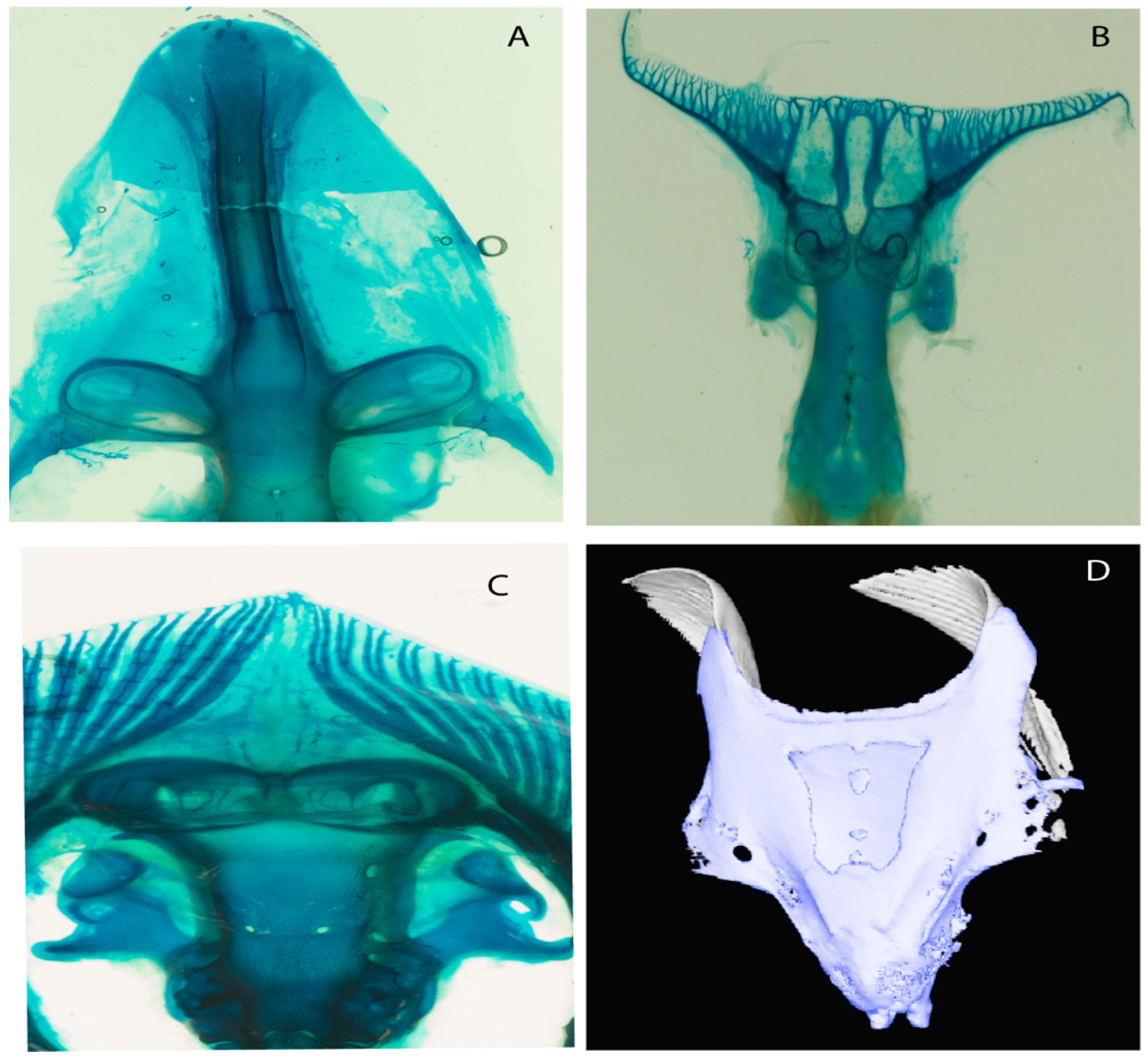
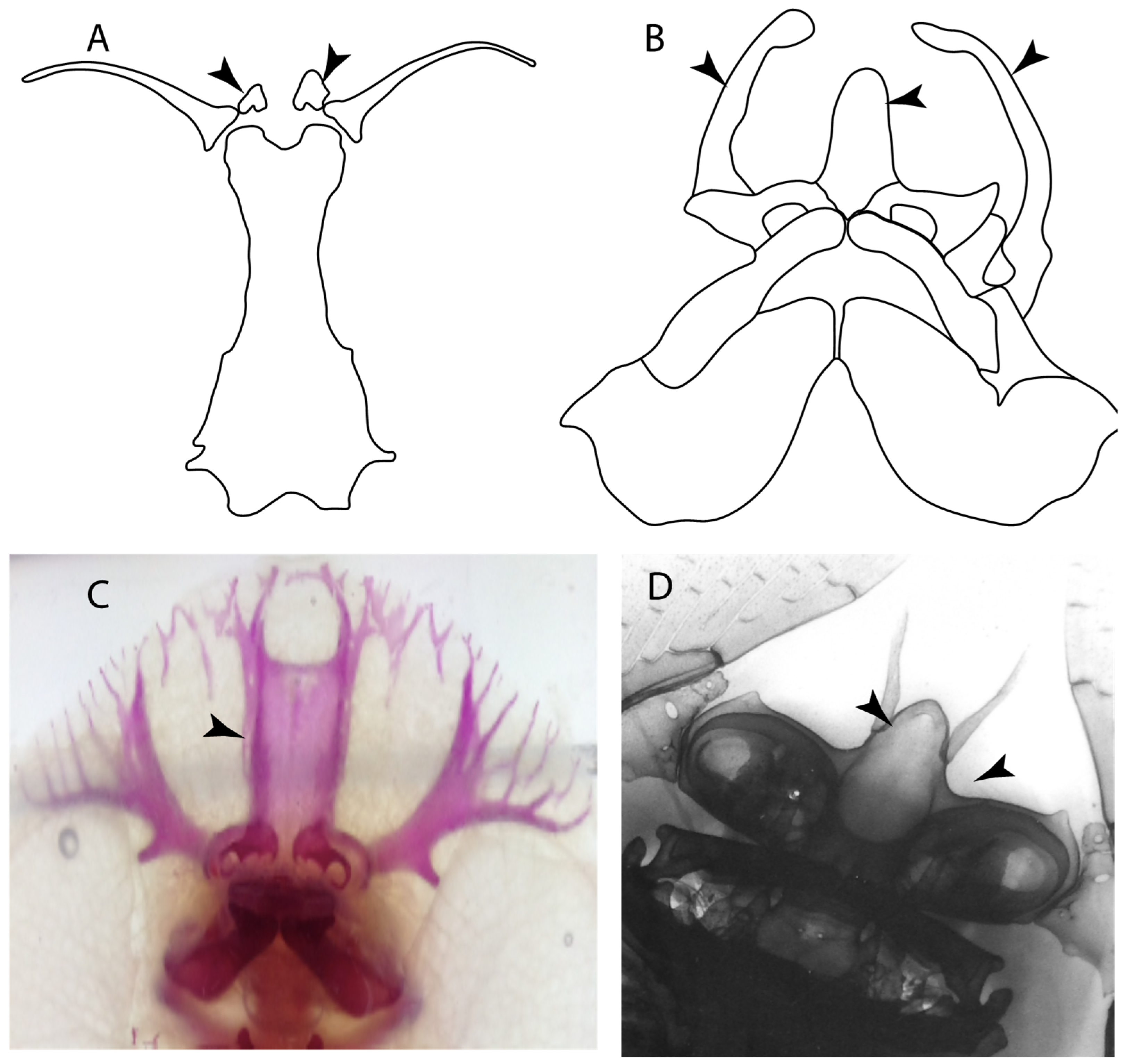
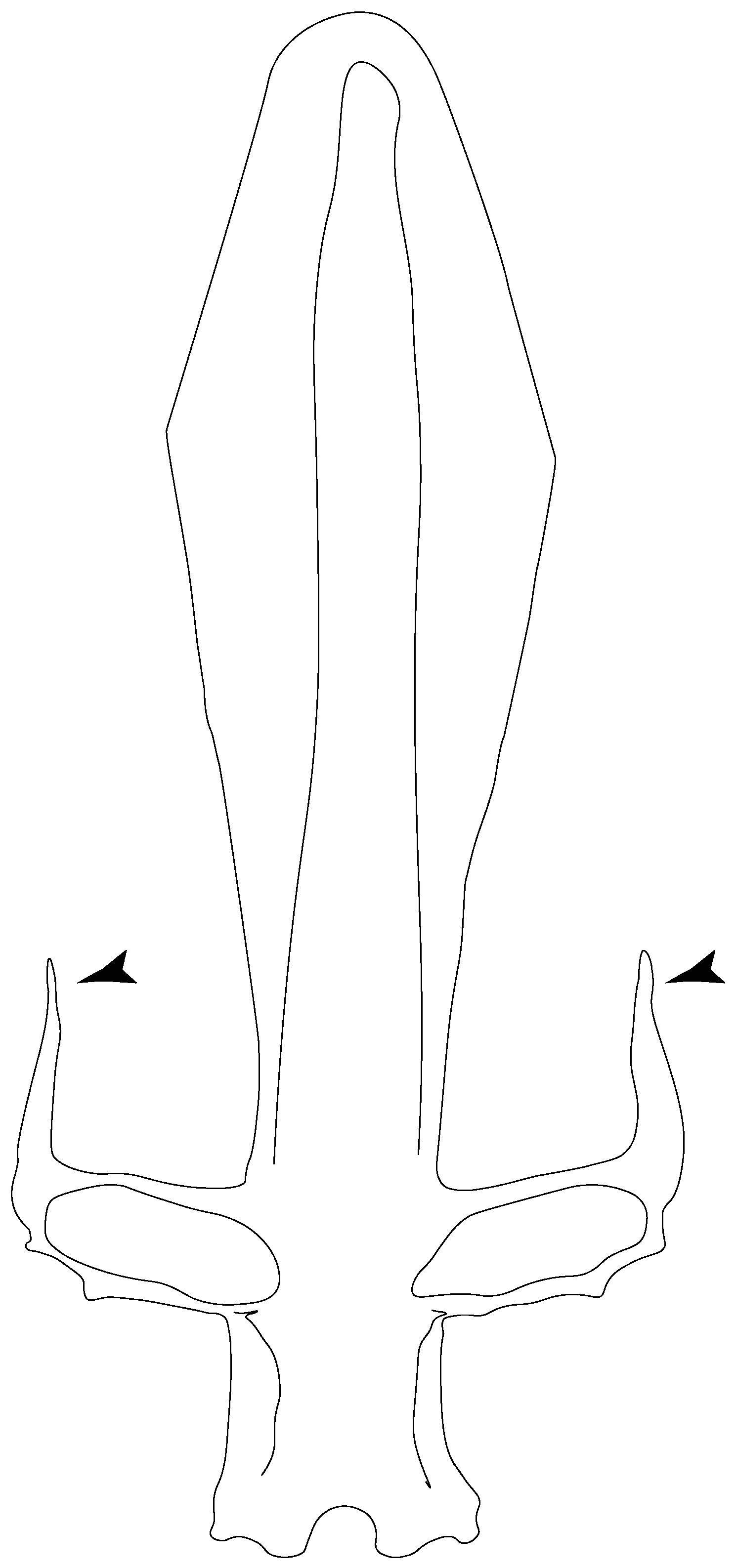
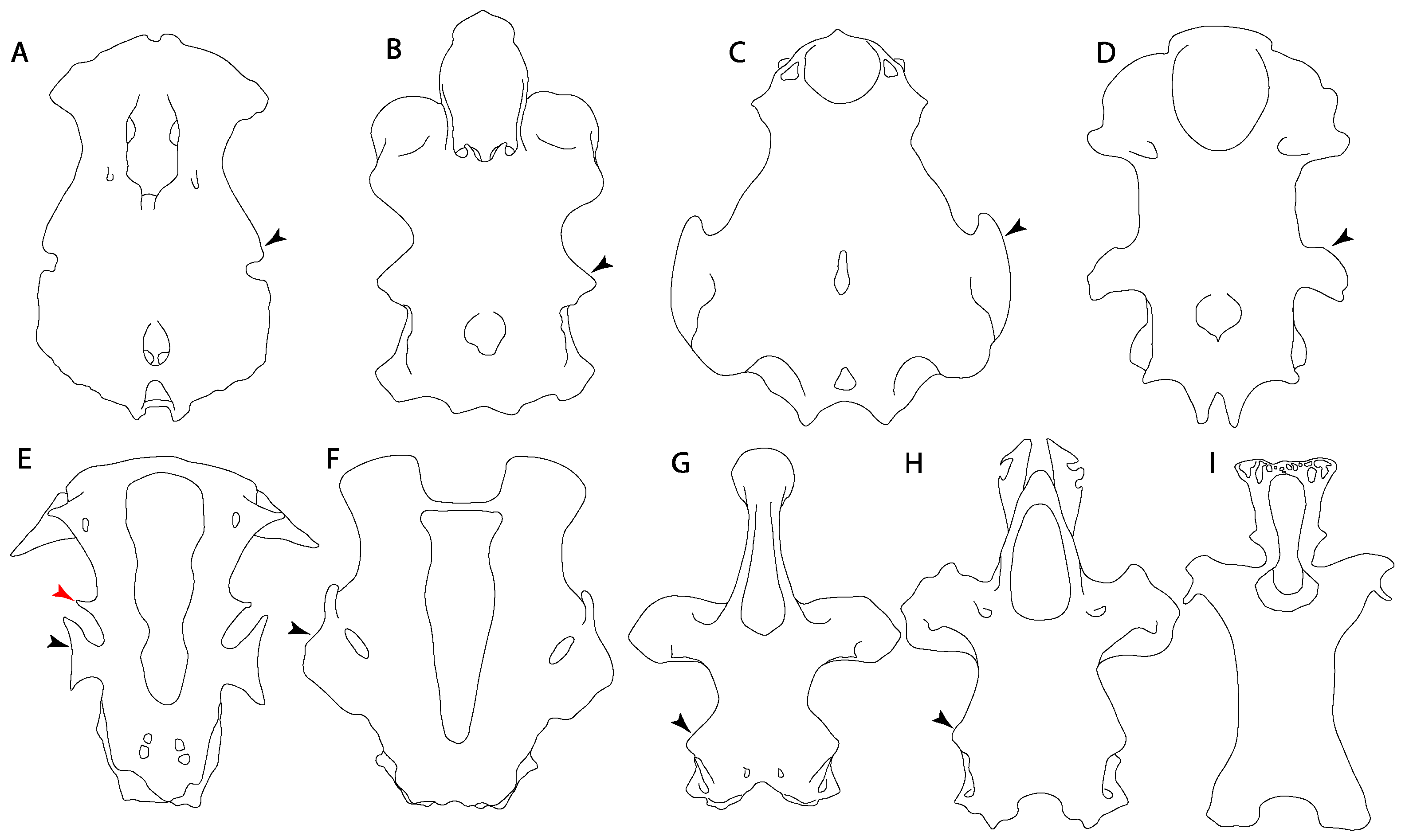
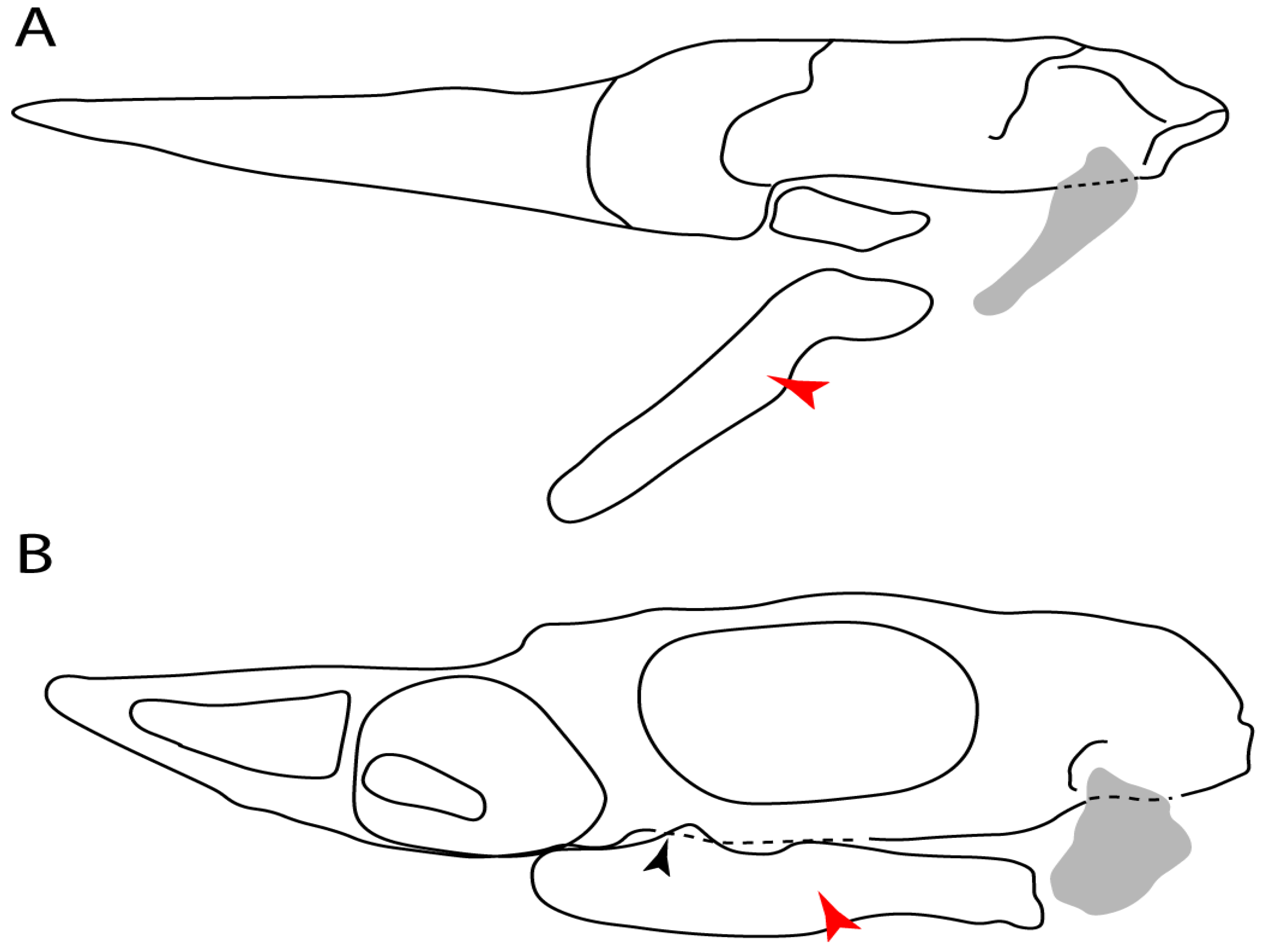
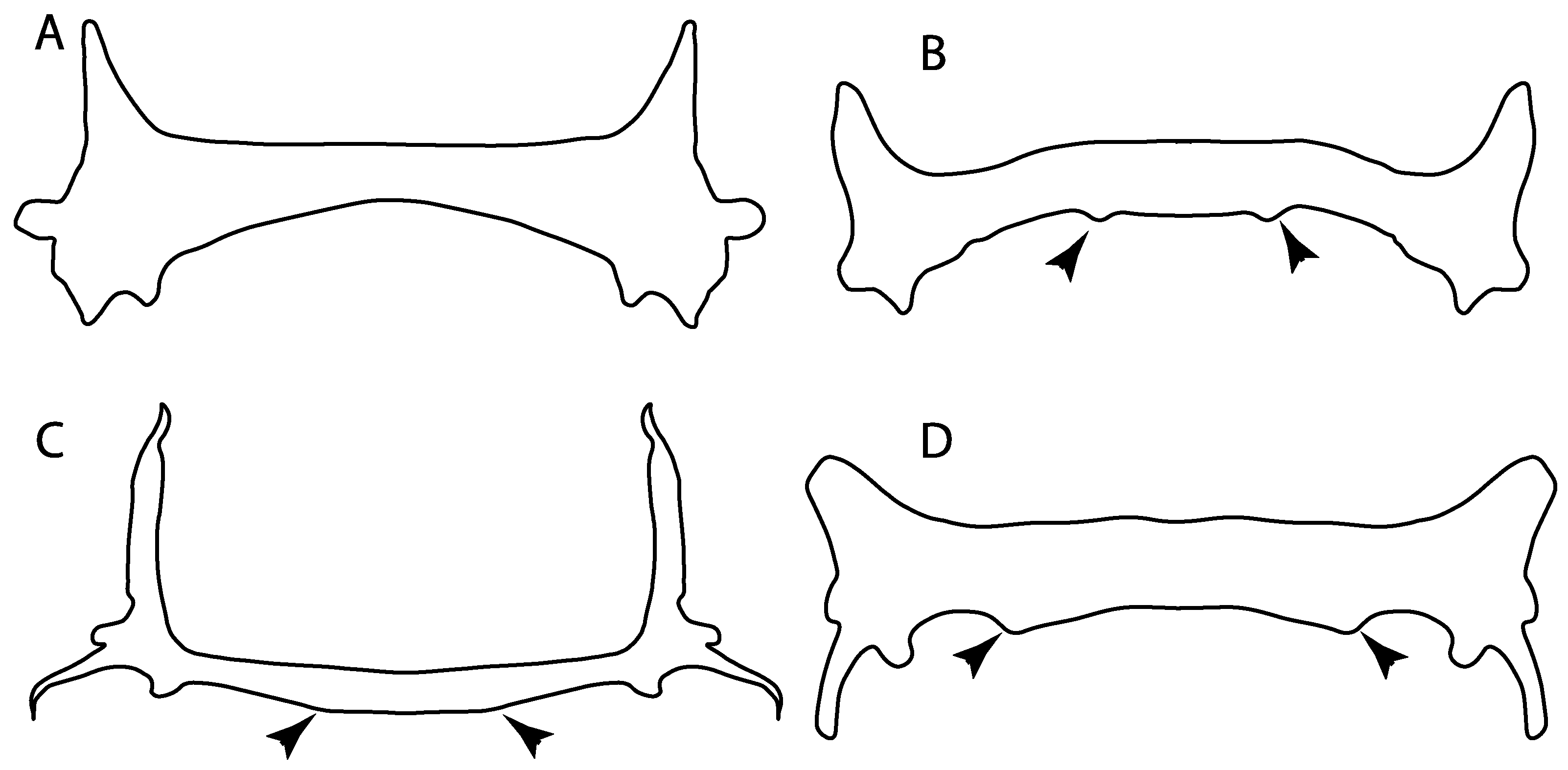
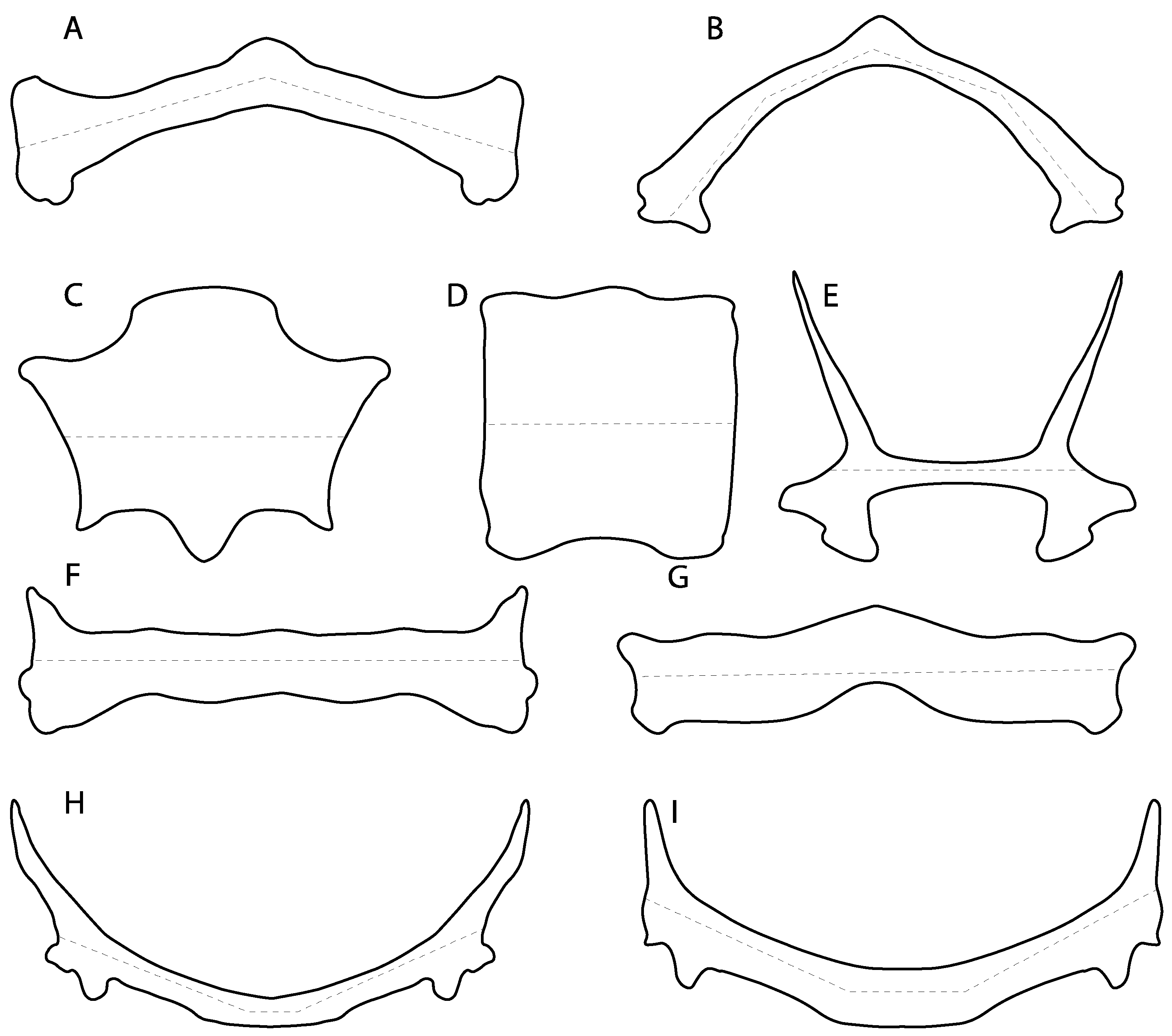
- Medial growth of rostral cartilage: (0) Inconspicuous; (1) conspicuous (noticeable). Modified from Aschliman et al. [7] (char. 26), Villalobos-Segura et al. [32] (char. 27) and Claeson et al. [23] (char. 2). De Beer and Moy-Thomas [55] named the cranial projections observed in chimaeroids (Figure 6A–C) as “medial and lateral rostral processes”. This nomenclature is kept by Claeson [60] and used to recognize the structures on the rostral cartilages of electric rays. However, the topological homology of these structures is unclear. Because of this, we coded the medial growth of rostral cartilage in chimaeroids as inapplicable (-). The coding of this character for Ginglymostoma follows Motta and Wilga Text-Figures 8 and 9 in [61]. The character state is inconspicuous (0) for both Torpedo and Hypnos (Figure 6E,F); in Hypnos, the antorbital cartilages and nasal capsules support the anterior extension of the pectoral disc, and in Torpedo, the rostral cartilages seem to arise primary from the anterior wall of the nasal capsules and interact with the antorbital cartilages. In Narke, there are tripodal rostral cartilages with lateral and medial growths. Narcine also shows a noticeable development of its rostral cartilages. We consider the observations by Miyake [62] in Urolophus, Urotrygon and Potamotrygon to be correct, as these taxa present evident vestiges of the rostral cartilages, such as those in Gymnura Text-Figure 1 in [63], Text-Figure 1 in [64]. Aschliman et al.’s [7] coding for Urobatis was kept, as we were unable to confirm the observations of Miyake et al. [56] and McEachran et al. [6] on the presence of these vestiges. Rhinoptera and Mobula (Figure 6F) present a lateral growth on the trabecula and lamina orbitonasalis to support the cephalic fins [56]. Zanobatus and †Plesiozanobatus show a small medial growth of the rostrum (Figure 6D). De Carvalho et al. [65] noticed in specimens of †Asterotrygon (NHMUK P 61244; PF 15180) the rostral projections and medial growth. The rostral cartilages in †Heliobatis, Plesiobatis, Hexatrygon, Hypanus, Potamotrygon, Neotrygon, Aetobatus, Myliobatis, †Lessiniabatis, †Tethytrygon and †Promyliobatis are inconspicuous. The rostral cartilages in the most rajoids (not in Sympterygia), and in all sclerorhynchoids, extant and extinct platyrhinids and Rhinopristiformes, are noticeable.
