
How Trophic Conditions Affect Development of Fire Salamander (Salamandra salamandra) Larvae: Two Extreme Cases

 , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
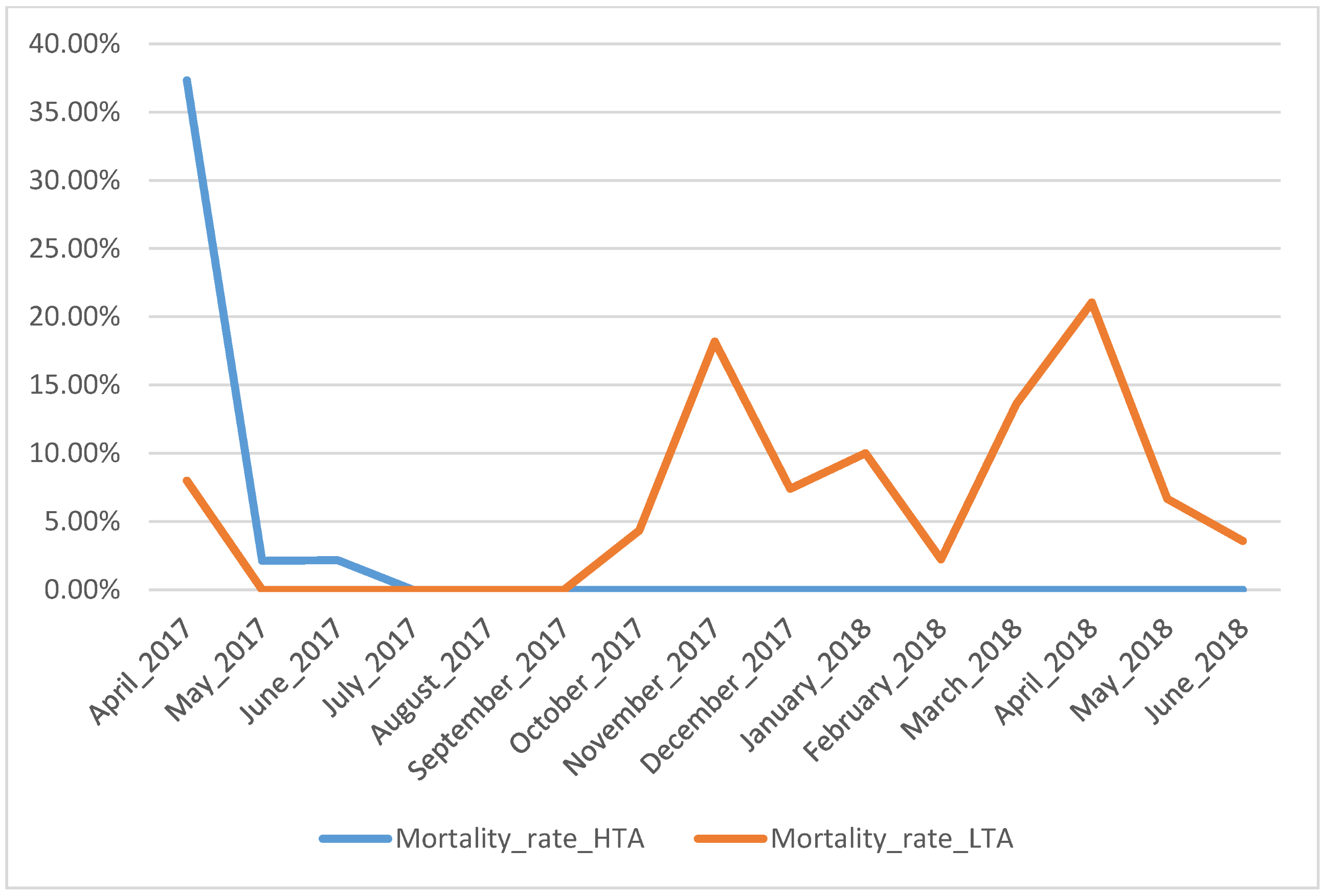
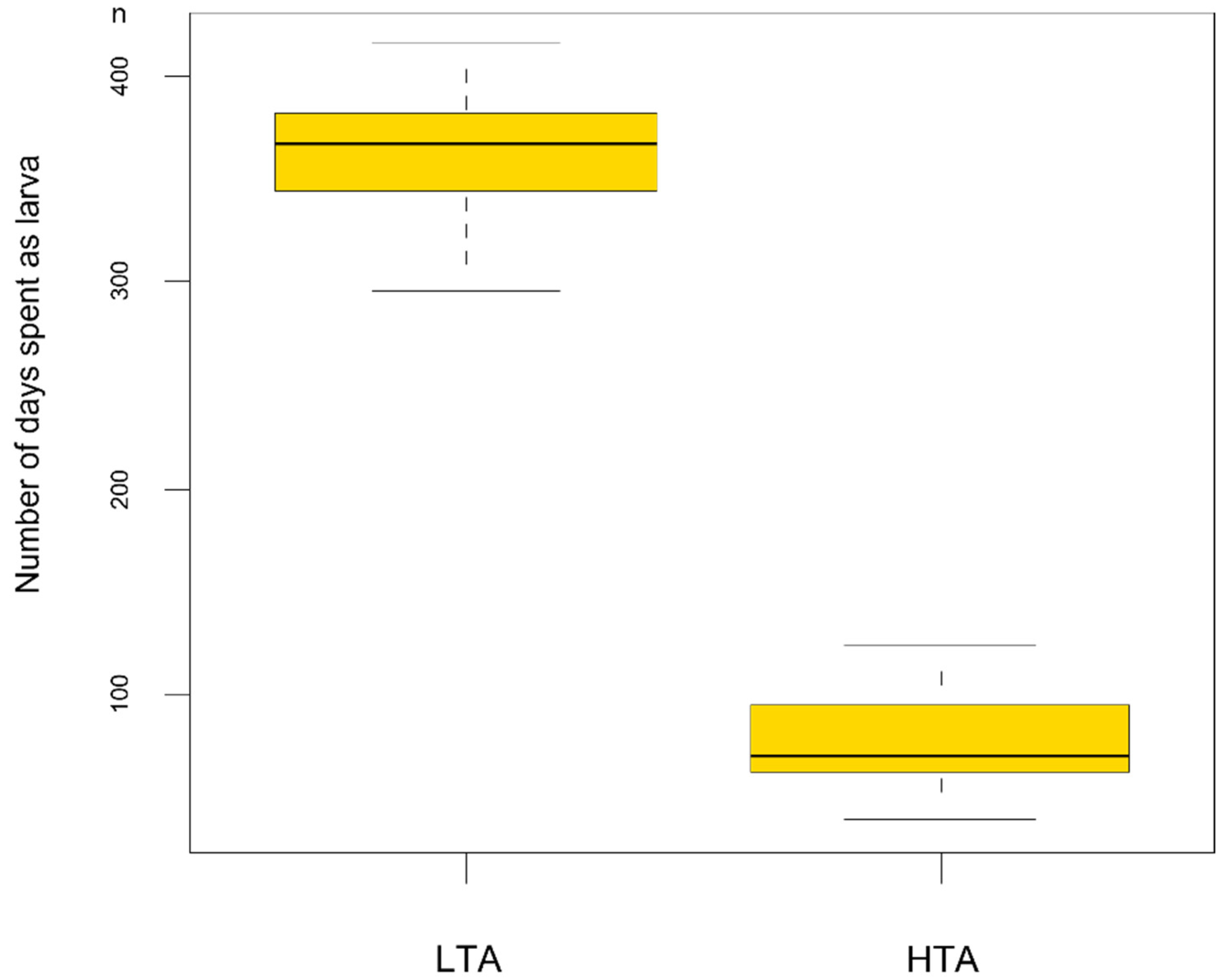
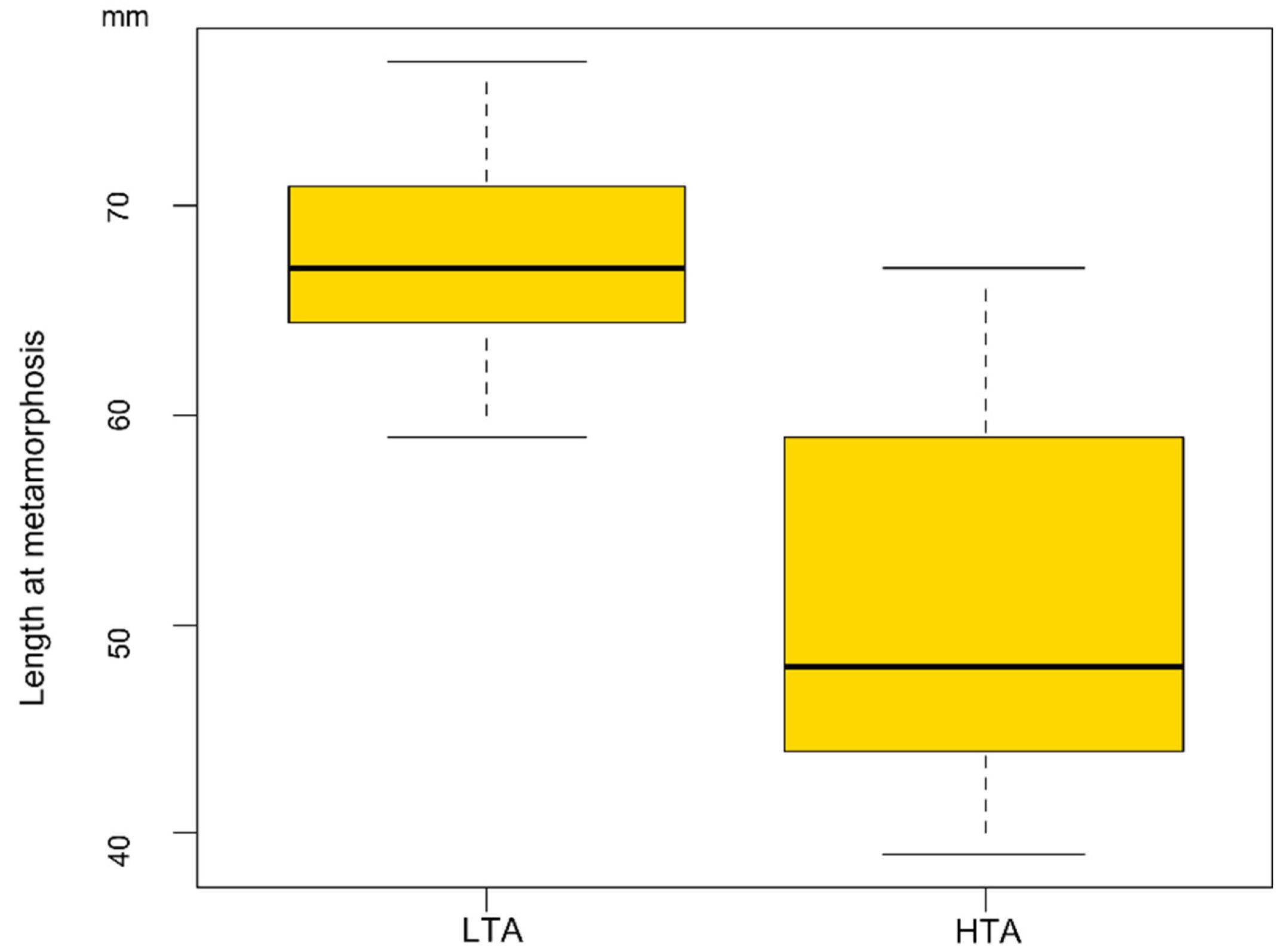
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moran, N.A. Adaptation and constraint in the complex life cycles of animals. Annu. Rev. Ecol. Evol. Syst. 1994, 25, 573–600. [Google Scholar] [CrossRef]
- Ficetola, G.F.; De Bernardi, F. Trade-off between larval development rate and post-metamorphic traits in the frog Rana latastei. Evol. Ecol. 2006, 20, 143–158. [Google Scholar] [CrossRef]
- Melotto, A.; Manenti, R.; Ficetola, G.F. Rapid adaptation to invasive predators overwhelms natural gradients of intraspecific variation. Nat. Commun. 2020, 11, 3608. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.S.; Monaghan, P.; Metcalfe, N.B. The trade-off between growth rate and locomotor performance varies with perceived time until breeding. J. Exp. Biol. 2010, 213, 3289–3298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, T.L.; Rowland, F.E.; Semlitsch, R.D. Variation in phenology and density differentially affects predator-prey interactions between salamanders. Oecologia 2017, 185, 475–486. [Google Scholar] [CrossRef]
- Reinhardt, T.; Steinfartz, S.; Paetzold, A.; Weitere, M. Linking the evolution of habitat choice to ecosystem functioning: Direct and indirect effects of pond-reproducing fire salamanders on aquatic-terrestrial subsidies. Oecologia 2013, 173, 281–291. [Google Scholar] [CrossRef]
- Rowland, F.E.; Rawlings, M.B.; Semlitsch, R.D. Joint effects of resources and amphibians on pond ecosystems. Oecologia 2017, 183, 237–247. [Google Scholar] [CrossRef]
