
Photoautotrophic Euendoliths and Their Complex Ecological Effects in Marine Bioengineered Ecosystems

 , ,
, , 
Abstract
:1. Introduction
2. What Are Euendoliths and How Are They Observed?
- Epiliths that live on the surface of the substrate;
- Chasmoliths that adhere to the surface of fissures and cracks in the substrate;
- Cryptoendoliths that adhere to the surface of pre-existing cavities within porous rocks, including spaces produced and vacated by euendoliths, with no dissolution action;
- Euendoliths that actively penetrate carbonate (and phosphate) substrates and reside partially or completely inside cavities of their own making.
- Isolation of endoliths. After fixing the sample (substrate and euendoliths of interest) in formaldehyde solution to prevent structural damage to enclosed euendolithic organisms, the surrounding carbonate substrate is usually dissolved using dilute acid. The released endolithic filaments can then be observed and identified under light microscopy or transmission electron microscopy (TEM) [8]. As the hard matrix supporting the euendolithic filaments is dissolved, the organic components of the boring collapse, making spatial relationships and growth arrangements difficult to reconstruct. This technique only allows a qualitative assessment of euendolithic communities but is still widely used to detect and identify euendoliths of interest [15,48,49].
- In situ observations. Microborings can be studied in standard petrographic thin sections or sufficiently transparent fragments of shells using light microscopy [50] or scanning electron microscopy (SEM). The 2D visualization of microborings by petrographic thin sections is of limited use for the study of 3D objects, such as empty or filled euendolithic microborings, but allows their observation and the estimation of their true penetration depth in association with the surrounding matrix [42].
- Cast-embedding of microboring networks [8,42,51]. Combined embedding and casting in polymerized resins preserves the spatial arrangements of boring tunnels (3D architecture) and the euendolithic organisms in situ [51], allowing proper examination after the dissolution of the surrounding substrate (routine protocol in [8], see Figure 2 in [42]). Such casts can then be observed using light microscopy or SEM. However, this technique is limited when microborings are filled by secondary carbonate precipitation following the death of the euendoliths [42].
- Cultivation [48,52,53]. Natural samples (e.g., shell fragments, ooids) harboring euendoliths are used to inoculate agar plates containing a chemically defined culture medium. Once single colonies grow enough to be mechanically isolated, each colony is transferred into a liquid medium and stored under specific environmental conditions. While cultivation selects heavily for fast-growing microorganisms, it is a valuable tool to confirm taxonomic identification and to investigate the life history and physiology of euendoliths [44,45,54]. Cultivation also represents a necessary step to build the databases used for the identification of environmental DNA sequences [52].
- X-ray computed tomography (CT) and micro-computed tomography (micro-CT) (reviewed in [55]). These non-destructive 3D-visualization tools are firmly established for paleontological investigations, with their highest resolution encompassing the spatial magnitude of microborings [56,57,58]. They are, however, still relatively expensive and time-consuming, and require a high level of technical skill [55,58]. More affordable micro-CT and associated analyses can underestimate the extent of bioerosion by photoautotrophic euendoliths [57], as microborings range between 1 and 100 µm in diameter [32]. Micro-CT does, nonetheless, allow the investigation of microborings (empty or filled) within substrates that cannot be altered or dissolved (e.g., type material).