- 6.
- Different cartilage structures on rostrum (highly porous peripherical and fibrous at the central portion): (0) Absent, (1) present (new).
- 7.
- Rostral processes: (0) Absent; (1) present. Modified from Aschliman et al. [7] (char. 29); Claeson et al. [23] (char. 12). The presence of hyaline and poorly calcified structures called “rostral processes”, are shared features of platyrhinids according to de Carvalho [67]. These structures are not considered homologous to the rostral appendices of Rajiformes and Rhinopristiformes [7]. The rostral appendix in skates and guitarfishes is formed de novo on the proximal sides of the growing rostral plate [68]. Meanwhile, according to Miyake et al. [56], the paired “rostral cartilages” that are equivalent to Holmgren’s [69] “rostral appendices”, develop during early ontogeny and arise from the ventromedial area of the lamina orbitonasalis (at least in Torpedo). The area of development of these structures in Torpedo is topologically similar to the areas where the rostral processes of platyrhinids are formed, and might indicate a homologous relationship between the “lateral rostral cartilages” of Baranes and Randal [70] and Claeson [60], which are present in Torpedo, Electrolux, Typhlonarke, Temera and Narke, and the “rostral processes” of Platyrhina and Platyrhinoidis of de Carvalho [67]. The presence of rostral processes is unknown (?) in some fossils recognized as sister taxa (i.e., †Britobatos) or belonging to Platyrhinidae (i.e., †Tingitanius; [23]), but rostral processes are present in the Eocene platyrhinid †Eoplatyrhina [33]. We kept holocephalians as unknown (?) as we still have doubts about the homology of the “process” recognized by de Beer and Moy-Thomas [55].
- 8.
- Rostral processes (proximal articulation): (0) Articulated with nasal capsules; (1) continuous with neurocranium; (2) articulated with ventral aspect of rostral cartilage. Based on Claeson [60] (Supporting Information char. 50) and Claeson [71] (char. 48). This character seeks to include the variation on the articulation between the rostral processes and the neurocranium following Marramà et al. [33].
- 9.