- Benard, M.F. Warmer winters reduce frog fecundity and shift breeding phenology, which consequently alters larval development and metamorphic timing. Glob. Change Biol. 2015, 21, 1058–1065. [Google Scholar] [CrossRef]
- Bohenek, J.R.; Leary, C.J.; Resetarits, W.J. Exposure to glucocorticoids alters life history strategies in a facultatively paedomorphic salamander. J. Exp. Zool. Part A—Ecol. Integr. Physiol. 2021, 335, 329–338. [Google Scholar] [CrossRef]
- Dahl, E.; Orizaola, G.; Nicieza, A.G.; Laurila, A. Time constraints and flexibility of growth strategies: Geographic variation in catch-up growth responses in amphibian larvae. J. Anim. Ecol. 2012, 81, 1233–1243. [Google Scholar] [CrossRef]
- Van Buskirk, J. Amphibian phenotypic variation along a gradient in canopy cover: Species differences and plasticity. Oikos 2011, 120, 906–914. [Google Scholar] [CrossRef]
- Denoel, M.; Hervant, F.; Schabetsberger, R.; Joly, P. Short- and long-term advantages of an alternative ontogenetic pathway. Biol. J. Linn. Soc. 2002, 77, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Denoël, M.; Ficetola, G.F.; Cirovic, R.; Radovic, D.; Dzukic, G.; Kalezic, M.L.; Vukov, T.D. A multi-scale approach to facultative paedomorphosis of European newts (Salamandridae) in the Montenegrin karst: Distribution pattern, environmental variables, and conservation. Biol. Conserv. 2009, 142, 509–517. [Google Scholar] [CrossRef]
- Ringia, A.M.; Lips, K.R. Oviposition, early development and growth of the cave salamander, Eurycea lucifuga: Surface and subterranean influences on a troglophilic species. Herpetologica 2007, 63, 258–268. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Scali, S.; Denoel, M.; Montinaro, G.; Vukov, T.D.; Zuffi, M.A.L.; Padoa-Schioppa, E. Ecogeographical variation of body size in the newt Triturus carnifex: Comparing the hypotheses using an information-theoretic approach. Glob. Ecol. Biogeogr. 2010, 19, 485–495. [Google Scholar] [CrossRef]
