
Effects of the Climatic Region on Richness Correlations between Vascular Plants and Vertebrates in Nature Reserves of China


Abstract
:1. Introduction
2. Materials and Methods
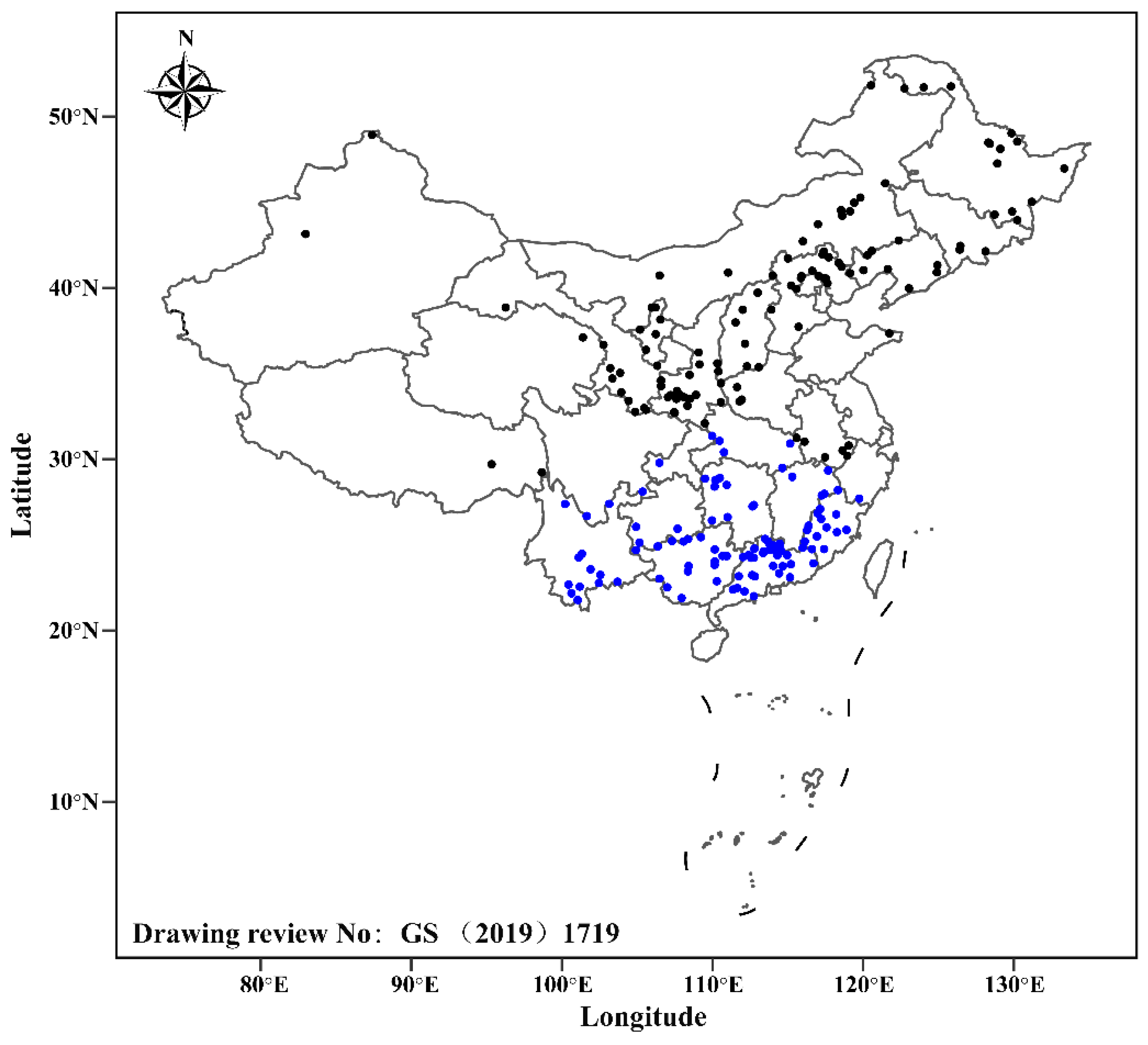
2.1. Study Area
2.2. Data Sources
- Monographs on national nature reserves [42];
- Scientific research reports on the nature reserves.