- Rostral appendix: (0) Absent; (1) present. Modified from Aschliman et al. [7] (char. 28). The presence of rostral appendices is considered a shared feature between rhinopristisforms and rajiforms [7]. McEachran et al. [72] and Claeson et al. [23] also recognized their presence in fossil taxa such as †“Rhinobatos”maronita, †“R.” latus, †Britobatos and †Rhombopterygia. The coding from previous works [23,59] for †Spathobatis was changed to “present” after new observations were made on several fossil specimens (e.g., BMNH P. 12067, BSPG 1952-I-82, BSPG -AS-I-505) (EV, pers. observ.). Although rostral appendices might be present in Diplobatis, Benthobatis, Narcine, Discopyge and †Titanonarke [28,69,73,74], their homology with rostral appendices of skates and guitarfishes is unclear, and we therefore coded the state as (0) in these taxa. Considering that the subtriangular rostral extremity reported and observed in Urolophus, Gymnura, Urotrygon, Plesiobatis and Potamotrygon is in connection with the anterior medial outgrowth of the trabecula [67] (one of the embryological cartilages that forming the rostral cartilages along with the lamina orbitonasalis), it is very likely that these vestigial cartilages correspond to the rostral node and rostral shaft (sensu McEachran and Compagno [75]) and that the rostral appendices are involved, considering the presence of small posterior projections parallel to the rostral shaft. However, considering the lack of agreement about the presence of this structure in the literature [56,67,72] we coded this character as unknown for these taxa (?).
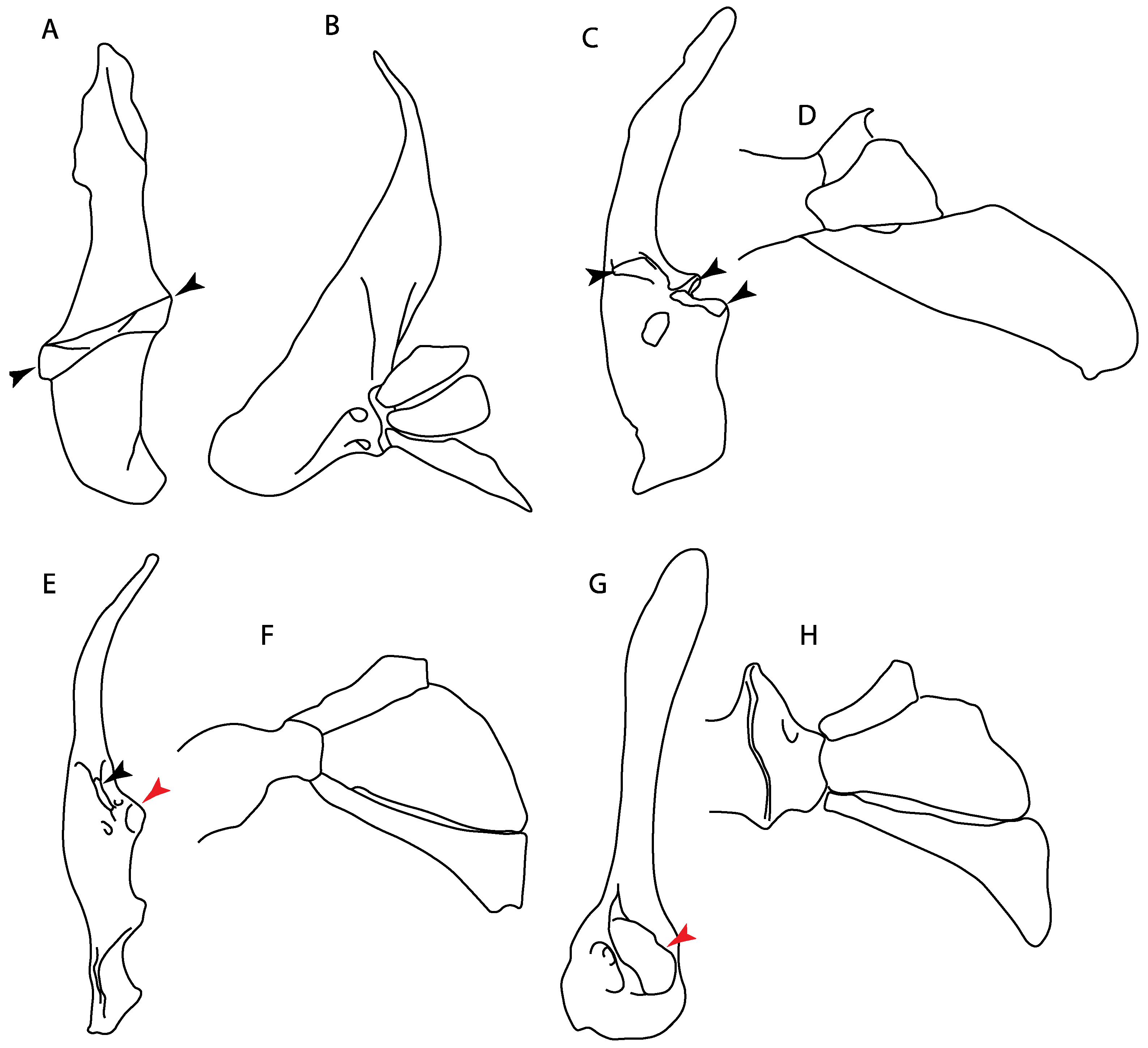
- 11.
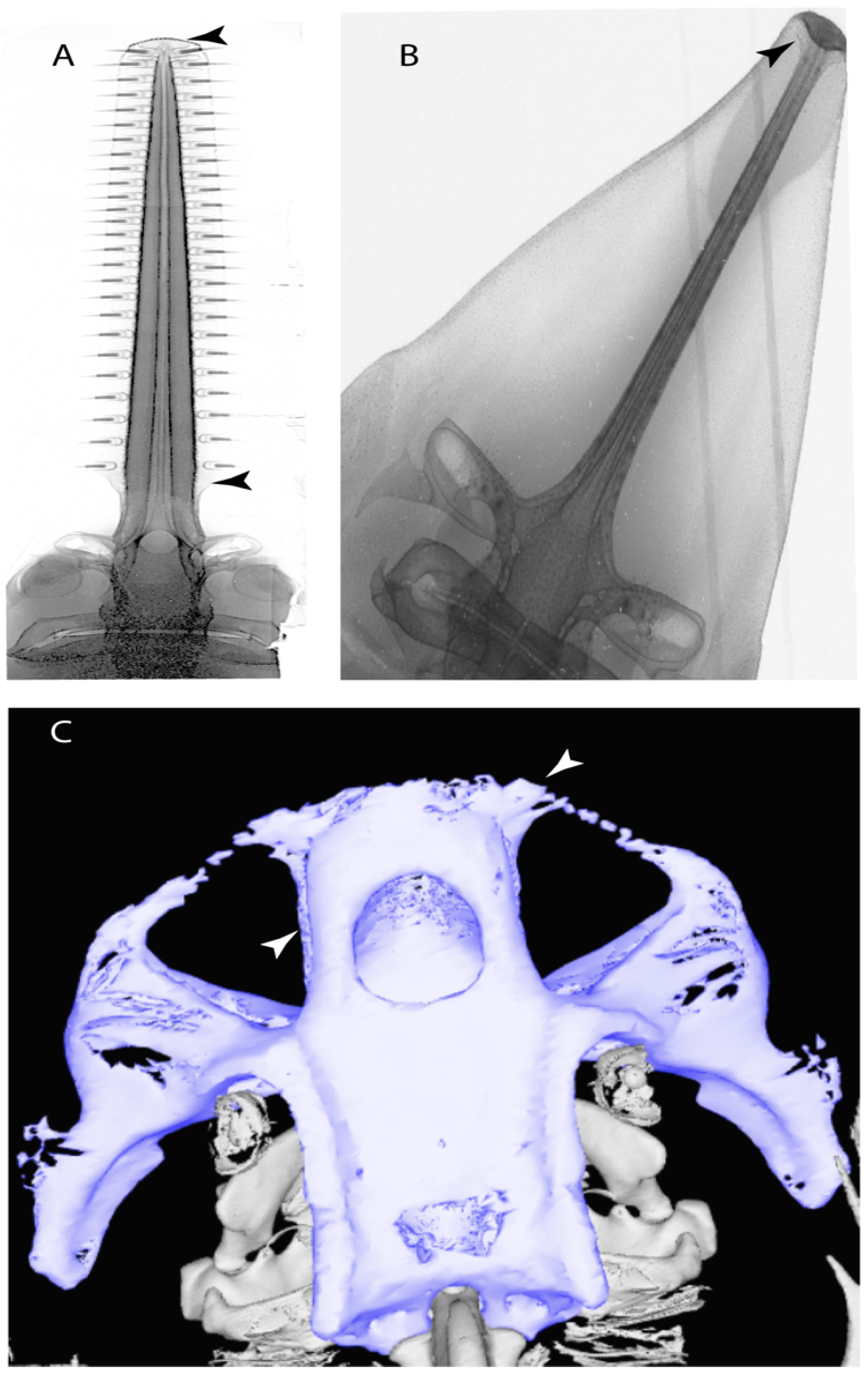
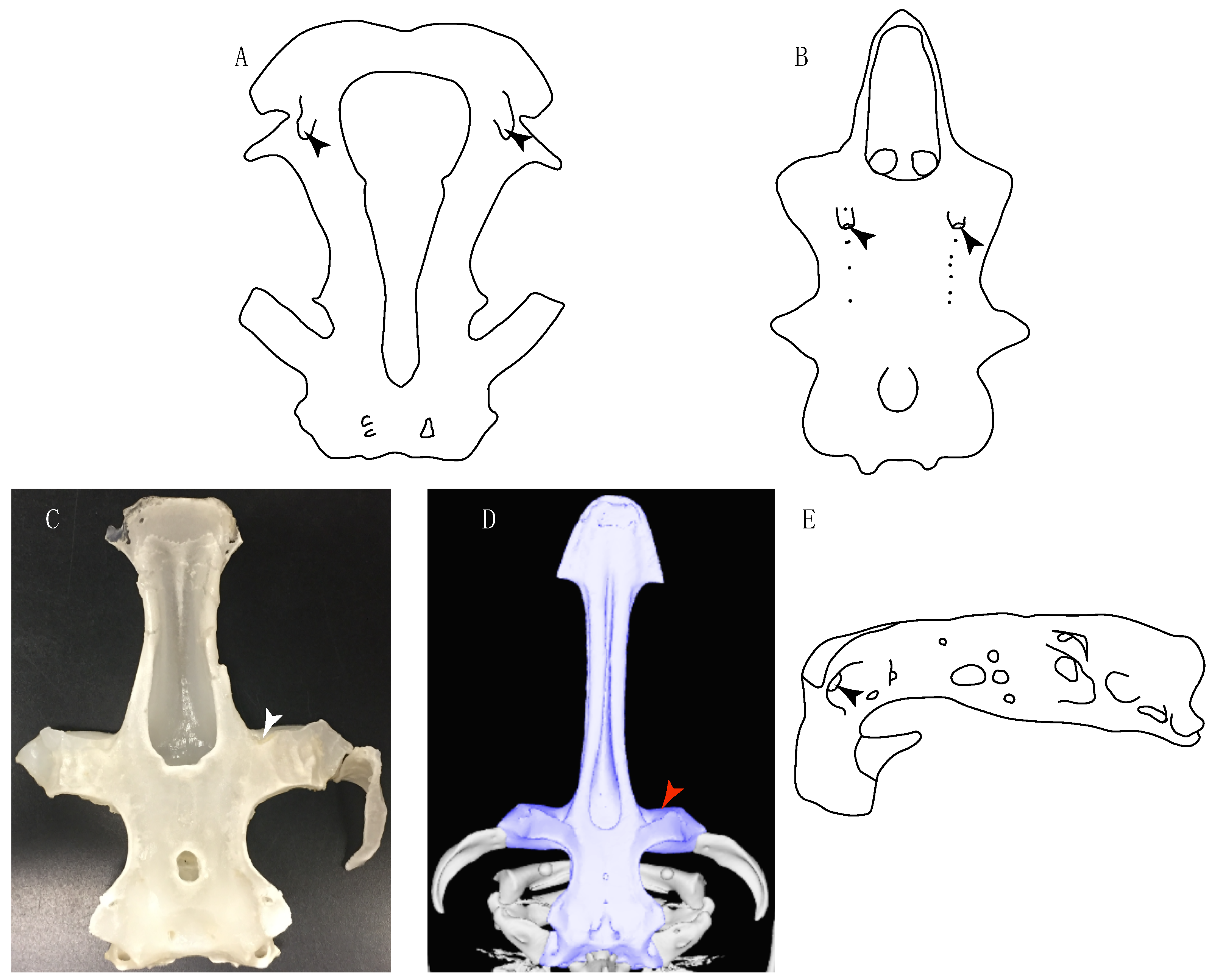
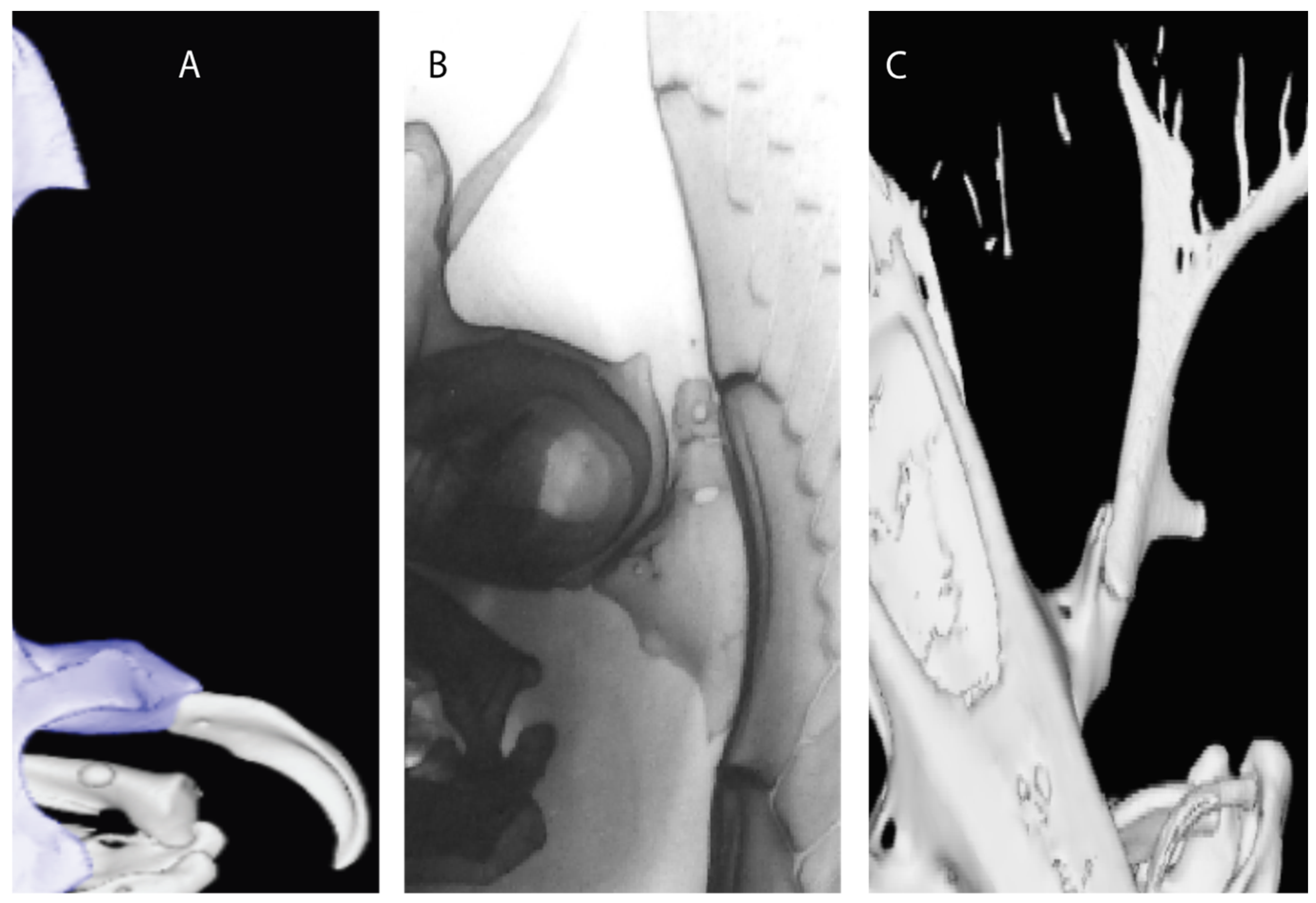
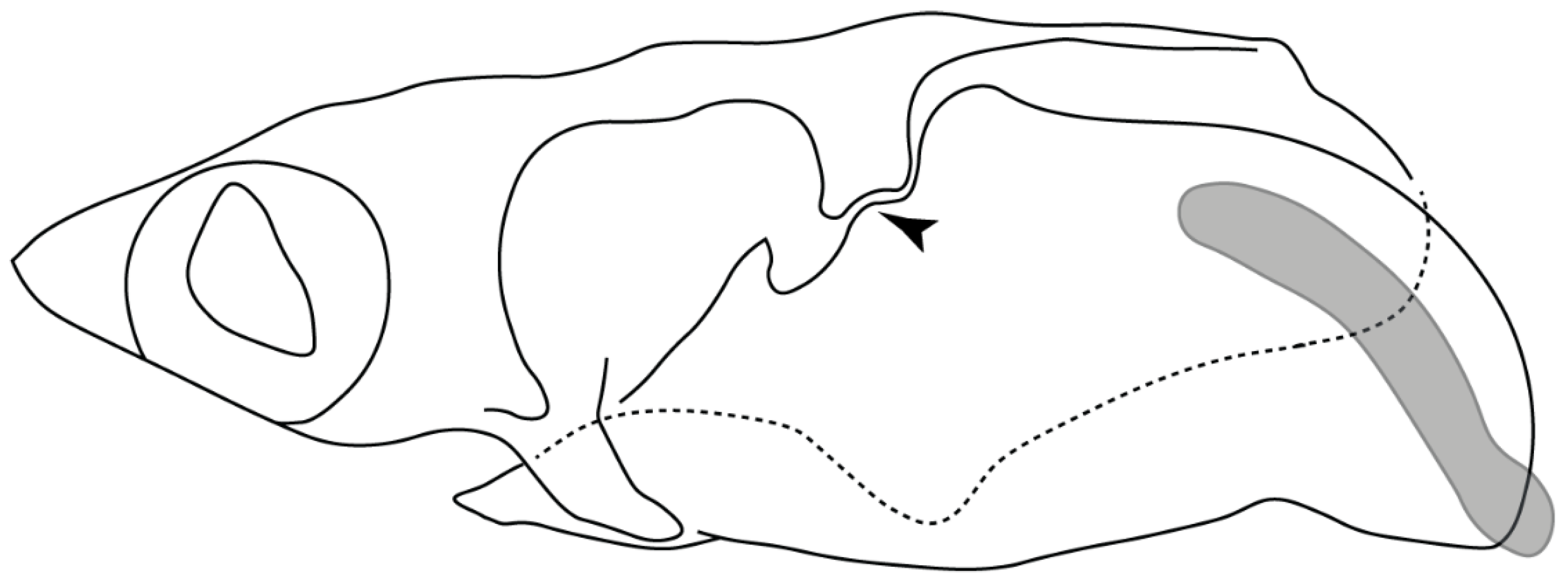
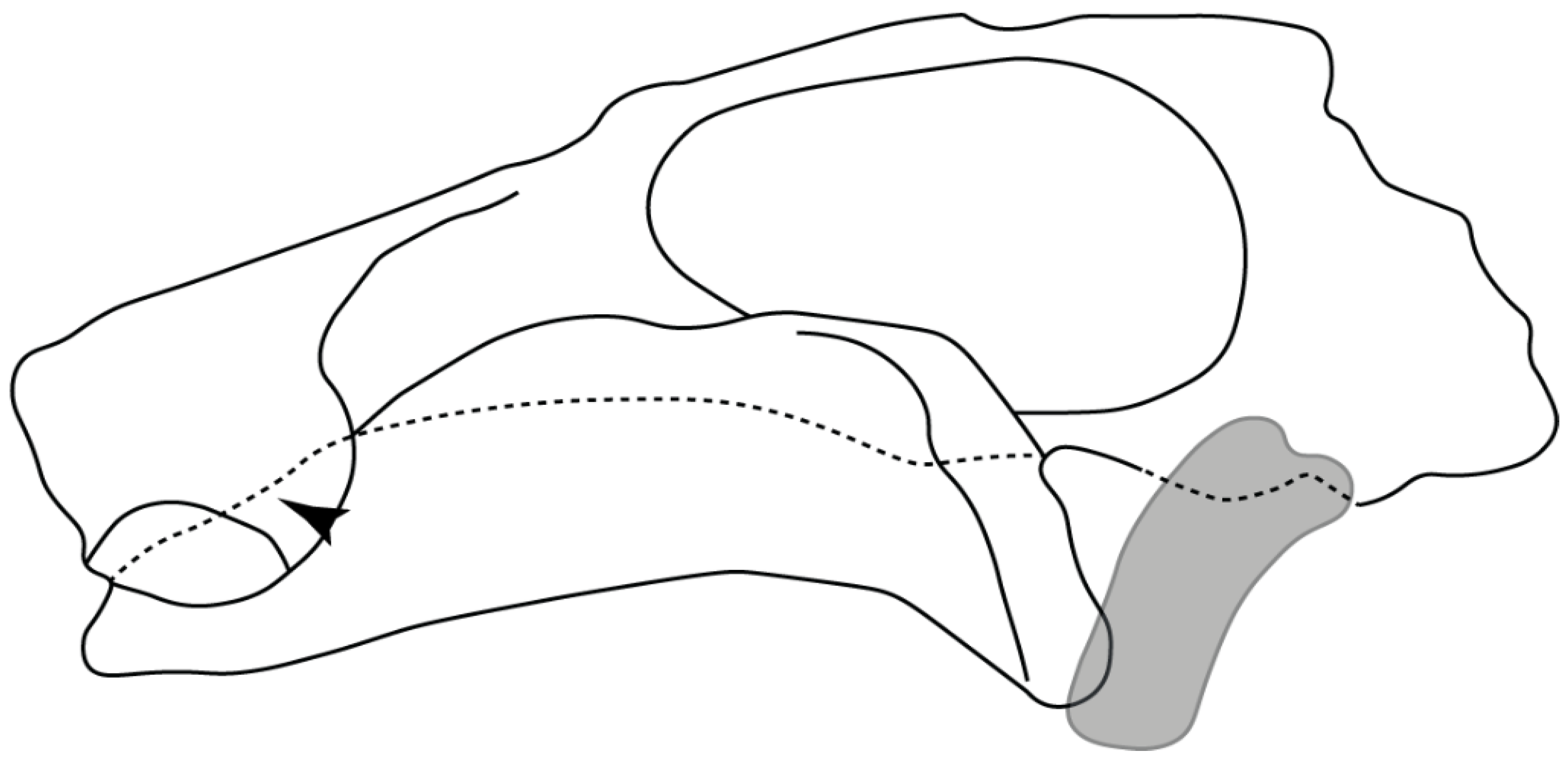
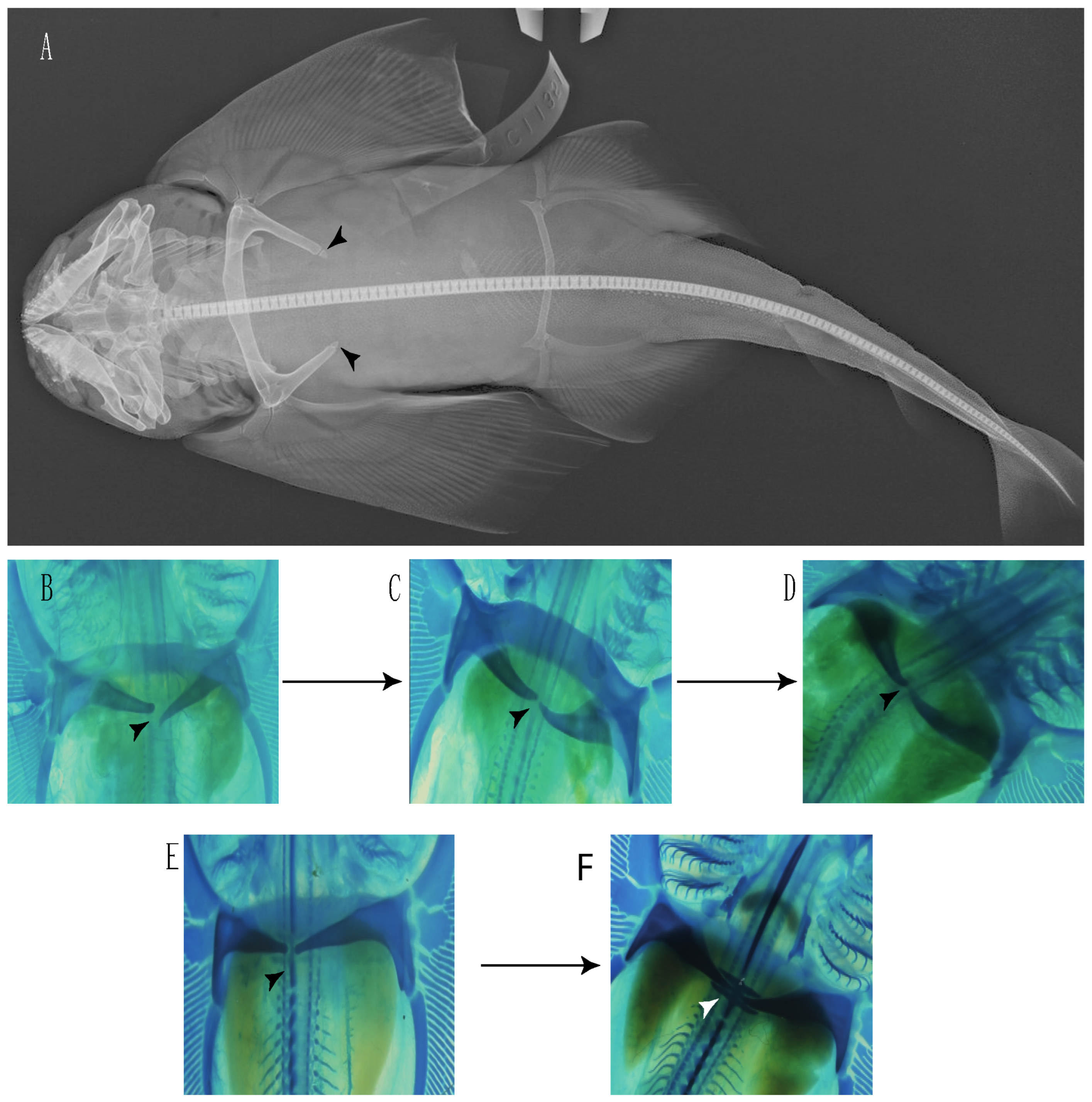
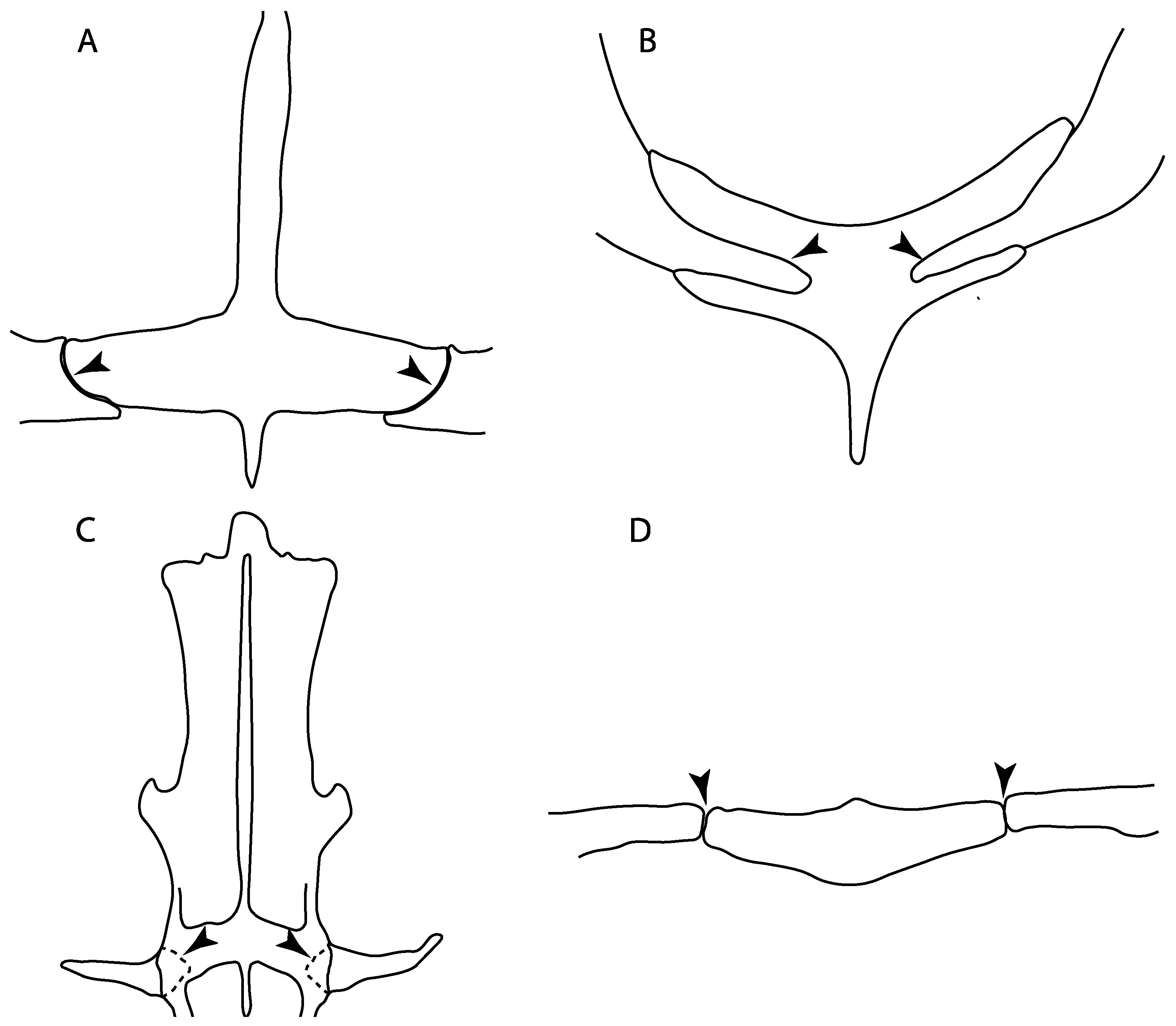
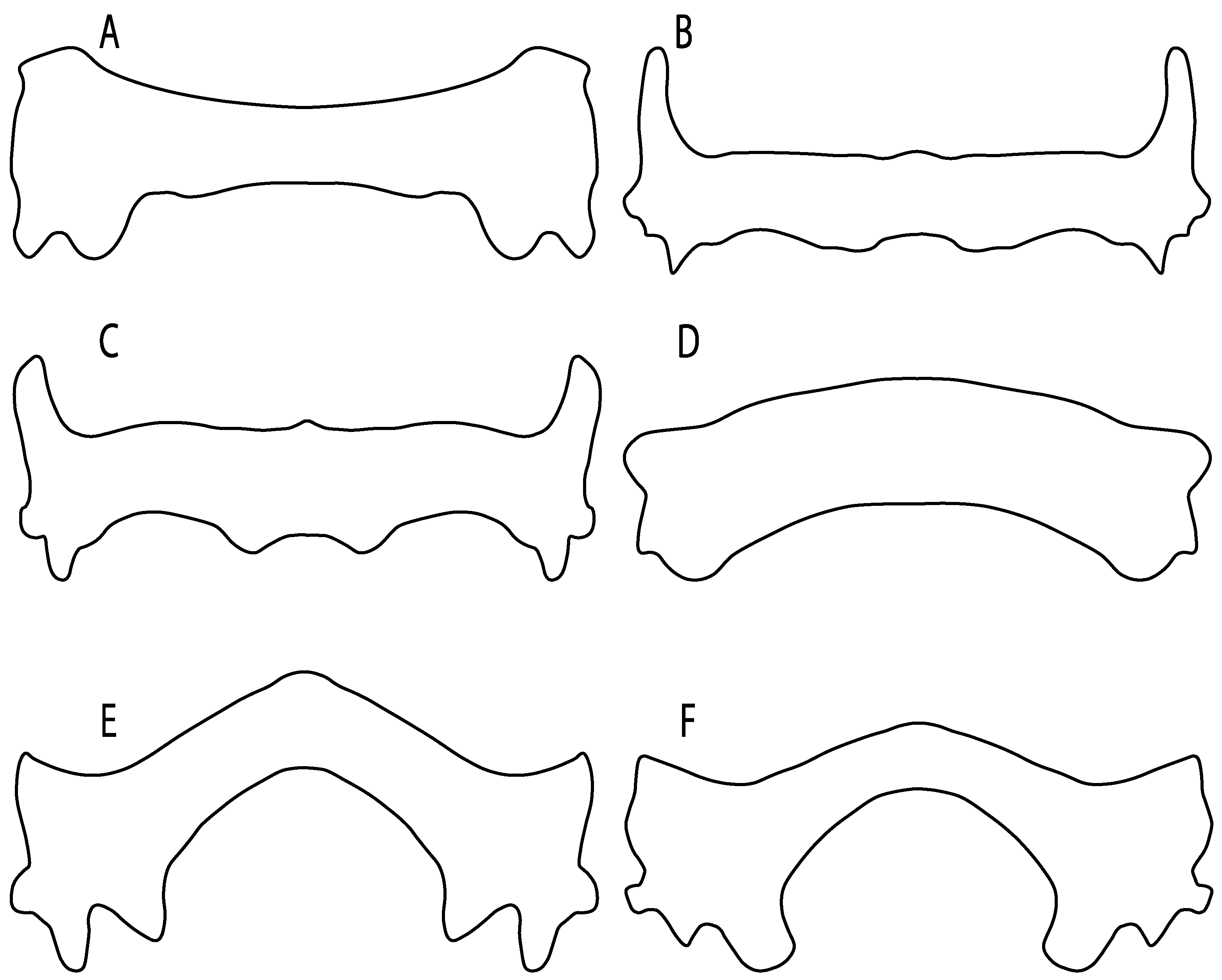
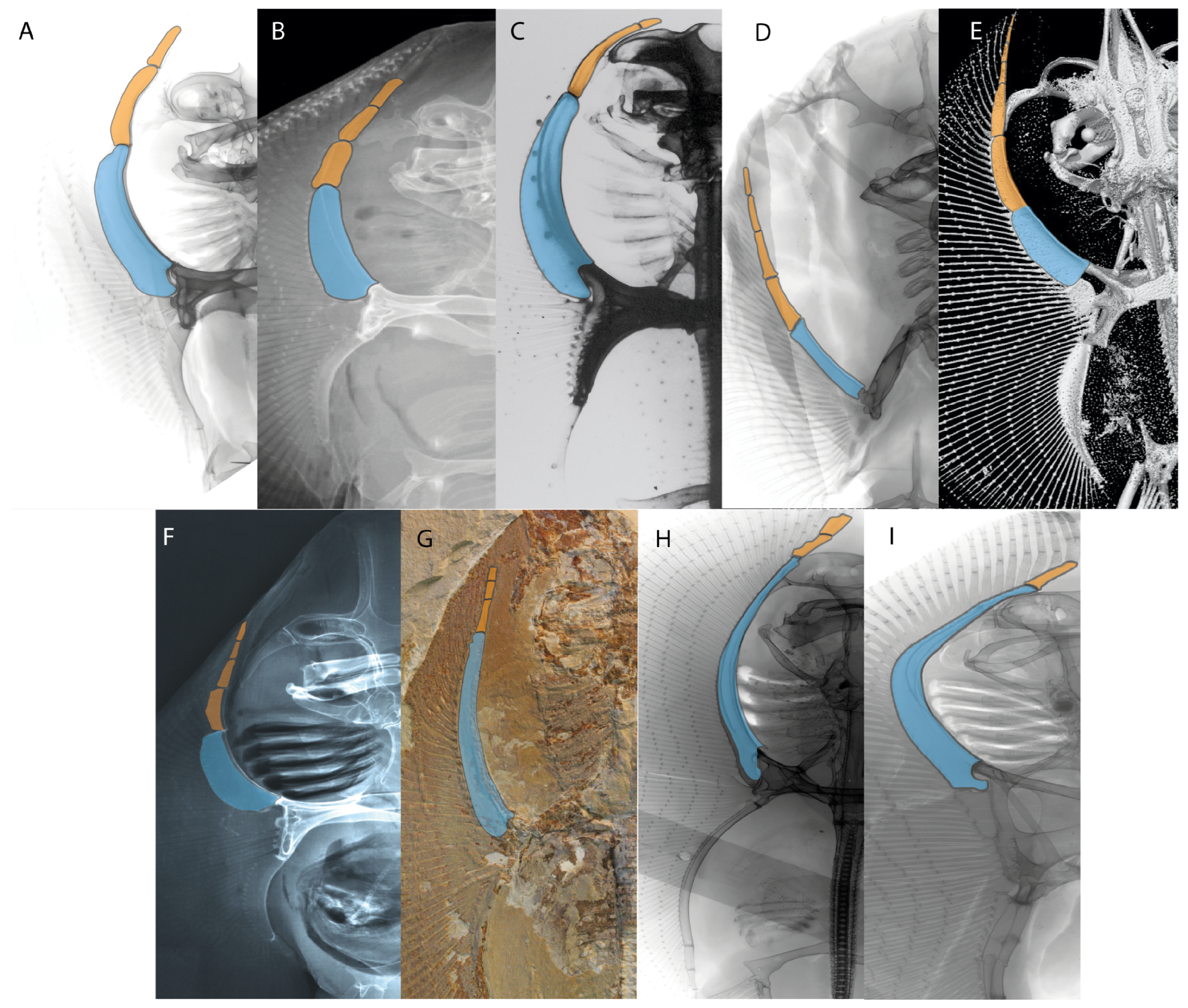
- Rostral passage of superficial ophthalmic nerve: (0) Covered; (1) open. Based on Wueringer et al. [76] and Cappetta [77]. In Chimaera and Harriotta, the ramus of the superficial ophthalmic nerves runs across the anterior and posterior opening of the ethmoidal canal and is covered by the lamina orbitonasalis anteriorly and by the orbital cartilage posteriorly [55]. Coding for Chlamydoselachus follows Allis’s [78] observations, coding in Torpedo follows Ewart’s [79] observations. Rhinobatos, Aptychotrema and Pristis were used to illustrate the state found in most batomorphs [76] (Figure 10A–C). In the rostral cartilage of sclerorhynchoids, the supraophthalmic nerve canal is open, like in Pristiophorus (Figure 10D–G) [76,77].