3. How Do Euendoliths Erode Calcium Carbonate?
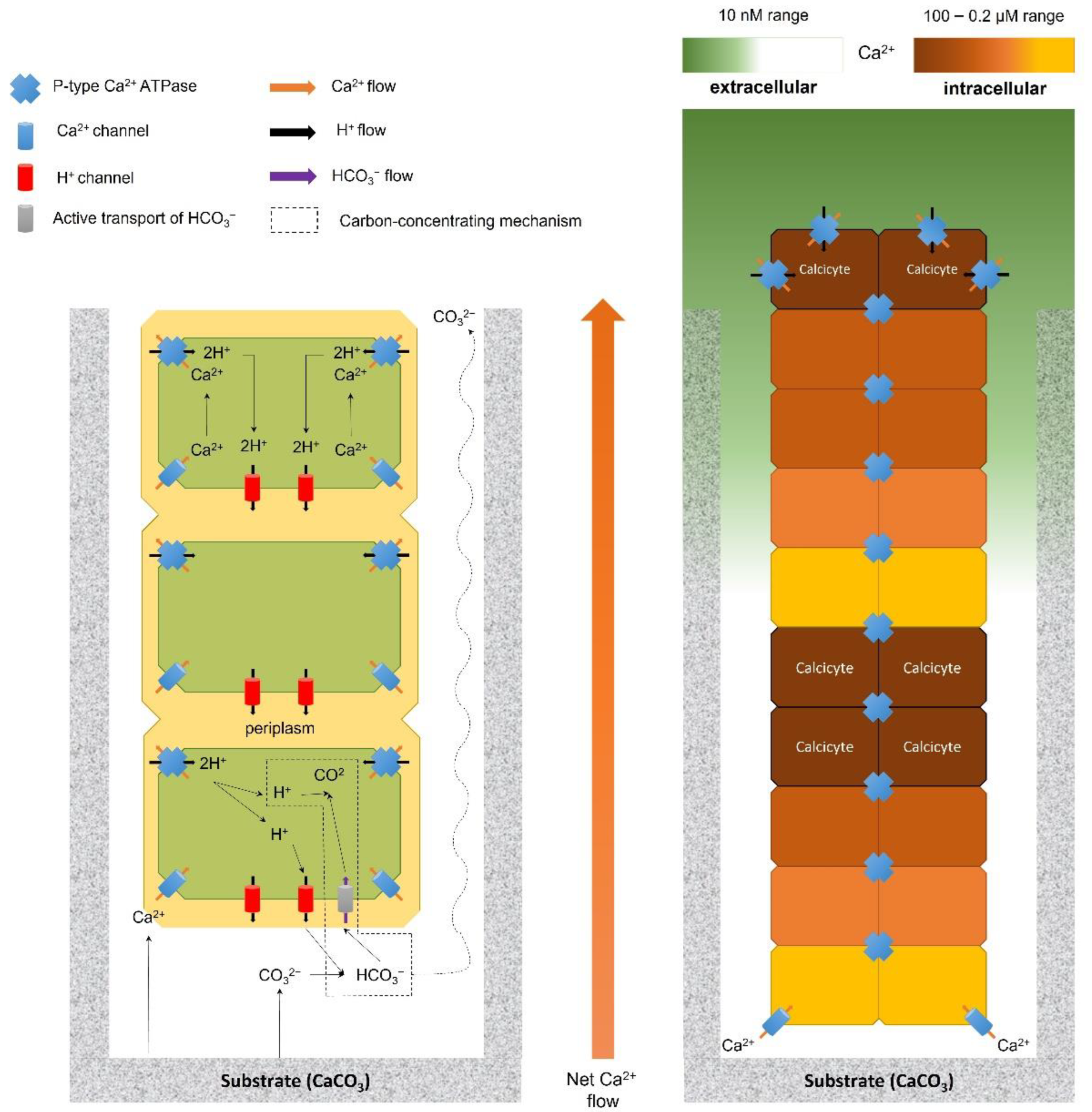
- The dissolution process is thermodynamically unfavorable, as it mainly occurs in waters saturated with calcium carbonate (i.e., calcite and aragonite). Excavation then becomes an ATP-driven active process with an energetic cost;
- The carbonate-dissolving mechanisms must be localized at the “head” of the microborer (i.e., apical cells) as dissolution produces true tunnels, with the typical “negative” shape of the borer [59];
- The proposed mechanisms must allow for the conservation of mass and electrical charge as in chemical reactions, the mass and the electrical charge of the components before the reaction must be equal to the mass and the electrical charge of the components after the reaction.

4. Incidence of Photoautotrophic Euendoliths in Marine Ecosystems
4.1. Light Availability
4.2. Nature of the Substrate
4.3. Biotic and Abiotic Environmental Factors
- Grazers are attracted to the substrate by the presence of photoautotrophic euendoliths, as these represent a renewable source of food [15,19,151]. The boring activity of euendoliths weakens the superficial layers of the substrate, which can facilitate the settlement of macroborers with their own bioerosive activity, as well as grazing;
- On the one hand, macrograzers constantly remove the superficial layers of the substrate, thus extending the depth to which the light can penetrate and, therefore, the depth to which the endoliths can bore, increasing microboring rates [150,152]. Grazing also reduces the settlement and growth of epilithic organisms that compete with euendoliths for space and diminish light availability [17]. On the other hand, macroborers excrete different waste products within the infested substrate, such as ammonium, phosphates, or CO2. Such waste products act as fertilizer for euendolithic communities, which increase in abundance, biomass, and productivity in the vicinity of macroborers [149,152,153].
4.4. Photoautotrophic Euendoliths as Bioindicators
5. Photoautotrophic Euendoliths in Marine Bioengineered Ecosystems
5.1. Corals and Crustose Coralline Algae
- Light transmission is affected by the architecture of the coral’s skeleton. Corallites on the top of coral colonies guide light deeper into the coral skeleton, while for corallites on the side of the colony, light enters at an angle, reducing its penetration into the skeleton [193];
- Water depth.
5.2. Bivalves
5.3. Other Groups
6. Photoautotrophic Euendoliths in the Anthropocene
7. Conclusions
- Update and standardize the taxonomy of known photoautotrophic euendolithic species, including morphological descriptions, ecological requirements, and molecular-based approaches to taxonomy;
- Determine the boring mechanisms for euendolithic species, such as red and green algae, and how these processes may be affected by environmental change, including OA and GCC;
- Investigate euendolithic communities (e.g., species composition, abundance, biomass), and the prevalence and severity of euendolithic infestation in less-investigated marine calcifiers (e.g., brachiopods, barnacles, limpets, snails), under different environmental conditions and timescales, and using a combination of approaches;
- Develop a standardized experimental framework, including ex situ and in situ experiments, to evaluate and compare the modalities of euendolithic infestation and how infestation may be influenced by various abiotic and biotic parameters (e.g., nutrient concentrations, light availability, presence of grazers and predators);
- Determine the detrimental and beneficial effects of euendolithic infestation on live marine calcifiers, using manipulative experiments when possible;
- Centralize available information on euendoliths (including heterotrophs) in a publicly accessible database (e.g., Ocean Acidification International Coordination Centre).