- Denoël, M.; Ficetola, G.F. Using kernels and ecological niche modeling to delineate conservation areas in an endangered patch-breeding phenotype. Ecol. Appl. 2015, 25, 1922–1931. [Google Scholar] [CrossRef] [Green Version]
- Hendrix, R.; Hauswaldt, J.S.; Veith, M.; Steinfartz, S. Strong correlation between cross-amplification success and genetic distance across all members of ’True Salamanders’ (Amphibia: Salamandridae) revealed by Salamandra salamandra-specific microsatellite loci. Mol. Ecol. Resour. 2010, 10, 1038–1047. [Google Scholar] [CrossRef]
- Najbar, A.; Konowalik, A.; Halupka, K.; Najbar, B.; Ogielska, M. Body size and life history traits of the fire salamander Salamandra salamandra from Poland. Amphib. -Reptil. 2020, 41, 63–74. [Google Scholar] [CrossRef]
- Steinfartz, S.; Veith, M.; Tautz, D. Mitochondrial sequence analysis of Salamandra taxa suggests old splits of major lineages and postglacial recolonizations of Central Europe from distinct source populations of Salamandra salamandra. Mol. Ecol. 2000, 9, 397–410. [Google Scholar] [CrossRef]
- Caspers, B.A.; Junge, C.; Weitere, M.; Steinfartz, S. Habitat adaptation rather than genetic distance correlates with female preference in fire salamanders (Salamandra salamandra). Front. Zool. 2009, 6, 13. [Google Scholar] [CrossRef] [Green Version]
- Manenti, R.; Conti, A.; Pennati, R. Fire salamander (Salamandra salamandra) males’ activity during breeding season: Effects of microhabitat features and body size. Acta Herpetol. 2017, 12, 29–36. [Google Scholar]
- Balogova, M.; Jelic, D.; Kyselova, M.; Uhrin, M. Subterranean systems provide a suitable overwintering habitat for Salamandra salamandra. Int. J. Speleol. 2017, 46, 321–329. [Google Scholar] [CrossRef]
- Manenti, R.; Ficetola, G.F. Salamanders breeding in subterranean habitats: Local adaptations or behavioural plasticity? J. Zool. 2013, 289, 182–188. [Google Scholar] [CrossRef]
