
Genome-Wide Screening for SNPs Associated with Stature in Diverse Cattle Breeds †


 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
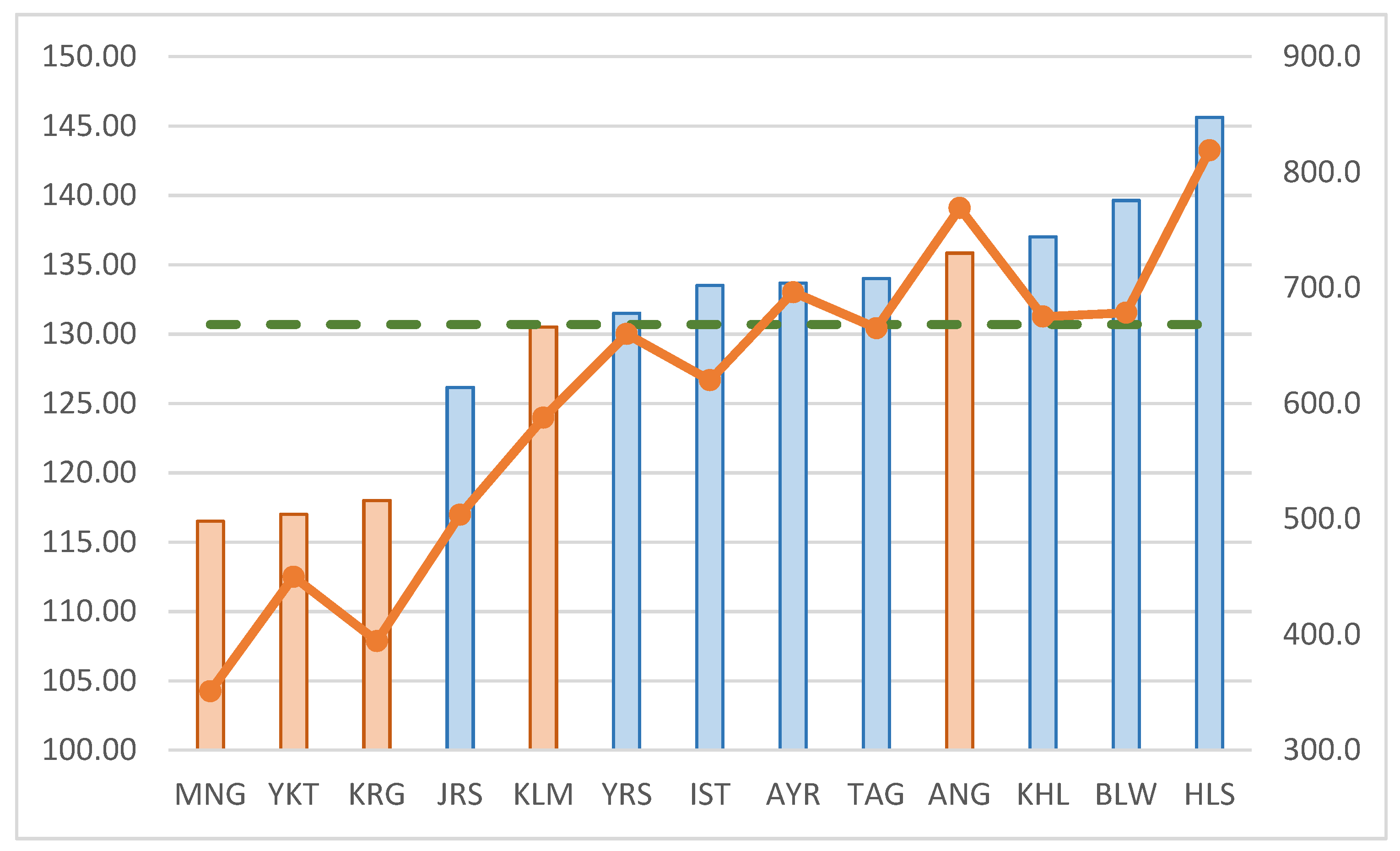
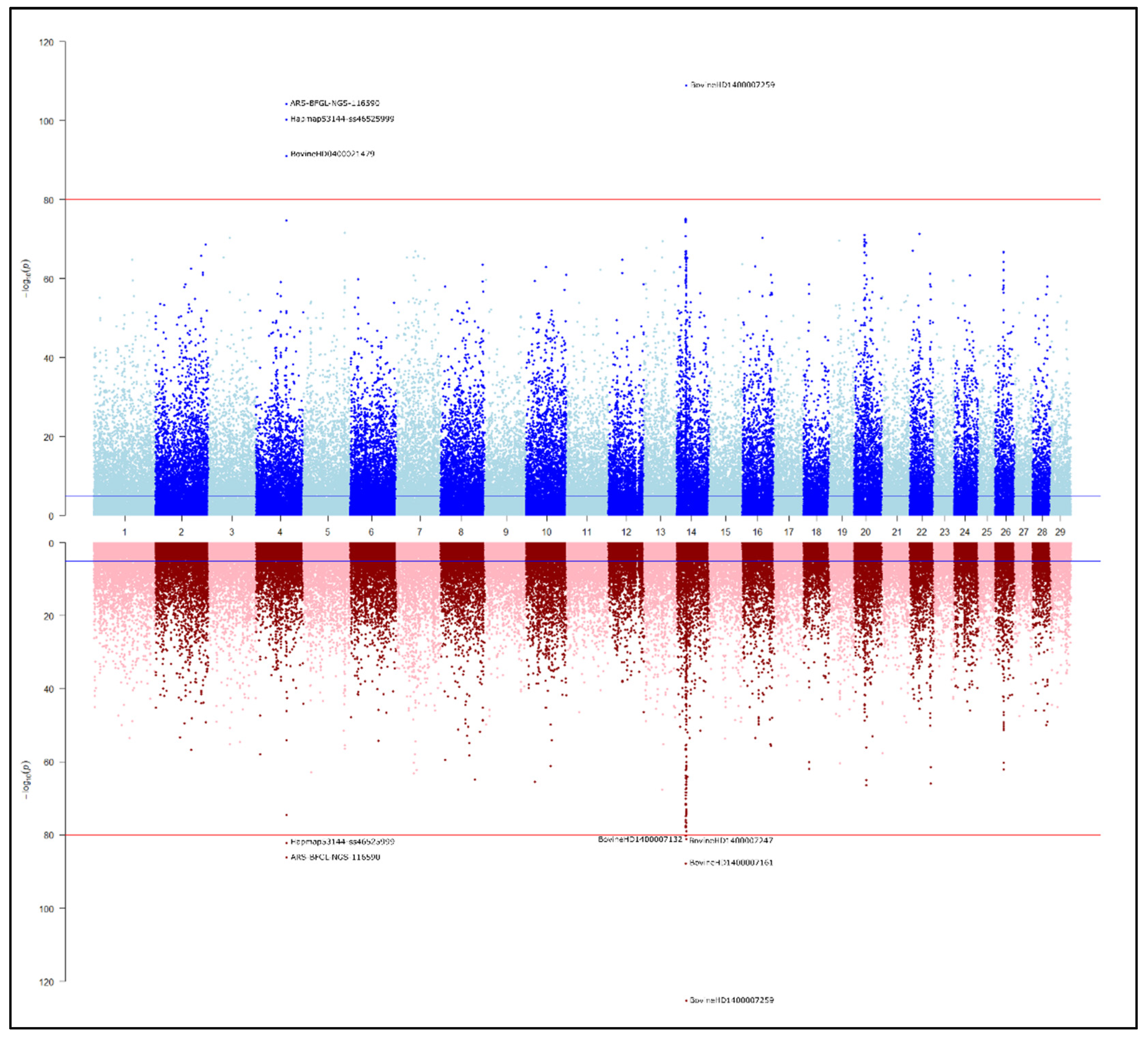
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kemper, K.E.; Visscher, P.M.; Goddard, M.E. Genetic architecture of body size in mammals. Genome Biol. 2012, 13, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galton, F. Regression towards mediocrity in hereditary stature. J. Anthropol. Inst. 1886, 15, 246–263. [Google Scholar] [CrossRef]
- Fink, T.; Tiplady, K.; Lopdell, T.; Johnson, T.; Snell, R.G.; Spelman, R.J.; Davis, S.R.; Littlejohn, M.D. Functional confirmation of PLAG1 as the candidate causative gene underlying major pleiotropic effects on body weight and milk characteristics. Sci. Rep. 2017, 21, 44793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brotherstone, S. Genetic and phenotypic correlations between linear type traits and production traits in Holstein-Friesian dairy cattle. Anim. Sci. 1994, 59, 183–187. [Google Scholar] [CrossRef]
- Bouwman, A.C.; Daetwyler, H.D.; Chamberlain, A.J.; Ponce, C.H.; Sargolzaei, M.; Schenkel, F.S.; Sahana, G.; Govignon-Gion, A.; Boitard, S.; Dolezal, M.; et al. Meta-analysis of genome-wide association studies for cattle stature identifies common genes that regulate body size in mammals. Nat. Genet. 2018, 50, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Takasuga, A. PLAG1 and NCAPG-LCORL in livestock. Anim. Sci. J. 2016, 87, 159–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purfield, D.C.; Evans, R.D.; Berry, D.P. Reaffirmation of known major genes and the identification of novel candidate genes associated with carcass-related metrics based on whole genome sequence within a large multi-breed cattle population. BMC Genom. 2019, 20, 720. [Google Scholar] [CrossRef]
- Zepeda-Batista, J.L.; Núñez-Domínguez, R.; Ramírez-Valverde, R.; Jahuey-Martínez, F.J.; Herrera-Ojeda, J.B.; Parra-Bracamonte, G.M. Discovering of Genomic Variations Associated to Growth Traits by GWAS in Braunvieh Cattle. Genes 2021, 12, 1666. [Google Scholar] [CrossRef]
- McClure, M.C.; Morsci, N.S.; Schnabel, R.D.; Kim, J.W.; Yao, P.; Rolf, M.M.; McKay, S.D.; Gregg, S.J.; Chapple, R.H.; Northcutt, S.L.; et al. A genome scan for quantitative trait loci influencing carcass, post-natal growth and reproductive traits in commercial Angus cattle. Anim. Genet. 2010, 41, 597–607. [Google Scholar] [CrossRef]