2.3. Spearman Rank Correlation Analysis
2.4. Collinearity Pre-Processing and Variables Standardization
2.5. Model Selection
3. Results
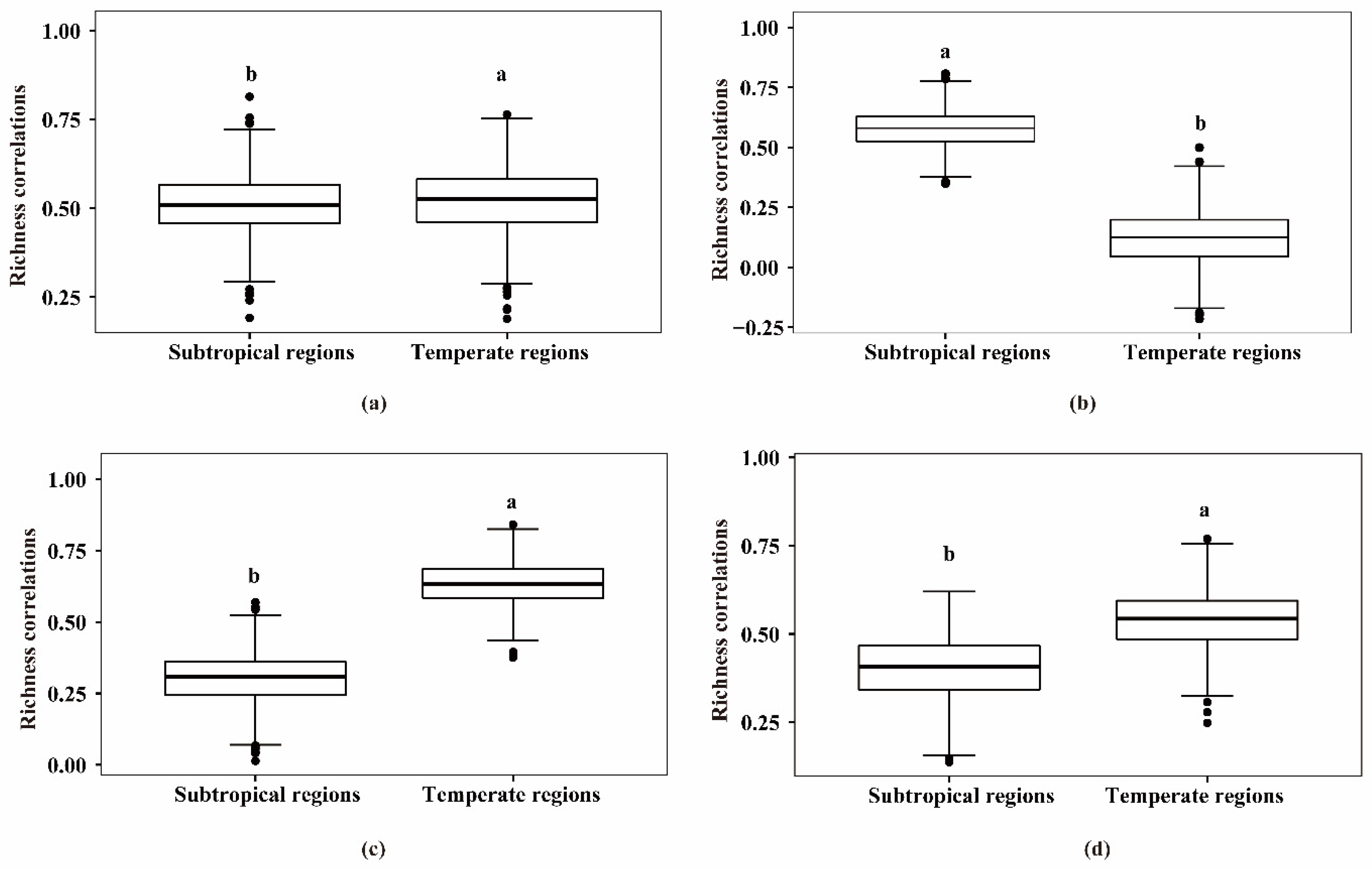
3.1. Richness Correlations between Vascular Plants and Vertebrates in Subtropical and Temperate Regions
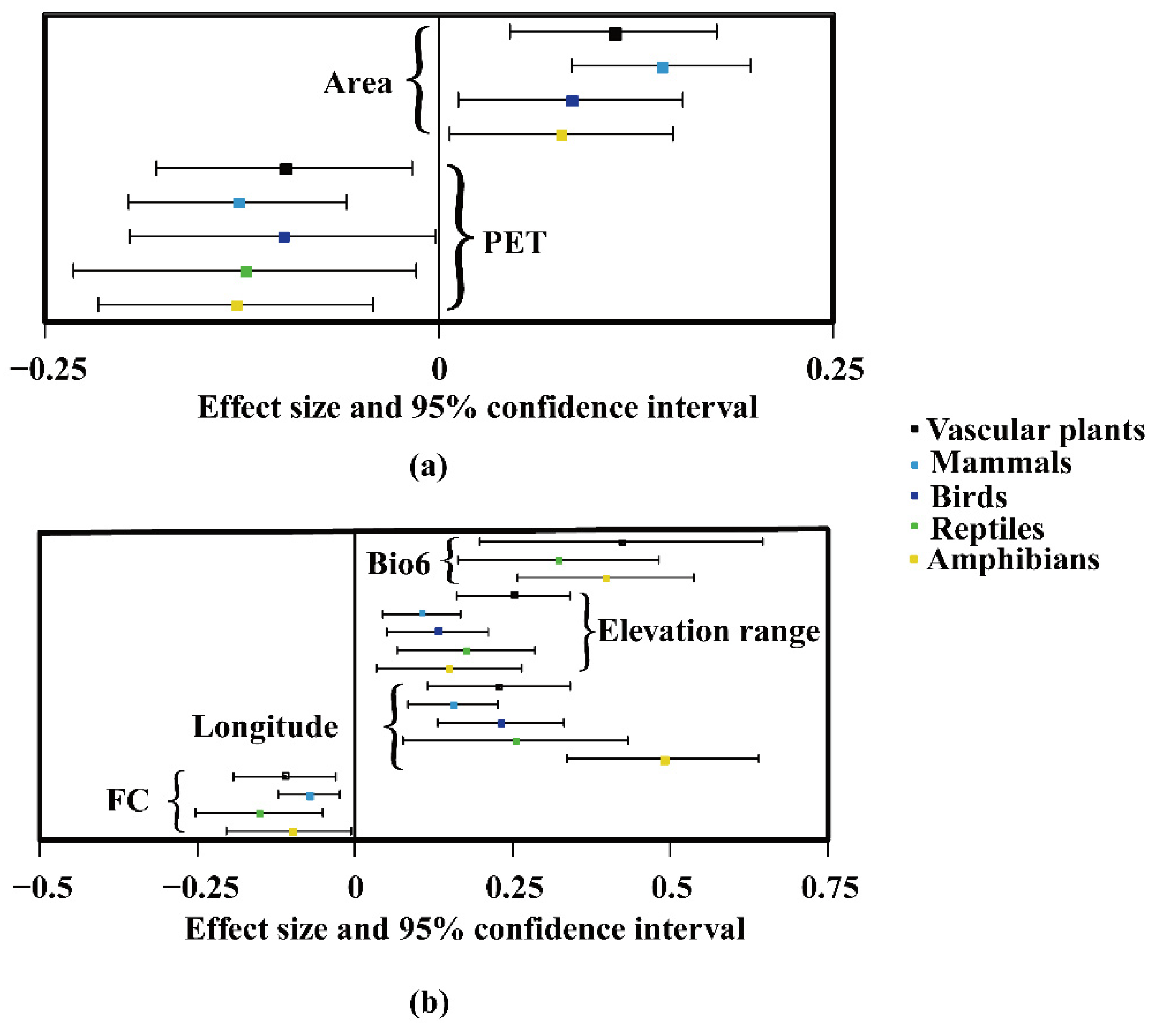
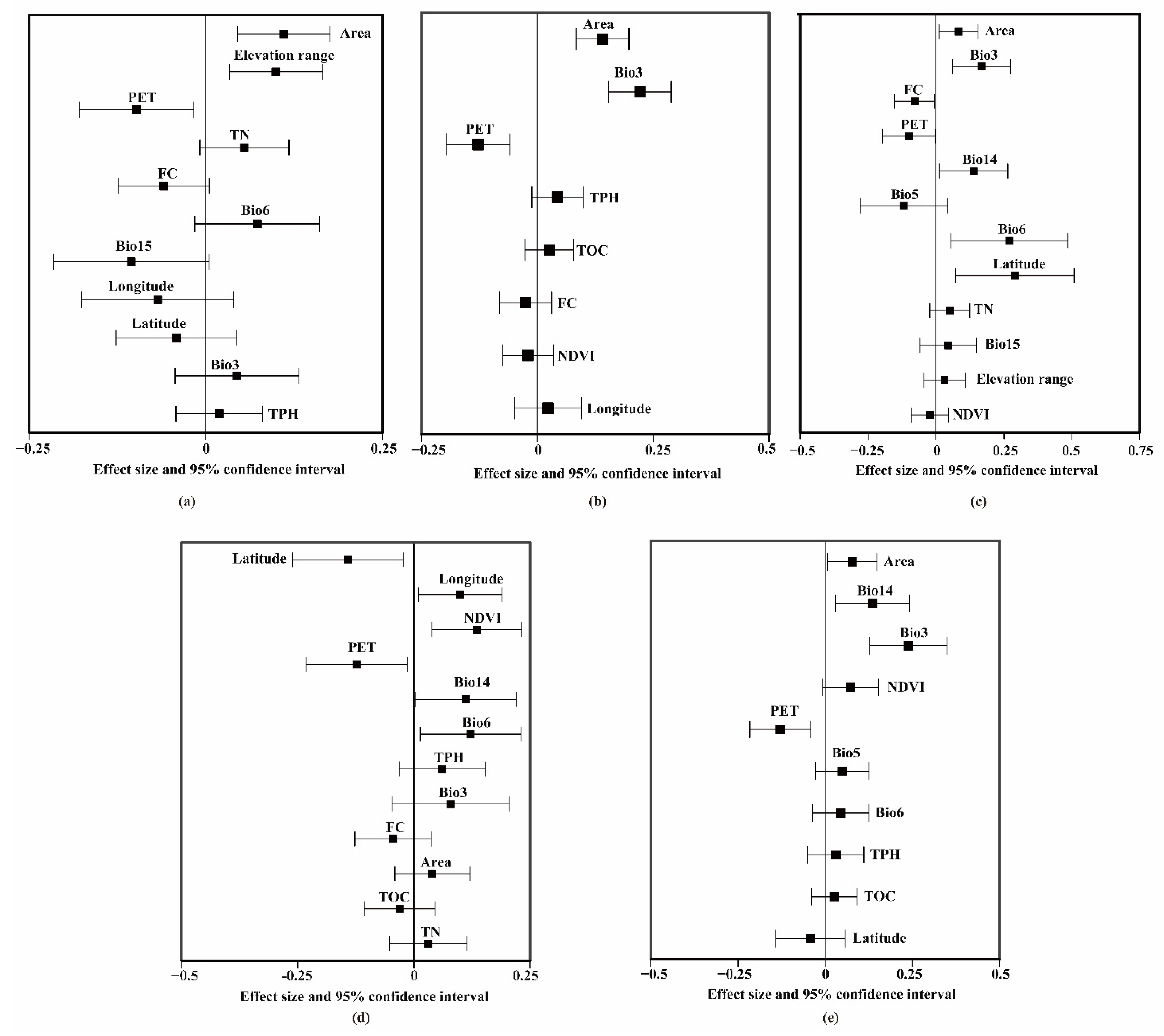
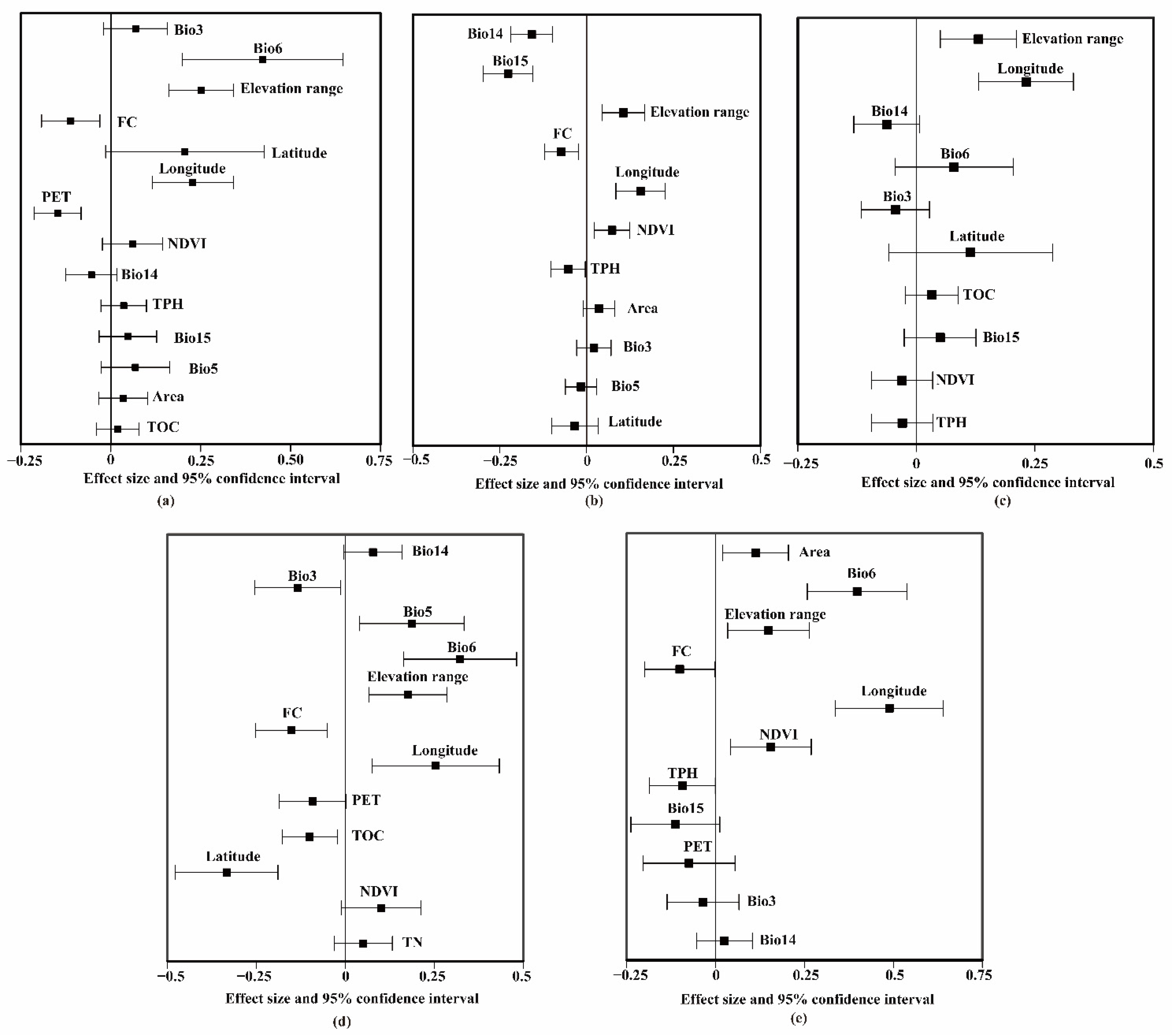
3.2. Common Environmental Factors Contributing to Richness Patterns of Vascular Plants and Vertebrates