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carpenter, W. On the Microscopic Structure of Shells. Rep. Br. Assoc. Adv. Sci. 1845, 14, 1–24. [Google Scholar]
- Golubic, S.; Friedmann, I.; Schneider, J. The Lithobiontic Ecological Niche, with Special Reference to Microorganisms. J. Sediment. Petrol. 1981, 51, 475–478. [Google Scholar]
- Zhang, Y.; Golubic, S. Endolithic Microfossils (Cyanophyta) from Early Proterozoic Stromatolites, Hebei, China. Acta Micropalaeontologica Sin. 1987, 4, 1–3. [Google Scholar]
- Schneider, J.; Le Campion-Alsumard, T. Construction and Destruction of Carbonates by Marine and Freshwater Cyanobacteria. Eur. J. Phycol. 1999, 34, 417–426. [Google Scholar] [CrossRef]
- Knoll, A.H. Cyanobacteria and Earth History. In The Cyanobacteria: Molecular Biology, Genomics, and Evolution; Herrero, A., Flores, E., Eds.; Caister Academic Press: Poole, UK, 2008; pp. 1–19. ISBN 978-1-904455-15-8. [Google Scholar]
- Sánchez-Baracaldo, P.; Bianchini, G.; Wilson, J.D.; Knoll, A.H. Cyanobacteria and Biogeochemical Cycles through Earth History. Trends Microbiol. 2022, 30, 143–157. [Google Scholar] [CrossRef]
- Frémy, P. Contribution à La Physiologie Des Thallophytes Marines Perforant et Cariant Les Roches Calcaires et Les Coquilles. Ann. Inst. Océan. 1945, 22, 107–144. [Google Scholar]
- Golubic, S.; Perkins, R.D.; Lukas, K.J. Boring Microorganisms and Microborings in Carbonate Substrates. In The Study of Trace Fossils; Frey, R.W., Ed.; Springer: Berlin/Heidelberg, Germany, 1975; pp. 229–259. ISBN 978-3-642-65925-6. [Google Scholar]
- Friedmann, I.E.; Hua, M.; Ocampo-Friedmann, R. Cryptoendolithic Lichen and Cyanobacterial Communities of the Ross Desert, Antarctica. Polarforschung 1988, 58, 251–259. [Google Scholar] [PubMed]
- Golubic, S.; Schneider, J. Microbial Endoliths as Internal Biofilms. In Fossil and Recent Biofilms: A Natural History of Life on Earth; Krumbein, W.E., Paterson, D.M., Zavarzin, G.A., Eds.; Springer: Dordrecht, The Netherlands, 2003; pp. 249–263. ISBN 978-90-481-6412-7. [Google Scholar]
- Gaylarde, P.M.; Jungblut, A.-D.; Gaylarde, C.C.; Neilan, B.A. Endolithic Phototrophs from an Active Geothermal Region in New Zealand. Geomicrobiol. J. 2006, 23, 579–587. [Google Scholar] [CrossRef]
- Musso, B. Internal Bioerosion In Situ Living and Dead Corals on the Great Barrier Reef. Ph.D. Thesis, James Cook University, Townsville, Australia, 1994. [Google Scholar]
- Le Campion-Alsumard, T.; Golubic, S.; Hutchings, P. Microbial Endoliths in Skeletons of Live and Dead Corals: Porites Lobata (Moorea, French Polynesia). Mar. Ecol. Prog. Ser. 1995, 117, 149–157. [Google Scholar] [CrossRef]
- Kaehler, S. Incidence and Distribution of Phototrophic Shell-Degrading Endoliths of the Brown Mussel Perna Perna. Mar. Biol. 1999, 135, 505–514. [Google Scholar] [CrossRef]
- Tribollet, A.; Payri, C. Bioerosion of the Coralline Alga Hydrolithon Onkodes by Microborers in the Coral Reefs of Moorea, French Polynesia. Oceanol. Acta 2001, 24, 329–342. [Google Scholar] [CrossRef]
- Ghirardelli, L.A. Endolithic Microorganisms in Live and Dead Thalli of Coralline Red Algae (Corallinales, Rhodophyta) in the Northern Adriatic Sea. Acta Geol. Hisp. 2002, 37, 53–60. [Google Scholar]
- Golubic, S.; Schneider, J. Carbonate Dissolution. Stud. Environ. Sci. 1979, 3, 107–129. [Google Scholar]
- Kiene, W.E.; Hutchings, P.A. Bioerosion Experiments at Lizard Island, Great Barrier Reef. Coral Reefs 1994, 13, 91–98. [Google Scholar] [CrossRef]
- Chazottes, V.; Le Campion-Alsumard, T.; Peyrot-Clausade, M. Bioerosion Rates on Coral Reefs: Interactions between Macroborers, Microborers and Grazers (Moorea, French Polynesia). Palaeogeogr. Palaeoclimatol. Palaeoecol. 1995, 113, 189–198. [Google Scholar] [CrossRef]
- Glaub, I.; Vogel, K.; Gektidis, M. The Role of Modern and Fossil Cyanobacterial Borings in Bioerosion and Bathymetry. Ichnos 2001, 8, 185–195. [Google Scholar] [CrossRef]
- Laukner, G. Diseases of Mollusca: Bivalvia. In Diseases of Marine Animals; Kinne, O., Ed.; Biologische Anstalt Helgoland: Hamburg, Germany, 1983; Volume II, pp. 477–961. [Google Scholar]
- Kaehler, S.; McQuaid, C.D. Lethal and Sub-Lethal Effects of Phototrophic Endoliths Attacking the Shell of the Intertidal Mussel Perna Perna. Mar. Biol. 1999, 135, 497–503. [Google Scholar] [CrossRef]
- Fine, M.; Loya, Y. Endolithic Algae: An Alternative Source of Photoassimilates during Coral Bleaching. Proc. R. Soc. Lond. B Biol. Sci. 2002, 269, 1205–1210. [Google Scholar] [CrossRef] [PubMed]
- Hassenrück, C.; Jantzen, C.; Försterra, G.; Häussermann, V.; Willenz, P. Rates of Apical Septal Extension of Desmophyllum Dianthus: Effect of Association with Endolithic Photo-Autotrophs. Mar. Biol. 2013, 160, 2919–2927. [Google Scholar] [CrossRef]