- Karim, L.; Takeda, H.; Lin, L.; Druet, T.; Arias, J.A.; Baurain, D.; Cambisano, N.; Davis, S.R.; Farnir, F.; Grisart, B.; et al. Variants modulating the expression of a chromosome domain encompassing PLAG1 influence bovine stature. Nat. Genet. 2011, 43, 405–413. [Google Scholar] [CrossRef]
- Littlejohn, M.; Grala, T.; Sanders, K.; Walker, C.; Waghorn, G.; Macdonald, K.; Coppieters, W.; Georges, M.; Spelman, R.; Hillerton, E. Genetic variation in PLAG1 associates with early life body weight and peripubertal weight and growth in Bos taurus. Anim. Genet. 2012, 43, 591–594. [Google Scholar] [CrossRef] [PubMed]
- Hoshiba, H.; Setoguchi, K.; Watanabe, T.; Kinoshita, A.; Mizoshita, K.; Sugimoto, Y.; Takasuga, A. Comparison of the effects explained by variations in the bovine PLAG1 and NCAPG genes on daily body weight gain, linear skeletal measurements and carcass traits in Japanese Black steers from a progeny testing program. Anim. Sci. J. 2013, 84, 529–534. [Google Scholar] [CrossRef] [PubMed]
- Naserkheil, M.; Bahrami, A.; Lee, D.; Mehrban, H. Integrating Single-Step GWAS and Bipartite Networks Reconstruction Provides Novel Insights into Yearling Weight and Carcass Traits in Hanwoo Beef Cattle. Animals 2020, 10, 1836. [Google Scholar] [CrossRef] [PubMed]
- Fortes, M.R.S.; Kemper, K.; Sasazaki, S.; Reverter, A.; Pryce, J.E.; Barendse, W.; Bunch, R.; McCulloch, R.; Harrison, B.; Bolormaa, S.; et al. Evidence for pleiotropism and recent selection in the PLAG1 region in Australian Beef cattle. Anim. Genet. 2013, 44, 636–647. [Google Scholar] [CrossRef]
- Randhawa, I.A.S.; Khatkar, M.S.; Thomson, P.C.; Raadsma, H.W. Composite selection signals for complex traits exemplified through bovine stature using multibreed cohorts of European and African Bos taurus. G3 Genes Genomes Genet. 2015, 5, 1391–1401. [Google Scholar] [CrossRef] [Green Version]
- Zimin, A.V.; Delcher, A.L.; Florea, L.; Kelley, D.R.; Schatz, M.C.; Puiu, D.; Hanrahan, F.; Pertea, G.; Van Tassell, C.P.; Sonstegard, T.S.; et al. A whole-genome assembly of the domestic cow, Bos taurus. Genome Biol. 2009, 10, R42. [Google Scholar] [CrossRef] [Green Version]