- Subtropical regions (Figure 3a): Area for vascular plants (β = 0.11, p < 0.001) and mammals (β = 0.14, p < 0.001), and PET for vascular plants (β = −0.1, p < 0.001) and mammals (β = −0.13, p < 0.001).
- Temperate regions (Figure 3b): Elevation ranges for vascular plants (β = 0.25, p < 0.001) and mammals (β = 0.11, p < 0.001), longitude for vascular plants (β = 0.23, p < 0.001) and mammals (β = 0.16, p < 0.001), and FC for vascular plants (β = −0.11, p < 0.001) and mammals (β = −0.07, p < 0.001).
- Subtropical regions (Figure 3a): Area for vascular plants (β = 0.11, p < 0.001) and birds (β = 0.08, p < 0.001), and PET for vascular plants (β = −0.1, p < 0.001) and birds (β = −0.09, p < 0.001).
- Temperate regions (Figure 3b): Elevation range for vascular plants (β = 0.25, p < 0.001) and birds (β = 0.23, p < 0.001), and longitude for vascular plants (β = 0.23, p < 0.001) and birds (β = 0.23, p < 0.001).
- Subtropical regions (Figure 3a): PET for vascular plants (β = −0.1, p < 0.001) and reptiles (β = −0.12, p < 0.001).