- Zardi, G.I.; Nicastro, K.R.; McQuaid, C.D.; Ng, T.P.T.; Lathlean, J.; Seuront, L. Enemies with Benefits: Parasitic Endoliths Protect Mussels against Heat Stress. Sci. Rep. 2016, 6, 31413. [Google Scholar] [CrossRef] [PubMed]
- Crooks, J.A.; Khim, H.S. Architectural vs. Biological Effects of a Habitat-Altering, Exotic Mussel, Musculista Senhousia. J. Exp. Mar. Biol. Ecol. 1999, 240, 53–75. [Google Scholar] [CrossRef]
- Knowlton, N.; Brainard, R.E.; Fisher, R.; Moews, M.; Plaisance, L.; Caley, M.J. Coral Reef Biodiversity. In Life in the World’s Oceans; McIntyre, A.D., Ed.; Wiley-Blackwell: Oxford, UK, 2010; pp. 65–78. ISBN 978-1-4443-2550-8. [Google Scholar]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as Ecosystem Engineers. Oikos 1994, 69, 373. [Google Scholar] [CrossRef]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Positive and Negative Effects of Organisms as Physical Ecosystem Engineers. Ecology 1997, 78, 1946–1957. [Google Scholar] [CrossRef]
- Helmuth, B.; Mieszkowska, N.; Moore, P.; Hawkins, S.J. Living on the Edge of Two Changing Worlds: Forecasting the Responses of Rocky Intertidal Ecosystems to Climate Change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 373–404. [Google Scholar] [CrossRef]
- Wild, C.; Hoegh-Guldberg, O.; Naumann, M.S.; Colombo-Pallotta, M.F.; Ateweberhan, M.; Fitt, W.K.; Iglesias-Prieto, R.; Palmer, C.; Bythell, J.C.; Ortiz, J.-C.; et al. Climate Change Impedes Scleractinian Corals as Primary Reef Ecosystem Engineers. Mar. Freshw. Res. 2011, 62, 205. [Google Scholar] [CrossRef]
- Tribollet, A. The Boring Microflora in Modern Coral Reef Ecosystems: A Review of Its Roles. In Current Developments in Bioerosion; Wisshak, M., Tapanila, L., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 67–94. ISBN 978-3-540-77597-3. [Google Scholar]
- Lourenço, C.R.; Nicastro, K.R.; McQuaid, C.D.; Sabour, B.; Zardi, G.I. Latitudinal Incidence of Phototrophic Shell-Degrading Endoliths and Their Effects on Mussel Bed Microclimates. Mar. Biol. 2017, 164, 129–139. [Google Scholar] [CrossRef]
- Fedor, P.J.; Spellerberg, I.F. Shannon–Wiener Index. In Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2013; p. 4. ISBN 978-0-12-409548-9. [Google Scholar]
- Bornet, M.E.; Flahault, C. Sur Quelques Plantes Vivant Dans Le Test Calcaire Des Mollusques. Bull. Soc. Bot. Fr. 1889, 36, CXLVII–CLXXVI. [Google Scholar] [CrossRef]
- Kölliker, A.V. On the Frequent Occurrence of Vegetable Parasites in the Hard Structures of Animals. Proc. R. Soc. Lond. 1860, 10, 95–99. [Google Scholar] [CrossRef]
- Wedl, C. On the Significance of the Canals Found in Many Mollusc and Gastropod Shells. Sitzungsberichte Kais. Akad. Wiss. 1859, 33, 451–472. [Google Scholar]
- Gary, M.; McAfee, R.; Wolf, C.L. Glossary of Geology; American Geological Institute: Washington, WA, USA, 1973. [Google Scholar]
- Couradeau, E.; Roush, D.; Guida, B.S.; Garcia-Pichel, F. Diversity and Mineral Substrate Preference in Endolithic Microbial Communities from Marine Intertidal Outcrops (Isla de Mona, Puerto Rico). Biogeosciences 2017, 14, 311–324. [Google Scholar] [CrossRef]
- Amarelle, V.; Carrasco, V.; Fabiano, E. The Hidden Life of Antarctic Rocks. In The Ecological Role of Micro-organisms in the Antarctic Environment; Castro-Sowinski, S., Ed.; Springer Polar Sciences; Springer International Publishing: Cham, Switzerland, 2019; pp. 221–237. ISBN 978-3-030-02785-8. [Google Scholar]
- Ercegović, A. Études Écologiques et Sociologiques Des Cyanophycées Lithophytes de La Côte Yougoslave de l’Adriatique. Bull. Int. Acad. Yougosl. Sci. B-Arts 1932, 26, 33–56. [Google Scholar]
- Wisshak, M. Microbioerosion. In Developments in Sedimentology; Elsevier: Amsterdam, The Netherlands, 2012; Volume 64, pp. 213–243. ISBN 978-0-444-53813-0. [Google Scholar]
- Schroeder, J.H. Calcified Filaments of an Endolithic Alga in Recent Bermuda Reefs. Neues Jahrb. Geol. Palaontol. Mon. 1972, 1972, 16–33. [Google Scholar]
- Drew, K.M. Studies in the Bangioideae. III. The Life-History of Porphyra Umbilicalis (L.) Kütz. Var. Laciniata (Lighf.) J. Ag. A. The Conchocelis-Phase in Culture. Ann. Bot. 1954, XVIII, 184–209. [Google Scholar]
- Drew, K.M. Studies in the Bangiophycidae. IV. The Conchocelis-Phase of Bangia Fuscopurpurea (Dillw.) Lyngbye in Culture. Publ. Sta. Zool. Napoli. 1958, 30, 358–372. [Google Scholar]
- Lagerheim, G. Note Sur Le Mastigocoleus, Nouveau Genre Des Algues Marines de l’Ordre Des Phycochromacées. Notarisia 1886, 1, 65–69. [Google Scholar]
- Golubic, S.; Schneider, J.; Le Campion-Alsumard, T.; Campbell, S.E.; Hook, J.E.; Radtke, G. Approaching Microbial Bioerosion. Facies 2019, 65, 25. [Google Scholar] [CrossRef]