- Hensen, K.; Braem, C.; Declercq, J.; Van Dyck, F.; Dewerchin, M.; Fiette, L.; Denef, C.; Van de Ven, W.J. Targeted disruption of the murine Plag1 proto-oncogene causes growth retardation and reduced fertility. Dev. Growth Differ. 2004, 46, 459–470. [Google Scholar] [CrossRef]
- Wei, Z.; Wang, K.; Wu, H.; Wang, Z.; Pan, C.; Chen, H.; Lan, X. Detection of 15-bp Deletion Mutation within PLAG1 Gene and Its Effects on Growth Traits in Goats. Animals 2021, 11, 2064. [Google Scholar] [CrossRef]
- Voz, M.L.; Agten, N.S.; Van de Ven, W.J.; Kas, K. PLAG1, the main translocation target in pleomorphic adenoma of the salivary glands, is a positive regulator of IGF-II. Cancer Res. 2000, 60, 106–113. [Google Scholar]
- Van Dyck, F.; Declercq, J.; Braem, C.V.; Van de Ven, W.J.M. PLAG1, the prototype of the PLAG gene family: Versatility in tumour development (review). Int. J. Oncol. 2007, 30, 765–774. [Google Scholar] [CrossRef] [Green Version]
- De Vos, L.; Declercq, J.; Rosas, G.G.; Van Damme, B.; Roebroek, A.; Vermorken, F.; Ceuppens, J.; van de Ven, W.; Creemers, J. MMTV-cre-mediated fur inactivation concomitant with PLAG1 proto-oncogene activation delays salivary gland tumorigenesis in mice. Int. J. Oncol. 2008, 32, 1073–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeChiara, T.M.; Efstratiadis, A.; Robertson, E.J. A growth-deficiency phenotype in heterozygous mice carrying an insulin-like growth factor II gene disrupted by targeting. Nature 1990, 345, 78–80. [Google Scholar] [CrossRef] [PubMed]
- Boitard, S.; Boussaha, M.; Capitan, A.; Rocha, D.; Servin, B. Uncovering adaptation from sequence data: Lessons from genome resequencing of four cattle breeds. Genetics 2016, 203, 433–450. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Qu, K.; Jia, P.; Hanif, Q.; Zhang, J.; Chen, N.; Dang, R.; Chen, H.; Huang, B.; Lei, C. A SNP in PLAG1 is associated with body height trait in Chinese cattle. Anim. Genet. 2020, 51, 87–90. [Google Scholar] [CrossRef]
- Abdelmanova, A.S.; Kharzinova, V.R.; Volkova, V.V.; Mishina, A.I.; Dotsev, A.V.; Sermyagin, A.A.; Boronetskaya, O.I.; Petrikeeva, L.V.; Chinarov, R.Y.; Brem, G.; et al. Genetic diversity of historical and modern populations of Russian cattle breeds revealed by microsatellite analysis. Genes 2020, 11, 940. [Google Scholar] [CrossRef] [PubMed]