- 12.
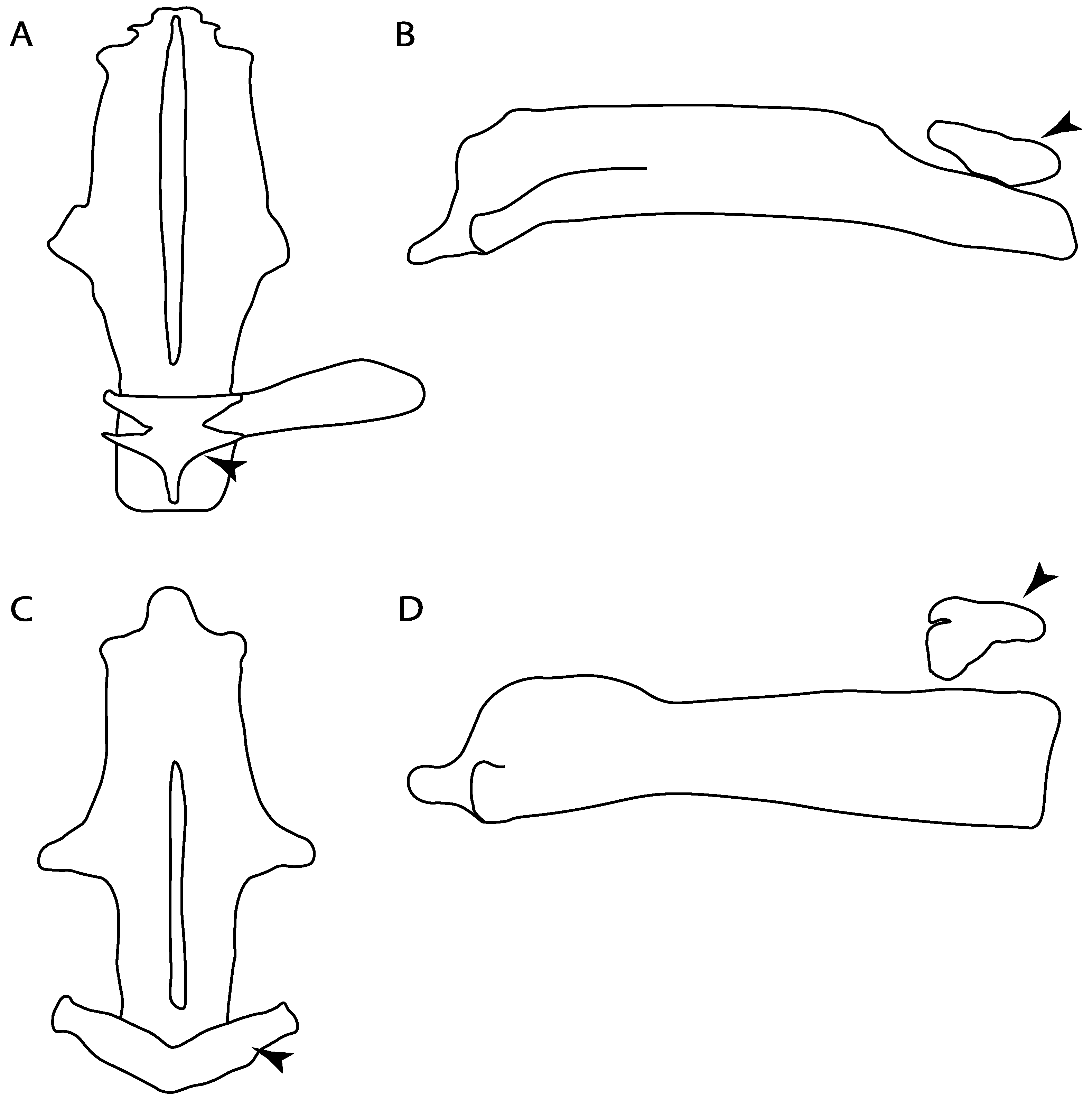
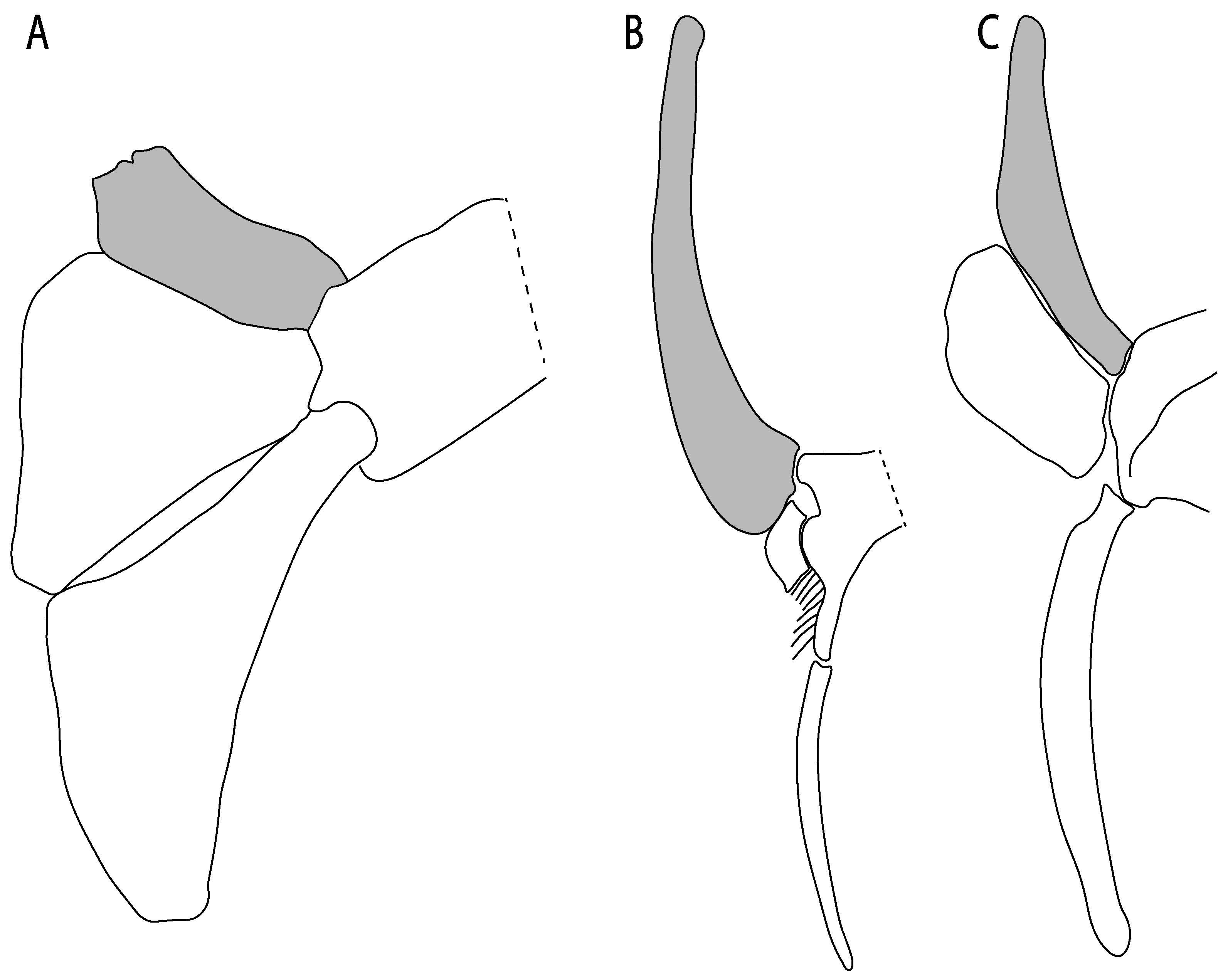
- Anterior preorbital foramen: (0) Dorsally located; (1) anteriorly located. Modified from Aschliman et al. [7] (char. 35); Villalobos-Segura et al. [32] (char. 37). We modified the previous coding for Rhinopristiformes, as the foramina in †Spathobatis, †Stahlraja, †Tlalocbatus, Pristis, Rhynchobatus, Glaucostegus, †“Rhinobatos” latus, Rhina, Rhinobatos, Pseudobatos, †Eorhinobatos, †Pseudorhinobatos, †Iansan, Trygonorrhina, Zapteryx and Aptychotrema are located near the base of the rostral cartilage, but more anteriorly directed in Myliobatis, Aetobatus, Rhinoptera and Mobula (Figure 11C–E). Aschliman et al. [7] coding was kept, except for Chimaera, Harriotta, Temera and Torpedo, as we could not observe the foramen (?).
- 13.
- Preorbital process: (0) Present; (1) absent. Modified from Aschliman et al. [7] (char. 33), based on a new reinterpretation of the specimens in the literature [24] and a re-examination of the specimens (BSP AS 1952-I-82 and AS-I-505), the coding of Villalobos-Segura et al. [32] for †Kimmerobatis and †Spathobatis was changed from absent (1) to present (0).
- 34.
- Nasal capsules: (0) Laterally expanded; (1) ventrolaterally expanded; (2) anterolaterally expanded; (3) prolonged interorbitonasal region, which forms a pedicel (“trumpet shaped nasal capsule”). Aschliman et al. [7] (char. 31) was modified, adding Maisey et al.’s [83] (char. 4) observations on the nasal capsules in Squatina and †Pseudorhina (2) and Compagno’s [7] and Shirai’s [35,37] characterization of these structures in the orectolobids and heterodontids (3).
- 35.
- Nasal capsule margin: (0) Straight; (1) horn-like process. Based on Villalobos-Segura et al. [32] (char. 83).
- 110.
- Position of the articulation of the antorbital cartilage on nasal capsule: (0) Lateral, (1) anterolateral; (2) posterolateral. Modified from de Carvalho [67] (char. 2).
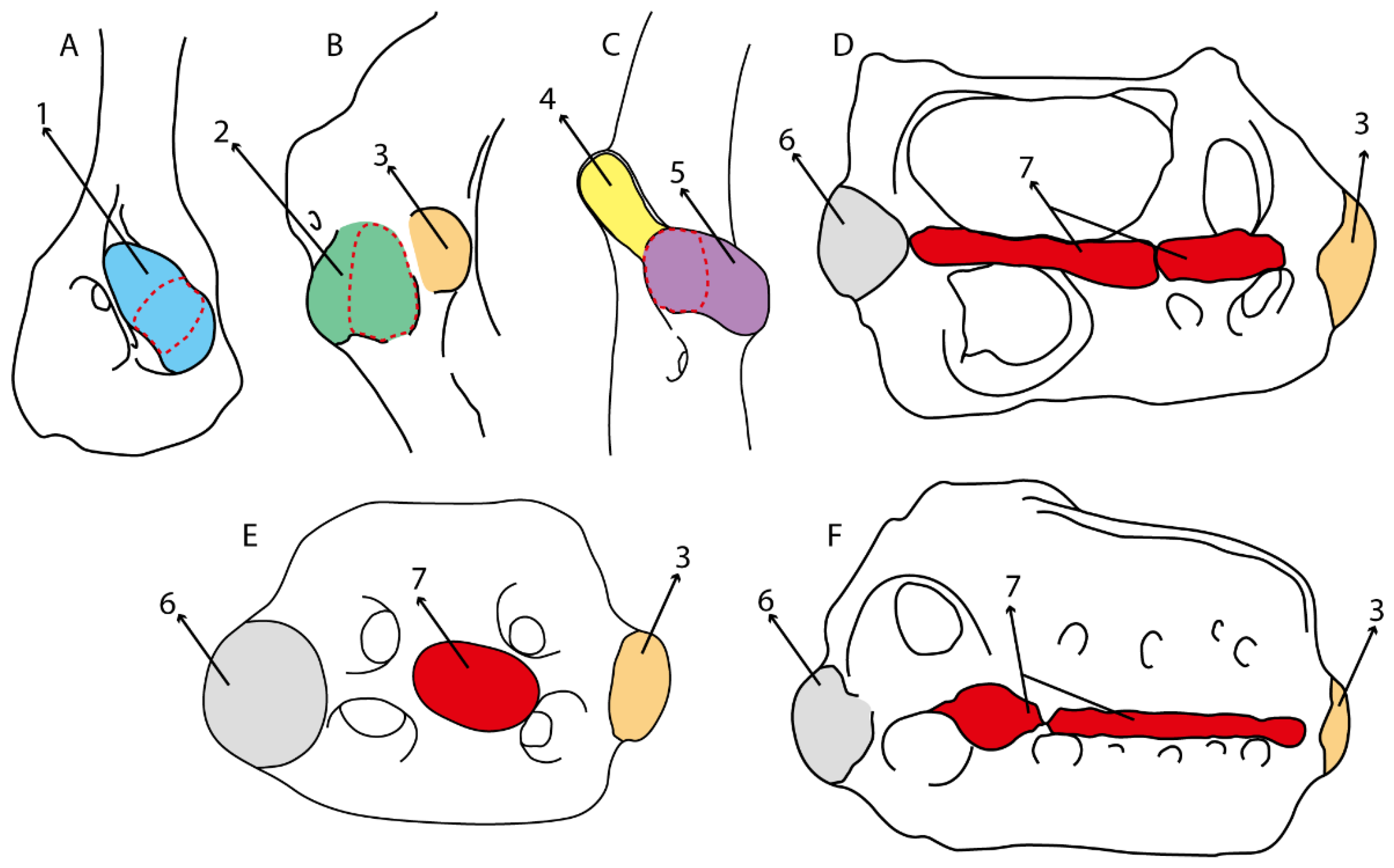
- 23.
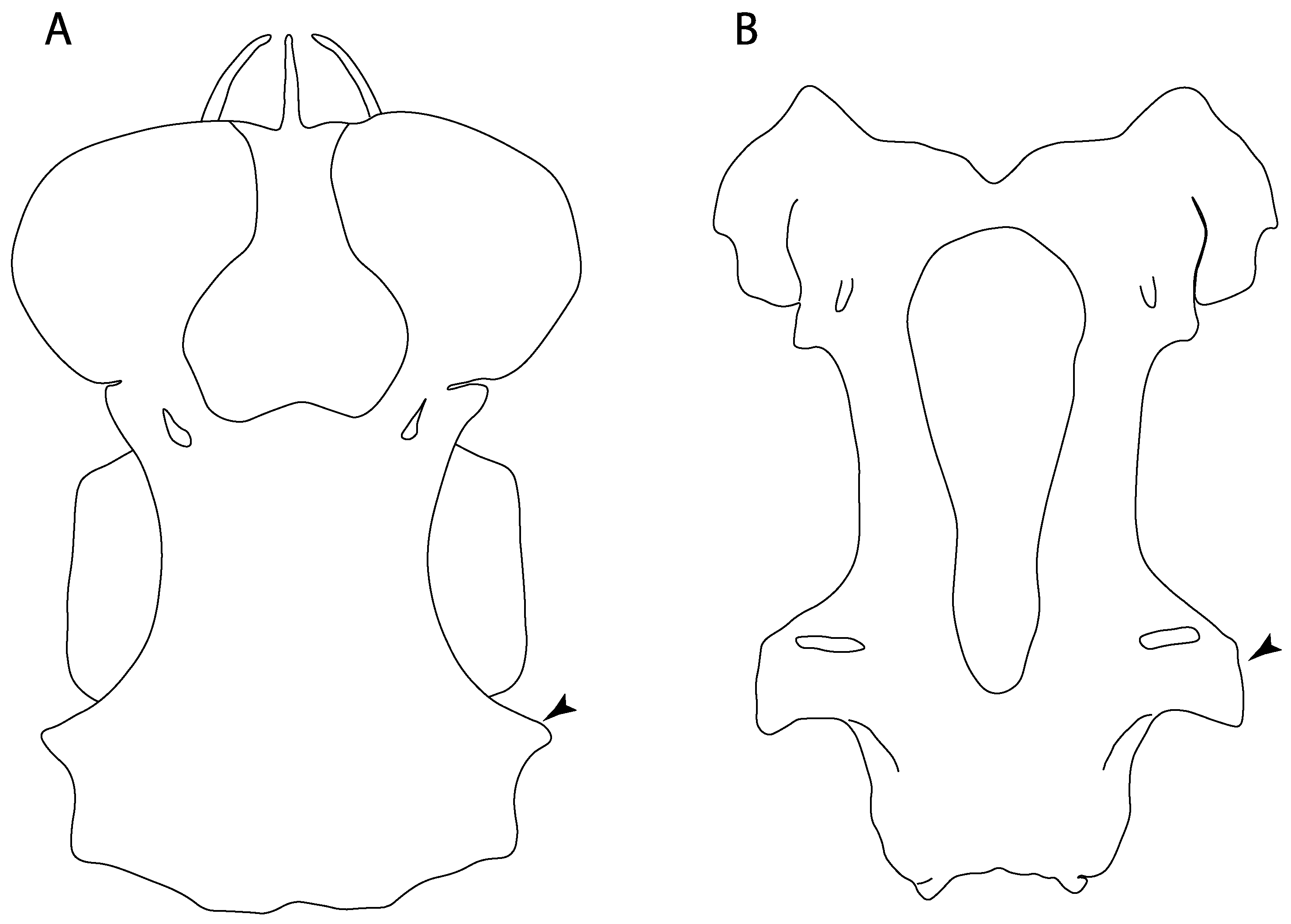
- Antorbital cartilages: (0) Absent; (1) present. Modified from Aschliman et al. [7] (char. 8). Shirai [35] suggests the presence of antorbital cartilages in Pristiophorus. However, no evidence of these cartilages was observed in the Pristiophorus specimens CSIRO 3731 and CAS 4942. The previously illustrated lack of antorbital cartilages in †Cyclobatis in Cappetta Text-Figure 355A in [85] seems to be caused by the position of these cartilages, which appear to be overlapped by the propterygium, like in stingrays (Figure 15).
- 24.
- Antorbital cartilage (shape): (0) Triangular-shaped with regular outline; (1) variously shaped and with an irregular outline. Modified from Villalobos-Segura et al. [32] (char. 9), based on observations on †Titanonarke, Narcine, Narke, Temera, †Eoplatyrhina, Platyrhinoidis and †Tingitanius, which present antorbital cartilages with an irregular outline and various projections. Due to preservation or damage, †Kimmerobatis, †Promyliobatis and †Ischyrhiza are coded as (?).
- 25.
- Antorbital cartilages (with regular outline): (0) Well-developed; (1) reduced. Modified from Villalobos-Segura et al. [32] (char. 9), split in two characters (25 and 26). This character includes the variation observed in the size of the antorbital cartilages with regular outlines in batomorphs. Taxa with irregular outlines of antorbital cartilages (i.e., †Titanonarke, Narcine, Narke, Temera, †Eoplatyrhina, Platyrhinoidis and †Tingitanius) and taxa lacking antorbital cartilages (i.e., holocephalians and sharks), were coded as inapplicable (-). Due to taphonomic loss or damage in †Kimmerobatis, †Promyliobatis and †Ischyrhiza, this character is unknown (?).
- 26.
- Anterior process of antorbital cartilage (if regular outline): (0) Absent; (1) present. This character includes the variation observed in the anterior portion of the antorbital cartilage of batomorphs.
- 27.
- Postorbital process: (0) Well-developed; (1) reduced. Based on Claeson et al. [23] (char. 12). This process cannot be observed in †Rhombopterygia, †Ischyrhiza and †Lessiniabatis (?).
- 28.
- Postorbital process: (0) Narrow; (1) broad and shelf-like. Taken from Aschliman et al. [7], (char. 36). †Cyclobatis presents a very narrow and laterally projected postorbital process.
- 40.
- Suborbital shelf: (0) Absent; (1) present. Based on observations by Shirai [35,37] and Klug [20]. The suborbital shelf is a horizontal plate on the ventral junction of the orbital wall and basal plate that is the floor of the orbit. It runs from the nasal capsule to the otic capsule and is penetrated posteriorly by the stapedial foramen and sometimes laterally by a notch, foramen, or fenestra for the palatine branch of the facial nerve.
- 41.