- Temperate regions (Figure 3b): Bio6 for vascular plants (β = 0.42, p < 0.001) and reptiles (β = 0.32, p < 0.001), elevation range for vascular plants (β = 0.25, p < 0.001) and reptiles (β = 0.18, p <0.01), longitude for vascular plants (β = 0.23, p < 0.001) and reptiles (β = 0.25, p < 0.001), and FC for vascular plants (β = −0.11, p < 0.001) and reptiles (β = −0.15, p < 0.01).
- Subtropical regions (Figure 3a): Area for vascular plants (β = 0.11, p < 0.001) and amphibians (β = 0.08, p < 0.001), and PET for vascular plants (β = −0.1, p < 0.001) and amphibians (β = −0.12, p < 0.001).
- Temperate regions (Figure 3b): Bio6 for vascular plants (β = 0.42, p < 0.001) and amphibians (β = 0.40, p < 0.05), elevation range for vascular plants (β = 0.25, p < 0.001) and amphibians (β = 0.15, p < 0.05), longitude for vascular plants (β = 0.23, p < 0.001) and amphibians (β = 0.49, p < 0.001), and FC for vascular plants (β = −0.11, p < 0.001) and amphibians (β = −0.10, p < 0.05).
4. Discussion
4.1. Difference between Climatic Regions in the Effectiveness of Vascular Plants as Indicator Taxa
4.2. Ecological Mechanisms Promoting RCs between Vascular Plants and Vertebrates in Subtropical and Temperate Regions
4.3. Differences in Richness Correlations in Subtropical and Temperate Regions Caused by Abiotic Factors
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A


References
- Scheffers, B.R.; Joppa, L.N.; Pimm, S.L.; Laurance, W.F. What we know and don’t know about Earth’s missing biodiversity. Trends Ecol. Evol. 2012, 27, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Souza, J.L.P.; Baccaro, F.B.; Landeiro, V.L.; Franklin, E.; Magnusson, W.E.; Pequeno, P.A.C.L.; Fernandes, I.O. Taxonomic sufficiency and indicator taxa reduce sampling costs and increase monitoring effectiveness for ants. Divers. Distrib. 2016, 22, 111–122. [Google Scholar] [CrossRef]
- Howard, P.C.; Viskanic, P.; Davenport, T.R.B.; Kigenyi, F.W.; Baltzer, M.; Dickinson, C.J.; Lwanga, J.S.; Matthews, R.A.; Balmford, A. Complementarity and the use of indicator groups for reserve selection in Uganda. Nature 1998, 394, 472–475. [Google Scholar] [CrossRef]
- Caro, T.; Girling, S. Conservation by Proxy: Indicator, Umbrella, Keystone, Flagship, and Other Surrogate Species; The University of Chicago Press: Chicago, IL, USA, 2010. [Google Scholar]
- Santi, E.; Maccherini, S.; Rocchini, D.; Bonini, I.; Brunialti, G.; Favilli, L.; Perini, C.; Pezzo, F.; Piazzini, S.; Rota, E.; et al. Simple to sample: Vascular plants as surrogate group in a nature reserve. J. Nat. Conserv. 2010, 18, 2–11. [Google Scholar] [CrossRef]
- Hunter, M.; Westgate, M.; Barton, P.; Calhoun, A.; Pierson, J.; Tulloch, A.; Beger, M.; Branquinho, C.; Caro, T.; Gross, J.; et al. Two roles for ecological surrogacy: Indicator surrogates and management surrogates. Ecol. Indic. 2016, 63, 121–125. [Google Scholar] [CrossRef]
- Bal, P.; Tulloch, A.I.; Addison, P.F.; McDonald-Madden, E.; Rhodes, J.R. Selecting indicator species for biodiversity management. Front. Ecol. Environ. 2018, 16, 589–598. [Google Scholar] [CrossRef]
- de Morais, G.F.; dos Santos Ribas, L.G.; Ortega, J.C.G.; Heino, J.; Bini, L.M. Biological surrogates: A word of caution. Ecol. Indic. 2018, 88, 214–218. [Google Scholar] [CrossRef]
- Pharo, E.J.; Beattie, A.J.; Binns, D. Vascular plant diversity as a surrogate for bryophyte and lichen fiversity. Conserv. Biol. 1999, 13, 282–292. [Google Scholar] [CrossRef]
- Lovell, S.; Hamer, M.; Slotow, R.; Herbert, D. Assessment of congruency across invertebrate taxa and taxonomic levels to identify potential surrogates. Biol. Conserv. 2007, 139, 113–125. [Google Scholar] [CrossRef]
- Kirkman, L.K.; Smith, L.L.; Quintana-Ascencio, P.F.; Kaeser, M.J.; Golladay, S.W.; Farmer, A.L. Is species richness congruent among taxa? Surrogacy, complementarity, and environmental correlates among three disparate taxa in geographically isolated wetlands. Ecol. Indic. 2012, 18, 131–139. [Google Scholar] [CrossRef]
- Ruhí, A.; Batzer, D.P. Assessing congruence and surrogacy among wetland macroinvertebrate taxa towards efficiently measuring biodiversity. Wetlands 2014, 34, 1061–1071. [Google Scholar] [CrossRef]
- Larrieu, L.; Gosselin, F.; Archaux, F.; Chevalier, R.; Corriol, G.; Dauffy-Richard, E.; Deconchat, M.; Gosselin, M.; Ladet, S.; Savoie, J.-M.; et al. Cost-efficiency of cross-taxon surrogates in temperate forests. Ecol. Indic. 2018, 87, 56–65. [Google Scholar] [CrossRef]