- Zinovieva, N.A.; Sermyagin, A.A.; Dotsev, A.V.; Boronetslaya, O.I.; Petrikeeva, L.V.; Abdelmanova, A.S.; Brem, G. Animal Genetic resources: Developing the research of allele pool of Russian cattle breeds—minireview. Sel’skokhozyaistvennaya Biol. Agric. Biol. 2019, 54, 631–641. [Google Scholar] [CrossRef] [Green Version]
- Sermyagin, A.A.; Dotsev, A.V.; Gladyr, E.A.; Traspov, A.A.; Deniskova, T.E.; Kostyunina, O.V.; Reyer, H.; Wimmers, K.; Barbato, M.; Paronyan, I.A.; et al. Whole-genome SNP analysis elucidates the genetic structure of Russian cattle and its relationship with Eurasian taurine breeds. Genet. Sel. Evol. 2018, 50, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. Gigascience 2015, 4, s13742-015. [Google Scholar] [CrossRef]
- Abdelmanova, A.S.; Sermyagin, A.A.; Dotsev, A.V.; Bardukov, N.V.; Fornara, M.S.; Brem, G.; Zinovieva, N.A. Genome-Wide Screening for SNPs Associated with Stature in Diverse Cattle Breeds. Biol. Life Sci. Forum 2022, 15, 24. [Google Scholar] [CrossRef]
- Lee, S.H.; Zhu, C.; Peng, Y.; Johnson, D.T.; Lehmann, L.; Sun, Z. Identification of a novel role of ZMIZ2 protein in regulating the activity of the Wnt/β-catenin signaling pathway. J. Biol. Chem. 2013, 288, 35913–35924. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, M.-P.; Ramayo-Caldas, Y.; Wolf, V.; Laithier, C.; El Jabri, M.; Michenet, A.; Boussaha, M.; Taussat, S.; Fritz, S.; Delacroix-Buchet, A.; et al. Sequence-based GWAS, network and pathway analyses reveal genes co-associated with milk cheese-making properties and milk composition in Montbéliarde cows. Genet. Sel. Evol. 2019, 51, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, M.P.; Govignon-Gion, A.; Croiseau, P.; Fritz, S.; Hozé, C.; Miranda, G.; Martin, P.; Barbat-Leterrier, A.; Letaïef, R.; Rocha, D.; et al. Within-breed and multi-breed GWAS on imputed whole-genome sequence variants reveal candi-date mutations affecting milk protein composition in dairy cattle. Genet. Sel. Evol. 2017, 49, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huo, N.; Yu, M.; Li, X.; Zhou, C.; Jin, X.; Gao, X. PURB is a positive regulator of amino acid-induced milk synthesis in bovine mammary epithelial cells. J. Cell. Physiol. 2019, 234, 6992–7003. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Miller, S.; Sargolzaei, M.; Kelly, M.; Vander Voort, G.; Caldwell, T.; Wang, Z.; Plastow, G.; Moore, S. Genome-wide association analyses for growth and feed efficiency traits in beef cattle. J. Anim. Sci. 2013, 91, 3612–3633. [Google Scholar] [CrossRef] [Green Version]