- Basitrabecular process: (0) Absent, (1) present. Based on de Carvalho [38] (char. 11), de Carvalho and Maisey [15] (char. 21) and Klug [20] (char. 10). This character is interpreted as a separate feature from the suborbital shelf based on its topographic relationships and development [90]. The basitrabecular process derives from a lateral expansion of the polar cartilage just anterior to the auditory capsules and articulates anteriorly with the orbital process of the palatoquadrate [15,90,91].
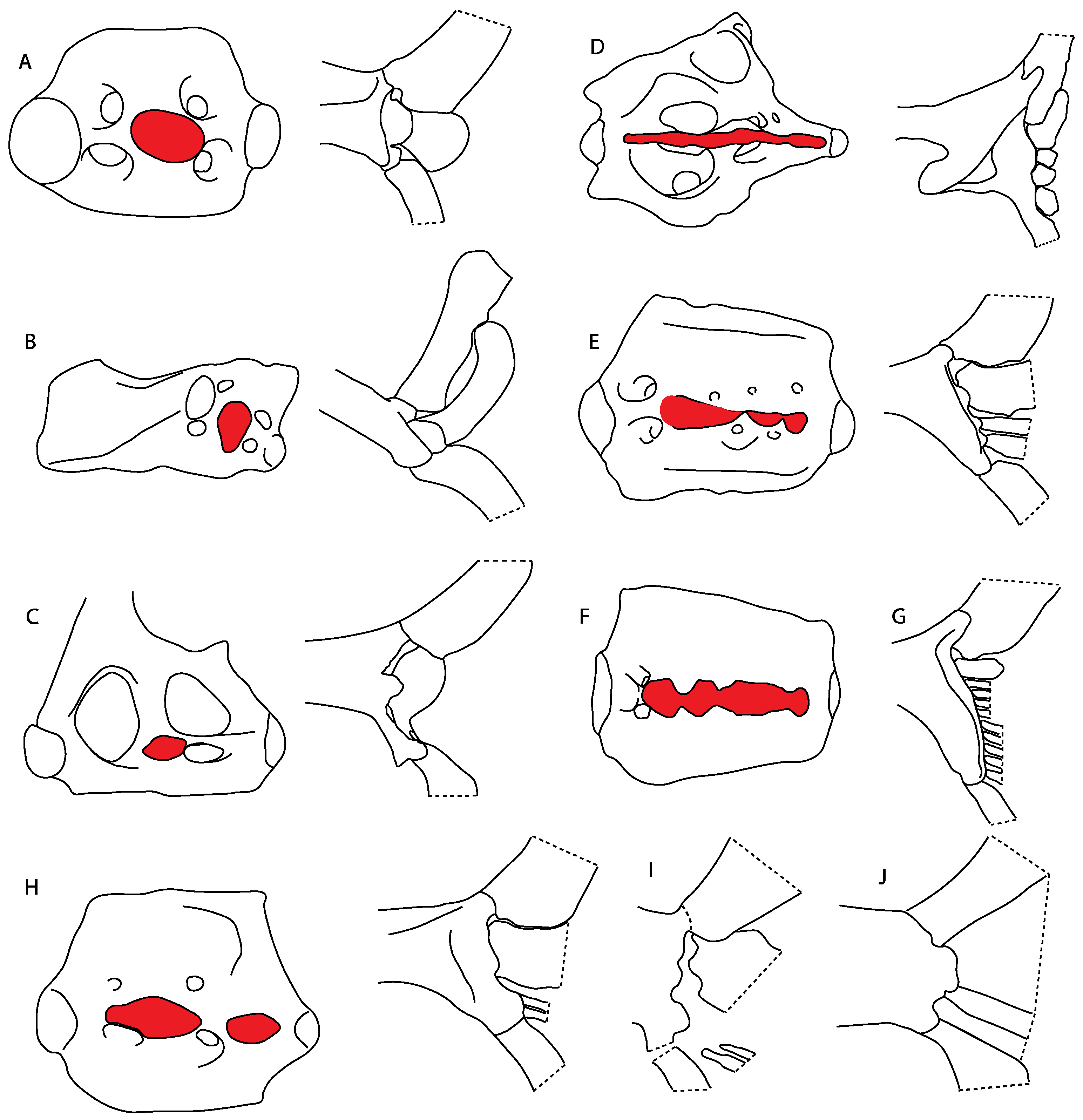
3.1.4. Viceral Arches
- 14.
- Jaw support: (0) Holostyly; (1) hyostyly; (2) archaeostylic. Based on observations by Maisey [92] and Wilga and Ferry [93]. In holocephalians, the neurocranium and the palatoquadrate are fused (i.e., holostylic jaw suspension). The term archaeostylic (sensu Maisey [94]) refers to those taxa with a postorbital articulation on the ventrolateral part of the lateral commissure. Both †Cobelodus and †Ozarcus lack a distinct hyomandibular facet [18,94], indicating a loose attachment to the neurocranium. Hybodontiforms and elasmobranchs present various articulation patterns between the upper jaw (palatoquadrate) and the neurocranium, but all share a close interaction with the hyomandibula (i.e., hyostylic) [87].
- 15.
- 17.
- 18.
- 19.
- Quadrate flange: (0) Absent; (1) present. Modified from Maisey et al. [83] (char. 6). The flange on the palatoquadrate is a characteristic feature in hybodontiforms (Figure 24A). This ledge is located laterally to the mandibular cartilage and does not interact with the Meckel’s cartilage. This process corresponds to the “quadrate process” of de Carvalho et al. [97] and Maisey et al. [83] in squatinids and pristiophorids (Figure 24B,C).
- 44.
- Basihyal: (0) Present; (1) absent. Modified from Aschliman et al. [40] (char. 48), Villalobos-Segura et al. [32] (char. 48) and Claeson et al. [23] (char. 27). Previous analyses placed both basihyal and first hypobranchial together in a single character, resulting in a mix of neomorphic and transformational characters. We therefore propose independence among these structures, providing separate characters for their presence/absence and interaction (char. 45–46).
- 39.
- Fourth hypobranchial: (0) Well-developed; (1) reduced (new).
- 37.
- Basibranchial: (0) Segmented; (1) unsegmented (new). According to Shirai [35] Pristiophorus presents an unsegmented basibranchial.
3.1.5. Jaws and Branchial Muscles
- 65.
- Spiracularis: (0) Undivided; (1) divided. Modified from Aschliman et al. [7] (char. 85), divided here into two separate characters, 65 and 66.
- 66.
- Spiracularis (if divided): (0) One bundle enters the dorsal oral membrane underlying the neurocranium; (1) spiracularis splits into lateral and medial bundles, with the medial bundles inserting onto the posterior surface of the Meckel’s cartilage and the lateral bundle onto the dorsal edge of the hyomandibula; (2) spiracularis subdivided proximally and inserts separately into the palatoquadrate and the hyomandibula. This character is proposed to include the character states recognized by Aschliman et al. [7] (char. 85) except for the third state, which seems to be a variation of the first state (splits into lateral and medial bundles).
- 68.
- Coracohyomandibularis: (0) Single origin; (1) separate origins. Modified from Aschliman et al. [7] (char. 88). The character is separated here into two separate characters, 68 and 69, aiming to increase the grouping information on the separate origins of the coracohyomandibularis.
- 69.
- Coracohyomandibularis (if separate origins): (0) Originates in the facia supporting the insertion of the coracoarcualis and on the pericardial membrane; (1) originates on the anterior portion of the ventral gill arch region and on the pericardial membrane.
- 70.
- Coracohyoideus: (0) Present; (1) absent. Modified from Aschliman et al. [7] (char. 89). The character is separated into two different characters, 70 and 71, aiming to increase the grouping information.
- 71.
- Coracohyoideus (if present): (0) Parallel to body axis; (1) runs parallel to the body axis and is very short; (2) runs diagonally from the wall of the first two gill slits to the posteromedial aspect of the basihyal or first basibranchial; (3) each muscle fuses with its antimere at a raphe near its insertion on the first hypobranchial.
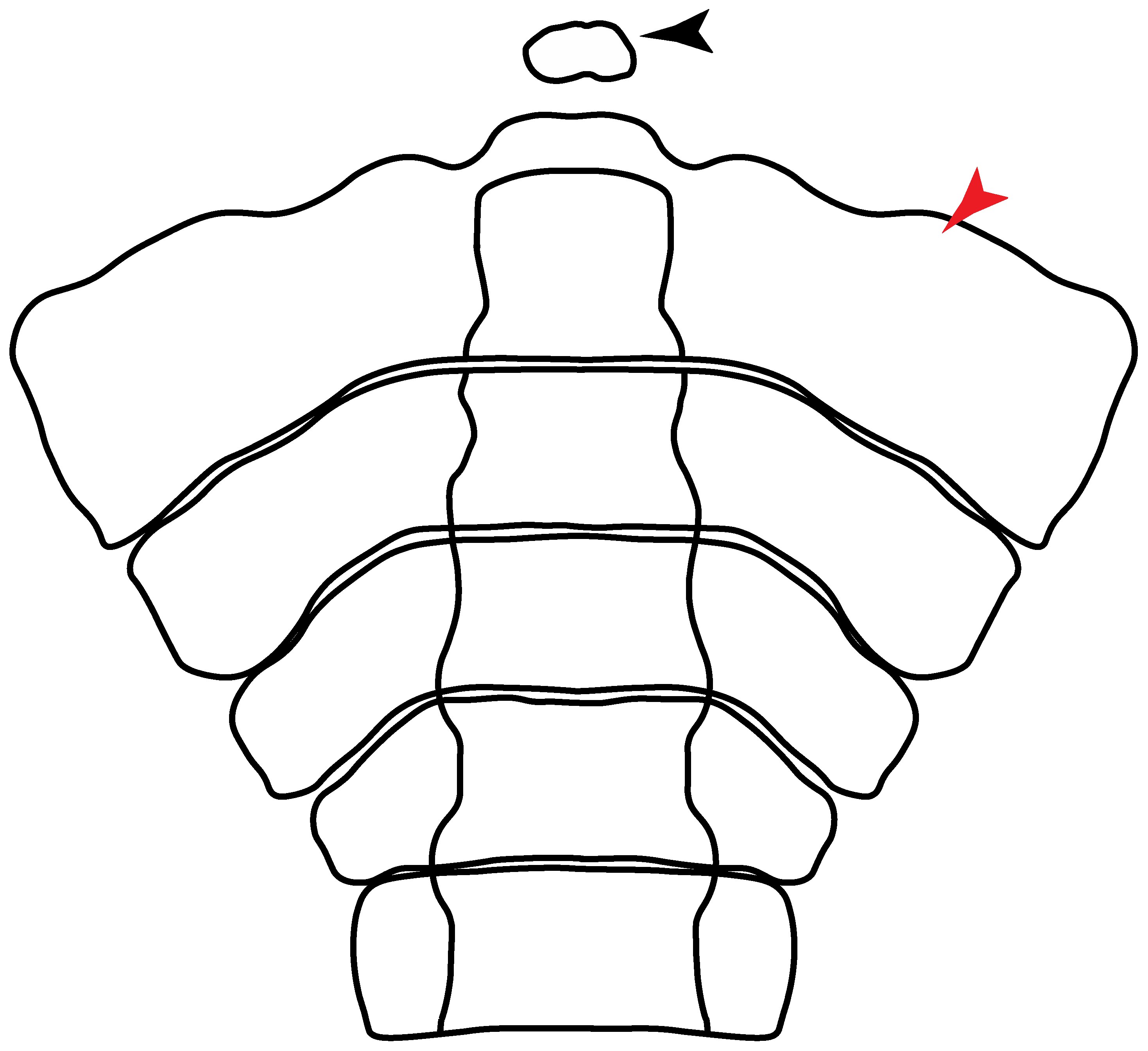
3.1.6. Synarcual and Axial Skeleton
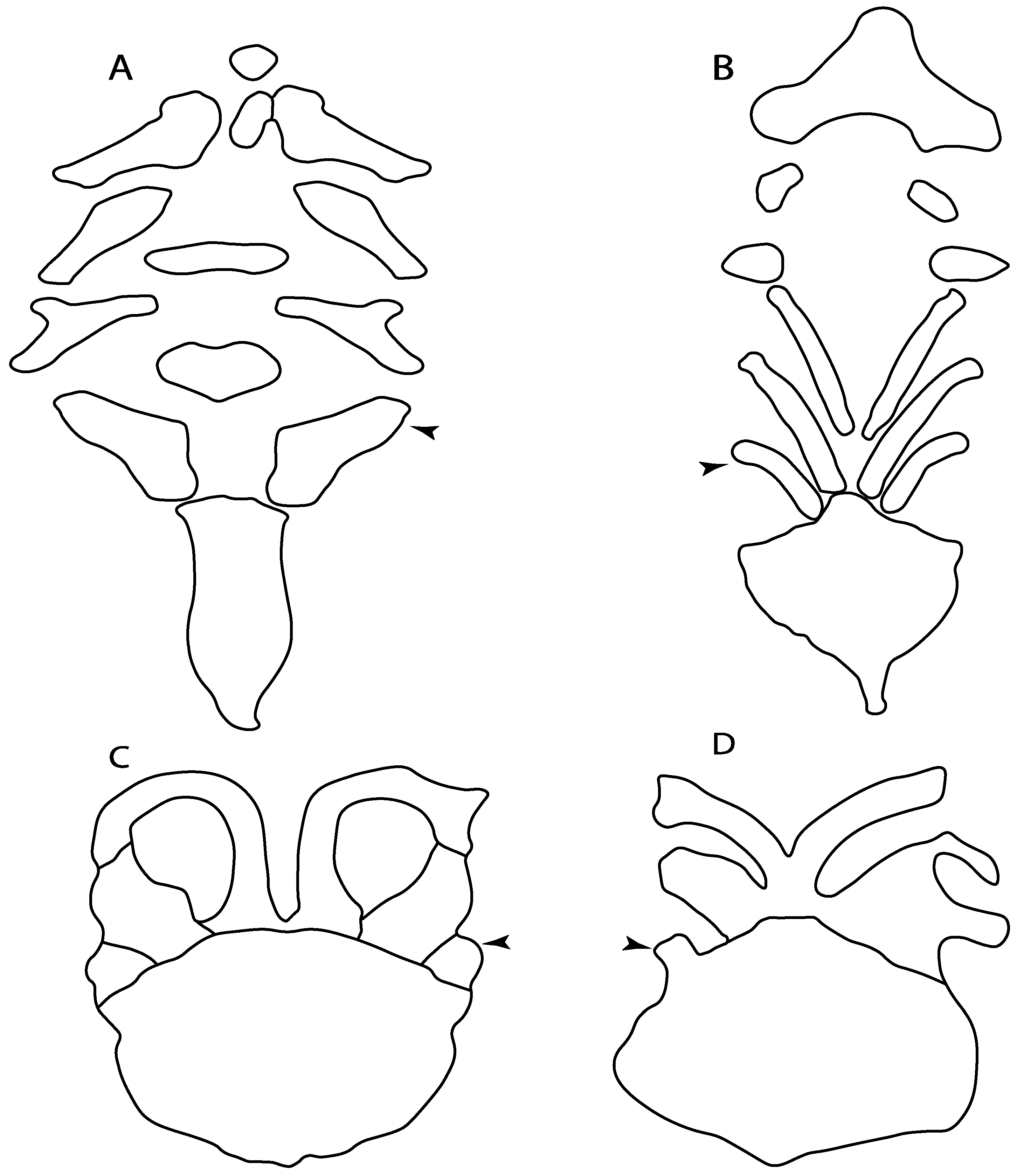
- 48.
- 49.
- Expanded basiventral process of cervical vertebrae: (0) Absent; (1) present. Taken from Maisey et al. [63] (chars. 16–18).
- 50.
- 51.
- Lateral stays: (0) Fused distally with medial crest; (1) free of medial crest (new). Taxa with no synarcual (i.e., outgroups) or with no lateral stays on the cervicothoracic synarcual (i.e., Chimaera and Harriotta) are coded as inapplicable (-), which makes the reconstruction of this character for basal chondrichthyans in the trees impossible.
3.1.7. Suprascapula and Pectoral Girdle
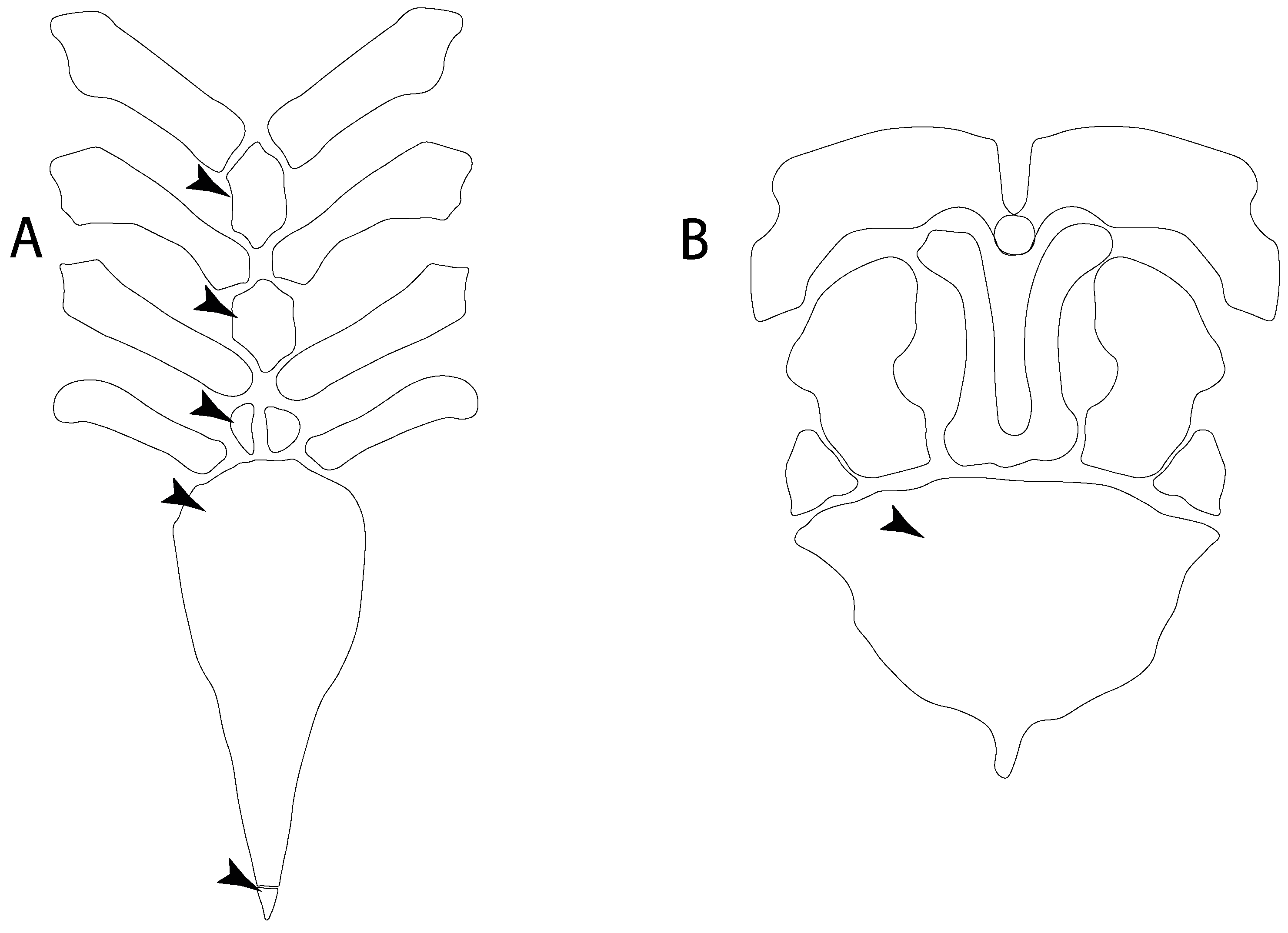
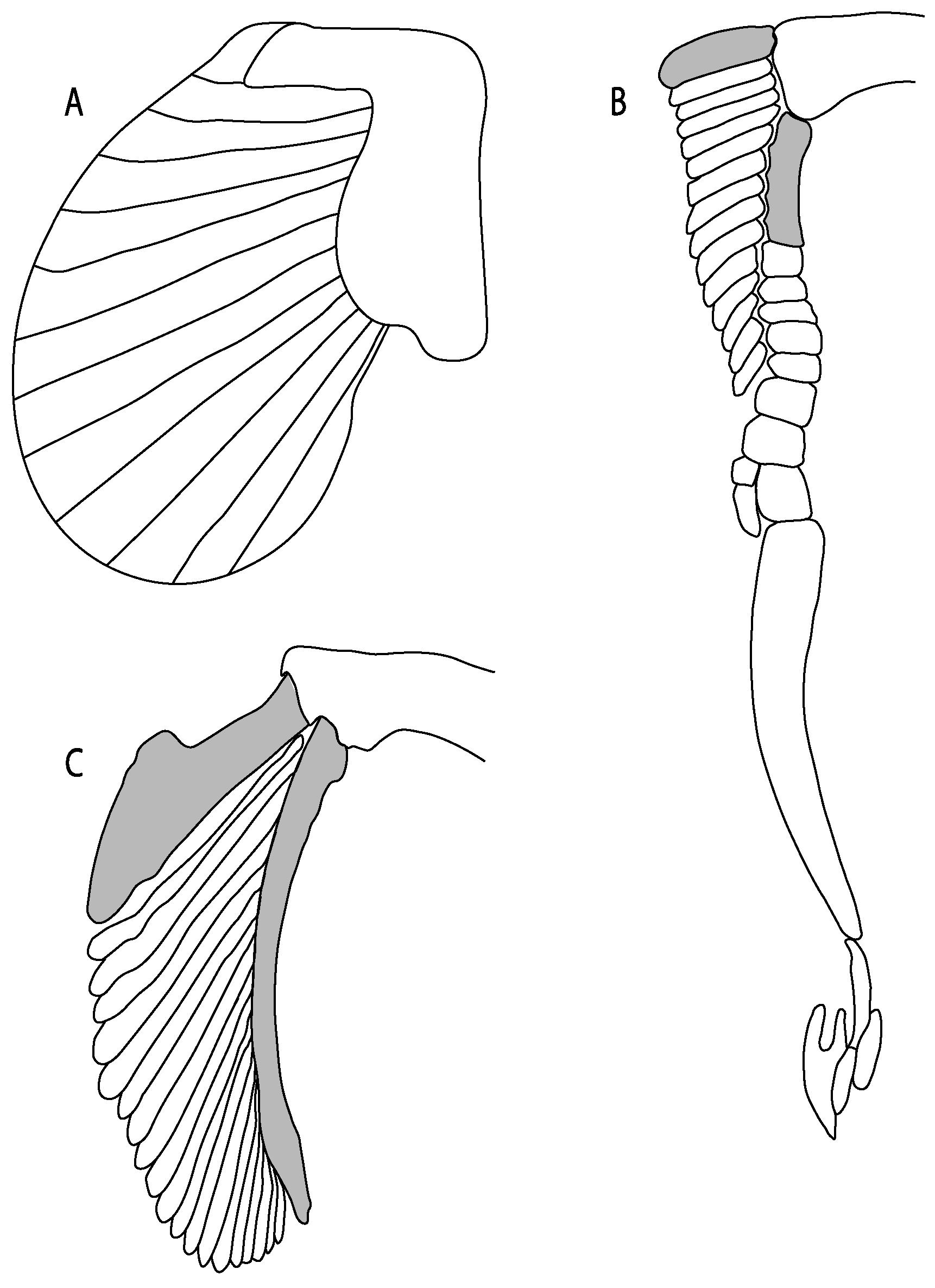
- 93.