- Zhang, K.; Lin, S.; Ji, Y.; Yang, C.; Wang, X.; Yang, C.; Wang, H.; Jiang, H.; Harrison, R.D.; Yu, D.W. Plant diversity accurately predicts insect diversity in two tropical landscapes. Mol. Ecol. 2016, 25, 4407–4419. [Google Scholar] [CrossRef] [PubMed]
- Brunbjerg, A.K.; Bruun, H.H.; Dalby, L.; Fløjgaard, C.; Frøslev, T.G.; Høye, T.T.; Goldberg, I.; Læssøe, T.; Hansen, M.D.D.; Brøndum, L.; et al. Vascular plant species richness and bioindication predict multi-taxon species richness. Methods Ecol. Evol. 2018, 9, 2372–2382. [Google Scholar] [CrossRef]
- Eglington, S.M.; Noble, D.G.; Fuller, R.J. A meta-analysis of spatial relationships in species richness across taxa: Birds as indicators of wider biodiversity in temperate regions. J. Nat. Conserv. 2012, 20, 301–309. [Google Scholar] [CrossRef]
- Gossner, M.M.; Fonseca, C.R.; Pašalić, E.; Türke, M.; Lange, M.; Weisser, W.W. Limitations to the use of arthropods as temperate forests indicators. Biodivers. Conserv. 2014, 23, 945–962. [Google Scholar] [CrossRef]
- Wolters, V.; Bengtsson, J.; Zaitsev, A. Relationship among species richness of different taxa. Ecology 2006, 87, 1886–1895. [Google Scholar] [CrossRef]
- Westgate, M.J.; Barton, P.S.; Lane, P.W.; Lindenmayer, D.B. Global meta-analysis reveals low consistency of biodiversity congruence relationships. Nat. Commun. 2014, 5, 3899. [Google Scholar] [CrossRef] [Green Version]
- Chave, J. The problem of pattern and scale in ecology: What have we learned in 20 years? Ecol. Lett. 2013, 16, 4–16. [Google Scholar] [CrossRef]
- Pearson, D.L.; Carroll, S.S. The Influence of spatial scale on cross-taxon congruence patterns and prediction accuracy of species richness. J. Biogeogr. 1999, 26, 1079–1090. [Google Scholar] [CrossRef]
- Burrascano, S.; de Andrade, R.B.; Paillet, Y.; Ódor, P.; Antonini, G.; Bouget, C.; Campagnaro, T.; Gosselin, F.; Janssen, P.; Persiani, A.M.; et al. Congruence across taxa and spatial scales: Are we asking too much of species data? Glob. Ecol. Biogeogr. 2018, 27, 980–990. [Google Scholar] [CrossRef]
- Qian, H.; Kissling, W.D. Spatial scale and cross-taxon congruence of terrestrial vertebrate and vascular plant species richness in China. Ecology 2010, 91, 1172–1183. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Cao, Y.; Cao, M.; Wu, J.; Wu, Y.; Le, Z.; Cui, P.; Li, J.; Ma, F.; Liu, L.; et al. Varying congruence among spatial patterns of vascular plants and vertebrates based on habitat groups. Ecol. Evol. 2017, 7, 8829–8840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillman, L.N.; Wright, S.D.; Cusens, J.; McBride, P.D.; Malhi, Y.; Whittaker, R.J. Latitude, productivity and species richness. Glob. Ecol. Biogeogr. 2015, 24, 107–117. [Google Scholar] [CrossRef]
- Willig, M.R.; Kaufman, D.M.; Stevens, R.D. Latitudinal gradients of biodiversity: Pattern, process, scale, and synthesis. Annu Rev. Ecol. Evol. Syst. 2003, 34, 273–309. [Google Scholar] [CrossRef]
- Hillebrand, H. On the generality of the latitudinal diversity gradient. Am. Nat. 2004, 163, 192–211. [Google Scholar] [CrossRef] [Green Version]
- Gaston, K.J. Biodiversity—Congruence. Prog. Phys. Geog. 1996, 20, 105–112. [Google Scholar] [CrossRef]
- Toranza, C.; Arim, M. Cross-taxon congruence and environmental conditions. BMC Ecol. 2010, 10, 18. [Google Scholar] [CrossRef] [Green Version]
- Duan, M.; Liu, Y.; Yu, Z.; Baudry, J.; Li, L.; Wang, C.; Axmacher, J.C. Disentangling effects of abiotic factors and biotic interactions on cross-taxon congruence in species turnover patterns of plants, moths and beetles. Sci. Rep. 2016, 6, 23511. [Google Scholar] [CrossRef] [Green Version]
- de Andrade, R.B.; Barlow, J.; Louzada, J.; Mestre, L.; Silveira, J.; Vaz-de-Mello, F.Z.; Cochrane, M.A. Biotic congruence in humid tropical forests: A multi-taxa examination of spatial distribution and responses to forest disturbance. Ecol. Indic. 2014, 36, 572–581. [Google Scholar] [CrossRef]
- Gao, J.; Liu, Y. Climate stability is more important than water–energy variables in shaping the elevational variation in species richness. Ecol. Evol. 2018, 8, 6872–6879. [Google Scholar] [CrossRef] [PubMed]
- Werneck, F.P.; Nogueira, C.; Colli, G.R.; Sites, J.W., Jr.; Costa, G.C. Climatic stability in the Brazilian Cerrado: Implications for biogeographical connections of south American savannas, species richness and conservation in a biodiversity hotspot. J. Biogeogr. 2012, 39, 1695–1706. [Google Scholar] [CrossRef]