- Marete, A.G.; Guldbrandtsen, B.; Lund, M.S.; Fritz, S.; Sahana, G.; Boichard, D. A Meta-Analysis Including Pre-selected Sequence Variants Associated With Seven Traits in Three French Dairy Cattle Populations. Front. Genet. 2018, 9, 522. [Google Scholar] [CrossRef] [Green Version]
- Ghoreishifar, S.M.; Eriksson, S.; Johansson, A.M.; Khansefid, M.; Moghaddaszadeh-Ahrabi, S.; Parna, N.; Davoudi, P.; Javanmard, A. Signatures of selection reveal candidate genes involved in economic traits and cold acclimation in five Swedish cattle breeds. Genet. Sel. Evol. 2020, 52, 1–15. [Google Scholar] [CrossRef]
- Smith, J.L.; Wilson, M.L.; Nilson, S.M.; Rowan, T.N.; Oldeschulte, D.L.; Schnabel, R.D.; Decker, J.E.; Seabury, C.M. Genome-wide association and genotype by environment interactions for growth traits in U.S. Gelbvieh cattle. BMC Genom. 2019, 20, 926. [Google Scholar] [CrossRef] [Green Version]
- Saatchi, M.; Schnabel, R.D.; Taylor, J.F.; Garrick, D.J. Large-effect pleiotropic or closely linked QTL segregate within and across ten US cattle breeds. BMC Genom. 2014, 15, 442. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Choi, B.H.; Lim, D.; Gondro, C.; Cho, Y.M.; Dang, C.G.; Sharma, A.; Jang, G.W.; Lee, K.T.; Yoon, D.; et al. Genome-wide association study identifies major loci for carcass weight on BTA14 in Hanwoo (Korean Cattle). PLoS ONE 2013, 8, e74677. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, S.; Watanabe, T.; Mizoshita, K.; Tatsuda, K.; Fujita, T.; Watanabe, N.; Sugimoto, Y.; Takasuga, A. Genome-wide association study identified three major QTL for carcass weight including the PLAG1-CHCHD7 QTN for stature in Japanese Black cattle. BMC Genet. 2012, 13, 40. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Miao, J.; Song, Y.; Zhang, W.; Xu, L.; Chen, Y.; Zhang, L.; Gao, H.; Zhu, B.; Li, J.; et al. Genome-wide association study identifies the PLAG1-OXR1 region on BTA14 for carcass meat yield in cattle. Physiol. Genom. 2019, 51, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Rowan, T.N.; Schnabel, R.D.; Decker, J.E. Uncovering the architecture of selection in two Bos taurus cattle breeds. bioRxiv 2021. [Google Scholar] [CrossRef]