- Suprascapula: (0) Absent; (1) fused medially; (2) unfused medially. Modified from Aschliman et al. [7] (char. 6). In some sharks, there seems to be an anterior portion of the scapular process that is detached from the scapula, referred to as suprascapular by Marramà et al. [59]. While this element is dorsal to the scapula, its interaction with other skeletal elements and its development seems to be different from that of the suprascapula of batomorphs.
- 94.
- Suprascapula interaction with axial skeleton (if fused medially): (0) Interacts with axial skeleton (articulated or fused); (1) free from axial skeleton (new). This character is proposed to include the variation observed in the suprascapula articulation in batomorphs.
- 95.
- Suprascapula (if interacts with axial skeleton): (0) Articulates with vertebral column; (1) fused medially to synarcual; (2) fused medially and laterally to synarcual (new). This character is proposed to account for the variation observed in the interaction between the suprascapula and axial skeleton in batomorphs.
- 96.
- Suprascapula-scapula articulation: (0) Curved; (1) crenate; (2) ball and socket; (3) straight. Modified from Aschliman et al. [7] (char. 53) to include the variation observed in the articulation between the suprascapula and scapula in batomorphs.
- 97.
- Crenated suprascapula (variations): (0) With lateral projections; (1) thin upper and lower lobes; (2) upper lobe wider than lower; (3) of similar size and width (new). This character is proposed to include for the variation observed in the suprascapula of Platyrhinidae and Rhinopristiformes.
- 98.
- Scapular process-scapula: (0) Fused; (1) articulated (new). The interaction between the scapula and scapular process is a rather variable within sharks.
- 99.
- Scapular process: (0) Short and dorsally directed; (1) long, U-curved and posteriorly directed; (2) short and posterodorsally directed. Modified from Aschliman et al. [7] (char. 56). The short, posterodorsally directed state was included to account for the variation observed in Pseudobatos.
- 101.
- 102.
- 103.
- Mesocondyle: (0) Single and small; (1) segmented and small; (2) forming an elongated ridge. Modified from Aschliman et al. [7] (char. 59). This character is proposed to include the variation observed in the mesocondyle of the taxa with three condyles (i.e., separated condyles).
3.1.8. Pelvic Girdle and Claspers
- 117.
- Lateral prepelvic processes: (0) Absent; (1) present. The modification of this character from the multistate coding used in McEachran and Dunn [107] (char. 36) is because the three proposed states (i.e., short to moderately long; extremely long with acute tips; and extremely long with biramous tips) are difficult to interpret in fossil specimens. Consequently, binary coding (presence/absence) is used here.
- 118.
- Postpelvic processes: (0) Absent; (1) present. Modified from Claeson et al. [23] (char. 37). Initially observed in Platyrhina and Platyrhinoidis only by de Carvalho [67], the coding of this character was changed for Pseudobatos and Torpedo based on observations by da Silva et al. [10], as these two taxa also show postpelvic processes.
- 119.
- Posterior margin of puboischiadic bar: (0) Straight or bending anteriorly; (1) convex (new).
- 120.
- Anterior margin of puboischiadic bar (if posterior margin straight or concave): (0) Straight; (1) anteriorly arched (new). This character is proposed to group the different patterns of the anterior margin of puboischiadic bar in taxa with a straight or anteriorly bending posterior margin.
- 124.
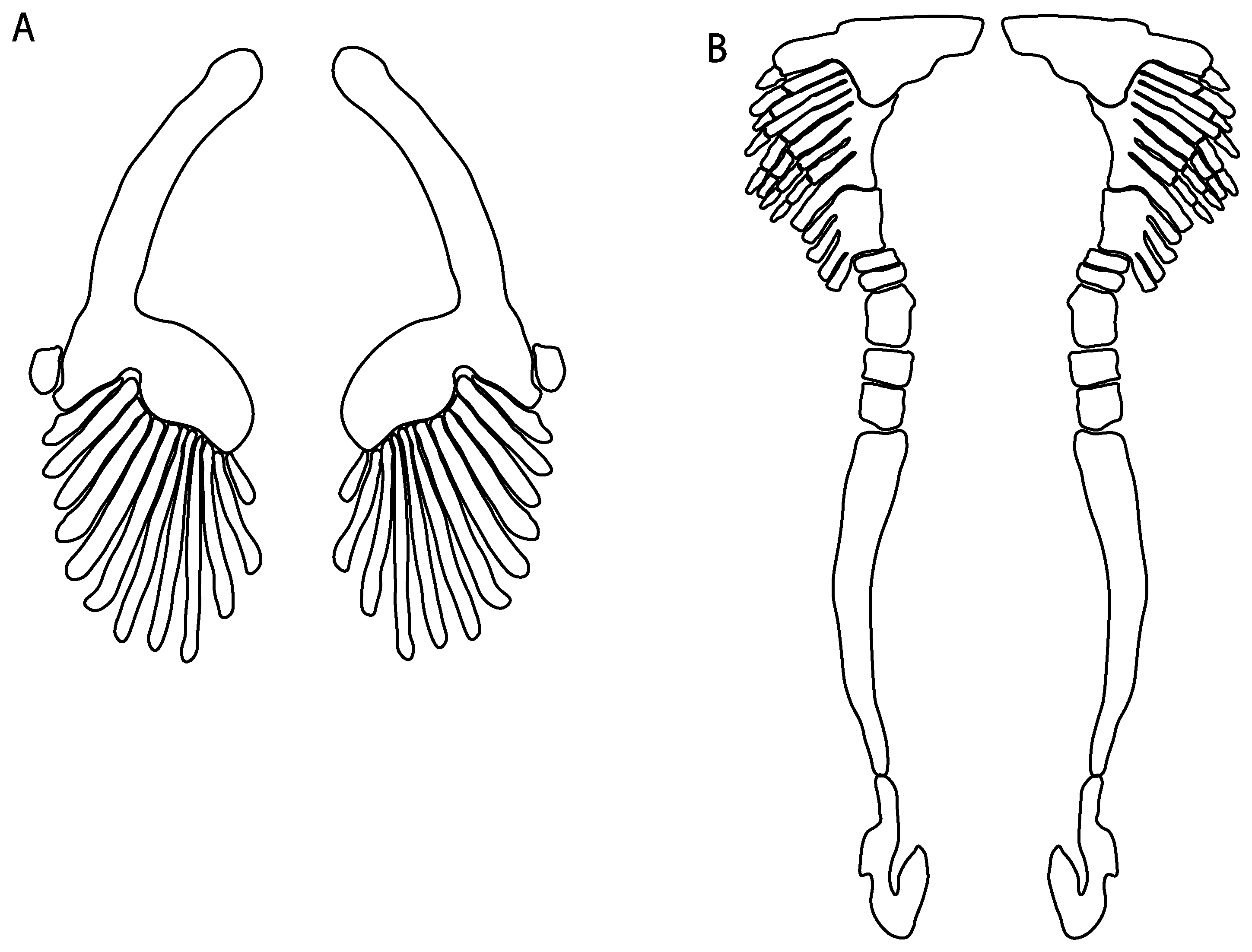
- Pelvic girdle: (0) Separated; (1) fused. Modified from Maisey [13] (char, 37), based on observations by Klug et al. [20] (Figure 2), Stumpf et al. [108] and Coates et al. [109]. Current fossil evidence suggests that the separation of two halves, or at least a not very well-mineralized mid-bar of the pelvic girdle, is the basal state across hybodontiform-like sharks (Figure 42A,B) (SMNS 10062) (NHMUK PV P 339).
3.1.9. Paired Fins
- 91.
- Radial calcification: (0) Crustal; (1) catenated. Taken from Marramà et al. [33] (char. 104).
- 92.
- Radial calcification (if catenated): (0) Two chains; (1) four chains (new). This character includes the remaining variation observed by Schaefer and Summers [63] regarding the number of chains.
3.1.10. Pectoral Fins
- 107.
- Propterygium extending anteriorly: (0) Absent; (1) present. Modified from Aschliman et al. [7] (char. 62), based on de Carvalho and Maisey [15] (char. 65). The original character was proposed as a synapomorphy for platyrhinids or as a shared feature between platyrhinids and Zanobatus, which according to de Carvalho [66] also present the following condition: extension of the propterygium and its associated radials to the anterior margin of the disc on both sides of the snout and rostrum. Aschliman et al. [7] suggested that the extension of the propterygium observed in platyrhinids and Zanobatus is similar to the condition present in Myliobatiformes and Bathyraja. However, in pelagic stingrays (e.g., Myliobatis Aetobatus, Rhinoptera and Mobula), the head stands out of the pectoral disc, causing modifications to the neurocranium and pectoral disc, suggesting differences in this structure. In contrast, the condition of Rajiformes resembles that of the remaining batomorphs.
- 108.
- First segment of propterygium (if propterygium extends anteriorly): (0) Not reaching the nasal capsules; (1) reaches the level of nasal capsules; (2) extending well beyond the nasal capsules. This character recovers the variation of the placement of the first propterygial segment with respect to the nasal capsules of Aschliman et al. [7] (char. 62 and 63).
- 109.
- 111.
3.1.11. Pelvic Fins
- 122.
- Overdevelopment of first pelvic radial: (0) Absent; (1) present (new).
- 60.
- Pelvic basipterygium: (0) Fused to first radial; (1) separated from first radial. Based on Riley et al. [110].
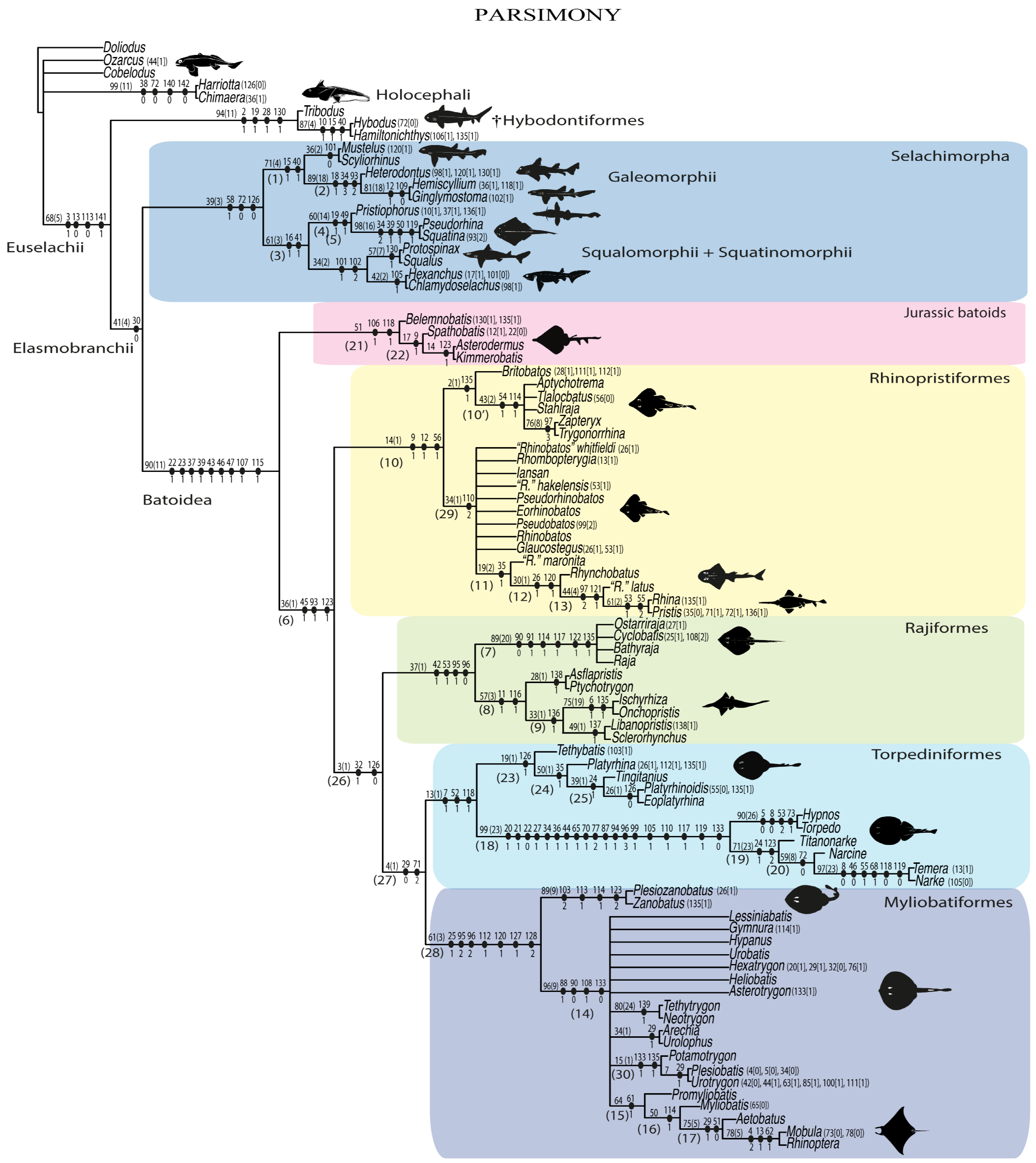
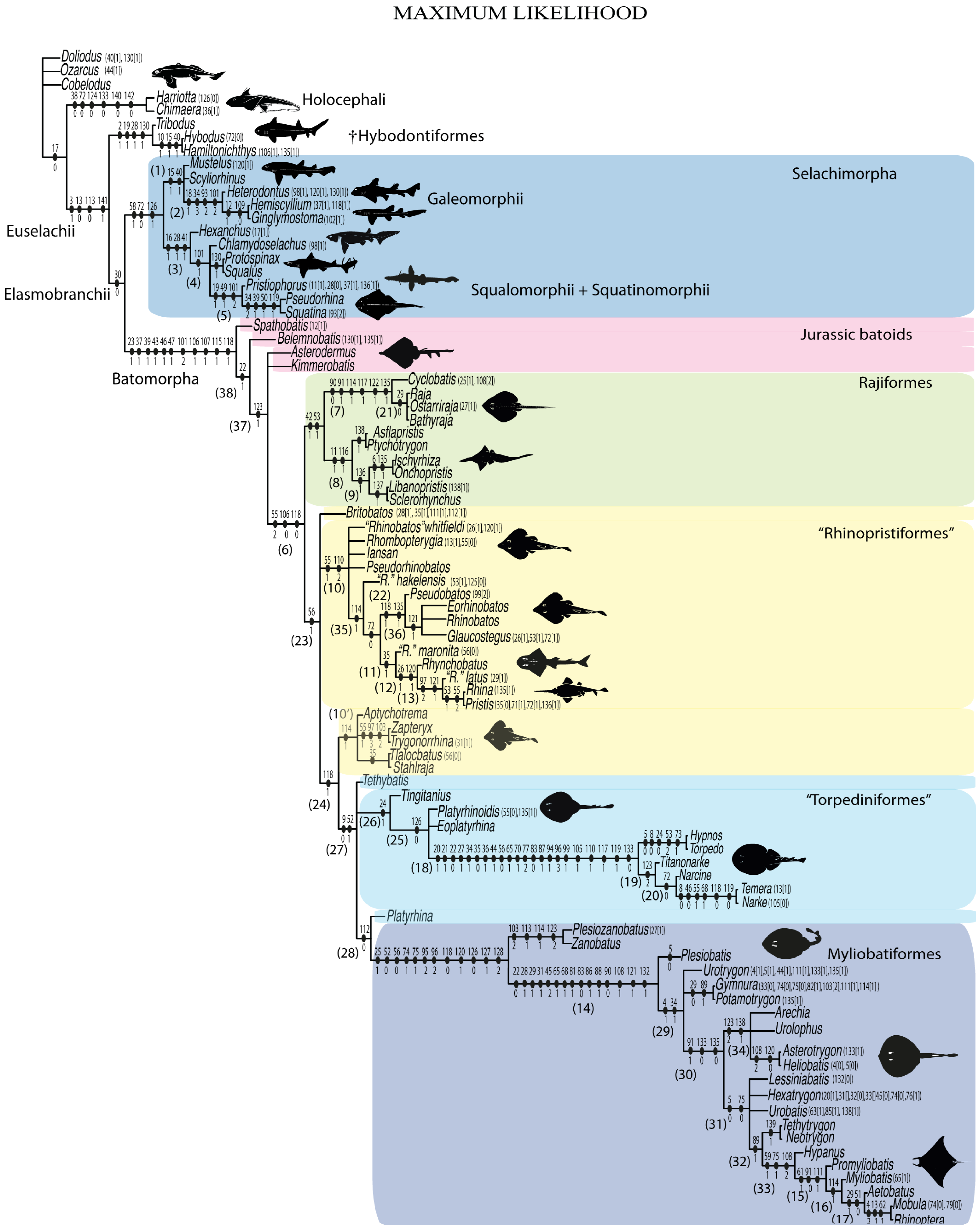
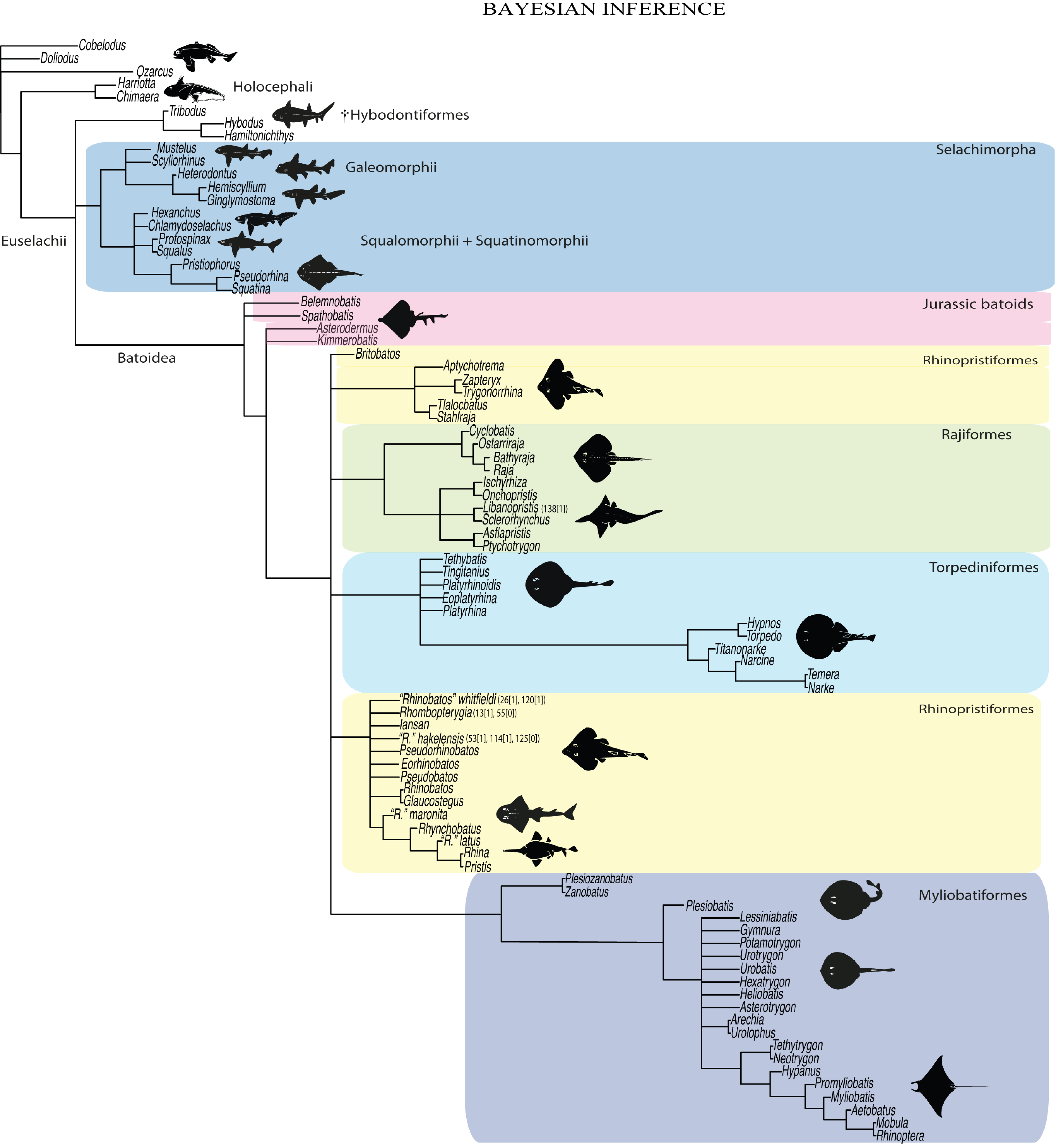
3.2. Phylogenetic Analyses
4. Discussion
4.1. Phylogenetic Analyses
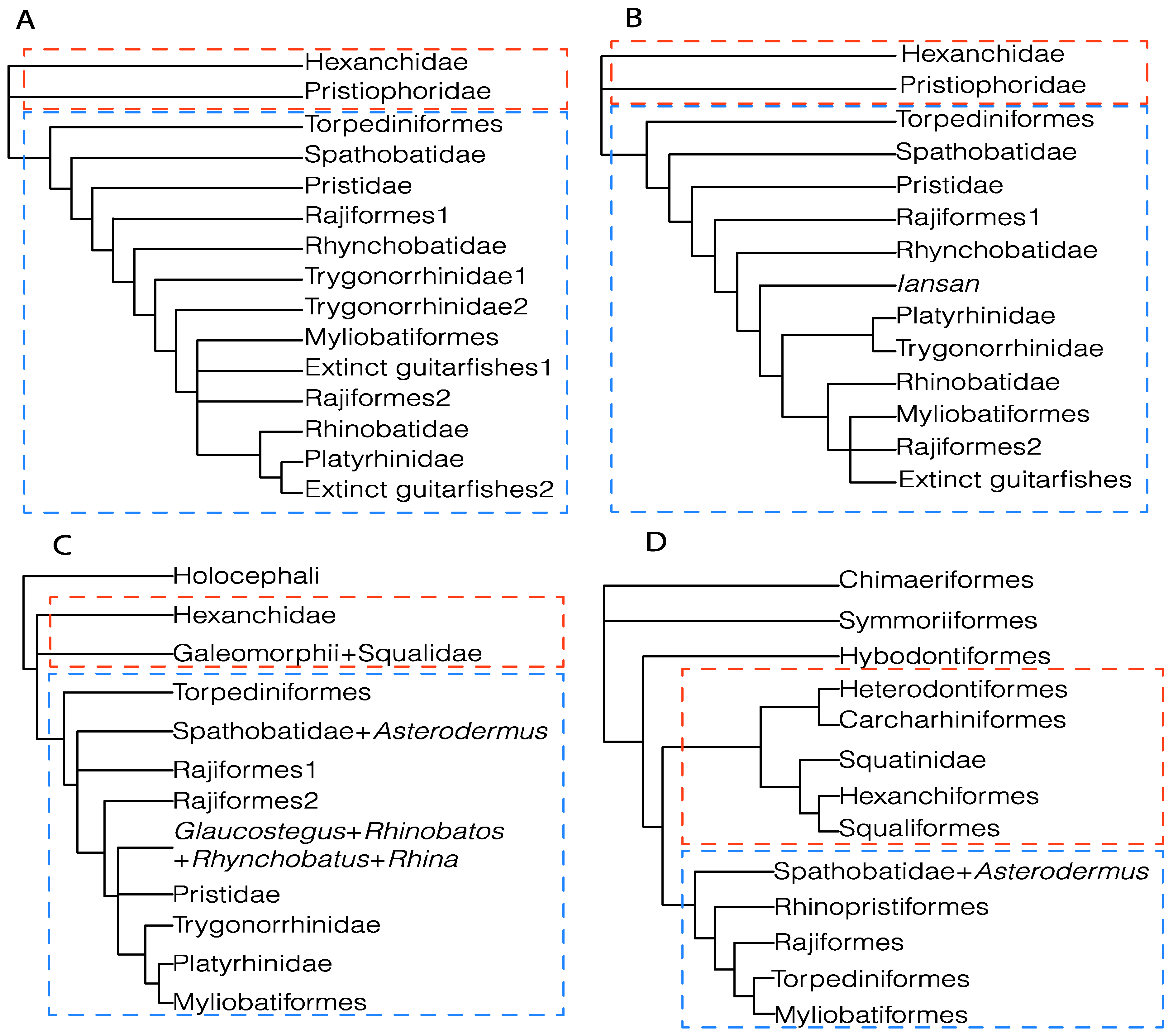
4.1.1. Parsimony
4.1.2. Maximum-Likelihood Analysis (ML)
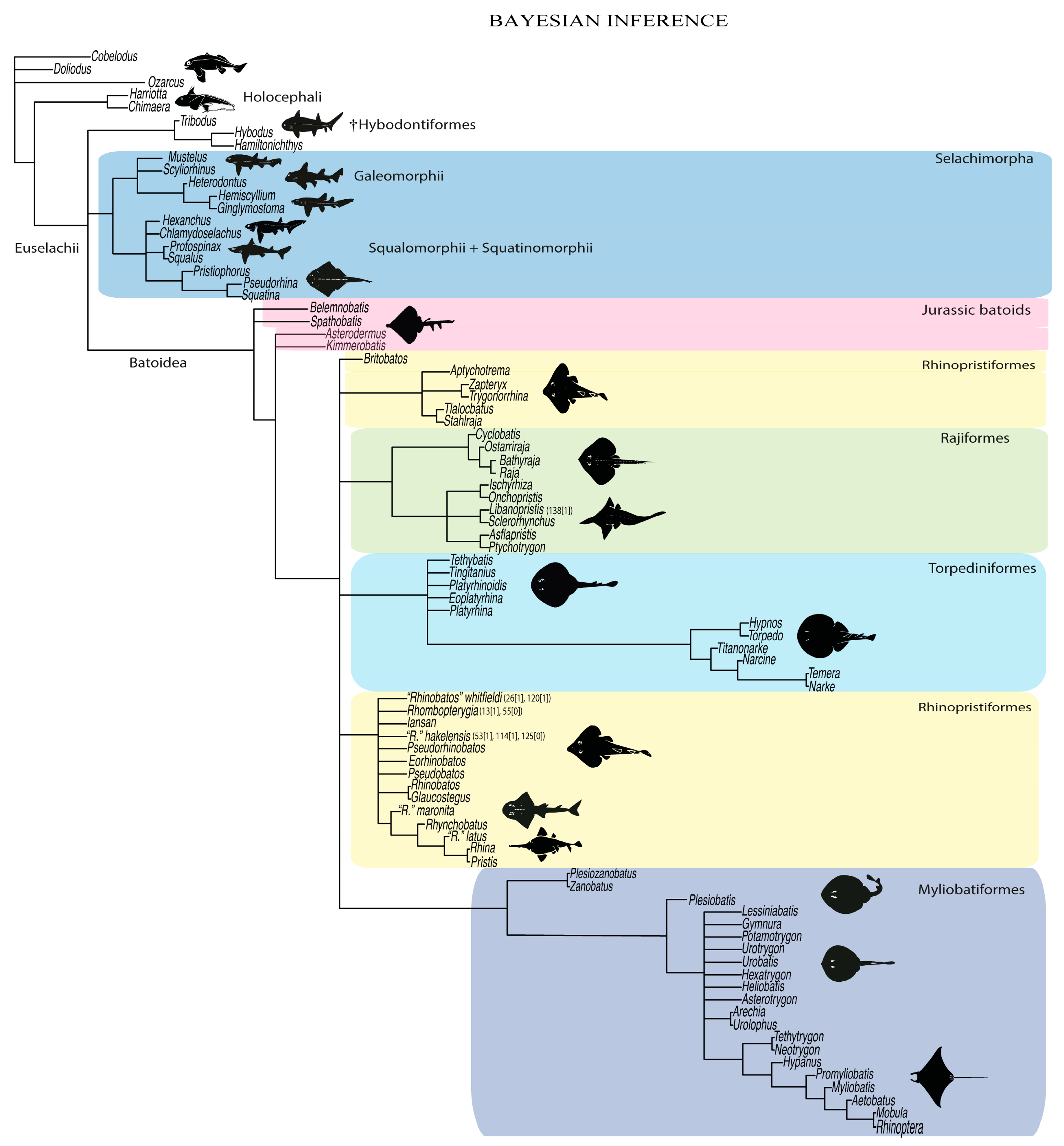
4.1.3. Bayesian Inference Analysis (BI)
4.2. Phylogenetic Considerations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maisey, J.G. What is an “elasmobranch”? The impact of palaeontology in understanding elasmobranch phylogeny and evolution. J. Fish Biol. 2012, 80, 918–951. [Google Scholar] [CrossRef]
- Fricke, R.; Eschmeyer, W.N.; Fong, J.D. Eschmeyer’s Catalog of Fishes: Genera/Species by Family/Subfamily. Available online: https://researcharchive.calacademy.org/research/ichthyology/catalog/SpeciesByFamily.asp (accessed on 28 February 2022).
- Last, P.; Naylor, G.J.; Bernard, S.; White, W.; de Carvalho, M.R.; Stehmann, M. Rays of the World; Csiro Publishing: Clayton South, Australia, 2016; 790p. [Google Scholar]
- Compagno, L.J. Phyletic relationships of living sharks and rays. Am. Zool. 1977, 17, 303–322. [Google Scholar] [CrossRef] [Green Version]
- Compagno, L.J. Interrelationships of living elasmobranchs. Zool. J. Linn. Soc. 1973, 53, 15–61. [Google Scholar]
- McEachran, J.D.; Aschliman, N. Phylogeny of Batoidea. In Biology of Sharks and Their Relatives; Carrier, J.C., Musick, J.A., Heithaus, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2004; Volume 1, pp. 79–113. [Google Scholar]
- Aschliman, N.C.; Claeson, K.M.; McEachran, J.D. Phylogeny of Batoidea. In Biology of Sharks and Their Relatives; Carrier, J.C., Musick, J.A., Heithaus, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2012; Volume 2, pp. 57–95. [Google Scholar]
- Da Silva, J.P.C.; Carvalho, M.R. Morphology and phylogenetic significance of the pectoral articular region in elasmobranchs (Chondrichthyes). Zool. J. Linn. Soc. 2015, 175, 525–568. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, J.P.C.; Vaz, D.F.; Carvalho, M.R. Phylogenetic inferences on the systematics of squaliform sharks based on elasmobranch scapular morphology (Chondrichthyes: Elasmobranchii). Zool. J. Linn. Soc. 2018, 182, 614–630. [Google Scholar] [CrossRef]