- Al-Thukair, A.A.; Golubic, S.; Rosen, G. New Euendolithic Cyanobacteria from the Bahama Bank and the Arabian Gulf: Hyella Racemus Sp. Nov. 1. J. Phycol. 1994, 30, 764–769. [Google Scholar] [CrossRef]
- Ndhlovu, A.; McQuaid, C.D.; Nicastro, K.R.; Zardi, G.I. Community Succession in Phototrophic Shell-Degrading Endoliths Attacking Intertidal Mussels. J. Molluscan Stud. 2021, 87, eyaa036. [Google Scholar] [CrossRef]
- Rooney, W.S.J.; Perkins, R.D. Distribution and Geologic Significance of Microboring Organisms within Sediments of the Arlington Reef Complex, Australia. Geol. Soc. Am. Bull. 1972, 83, 1139–1150. [Google Scholar] [CrossRef]
- Golubic, S.; Brent, G.; Le Campion, T. Scanning Electron Microscopy of Endolithic Algae and Fungi Using a Multipurpose Casting-embedding Technique. Lethaia 1970, 3, 203–209. [Google Scholar] [CrossRef]
- Chacón, E.; Berrendero, E.; Garcia-Pichel, F. Biogeological Signatures of Microboring Cyanobacterial Communities in Marine Carbonates from Cabo Rojo, Puerto Rico. Sediment. Geol. 2006, 185, 215–228. [Google Scholar] [CrossRef]
- Al-Thukair, A.A. Calculating Boring Rate of Endolithic Cyanobacteria Hyella Immanis under Laboratory Conditions. Int. Biodeterior. Biodegrad. 2011, 65, 664–667. [Google Scholar] [CrossRef]
- Pasella, M.M.; Lee, M.-F.E.; Marcelino, V.R.; Willis, A.; Verbruggen, H. Ten Ostreobium (Ulvophyceae) Strains from Great Barrier Reef Corals as a Resource for Algal Endolith Biology and Genomics. Phycologia 2022, 61, 452–458. [Google Scholar]
- Cunningham, J.A.; Rahman, I.A.; Lautenschlager, S.; Rayfield, E.J.; Donoghue, P.C.J. A Virtual World of Paleontology. Trends Ecol. Evol. 2014, 29, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Sutton, M.D. Tomographic Techniques for the Study of Exceptionally Preserved Fossils. Proc. R. Soc. B Biol. Sci. 2008, 275, 1587–1593. [Google Scholar] [CrossRef] [PubMed]
- Silbiger, N.; Guadayol, Ò.; Thomas, F.; Donahue, M. Reefs Shift from Net Accretion to Net Erosion along a Natural Environmental Gradient. Mar. Ecol. Prog. Ser. 2014, 515, 33–44. [Google Scholar] [CrossRef]
- Wisshak, M.; Titschack, J.; Kahl, W.-A.; Girod, P. Classical and New Bioerosion Trace Fossils in Cretaceous Belemnite Guards Characterised via Micro-CT. Foss. Rec. 2017, 20, 173–199. [Google Scholar] [CrossRef]
- Golubic, S. Distribution, Taxonomy, and Boring Patterns of Marine Endolithic Algae. Am. Zool. 1969, 9, 747–751. [Google Scholar] [CrossRef]
- Verbruggen, H. Morphological Complexity, Plasticity, and Species Diagnosability in the Application of Old Species Names in DNA-Based Taxonomies. J. Phycol. 2014, 50, 26–31. [Google Scholar] [CrossRef]
- Lukas, K.J. Two Species of the Chlorophyte Genus Ostreobium from Skeletons of Atlantic and Caribbean Reef Corals. J. Phycol. 1974, 10, 331–335. [Google Scholar]
- Verbruggen, H.; Ashworth, M.; LoDuca, S.T.; Vlaeminck, C.; Cocquyt, E.; Sauvage, T.; Zechman, F.W.; Littler, D.S.; Littler, M.M.; Leliaert, F. A Multi-Locus Time-Calibrated Phylogeny of the Siphonous Green Algae. Mol. Phylogenet. Evol. 2009, 50, 642–653. [Google Scholar] [CrossRef] [PubMed]
- Gutner-Hoch, E.; Fine, M. Genotypic Diversity and Distribution of Ostreobium Quekettii within Scleractinian Corals. Coral Reefs 2011, 30, 643–650. [Google Scholar] [CrossRef]
- Marcelino, V.R.; Verbruggen, H. Multi-Marker Metabarcoding of Coral Skeletons Reveals a Rich Microbiome and Diverse Evolutionary Origins of Endolithic Algae. Sci. Rep. 2016, 6, 31508. [Google Scholar] [CrossRef]
- Sauvage, T.; Schmidt, W.E.; Suda, S.; Fredericq, S. A Metabarcoding Framework for Facilitated Survey of Endolithic Phototrophs with TufA. BMC Ecol. 2016, 16, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Zapata, F.L.; Gómez-Osorio, S.; Sánchez, J.A. Conspicuous Endolithic Algal Associations in a Mesophotic Reef-Building Coral. Coral Reefs 2018, 37, 705–709. [Google Scholar] [CrossRef]
- Ricci, F.; Fordyce, A.; Leggat, W.; Blackall, L.L.; Ainsworth, T.; Verbruggen, H. Multiple Techniques Point to Oxygenic Phototrophs Dominating the Isopora Palifera Skeletal Microbiome. Coral Reefs 2021, 40, 275–282. [Google Scholar] [CrossRef]
- Taberlet, P.; Coissac, E.; Pompanon, F.; Brochmann, C.; Willerslev, E. Towards Next-Generation Biodiversity Assessment Using DNA Metabarcoding. Mol. Ecol. 2012, 21, 2045–2050. [Google Scholar] [CrossRef]
- Yang, S.-H.; Tandon, K.; Lu, C.-Y.; Wada, N.; Shih, C.-J.; Hsiao, S.S.-Y.; Jane, W.-N.; Lee, T.-C.; Yang, C.-M.; Liu, C.-T.; et al. Metagenomic, Phylogenetic, and Functional Characterization of Predominant Endolithic Green Sulfur Bacteria in the Coral Isopora Palifera. Microbiome 2019, 7, 3. [Google Scholar] [CrossRef]
- Roush, D.; Giraldo-Silva, A.; Garcia-Pichel, F. Cydrasil 3, a Curated 16S rRNA Gene Reference Package and Web App for Cyanobacterial Phylogenetic Placement. Sci. Data 2021, 8, 230. [Google Scholar] [CrossRef]