- Naserkheil, M.; Mehrban, H.; Lee, D.; Park, M.N. Genome-wide Association Study for Carcass Primal Cut Yields Using Single-step Bayesian Approach in Hanwoo Cattle. Front. Genet. 2021, 12, 752424. [Google Scholar] [CrossRef] [PubMed]
- Setoguchi, K.; Furuta, M.; Hirano, T.; Nagao, T.; Watanabe, T.; Sugimoto, Y.; Takasuga, A. Cross-breed comparisons identified a critical 591-kb region for bovine carcass weight QTL (CW-2) on chromosome 6 and the Ile-442-Met substitution in NCAPG as a positional candidate. BMC Genet. 2009, 10, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanvanhossou, S.; Scheper, C.; Dossa, L.H.; Yin, T.; Brügemann, K.; König, S. A multi-breed GWAS for morphometric traits in four Beninese indigenous cattle breeds reveals loci associated with conformation, carcass and adaptive traits. BMC Genom. 2020, 21, 783. [Google Scholar] [CrossRef]
- Doyle, J.L.; Berry, D.P.; Veerkamp, R.F.; Carthy, T.R.; Walsh, S.W.; Evans, R.D.; Purfield, D.C. Genomic regions associated with skeletal type traits in beef and dairy cattle are common to regions associated with carcass traits, feed intake and calving difficulty. Front. Genet. 2020, 11, 20. [Google Scholar] [CrossRef] [Green Version]
- Pryce, J.E.; Hayes, B.J.; Bolormaa, S.; Goddard, M.E. Polymorphic regions affecting human height also control stature in cattle. Genetics 2011, 187, 981–984. [Google Scholar] [CrossRef] [Green Version]
- An, B.; Xu, L.; Xia, J.; Wang, X.; Miao, J.; Chang, T.; Song, M.; Ni, J.; Xu, L.; Zhang, L.; et al. Multiple association analysis of loci and candidate genes that regulate body size at three growth stages in Simmental beef cattle. BMC Genet. 2020, 21, 32. [Google Scholar] [CrossRef] [Green Version]
- Abdelmanova, A.S.; Kharzinova, V.R.; Volkova, V.V.; Dotsev, A.V.; Sermyagin, A.A.; Chinarov, R.Y.; Zinovieva, N.A.; Boronetskaya, O.I.; Lutshikhina, E.M.; Sölkner, J.; et al. Comparative study of the genetic diversity of local steppe cattle breeds from Russia, Kazakhstan and Kyrgyzstan by microsatellite analysis of museum and modern samples. Diversity 2021, 13, 351. [Google Scholar] [CrossRef]
- Bown, M.D.; Muir, P.D.; Thomson, B.C. Dairy and beef breed effects on beef yield, beef quality and profitability: A review. New Zealand J. Agric. Res. 2016, 59, 74–184. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Localization | Trait | SNP | Allele | Allele Frequencies | |
|---|---|---|---|---|---|