- Pandey, B.; Khatiwada, J.R.; Zhang, L.; Pan, K.; Dakhil, M.A.; Xiong, Q.; Yadav, R.K.P.; Siwakoti, M.; Tariq, A.; Olatunji, O.A.; et al. Energy–water and seasonal variations in climate underlie the spatial distribution patterns of gymnosperm species richness in China. Ecol. Evol. 2020, 10, 9474–9485. [Google Scholar] [CrossRef] [PubMed]
- Peppler-Lisbach, C.; Kleyer, M. Patterns of species richness and turnover along the pH gradient in deciduous forests: Testing the continuum hypothesis. J. Veg. Sci. 2009, 20, 984–995. [Google Scholar] [CrossRef]
- Palpurina, S.; Wagner, V.; von Wehrden, H.; Hájek, M.; Horsák, M.; Brinkert, A.; Hölzel, N.; Wesche, K.; Kamp, J.; Hájková, P.; et al. The relationship between plant species richness and soil pH vanishes with increasing aridity across Eurasian dry grasslands. Glob. Ecol. Biogeogr. 2017, 26, 425–434. [Google Scholar] [CrossRef]
- Jordan, S.; Palmquist, K.; Bradford, J.; Lauenroth, W. Soil water availability shapes species richness in mid-latitude shrub steppe plant communities. J. Veg. Sci. 2020, 31, 646–657. [Google Scholar] [CrossRef]
- Chase, J.M.; Leibold, M.A. Spatial scale dictates the productivity–biodiversity relationship. Nature 2002, 416, 427–430. [Google Scholar] [CrossRef]
- Xu, J.; García Molinos, J.; Su, G.; Matsuzaki, S.-i.S.; Akasaka, M.; Zhang, H.; Heino, J. Cross-taxon congruence of multiple diversity facets of freshwater assemblages is determined by large-scale processes across China. Freshw. Biol. 2019, 64, 1492–1503. [Google Scholar] [CrossRef]
- Zhang, B.; Zhu, G. Climate Regionalization in China (First Draft); Science Press: Beijing, China, 1959. [Google Scholar]
- Administration, N.M. Climatological Atlas for the People’s Republic of China; Maps Press: Beijing, China, 1979. [Google Scholar]
- Wang, K. National Nature Reserves of China; Anhui Scientific and Technology Press: Hefei, China, 2003. [Google Scholar]
- Zhang, J. Nature Reserves in Guangdong Province; China Forestry Publishing House: Beijing, China, 2016. [Google Scholar]
- Tang, W. Nature Reserves in Guangxi Province; China Environmental Press: Beijing, China, 2014. [Google Scholar]
- Ge, J.; Wang, X. Nature Reserves in Hubei Province; Hubei Science and Technology Press: Wuhan, China, 2014. [Google Scholar]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Grueber, C.E.; Nakagawa, S.; Laws, R.J.; Jamieson, I.G. Multimodel inference in ecology and evolution: Challenges and solutions. J. Evol. Biol. 2011, 24, 699–711. [Google Scholar] [CrossRef]
- Peters, M.K.; Hemp, A.; Appelhans, T.; Behler, C.; Classen, A.; Detsch, F.; Ensslin, A.; Ferger, S.W.; Frederiksen, S.B.; Gebert, F.; et al. Predictors of elevational biodiversity gradients change from single taxa to the multi-taxa community level. Nat. Commun 2016, 7, 13736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagella, S. Does cross-taxon analysis show similarity in diversity patterns between vascular plants and bryophytes? some answers from a literature review. Comptes Rendus. Biol. 2014, 337, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Lamoreux, J.F.; Morrison, J.C.; Ricketts, T.H.; Olson, D.M.; Dinerstein, E.; McKnight, M.W.; Shugart, H.H. Global tests of biodiversity concordance and the importance of endemism. Nature 2006, 440, 212–214. [Google Scholar] [CrossRef]
- Qian, H. Relationships between plant and animal species richness at a regional scale in China. Conserv. Biol. 2007, 21, 937–944. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Fang, J.; Peng, C.; Tang, Z. Relationships between species richness of vascular plants and terrestrial vertebrates in China: Analyses based on data of nature reserves. Divers. Distrib. 2006, 12, 189–194. [Google Scholar] [CrossRef]
- Shinohara, N.; Yoshida, T. Why species richness of plants and herbivorous insects do or do not correlate. Ecol. Res. 2021, 36, 258–265. [Google Scholar] [CrossRef]
- Lewandowski, A.S.; Noss, R.F.; Parsons, D.R. The effectiveness of surrogate taxa for the representation of biodiversity. Conserv. Biol. 2010, 24, 1367–1377. [Google Scholar] [CrossRef]
- Gioria, M.; Bacaro, G.; Feehan, J. Evaluating and interpreting cross-taxon congruence: Potential pitfalls and solutions. Acta OEcol. 2011, 37, 187–194. [Google Scholar] [CrossRef]
- van Weerd, M.; Udo de Haes, H.A. Cross-taxon congruence in tree, bird and bat species distributions at a moderate spatial scale across four tropical forest types in the Philippines. Biodivers. Conserv. 2010, 19, 3393–3411. [Google Scholar] [CrossRef] [Green Version]