- Tandon, K.; Pasella, M.M.; Iha, C.; Ricci, F.; Hu, J.; O’Kelly, C.J.; Medina, M.; Kühl, M.; Verbruggen, H. Every Refuge Has Its Price: Ostreobium as a Model for Understanding How Algae Can Live in Rock and Stay in Business. Semin. Cell Dev. Biol. 2022, S1084952122000775. [Google Scholar] [CrossRef]
- Behrendt, L.; Larkum, A.W.; Norman, A.; Qvortrup, K.; Chen, M.; Ralph, P.; Sørensen, S.J.; Trampe, E.; Kühl, M. Endolithic Chlorophyll d-Containing Phototrophs. ISME J. 2011, 5, 1072–1076. [Google Scholar] [CrossRef] [PubMed]
- Massé, A.; Tribollet, A.; Meziane, T.; Bourguet-Kondracki, M.; Yéprémian, C.; Sève, C.; Thiney, N.; Longeon, A.; Couté, A.; Domart-Coulon, I. Functional Diversity of Microboring Ostreobium Algae Isolated from Corals. Environ. Microbiol. 2020, 22, 4825–4846. [Google Scholar] [CrossRef]
- Iha, C.; Dougan, K.E.; Varela, J.A.; Avila, V.; Jackson, C.J.; Bogaert, K.A.; Chen, Y.; Judd, L.M.; Wick, R.; Holt, K.E.; et al. Genomic Adaptations to an Endolithic Lifestyle in the Coral-Associated Alga Ostreobium. Curr. Biol. 2021, 31, 1393–1402. [Google Scholar] [CrossRef]
- Golubic, S.; Seong-Joo, L.; Browne, K.M. Cyanobacteria: Architects of Sedimentary Structures. In Microbial Sediments; Riding, R.E., Awramik, S.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 57–67. ISBN 978-3-642-08275-7. [Google Scholar]
- Merz-Preiβ, M. Calcification in Cyanobacteria. In Microbial Sediments; Riding, R.E., Awramik, S.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 50–56. ISBN 978-3-642-08275-7. [Google Scholar]
- Stumm, W.; Morgan, J.J. Aquatic Chemistry. Chemical Equilibria and Rates in Natural Waters, 3rd ed.; Environmental Science and Technology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 1996. [Google Scholar]
- Garcia-Pichel, F. Plausible Mechanisms for the Boring on Carbonates by Microbial Phototrophs. Sediment. Geol. 2006, 185, 205–213. [Google Scholar] [CrossRef]
- Alexanderson, T.E. Marks of Unknown Carbonate-Decomposing Organelles in Cyanophyte Borings. Nature 1975, 254, 236–238. [Google Scholar] [CrossRef]
- Hatch, W.I. The Implication of Carbonic Anhydrase in the Physiological Mechanism of Penetration of Carbonate Substrata by the Marine Burrowing Sponge Cliona Celata (Demospongiae). Biol. Bull. 1980, 159, 135–147. [Google Scholar] [CrossRef]
- Cockell, C.S.; Herrera, A. Why Are Some Microorganisms Boring? Trends Microbiol. 2008, 16, 101–106. [Google Scholar] [CrossRef]
- Garcia-Pichel, F.; Ramirez-Reinat, E.; Gao, Q. Microbial Excavation of Solid Carbonates Powered by P-Type ATPase-Mediated Transcellular Ca2+ Transport. Proc. Natl. Acad. Sci. USA 2010, 107, 21749–21754. [Google Scholar] [CrossRef] [PubMed]
- Stal, L.J.; Krumbein, W.E. Temporal Separation of Nitrogen Fixation and Photosynthesis in the Filamentous, Non-Heterocystous Cyanobacterium Oscillatoria Sp. Arch. Microbiol. 1987, 149, 76–80. [Google Scholar] [CrossRef]
- Guida, B.S.; Garcia-Pichel, F. Extreme Cellular Adaptations and Cell Differentiation Required by a Cyanobacterium for Carbonate Excavation. Proc. Natl. Acad. Sci. USA 2016, 113, 5712–5717. [Google Scholar] [CrossRef]
- Fredd, C.N.; Fogler, H.S. The Influence of Chelating Agents on the Kinetics of Calcite Dissolution. J. Colloid Interface Sci. 1998, 204, 187–197. [Google Scholar] [CrossRef]
- Ramírez-Reinat, E.L.; Garcia-Pichel, F. Prevalence of Ca2+-ATPase-Mediated Carbonate Dissolution among Cyanobacterial Euendoliths. Appl. Environ. Microbiol. 2012, 78, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Krause, S.; Liebetrau, V.; Nehrke, G.; Damm, T.; Büsse, S.; Leipe, T.; Vogts, A.; Gorb, S.N.; Eisenhauer, A. Endolithic Algae Affect Modern Coral Carbonate Morphology and Chemistry. Front. Earth Sci. 2019, 7, 304. [Google Scholar] [CrossRef]
- Guida, B.S.; Bose, M.; Garcia-Pichel, F. Carbon Fixation from Mineral Carbonates. Nat. Commun. 2017, 8, 1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinfelder, J.R. Carbon Concentrating Mechanisms in Eukaryotic Marine Phytoplankton. Annu. Rev. Mar. Sci. 2011, 3, 291–315. [Google Scholar] [CrossRef] [PubMed]
- Guida, B.S. Unique Cellular, Physiological, and Metabolic Adaptations to the Euendolithic Lifestyle in a Boring Cyanobacterium. Ph.D. Thesis, Arizona State University, Tempe, AZ, USA, 2016. [Google Scholar]
- Ascaso, C.; Wierzchosb, J.; Castelloa, R. Study of the Biogenic Weathering of Calcareous Litharenite Stones Caused by Lichen and Endolithic Microorganisms. Int. Biodeterior. Biodegrad. 1998, 42, 29–38. [Google Scholar] [CrossRef]
- Huber, J.; Jadin, F. Sur Une Algue Perforante d’Eau Douce. C R. Hebd. Séances L’Académie Sci. Paris 1892, 115, 262–264. [Google Scholar]