- Manenti, R.; De Bernardi, F.; Ficetola, G.F. Pastures vs forests: Do traditional pastoral activities negatively affect biodiversity? The case of amphibians communities. North-West. J. Zool. 2013, 9, 284–292. [Google Scholar]
- Manenti, R.; Siesa, M.E.; Ficetola, G.F. Odonata occurence in caves: Active or accidentals? A new case study. J. Cave Karst Stud. 2013, 75, 205–209. [Google Scholar] [CrossRef]
- Weitere, M.; Tautz, D.; Neumann, D.; Steinfartz, S. Adaptive divergence vs. environmental plasticity: Tracing local genetic adaptation of metamorphosis traits in salamanders. Mol. Ecol. 2004, 13, 1665–1677. [Google Scholar] [CrossRef]
- Buckley, D.; Alcobendas, M.; Garcia-Paris, M.; Wake, M.H. Heterochrony, cannibalism, and the evolution of viviparity in Salamandra salamandra. Evol. Dev. 2007, 9, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Caspers, B.A.; Steinfartz, S. Preference for the other sex: Olfactory sex recognition in terrestrial fire salamanders (Salamandra salamandra). Amphib. -Reptil. 2011, 32, 503–508. [Google Scholar] [CrossRef]
- Caspers, B.A.; Steinfartz, S.; Krause, E.T. Larval deposition behaviour and maternal investment of females reflect differential habitat adaptation in a genetically diverging salamander population. Behav. Ecol. Sociobiol. 2015, 69, 407–413. [Google Scholar] [CrossRef]
- Krause, E.T.; Steinfartz, S.; Caspers, B.A. Poor nutritional conditionsduring the early larval stage reduce risk-taking activities of fire salamander larvae (Salamandra salamandra). Ethology 2011, 117, 416–421. [Google Scholar] [CrossRef]
- Limongi, L.; Ficetola, G.F.; Romeo, G.; Manenti, R. Environmental factors determining growth of salamander larvae: A field study. Curr. Zool. 2015, 61, 421–427. [Google Scholar] [CrossRef] [Green Version]