- Compton, T.J.; Rijkenberg, M.J.A.; Drent, J.; Piersma, T. Thermal tolerance ranges and climate variability: A comparison between bivalves from differing climates. J. Exp. Mar. Biol. Ecol. 2007, 352, 200–211. [Google Scholar] [CrossRef] [Green Version]
- Sunday, J.; Bennett, J.M.; Calosi, P.; Clusella-Trullas, S.; Gravel, S.; Hargreaves, A.L.; Leiva, F.P.; Verberk, W.C.E.P.; Olalla-Tárraga, M.Á.; Morales-Castilla, I. Thermal tolerance patterns across latitude and elevation. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20190036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, S.; Zhou, S.; Yin, X.; Zhang, C.; Li, R.; Chen, J.; Ma, D.; Wang, Y.; Yu, Z.; Chen, Y. Patterns of tree species richness in southwest China. Environ. Monit. Assess. 2021, 193, 97. [Google Scholar] [CrossRef]
- Moreno-Rueda, G.; Pizarro, M. Relative influence of habitat heterogeneity, climate, human disturbance, and spatial structure on vertebrate species richness in Spain. Ecol. Res. 2009, 24, 335–344. [Google Scholar] [CrossRef]
- Gillman, L.N.; Wright, S.D. Species richness and evolutionary speed: The influence of temperature, water and area. J. Biogeogr. 2014, 41, 39–51. [Google Scholar] [CrossRef]
- Stein, A.; Gerstner, K.; Kreft, H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecol. Lett. 2014, 17, 866–880. [Google Scholar] [CrossRef]
- Moser, D.; Dullinger, S.; Englisch, T.; Niklfeld, H.; Plutzar, C.; Sauberer, N.; Zechmeister, H.G.; Grabherr, G. Environmental determinants of vascular plant species richness in the Austrian Alps. J. Biogeogr. 2005, 32, 1117–1127. [Google Scholar] [CrossRef]
- Currie, D.J. Energy and large-scale patterns of animal- and plant-species richness. Am. Nat. 1991, 137, 27–49. [Google Scholar] [CrossRef]
- Qian, H.; Wang, X.; Wang, S.; Li, Y. Environmental determinants of amphibian and reptile species richness in China. Ecography 2007, 30, 471–482. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hypothesis | Variables |
|---|---|
| Geographic factors | Longitude |
| Latitude | |
| Habitat heterogeneity | Area |
| Elevation range | |
| Energy hypothesis | Max temperature of the warmest month (Bio5) |
| Min temperature of the coldest month (Bio6) | |
| Precipitation of the driest month (Bio14) | |
| Annual potential evapotranspiration (PET) | |
| Productivity hypothesis | Normalized difference vegetation index (NDVI) |
| Climatic stability | Isothermality (Bio3) |
| Precipitation seasonality (Bio15) | |
| Soil properties | Topsoil PH |
| Field capacity (FC) | |
| Topsoil organic carbon | |
| Total nitrogen density |
| Taxa | Climatic Regions | ρmean (±ρsd) | Pmean (±Psd) |
|---|---|---|---|
| Vascular plants–mammals | Subtropical regions | 0.51 (±0.084) | 1.95 × 10−3 (±0.008) |
| Vascular plants–mammals | Temperate regions | 0.52 (±0.089) | 2.24 × 10−3 (±0.011) |
| Vascular plants–birds | Subtropical regions | 0.58 (±0.075) | 2.13 × 10−4 (±0.001) |
| Vascular plants–birds | Temperate regions | 0.12 (±0.110) | 0.428 (±0.293) |
| Vascular plants–reptiles | Subtropical regions | 0.30 (±0.086) | 0.068 (±0.100) |
| Vascular plants–reptiles | Temperate regions | 0.63 (±0.073) | 4.86 × 10−5 (±0.001) |
| Vascular plants–amphibians | Subtropical regions | 0.40 (±0.089) | 0.02 (±0.004) |
| Vascular plants–amphibians | Temperate regions | 0.54 (±0.081) | 8.55 × 10−4 (±0.004) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, S.; Kong, F.; Hu, R.; Zhang, J.; Zhang, M. Effects of the Climatic Region on Richness Correlations between Vascular Plants and Vertebrates in Nature Reserves of China. Diversity 2022, 14, 499. https://doi.org/10.3390/d14060499
Jiang S, Kong F, Hu R, Zhang J, Zhang M. Effects of the Climatic Region on Richness Correlations between Vascular Plants and Vertebrates in Nature Reserves of China. Diversity. 2022; 14(6):499. https://doi.org/10.3390/d14060499
Chicago/Turabian StyleJiang, Sha, Fanhua Kong, Rui Hu, Jiaxin Zhang, and Minhua Zhang. 2022. "Effects of the Climatic Region on Richness Correlations between Vascular Plants and Vertebrates in Nature Reserves of China" Diversity 14, no. 6: 499. https://doi.org/10.3390/d14060499
APA StyleJiang, S., Kong, F., Hu, R., Zhang, J., & Zhang, M. (2022). Effects of the Climatic Region on Richness Correlations between Vascular Plants and Vertebrates in Nature Reserves of China. Diversity, 14(6), 499. https://doi.org/10.3390/d14060499