- Akpan, E.B. Bioerosion of Oyster Shells in Brackish Modern Mangrove Swamps, Nigeria. Ichnos 1990, 1, 125–132. [Google Scholar] [CrossRef]
- Cerrano, C.; Bavestrello, G.; Calcinai, B.; Cattaneo-Vietti, R.; Chiantore, M.; Guidetti, M.; Sarà, A. Bioerosive Processes in Antarctic Seas. Polar Biol. 2001, 24, 790–792. [Google Scholar] [CrossRef]
- Pantazidou, A.; Louvrou, I.; Economou-Amilli, A. Euendolithic Shell-Boring Cyanobacteria and Chlorophytes from the Saline Lagoon Ahivadolimni on Milos Island, Greece. Eur. J. Phycol. 2006, 41, 189–200. [Google Scholar] [CrossRef]
- Ćurin, M.; Peharda, M.; Calcinai, B.; Golubić, S. Incidence of Damaging Endolith Infestation of the Edible Mytilid Bivalve Modiolus Barbatus. Mar. Biol. Res. 2014, 10, 179–189. [Google Scholar] [CrossRef]
- Akpan, E.B.; Farrow, G.E. Shell-Boring Algae on the Scottish Continental Shelf: Identification, Distribution, Bathymetric Zonation. Trans. R. Soc. Edinb. Earth Sci. 1984, 75, 1–12. [Google Scholar] [CrossRef]
- Försterra, G.; Beuck, L.; Häussermann, V.; Freiwald, A. Shallow-Water Desmophyllum Dianthus (Scleractinia) from Chile: Characteristics of the Biocoenoses, the Bioeroding Community, Heterotrophic Interactions and (Paleo)-Bathymetric Implications. In Cold-Water Corals and Ecosystems; Freiwald, A., Roberts, J.M., Eds.; Erlangen Earth Conference Series; Springer-Verlag: Berlin/Heidelberg, Germany, 2005; pp. 937–977. ISBN 978-3-540-24136-2. [Google Scholar]
- Wilkinson, M.; Burrows, E.M. The Distribution of Marine Shell-Boring Green Algae. J. Mar. Biol. Assoc. UK 1972, 52, 59–65. [Google Scholar] [CrossRef]
- Mao Che, L.; Le Campion-Alsumard, T.; Boury-Esnault, N.; Payri, C.; Golubic, S.; Bézac, C. Biodegradation of Shells of the Black Pearl Oyster, Pinctada Margaritifera Var. Cumingii, by Microborers and Sponges of French Polynesia. Mar. Biol. 1996, 126, 509–519. [Google Scholar] [CrossRef]
- Perry, C.T. Grain Susceptibility to the Effects of Microboring: Implications for the Preservation of Skeletal Carbonates. Sedimentology 1998, 45, 39–51. [Google Scholar] [CrossRef]
- Tribollet, A. Dissolution of Dead Corals by Euendolithic Microorganisms Across the Northern Great Barrier Reef (Australia). Microb. Ecol. 2008, 55, 569–580. [Google Scholar] [CrossRef]
- Meyer, N.; Wisshak, M.; Freiwald, A. Ichnodiversity and Bathymetric Range of Microbioerosion Traces in Polar Barnacles of Svalbard. Polar Res. 2020, 39, 3766. [Google Scholar] [CrossRef]
- Campbell, S.E. The Modern Distribution and Geological History of Calcium Carbonate Boring Microorganisms. In Biomineralization and Biological Metal Accumulation; Westbroek, P., de Jong, E.W., Eds.; Springer: Dordrecht, The Netherlands, 1983; pp. 99–104. ISBN 978-94-009-7946-8. [Google Scholar]
- Gektidis, M. Development of Microbial Euendolithic Communities: The Influence of Light and Time. Bull. Geol. Soc. Den. 1999, 45, 147–150. [Google Scholar] [CrossRef]
- Tribollet, A.; Golubic, S.; Radtke, G.; Reitner, J. On Microbiocorrosion. In Advances in Stromatolite Geobiology; Lecture Notes in Earth Sciences; Springer: Berlin/Heidelberg, Germany, 2011; Volume 131, pp. 265–276. ISBN 978-3-642-10414-5. [Google Scholar]
- Zardi, G.I.; Nicastro, K.R.; McQuaid, C.D.; Gektidis, M. Effects of Endolithic Parasitism on Invasive and Indigenous Mussels in a Variable Physical Environment. PLoS ONE 2009, 4, e6560. [Google Scholar] [CrossRef]
- Gehman, A.M.; Harley, C.D.G. Symbiotic Endolithic Microbes Alter Host Morphology and Reduce Host Vulnerability to High Environmental Temperatures. Ecosphere 2019, 10, e02683. [Google Scholar] [CrossRef]
- Monsinjon, J.R.; McQuaid, C.D.; Nicastro, K.R.; Seuront, L.; Oróstica, M.H.; Zardi, G.I. Weather and Topography Regulate the Benefit of a Conditionally Helpful Parasite. Funct. Ecol. 2021, 35, 2691–2706. [Google Scholar] [CrossRef]
- Gektidis, M.; Dubinsky, Z.; Goffredo, S. Microendoliths of the Shallow Euphotic Zone in Open and Shaded Habitats at 30°N–Eilat, Israel – Paleoecological Implications. Facies 2007, 53, 43–55. [Google Scholar] [CrossRef]
- Wisshak, M.; Tribollet, A.; Golubic, S.; Jakobsen, J.; Freiwald, A. Temperate Bioerosion: Ichnodiversity and Biodiversity from Intertidal to Bathyal Depths (Azores). Geobiology 2011, 9, 492–520. [Google Scholar] [CrossRef]
- Le Bris, S.; Le Campion-Alsumard, T.; Romano, J.-C. Caractéristiques du feutrage algal des récifs coralliens de Polynésie française soumis à différentes intensités de bioérosion. Oceanol. Acta 1998, 21, 695–708. [Google Scholar] [CrossRef]
- Tribollet, A.; Golubic, S. Cross-Shelf Differences in the Pattern and Pace of Bioerosion of Experimental Carbonate Substrates Exposed for 3 Years on the Northern Great Barrier Reef, Australia. Coral Reefs 2005, 24, 422–434. [Google Scholar] [CrossRef]