- Da Silva, J.P.C.; Silva–Loboda, T.; Souza, R.R. A new synapomorphy in the pelvic girdle reinforces a close relationship of Zanobatus and Myliobatiformes (Chondrichthyes: Batoidea). J. Anat. 2021, 238, 874–885. [Google Scholar] [CrossRef]
- Mooi, R.D.; Gill, A.C. Phylogenies without synapomorphies—A crisis in fish systematics: Time to show some character. Zootaxa 2010, 2450, 26–40. [Google Scholar] [CrossRef] [Green Version]
- Maisey, J.G.; Naylor, G.J.; Ward, D.J. Mesozoic elasmobranchs, neoselachian phylogeny and the rise of modern elasmobranch diversity. In Mesozoic Fishes Systematics Paleoenvironments and Biodiversity; Arriatia, G., Tintori, A., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2004; Volume 3, pp. 17–56. [Google Scholar]
- Maisey, J.G. Chondrichthyan phylogeny: A look at the evidence. J. Vertebr. Paleontol. 1984, 4, 359–371. [Google Scholar] [CrossRef]
- Maisey, J.G. Higher elasmobranch phylogeny and biostratigraphy. Zool. J. Linn. Soc. 1984, 82, 33–54. [Google Scholar] [CrossRef]
- De Carvalho, M.R.; Maisey, J.G. The Phylogenetic Relationship of the Late Jurassic Shark Protospinax Woodward 1919 (Chondrichthyes: Elasmobranchii). In Mesozoic Fishes Systematics and Paleoecology; Arriatia, G., Viohl, G., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 1996; pp. 9–46. [Google Scholar]
- Brito, P.; Dutheil, D.B. A preliminary systematic analysis of Cretaceous guitarfishes from Lebanon. In Mesozoic Fishes Systematics Paleoenvironments and Biodiversity; Arriatia, G., Tintori, A., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2004; Volume 3, pp. 101–109. [Google Scholar]
- Maisey, J.G. Braincase of the Upper Devonian shark Cladodoides wildungensis (Chondrichthyes, Elasmobranchii), with observations on the braincase in early chondrichthyans. Bull. Am. Mus. Nat. Hist. 2005, 288, 1–103. [Google Scholar] [CrossRef]
- Maisey, J.G. The braincase in Paleozoic symmoriiform and cladoselachian sharks. Bull. Am. Mus. Nat. Hist. 2007, 2007, 1–122. [Google Scholar] [CrossRef] [Green Version]
- Klug, S. A new palaeospinacid shark (Chondrichthyes, Neoselachii) from the Upper Jurassic of southern Germany. J. Vertebr. Paleontol. 2009, 2, 326–335. [Google Scholar] [CrossRef] [Green Version]
- Klug, S. Monophyly, phylogeny and systematic position of the †Synechodontiformes (Chondrichthyes, Neoselachii). Zool. Scr. 2010, 39, 37–49. [Google Scholar] [CrossRef]
- Lane, J.A.; Maisey, J.G. The visceral skeleton and jaw suspension in the durophagous hybodontid shark Tribodus limae from the Lower Cretaceous of Brazil. J. Paleontol. 2012, 86, 886–905. [Google Scholar] [CrossRef]
- Pradel, A.; Maisey, J.G.; Tafforeau, P.; Mapes, R.H.; Mallatt, J. A Palaeozoic shark with osteichthyan-like branchial arches. Nature 2014, 509, 608–611. [Google Scholar] [CrossRef] [PubMed]
- Claeson, K.M.; Underwood, C.J.; Ward, D. †Tingitanius tenuimandibulus, a new platyrhinid batoid from the Turonian (Cretaceous) of Morocco and the Cretaceous radiation of the Platyrhinidae. J. Vertebr. Paleontol. 2013, 33, 1019–1036. [Google Scholar] [CrossRef]
- Underwood, C.J.; Claeson, K.M. The late Jurassic ray Kimmerobatis etchesi gen. et sp. nov. and the Jurassic radiation of the Batoidea. Proc. Geol. Assoc. 2017, 130, 345–354. [Google Scholar] [CrossRef]
- Coates, M.I.; Gess, R.W.; Finarelli, J.A.; Criswell, K.E.; Tietjen, K. A symmoriiform chondrichthyan braincase and the origin of chimaeroid fishes. Nature 2017, 541, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Coates, M.I.; Finarelli, J.A.; Sansom, I.J.; Andreev, P.S.; Criswell, K.E.; Tietjen, K.; Rivers, M.L.; La Riviere, P.J. An early chondrichthyan and the evolutionary assembly of a shark body plan. Proc. R. Soc. B. 2018, 285, 20172418. [Google Scholar] [CrossRef]
- Maisey, J.G.; Miller, R.; Pradel, A.; Denton, J.S.; Bronson, A.; Janvier, P. Pectoral morphology in Doliodus: Bridging the ‘acanthodian’-chondrichthyan divide. Am. Mus. Novit. 2017, 2017, 1–15. [Google Scholar] [CrossRef]
- Marramà, G.; Claeson, K.M.; Carnevale, G.; Kriwet, J. Revision of Eocene electric rays (Torpediniformes, Batomorphii) from the Bolca Konservat–Lagerstätte, Italy, reveals the first fossil embryo in situ in marine batomorphs and provides new insights into the origin of trophic novelties in coral reef fishes. J. Syst. Palaeontol. 2018, 16, 1189–1219. [Google Scholar] [CrossRef] [Green Version]
- Marramà, G.; Klug, S.; De Vos, J.; Kriwet, J. Anatomy, relationships and palaeobiogeographic implications of the first Neogene holomorphic stingray (Myliobatiformes: Dasyatidae) from the early Miocene of Sulawesi, Indonesia, SE Asia. Zool. J. Linn. Soc. 2018, 184, 1142–1168. [Google Scholar] [CrossRef] [Green Version]
- Marramà, G.; Schultz, O.; Kriwet, J. A new Miocene skate from the Central Paratethys (Upper Austria): The first unambiguous skeletal record for the Rajiformes (Chondrichthyes: Batomorphii). J. Syst. Palaeontol. 2019, 17, 937–960. [Google Scholar] [CrossRef] [Green Version]
- Marramà, G.; Carnevale, G.; Naylor, G.J.; Kriwet, J. Mosaic of plesiomorphic and derived characters in an Eocene myliobatiform batomorph (Chondrichthyes, Elasmobranchii) from Italy defines a new, basal body plan in pelagic stingrays. Zool. Lett. 2019, 5, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Villalobos-Segura, E.; Underwood, C.J.; Ward, D.J.; Claeson, K.M. The first three–dimensional fossils of Cretaceous sclerorhynchid sawfish: Asflapristis cristadentis gen. et sp. nov., and implications for the phylogenetic relations of the Sclerorhynchoidei (Chondrichthyes). J. Syst. Palaeontol. 2019, 17, 1847–1870. [Google Scholar] [CrossRef] [Green Version]
- Marramà, G.; Carnevale, G.; Claeson, K.M.; Naylor, G.J.; Kriwet, J. Revision of the Eocene “Platyrhina” species from the Bolca Lagerstätte (Italy) reveals the first panray (Batomorphii: Zanobatidae) in the fossil record. J. Syst. Palaeontol. 2020, 18, 1519–1542. [Google Scholar] [CrossRef] [PubMed]
- Underwood, C.J. Diversification of the Neoselachii (Chondrichthyes) during the Jurassic and Cretaceous. Paleobiology 2006, 32, 215–235. [Google Scholar] [CrossRef] [Green Version]
- Shirai, S. Squalean Phylogeny: A New Framework of “Squaloid” Sharks and Related Taxa. Ph.D. Thesis, Hokkaido University, Sapporo, Japan, 1992; 212p. [Google Scholar]
- Shirai, S. Phylogenetic relationships of the angel sharks, with comments on elasmobranch phylogeny (Chondrichthyes, Squatinidae). Copeia 1992, 1992, 505–518. [Google Scholar] [CrossRef]
- Shirai, S. Phylogenetic interrelationships of neoselachians (Chondrichthyes: Euselachii). In Interrelationships of Fishes; Stiassny, M.L., Parenti, L.R., Johnson, G.D., Eds.; Atlantic Press: New York, NY, USA, 1996; pp. 9–34. [Google Scholar]
- De Carvalho, M.R. Higher-level elasmobranch phylogeny, basal squaleans, and paraphyly. In Interrelationships of Fishes; Stiassny, M.L., Parenti, L.R., Johnson, G.D., Eds.; Atlantic Press: New York, NY, USA, 1996; pp. 593–660. [Google Scholar]
- Dunn, K.A.; Morrissey, J.F. Molecular phylogeny of elasmobranchs. Copeia 1995, 1995, 526–531. [Google Scholar] [CrossRef]
- Douady, C.J.; Dosay, M.; Shivji, M.S.; Stanhope, M.J. Molecular phylogenetic evidence refuting the hypothesis of Batoidea (rays and skates) as derived sharks. Mol. Phylogenet. Evol. 2003, 26, 215–221. [Google Scholar] [CrossRef]
- Naylor, G.J.; Caira, J.N.; Jensen, K.; Rosana, K.A.; Straube, N.; Lakner, C. Elasmobranch phylogeny: A mitochondrial estimate based on 595 species. In Biology of Sharks and Their Relatives; Carrier, J.C., Musick, J.A., Heithaus, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2012; Volume 2, pp. 31–56. [Google Scholar]
- Aschliman, N.C.; Nishida, M.; Miya, M.; Inoue, J.G.; Rosana, K.M.; Naylor, G.J. Body plan convergence in the evolution of skates and rays (Chondrichthyes: Batoidea). Mol. Phylogenet. Evol. 2012, 63, 28–42. [Google Scholar] [CrossRef] [PubMed]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 3.61. 2019. Available online: https://www.mesquiteproject.org (accessed on 21 April 2021).