| High-Stature Breeds | Low-Stature Breeds | ||||
| Chr 4 | Height | BovineHD0400021479 | T | 0.28 | 0.84 |
| C a | 0.72 | 0.16 | |||
| Height and Weight | ARS-BFGL-NGS-116590 | T | 0.29 | 0.92 | |
| G a,b | 0.71 | 0.08 | |||
| Hapmap53144-ss46525999 | A a,b | 0.71 | 0.09 | ||
| G | 0.29 | 0.91 | |||
| Chr 14 | BovineHD1400007259 | G a,b | 0.91 | 0.04 | |
| T | 0.09 | 0.96 | |||
| Weight | BovineHD1400007247 | A b | 0.95 | 0.45 | |
| G | 0.05 | 0.55 | |||
| BovineHD1400007132 | A b | 0.93 | 0.36 | ||
| G | 0.07 | 0.64 | |||
| BovineHD1400007161 | T b | 0.84 | 0.14 | ||
| G | 0.16 | 0.86 | |||
| Localization | SNP | Alleles | High-Stature Breeds | Low-Stature Breeds | ||
|---|---|---|---|---|---|---|
| Height | Weight | Height | Weight | |||
| Chr 4 | BovineHD0400021479 | T/C a | 0.708 ** | −0.498 ns | ||
| ARS-BFGL-NGS-116590 | T/G a | 0.722 ** | 0.216 ns | −0.172 ns | −0.233 ns | |
| Hapmap53144-ss46525999 | A a,b/G | 0.719 ** | 0.216 ns | −0.281 ns | −0.413 ns | |
| Chr 14 | BovineHD1400007259 | G a,b/T | 0.678 ** | 0.527 ns | −0.063 ns | −0.079 ns |
| BovineHD1400007247 | A b/G | 0.650 * | −0.062 ns | |||
| BovineHD1400007132 | A b/G | 0.584 * | −0.170 ns | |||
| BovineHD1400007161 | T b/G | 0.750 ** | −0.338 ns | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelmanova, A.S.; Sermyagin, A.A.; Dotsev, A.V.; Bardukov, N.V.; Fornara, M.S.; Brem, G.; Zinovieva, N.A. Genome-Wide Screening for SNPs Associated with Stature in Diverse Cattle Breeds. Diversity 2022, 14, 692. https://doi.org/10.3390/d14080692
Abdelmanova AS, Sermyagin AA, Dotsev AV, Bardukov NV, Fornara MS, Brem G, Zinovieva NA. Genome-Wide Screening for SNPs Associated with Stature in Diverse Cattle Breeds. Diversity. 2022; 14(8):692. https://doi.org/10.3390/d14080692
Chicago/Turabian StyleAbdelmanova, Alexandra S., Alexander A. Sermyagin, Arsen V. Dotsev, Nikolay V. Bardukov, Margaret S. Fornara, Gottfried Brem, and Natalia A. Zinovieva. 2022. "Genome-Wide Screening for SNPs Associated with Stature in Diverse Cattle Breeds" Diversity 14, no. 8: 692. https://doi.org/10.3390/d14080692
APA StyleAbdelmanova, A. S., Sermyagin, A. A., Dotsev, A. V., Bardukov, N. V., Fornara, M. S., Brem, G., & Zinovieva, N. A. (2022). Genome-Wide Screening for SNPs Associated with Stature in Diverse Cattle Breeds. Diversity, 14(8), 692. https://doi.org/10.3390/d14080692