- Melotto, A.; Ficetola, G.F.; Manenti, R. Safe as a cave? Intraspecific aggressiveness rises in predator-devoid and resource-depleted environments. Behav. Ecol. Sociobiol. 2019, 73, 68. [Google Scholar] [CrossRef]
- Berven, K.A.; Grudzen, T.A. Dispersal in the wood frog (Rana sylvatica): Implications for genetic population structure. Evolution 1990, 44, 2047–2056. [Google Scholar] [CrossRef] [PubMed]
- Ficetola, G.F.; Garner, T.W.J.; De Bernardi, F. Genetic diversity, but not hatching success, is jointly affected by postglacial colonization and isolation in the threatened frog. Mol. Ecol. 2007, 16, 3285. [Google Scholar] [CrossRef]
- Romeo, G.; Giovine, G.; Ficetola, G.F.; Manenti, R. Development of the fire salamander larvae at the altitudinal limit in Lombardy (north-western Italy): Effect of two cohorts occurrence on intraspecific aggression. North-West. J. Zool. 2015, 11, 234–240. [Google Scholar]
- Barzaghi, B.; Ficetola, G.F.; Pennati, R.; Manenti, R. Biphasic predators provide biomass subsidies in small freshwater habitats: A case study of spring and cave pools. Freshw. Biol. 2017, 62, 1637–1644. [Google Scholar] [CrossRef]
- Manenti, R.; Ficetola, G.F.; Marieni, A.; De Bernardi, F. Caves as breeding sites for Salamandra salamandra: Habitat selection, larval development and conservation issues. North-West. J. Zool. 2011, 7, 304–309. [Google Scholar]
- Manenti, R.; Ficetola, G.F.; Bianchi, B.; De Bernardi, F. Habitat features and distribution of Salamandra salamandra in underground springs. Acta Herpetol. 2009, 4, 143–151. [Google Scholar]
- Manenti, R.; Lunghi, E.; Ficetola, G.F. Cave exploitation by an usual epigean species: A review on the current knowledge on fire salamander breeding in cave. Biogeographia 2017, 32, 31–46. [Google Scholar] [CrossRef] [Green Version]
- Jusczcyk, W.; Zakrzewski, M. External morphology of larval stages of the spotted salamander Salamandra salamandra (L.). Acta Biol. Crac. 1981, 23, 127–135. [Google Scholar]