- Lyons–Weiler, J.; Hoelzer, G.A.; Tausch, R.J. Optimal outgroup analysis. Biol. J. Linn. Soc. 1998, 64, 493–511. [Google Scholar] [CrossRef]
- Wilberg, E.W. What’s in an outgroup? The impact of outgroup choice on the phylogenetic position of Thalattosuchia (Crocodylomorpha) and the origin of Crocodyliformes. Syst. Biol. 2015, 64, 621–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goloboff, P.A.; Farris, S.; Nixon, K. TNT, a free program for phylogenetic analysis. Cladistics 2008, 24, 774–786. [Google Scholar] [CrossRef]
- Goloboff, P.A.; Catalano, S.A. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 2016, 32, 221–238. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods); Version 4; Sinauer Associates: Sunderland, MA, USA, 2003. [Google Scholar]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MRBAYES 3.2: Efficient Bayesian phylogenetic inference and model selection across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Heemstra, P.C.; Smith, M.M. Hexatrygonidae, a new family of stingrays (Myliobatiformes: Batoidea) from South Africa, with comments on the classification of batoid fishes. Ichthyol. Bull. 1980, 43, 1–17. [Google Scholar]
- Pradeep, H.D.; Swapnil, S.S.; Nashad, M.; Venu, S.; Ravi, R.K.; Sumitha, G.; Monalisha, D.S.; Farejiya, M.K. First record and DNA Barcoding of Oman cownose ray, Rhinoptera jayakari Boulenger, 1895 from Andaman Sea, India. Zoosystema 2018, 40, 67–74. [Google Scholar] [CrossRef]
- Steven, G.A. Rays and skates of Devon and Cornwall. Methods of rapid identification on the fishmarket. J. Mar. Biolog. Assoc. 1931, 17, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Last, P.R.; Ho, H.C.; Chen, R.R. A new species of wedgefish, Rhynchobatus immaculatus (Chondrichthyes, Rhynchobatidae), from Taiwan. Zootaxa 2013, 3752, 185–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wueringer, B.; Peverell, S.C.; Seymour, J.; Squire, J.L.; Collin, S.P. Sensory systems in sawfishes. The lateral line. Brain Behav. Evol. 2011, 78, 150–161. [Google Scholar] [CrossRef]
- De Beer, G.R.; Moy–Thomas, J.A. VI—On the skull of Holocephali. Philos. Trans. R. Soc. B. 1935, 224, 287–312. [Google Scholar]
- Miyake, T.; McEachran, J.D.; Walton, P.J.; Hall, B.K. Development and morphology of rostral cartilages in batoid fishes (Chondrichthyes: Batoidea), with comments on homology within vertebrates. Biol. J. Linn. Soc. 1992, 46, 259–298. [Google Scholar] [CrossRef]
- Maisey, J.G. The anatomy and interrelationships of Mesozoic hybodont sharks. Am. Mus. Novit. 1982, 2724, 1–17. [Google Scholar]
- Lane, J.A. Morphology of the braincase in the Cretaceous hybodont shark Tribodus limae (Chondrichthyes: Elasmobranchii), based on CT scanning. Am. Mus. Novit. 2010, 2010, 1–70. [Google Scholar] [CrossRef] [Green Version]
- Marramà, G.; Carnevale, G.; Naylor, G.J.; Kriwet, J. Skeletal Anatomy, Phylogenetic Relationships, and Paleoecology of the Eocene Urolophid Stingray Arechia crassicaudata (Blainville, 1818) from Monte Postale (Bolca Lagerstätte, Italy). Zool. J. Linn. Soc. 2020, 40, e1803339. [Google Scholar] [CrossRef]
- Claeson, K.M. The impacts of comparative anatomy of electric rays (Batoidea: Torpediniformes) on their systematic hypotheses. J. Morphol. 2014, 275, 597–612. [Google Scholar] [CrossRef] [PubMed]
- Motta, P.J.; Wilga, C.D. Anatomy of the feeding apparatus of the nurse shark, Ginglymostoma cirratum. J. Morphol. 1999, 241, 33–60. [Google Scholar] [CrossRef]
- Miyake, T. The Systematics of the Stingray Genus Urotrygon with Comments on the Interrelationships within Urolophidae (Chondrichthyes, Myliobatiformes). Ph.D. Thesis, Texas A&M University, College Station, TX, USA, 1988. Volumes I and II. 518p. [Google Scholar]
- Schaefer, J.T.; Summers, A.P. Batoid wing skeletal structure: Novel morphologies, mechanical implications, and phylogenetic patterns. J. Morphol. 2005, 264, 34298–34313. [Google Scholar] [CrossRef]
- Seidel, R.; Jayasankar, A.K.; Dean, M.N. The multiscale architecture of tessellated cartilage and its relation to function. J. Fish Biol. 2021, 98, 942–955. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, M.R.; Maisey, J.G.; Grande, L. Freshwater stingrays of the Green River Formation of Wyoming (Early Eocene), with the description of a new genus and species and an analysis of its phylogenetic relationships (Chondrichthyes: Myliobatiformes). Bull. Am. Mus. Nat. Hist. 2004, 284, 1–136. [Google Scholar] [CrossRef]
- Mollen, F.H.; Wintner, S.; Iglesias, S.P.; Van Sommeran, S.R.; Jagt, J.W. Comparative morphology of rostral cartilages in extant mackerel sharks (Chondrichthyes, Lamniformes, Lamnidae) using CT scanning. Zootaxa 2012, 3340, 29–43. [Google Scholar] [CrossRef]
- De Carvalho, M.R. A Late Cretaceous thornback ray from southern Italy, with a phylogenetic reappraisal of the Platyrhinidae (Chondrichthyes: Batoidea). In Mesozoic fishes Systematics Paleoenvironments and Biodiversity; Arriatia, G., Tintori, A., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2004; Volume 3, pp. 75–100. [Google Scholar]
- De Carvalho, M.R. A Systematic Revision of the Electric Ray Genus Narcine Henle, 1834 (Chondrichthyes: Torpediniformes: Narcinidae), and the Higher-Level Phylogenetic Relationships of the Orders of Elasmobranch Fishes (Chondrichthyes). Ph.D. Thesis, City University of New York, New York, NY, USA, 1999; 741p. [Google Scholar]
- Holmgren, N. Studies on the head in fishes. Part 2. Comparative anatomy of the adult selachian skull with remarks on the dorsal fins in sharks. Acta Zool. 1941, 22, 1–100. [Google Scholar] [CrossRef]
- Baranes, A.; Randal, J.E. Narcine bentuviai, a new torpedinoid ray from the Northern Red Sea. Isr. J. Zool. 1989, 36, 85–101. [Google Scholar]
- Claeson, K.M. Trends in Evolutionary Morphology: A Case Study in the Relationships of Angel Sharks and Batoid Fishes. Ph.D. Thesis, The University of Texas at Austin, Austin, TX, USA, 2010; 254p. [Google Scholar]
- McEachran, J.D.; Dunn, K.A.; Miyake, T. Interrelationships of the batoid fishes (Chondrichthyes: Batoidea). In Interrelationships of Fishes; Stiassny, M.L., Parenti, L.R., Johnson, G.D., Eds.; Atlantic Press: New York, NY, USA, 1996; pp. 63–84. [Google Scholar]
- Fechhelm, J.D.; McEachran, J.D. A revision of the electric ray genus Diplobatis with notes on the interrelationships of Narcinidae (Chondrichthyes, Torpediniformes). Bull. Fla. State Mus. Biol. Sci. 1984, 29, 171–209. [Google Scholar]
- Rincon, G.; Stehmann, M.F.W.; Vooren, C.M. Results of the research cruises of FRV Walther Herwig to South America. LXXIV. Benthobatis kreffti sp. nov. (Chondrichthyes, Torpediniformes, Narcinidae), a new deep-water electric ray from off South Brazil and the third species of the genus. Arch. Fish. Mar. Res. 2001, 49, 45–60. [Google Scholar]
- McEachran, J.D.; Compagno, L.J. Interrelationships of and within Breviraja based on anatomical structures (Pisces: Rajoidei). Bull. Mar. Sci. 1982, 32, 399–425. [Google Scholar]
- Wueringer, B.; Squire, L.; Collin, S. The biology of extinct and extant sawfish (Batoidea: Sclerorhynchidae and Pristidae). Rev. Fish Biol. 2009, 19, 445–465. [Google Scholar] [CrossRef]
- Cappetta, H. Les Selaciens du Cretace superieur du Liban. II. Batoïdes. Palaeontogr. Abt. A 1980, 168, 149–229. [Google Scholar]
- Allis, E.P. The cranial anatomy of Chlamydoselachus anguineus. Acta Zool. 1923, 4, 123–221. [Google Scholar] [CrossRef]
- Ewart, J.C. IV. The cranial nerves of the Torpedo. (Preliminary note). Proc. R. Soc. Lond. 1890, 47, 290–291. [Google Scholar]
- Smith, M.M.; Riley, A.; Fraser, G.J.; Underwood, C.J.; Welten, M.; Kriwet, J.; Pfaff, C.; Johanson, Z. Early development of rostrum saw-teeth in a fossil ray tests classical theory of the evolution of vertebrate dentitions. Proc. R. Soc. B Biol. Sci. 2015, 282, 20151628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, D.B.; Hiscox, J.D.; Dixon, B.J.; Potgieter, J. 3D scanning and printing skeletal tissues for anatomy education. J. Anat. 2016, 229, 473–481. [Google Scholar] [CrossRef]
- Nishida, K. Phylogeny of the suborder Myliobatidoidei. Mem. Fac. Fish. Hokkaido Univ. 1990, 37, 1–108. [Google Scholar]
- Maisey, J.G.; Ehret, D.J.; Denton, J.S. A new genus of Late Cretaceous angel shark (Elasmobranchii; Squatinidae), with comments on squatinid phylogeny. Am. Mus. Novit. 2020, 2020, 1–29. [Google Scholar] [CrossRef]
- Iglésias, S.P.; Lévy–Hartmann, L. Bathyraja leucomelanos, a new species of softnose skate (Chondrichthyes: Arhynchobatidae) from New Caledonia. Ichthyol. Res. 2012, 59, 38–48. [Google Scholar] [CrossRef]
- Cappetta, H. Handbook of Paleoichthyology, Chondrichthyes II: Mesozoic and Cenozoic Elasmobrachii: Teeth; Schultze, H.P., Ed.; Gustav Fischer Verlag: Munich, Germany, 2012; Volume 3E, 512p. [Google Scholar]
- Maisey, J.G. Cranial anatomy of Hybodus basanus Egerton from the Lower Cretaceous of England. Am. Mus. Novit. 1983, 2758, 1–64. [Google Scholar]
- Ebert, D.A.; Compagno, L.J. Chlamydoselachus africana, a new species of frilled shark from southern Africa (Chondrichthyes, Hexanchiformes, Chlamydoselachidae). Zootaxa 2009, 217, 1–18. [Google Scholar] [CrossRef]
- Soares, K.D.; Gomes, U.L.; Carvalho, M.R. Taxonomic review of catsharks of the Scyliorhinus haeckelii group, with the description of a new species (Chondrichthyes: Carcharhiniformes: Scyliorhinidae). Zootaxa 2016, 4066, 501–534. [Google Scholar] [CrossRef] [PubMed]
- Ebert, D.A.; White, W.T.; Ho, H.C. Redescription of Hexanchus nakamurai Teng 1962, (Chondrichthyes: Hexanchiformes: Hexanchidae), with designation of a neotype. Zootaxa 2013, 3752, 20–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmgren, N. Studies on the head in fishes. Embryological, morphological, and phylogenetical researches. Part I: Development of the skull in sharks and rays. Acta Zool. 1940, 21, 51–267. [Google Scholar] [CrossRef]
- Jollie, M. Some developmental aspects of the head skeleton of the 35–37 mm Squalus acanthias foetus. J. Morphol. 1971, 133, 17–40. [Google Scholar] [CrossRef] [PubMed]
- Maisey, J.G. An evaluation of jaw suspension in sharks. Am. Mus. Novit. 1980, 2706, 1–48. [Google Scholar]
- Wilga, C.A.; Ferry, L.A. Functional anatomy and biomechanics of feeding in elasmobranchs. In Fish Physiology; Shadwick, R.E., Farrell, A.P., Brauner, C.J., Eds.; Academic Press: London, UK, 2015; Volume 34B, pp. 153–187. [Google Scholar] [CrossRef]
- Maisey, J.G. The postorbital palatoquadrate articulation in elasmobranchs. J. Morphol. 2008, 269, 1022–1040. [Google Scholar] [CrossRef]
- Pradel, A.; Tafforeau, P.; Maisey, J.G.; Janvier, P. A new Paleozoic Symmoriiformes (Chondrichthyes) from the Late Carboniferous of Kansas (USA) and cladistic analysis of early chondrichthyans. PLoS ONE 2011, 6, e24938. [Google Scholar] [CrossRef]
- Holmgren, N. Studies on the head in fishes. Part 3. The phylogeny of elasmobranch fishes. Acta Zool. 1942, 23, 129–261. [Google Scholar] [CrossRef]
- De Carvalho, M.D.; Kriwet, J.; Thies, D. A systematic and anatomical revision of Late Jurassic angelsharks (Chondrichthyes: Squatinidae). In Mesozoic Fishes Homology and Phylogeny; Arratia, G., Schultze, H.P., Wilson, M.V.H., Eds.; Verlag Dr. Friedrich Pfeil: Munich, Germany, 2008; Volume 4, pp. 469–502. [Google Scholar]
- Maisey, J.G. Anatomical revision of the fossil shark Hybodus fraasi (Chondrichthyes, Elasmobranchii). Am. Mus. Novit. 1986, 2857, 1–16. [Google Scholar]
- De Carvalho, M.; Bockmann, F.A.; de Carvalho, M.R. Homology of the fifth epibranchial and accessory elements of the ceratobranchials among Gnathostomes: Insights from the development of ostariophysans. PLoS ONE 2013, 8, e62389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavin, L.; Cappetta, H.; Séret, B. Révision de Belemnobatis morinicus (Sauvage, 1873) du Portladien du Boulonnais (Pas-de-Calais, France). Comparaison avec quelques Rhinobatidae Jursassiques. Geol. Palaeontol. 1995, 29, 245–267. [Google Scholar]
- Miyake, T.; McEachran, J.D. The morphology and evolution of the ventral gill arch skeleton in batoid fishes (Chondrichthyes: Batoidea). Zool. J. Linn. Soc. 1991, 102, 75–100. [Google Scholar] [CrossRef]
- Johanson, Z.; Trinajstic, K.; Carr, R.; Ritchie, A. Evolution and development of the synarcual in early vertebrates. Zoomorphology 2013, 132, 95–110. [Google Scholar] [CrossRef]
- Claeson, K.M.; Hilger, A. Morphology of the anterior vertebral region in elasmobranchs: Special focus, Squatiniformes. Foss. Rec. 2011, 14, 129–140. [Google Scholar] [CrossRef]
- A Two-Headed Shark and Other X-Rayed Beauties at the Smithsonian. Smithsonian Magazine. Available online: https://www.smithsonianmag.com/science-nature/a-two-headed-shark-and-other-x-rayed-beauties-at-the-smithsonian-180952186 (accessed on 15 June 2019).
- Lane, J.A.; Maisey, J.G. Pectoral anatomy of Tribodus limae (Elasmobranchii: Hybodontiformes) from the Lower Cretaceous of northeastern Brazil. J. Vertebr. Paleontol. 2009, 29, 25–38. [Google Scholar] [CrossRef]
- Brito, P.; Seret, B. The new genus Iansan (Chondrichthyes, Rhinobatoidea) from the Early Cretaceous of Brazil and its phylogenetic relationships. In Mesozoic Fishes Systematics and Paleoecology; Arriatia, G., Viohl, G., Eds.; Verlag Dr. Friedrich Pfeil: Munuch, Germany, 1996; pp. 47–62. [Google Scholar]
- McEachran, J.D.; Dunn, K.A. Phylogenetic analysis of skates, a morphologically conservative clade of elasmobranchs (Chondrichthyes: Rajidae). Copeia 1998, 1998, 271–290. [Google Scholar] [CrossRef]
- Stumpf, S.; Etches, S.; Underwood, C.J.; Kriwet, J. Durnonovariaodus maiseyi gen. et sp. nov., a new hybodontiform shark-like chondrichthyan from the Upper Jurassic Kimmeridge Clay Formation of England. PeerJ 2021, 9, e11362. [Google Scholar] [CrossRef] [PubMed]
- Coates, M.I.; Gess, R.W. A new reconstruction of Onychoselache traquairi, comments on early chondrichthyan pectoral girdles and hybodontiform phylogeny. Palaeontology 2007, 50, 1421–1446. [Google Scholar] [CrossRef]
- Riley, C.; Cloutier, R.; Grogan, E.D. Similarity of morphological composition and developmental patterning in paired fins of the elephant shark. Sci. Rep. 2017, 7, 9985. [Google Scholar] [CrossRef] [Green Version]
- Hall, B.G. Phylogenetic Trees Made Easy: A How to Manual; Sinauer: Sunderland, MA, USA, 2011; 282p. [Google Scholar]
- Maisey, J.G.; Janvier, P.; Pradel, A.; Denton, J.S.S.; Bronson, A.; Miller, R.; Burrow, C.J. Doliodus and pucapampellids: Contrasting perspectives on stem chondrichthyan morphology. In Evolution and Development of Fishes; Johanson, Z., Underwood, C., Richter, M., Eds.; Cambridge University Press: Cambridge, UK, 2019; pp. 87–109. [Google Scholar]
- Nelson, J.S. Fishes of the World; A Wiley Interscience Publication John Wily and Sons: Hoboken, NJ, USA, 1984; 523p. [Google Scholar]
- Frey, L.; Coates, M.I.; Tietjen, K.; Rücklin, M.; Klug, C. A symmoriiform from the Late Devonian of Morocco demonstrates a derived jaw function in ancient chondrichthyans. Commun. Biol. 2019, 3, 681. [Google Scholar] [CrossRef]
- Brito, P.; Leal, M.; Gallo, V. A new lower Cretaceous guitarfish (Chondrichthyes, Batoidea) from the Santana formation, Northeastern Brazil. Bol. Mus. Nac. Geol. 2013, 75, 1–13. [Google Scholar]
- Brito, P.; Villalobos-Segura, E.; Alvarado-Ortega, J. A new early cretaceous guitarfish (Chondrichthyes, Batoidea) from the Tlayua Formation, Puebla, Mexico. J. S. Am. Earth Sci. 2019, 90, 155–161. [Google Scholar] [CrossRef]
- Villalobos-Segura, E.; Kriwet, J.; Vullo, R.; Stumpf, S.; Ward, D.J.; Underwood, C.J. The skeletal remains of the euryhaline sclerorhynchoid† Onchopristis (Elasmobranchii) from the “Mid”–Cretaceous and their palaeontological implications. Zool. J. Linn. Soc. 2021, 193, 746–771. [Google Scholar] [CrossRef]
- Strong, E.E.; Lipscomb, D. Character coding and inapplicable data. Cladistics 1999, 15, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Brazeau, M.D.; Guillerme, T.; Smith, M.R. An algorithm for morphological phylogenetic analysis with inapplicable data. Syst. Biol. 2019, 68, 619–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villalobos-Segura, E.; Underwood, C.J.; Ward, D.J. The first skeletal record of the enigmatic Cretaceous sawfish genus Ptychotrygon (Chondrichthyes, Batoidea) from the Turonian of Morocco. Pap. Palaeontol. 2021, 7, 353–376. [Google Scholar] [CrossRef]
- O’Reilly, J.E.; Puttick, M.N.; Parry, L.; Tanner, A.R.; Tarver, J.E.; Fleming, J.; Pisani, D.; Donoghue, P.C. Bayesian methods outperform parsimony but at the expense of precision in the estimation of phylogeny from discrete morphological data. Biol. Lett. 2016, 12, 20160081. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, J.E.; Puttick, M.N.; Pisani, D.; Donoghue, P.C. Probabilistic methods surpass parsimony when assessing clade support in phylogenetic analyses of discrete morphological data. Palaeontology 2018, 61, 105–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goloboff, P.A.; Torres, A.; Arias, J.S. Weighted parsimony outperforms other methods of phylogenetic inference under models appropriate for morphology. Cladistics 2018, 34, 407–437. [Google Scholar] [CrossRef]
- Goloboff, P.A.; Torres Galvis, A.; Arias Becerra, J.S. Parsimony and model–based phylogenetic methods for morphological data: Comments on O’Reilly et al. Paleontology 2018, 61, 1–6. [Google Scholar] [CrossRef]
- Sansom, R.S.; Choate, P.G.; Keating, J.N.; Randle, E. Parsimony, not Bayesian analysis, recovers more stratigraphically congruent phylogenetic trees. Biol. Lett. 2018, 14, 20180263. [Google Scholar] [CrossRef]
- Smith, M.R. Bayesian and parsimony approaches reconstruct informative trees from simulated morphological datasets. Biol. Lett. 2019, 15, 20180632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villalobos-Segura, E.; Underwood, C.J. Radiation and divergence times of Batoidea. J. Vertebr. Paleontol. 2020, 40, e1777147. [Google Scholar] [CrossRef]
- Lovejoy, N.R. Systematics of myliobatoid elasmobranchs: With emphasis on the phylogeny and historical biogeography of neotropical freshwater stingrays (Potamotrygonidae: Rajiformes). Zool. J. Linn. Soc. 1996, 117, 207–257. [Google Scholar] [CrossRef]
- Reif, W.E. Tooth enameloid as a taxonomic criterion: 1. A new eusalachian shark from the Rhaetic-Liassic boundary. Neues Jahrb. fur Geol. Palaontol. Monatshefte 1977, 1977, 565–576. [Google Scholar]
- Aschliman, N.C. Interrelationships of the durophagous stingrays (Batoidea: Myliobatidae). Environ. Biol. Fishes 2014, 97, 967–979. [Google Scholar] [CrossRef]
- Welten, M.; Smith, M.; Underwood, C.J.; Johanson, Z. Evolutionary origins and development of saw-teeth on the sawfish and sawshark rostrum (Elasmobranchii; Chondrichthyes). R. Soc. Open Sci. 2015, 2, 2–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herman, J.; Hovestadt–Euler, M.; Hovestadt, D.C.; Stehmann, M. Contributions to the study of the comparative morphology of teeth and other relevant ichthyodorulites in living supraspecific taxa of Chondrichthyan fishes. Part B: Batomorphii 4c: Order Rajiformes-Suborder Myliobatoidei-Superfamily Dasyatoidea-Family Dasyatidae-Subfamily Dasyatinae-Genus: Urobatis, Subfamily Potamotrygoninae-Genus: Paratrygon, Superfamily Plesiobatoidea-Family Plesiobatidae-Genus: Plesiobatis, Superfamily Myliobatoidea-Family Myliobatidae-Subfamily Myliobatinae-Genera: Aetobatus, Aetomylaeus, Myliobatis and Pteromylaeus, Subfamily Rhinopterinae–Genus: Rhinoptera and Subfamily Mobulinae—Genera: Manta and Mobula. Addendum 1 to 4a: Erratum to Genus Pteroplatytrygon. Bull. K. Belg. Inst. Natuurwetenschappen-Biol. 2000, 70, 5–67. [Google Scholar]
- Zangerl, R.; Case, G.R. Cobelodus aculeatus (Cope), an Anacanthous shark from Pennsylvanian black shales of North America. Paleontol. Abt. A 1976, 154, 107–157. [Google Scholar]
- Dean, M.N.; Summers, A.P.; Ferry, L.A. Very low pressures drive ventilatory flow in chimaeroid fishes. J. Morphol. 2011, 273, 461–479. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.F.; Cloutier, R.; Turner, S. The oldest articulated chondrichthyan from the Early Devonian period. Nature 2003, 425, 501–504. [Google Scholar] [CrossRef]
- Marramà, G.; Carnevale, G.; Naylor, G.J.; Varese, M.; Giusberti, L.; Kriwet, J. Anatomy, taxonomy and phylogeny of the Eocene guitarfishes from the Bolca Lagerstätten, Italy, provide new insights into the relationships of the Rhinopristiformes (Elasmobranchii: Batomorphii). Zool. J. Linn. Soc. 2020, 192, 1090–1110. [Google Scholar] [CrossRef]




















































Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villalobos-Segura, E.; Marramà, G.; Carnevale, G.; Claeson, K.M.; Underwood, C.J.; Naylor, G.J.P.; Kriwet, J. The Phylogeny of Rays and Skates (Chondrichthyes: Elasmobranchii) Based on Morphological Characters Revisited. Diversity 2022, 14, 456. https://doi.org/10.3390/d14060456
Villalobos-Segura E, Marramà G, Carnevale G, Claeson KM, Underwood CJ, Naylor GJP, Kriwet J. The Phylogeny of Rays and Skates (Chondrichthyes: Elasmobranchii) Based on Morphological Characters Revisited. Diversity. 2022; 14(6):456. https://doi.org/10.3390/d14060456
Chicago/Turabian StyleVillalobos-Segura, Eduardo, Giuseppe Marramà, Giorgio Carnevale, Kerin M. Claeson, Charlie J. Underwood, Gavin J. P. Naylor, and Jürgen Kriwet. 2022. "The Phylogeny of Rays and Skates (Chondrichthyes: Elasmobranchii) Based on Morphological Characters Revisited" Diversity 14, no. 6: 456. https://doi.org/10.3390/d14060456
APA StyleVillalobos-Segura, E., Marramà, G., Carnevale, G., Claeson, K. M., Underwood, C. J., Naylor, G. J. P., & Kriwet, J. (2022). The Phylogeny of Rays and Skates (Chondrichthyes: Elasmobranchii) Based on Morphological Characters Revisited. Diversity, 14(6), 456. https://doi.org/10.3390/d14060456