- Reilly, S.M. The ontogeny of aquatic feeding behavior in Salamandra salamandra: Stereotypy and isometry in feeding kinematics. J. Exp. Biol. 1995, 198, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Giovine, G. Analisi dello sviluppo e osservazioni sullo svernamento delle larve di Salamandra salamandra salamandra (L.) nei colli di Bergamo (lombardia). Nat. Brescia. 1996, 31, 263–269. [Google Scholar]
- Lanza, B.; Andreone, F.; Bologna, M.A.; Corti, C.; Razzetti, E.P.C. Fauna d’Italia, vol. XLII, Amphibia; Calderini: Bologna, Italy, 2007; Volume XLII. [Google Scholar]
- Lanza, B.; Nistri, A.; Vanni, S. Anfibi d’Italia; Ministero dell’Ambiente e della Tutela del Territorio e del Mare, I.S.P.R.A.; Grandi & Grandi Editori: Avignano sul Panaro, Italy, 2009.




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cogliati, P.; Barzaghi, B.; Melotto, A.; Ficetola, G.F.; Manenti, R. How Trophic Conditions Affect Development of Fire Salamander (Salamandra salamandra) Larvae: Two Extreme Cases. Diversity 2022, 14, 487. https://doi.org/10.3390/d14060487
Cogliati P, Barzaghi B, Melotto A, Ficetola GF, Manenti R. How Trophic Conditions Affect Development of Fire Salamander (Salamandra salamandra) Larvae: Two Extreme Cases. Diversity. 2022; 14(6):487. https://doi.org/10.3390/d14060487
Chicago/Turabian StyleCogliati, Paola, Benedetta Barzaghi, Andrea Melotto, Gentile Francesco Ficetola, and Raoul Manenti. 2022. "How Trophic Conditions Affect Development of Fire Salamander (Salamandra salamandra) Larvae: Two Extreme Cases" Diversity 14, no. 6: 487. https://doi.org/10.3390/d14060487
APA StyleCogliati, P., Barzaghi, B., Melotto, A., Ficetola, G. F., & Manenti, R. (2022). How Trophic Conditions Affect Development of Fire Salamander (Salamandra salamandra) Larvae: Two Extreme Cases. Diversity, 14(6), 487. https://doi.org/10.3390/d14060487