- Wisshak, M.; Gektidis, M.; Freiwald, A.; Lundälv, T. Bioerosion along a Bathymetric Gradient in a Cold-Temperate Setting (Kosterfjord, SW Sweden): An Experimental Study. Facies 2005, 51, 93–117. [Google Scholar] [CrossRef]
- Zardi, G.I.; Monsinjon, J.R.; McQuaid, C.D.; Seuront, L.; Orostica, M.; Want, A.; Firth, L.B.; Nicastro, K.R. Foul-weather Friends: Modelling Thermal Stress Mitigation by Symbiotic Endolithic Microbes in a Changing Environment. Glob. Change Biol. 2021, 27, 2549–2560. [Google Scholar] [CrossRef]
- Ramírez-Reinat, E.L.; Garcia-Pichel, F. Characterization of a Marine Cyanobacterium That Bores into Carbonates and the Redescription of the Genus Mastigocoleus. J. Phycol. 2012, 48, 740–749. [Google Scholar] [CrossRef]
- Meyer, N.; Wisshak, M.; Freiwald, A. Bioerosion Ichnodiversity in Barnacles from the Ross Sea, Antarctica. Polar Biol. 2021, 44, 667–682. [Google Scholar] [CrossRef]
- Hutchings, P.A. Biological Destruction of Coral Reefs. Coral Reefs 1986, 4, 239–252. [Google Scholar] [CrossRef]
- Chazottes, V.; Le Campion-Alsumard, T.; Peyrot-Clausade, M.; Cuet, P. The Effects of Eutrophication-Related Alterations to Coral Reef Communities on Agents and Rates of Bioerosion (Reunion Island, Indian Ocean). Coral Reefs 2002, 21, 375–390. [Google Scholar] [CrossRef]
- Chazottes, V.; Cabioch, G.; Golubic, S.; Radtke, G. Bathymetric Zonation of Modern Microborers in Dead Coral Substrates from New Caledonia—Implications for Paleodepth Reconstructions in Holocene Corals. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 280, 456–468. [Google Scholar] [CrossRef]
- Bentis, C.; Kaufman, L.; Golubic, S. Endolithic Fungi in Reef-Building Corals (Order: Scleractinia) Are Common, Cosmopolitan, and Potentially Pathogenic. Biol. Bull. 2000, 198, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Vogel, K.; Gektidis, M.; Golubic, S.; Kiene, W.E.; Radtke, G. Experimental Studies on Microbial Bioerosion at Lee Stocking Island, Bahamas and One Tree Island, Great Barrier Reef, Australia: Implications for Paleoecological Reconstructions. Lethaia 2000, 33, 190–204. [Google Scholar] [CrossRef]
- Radtke, G.; Golubic, S. Microborings in Mollusk Shells, Bay of Safaga, Egypt: Morphometry and Ichnology. Facies 2005, 51, 118–134. [Google Scholar] [CrossRef]
- Golubic, S.; Campbell, S.E.; Drobne, K.; Cameron, B.; Balsam, W.L.; Cimerman, F.; Duboiss, L. Microbial Endoliths: A Benthic Overprint in the Sedimentary Record, and a Paleobathymetric Cross-Reference with Foraminifera. J. Paleontol. 1984, 58, 12. [Google Scholar]
- Kiene, W.E.; Radtke, G.; Gektidis, M.; Golubic, S.; Vogel, K. Factors Controlling the Distribution of Microborers in Bahamian Reef Environments. Facies 1995, 32, 176–188. [Google Scholar]
- Le Campion-Alsumard, T.; Campbell, S.E.; Golubic, S. Endoliths and the Depth of the Photic Zone: Discussion. J. Sediment. Petrol. 1982, 52, 1333–13338. [Google Scholar] [CrossRef]
- Lukas, K.J. Depth Distribution and Form among Common Microboring Algae from the Florida Continental Shelf. In Proceedings of the Abstract with Programs; Boulder: Toronto, ON, Canada, 1978; Volume 10, pp. 1–448. [Google Scholar]
- Försterra, G.; Häussermann, V. Unusual Symbiotic Relationships between Microendolithic Phototrophic Organisms and Azooxanthellate Cold-Water Corals from Chilean Fjords. Mar. Ecol. Prog. Ser. 2008, 370, 121–125. [Google Scholar] [CrossRef]
- Reyes-Nivia, C.; Diaz-Pulido, G.; Dove, S. Relative Roles of Endolithic Algae and Carbonate Chemistry Variability in the Skeletal Dissolution of Crustose Coralline Algae. Biogeosciences Discuss. 2014, 11, 2993–3021. [Google Scholar] [CrossRef]
- Tribollet, A.; Veinott, G.; Golubic, S.; Dart, R. Infestation of the North American Freshwater Mussel Elliptio Complanata (Head Lake, Canada) by the Euendolithic Cyanobacterium Plectonema Terebrans Bornet et Flahault. Algol. Stud. 2008, 128, 65–77. [Google Scholar] [CrossRef]
- Gaspard, D. Endolithic Algae, Fungi and Bacterial Activity in Holocene and Cretaceous Brachiopod Shells—Diagenetic Consequences. Mem. Assoc. Australas. Palaeontol. 2011, 41, 327–337. [Google Scholar]
- Lukas, K.J. Taxonomy and Ecology of the Endolithic Microflora of Reef Corals with a Review of the Literature on Endolithic Microphytes. Ph.D. Thesis, University of Rhode Island, Kingston, RI, USA, 1973. [Google Scholar]
- Perkins, R.D.; Halsey, S.D. Geologic Significance of Microboring Fungi and Algae in Carolina Shelf Sediments. J. Sediment. Res. 1971, 41, 843–853. [Google Scholar] [CrossRef]
- Nash, M.C.; Opdyke, B.N.; Troitzsch, U.; Russell, B.D.; Adey, W.H.; Kato, A.; Diaz-Pulido, G.; Brent, C.; Gardner, M.; Prichard, J.; et al. Dolomite-Rich Coralline Algae in Reefs Resist Dissolution in Acidified Conditions. Nat. Clim. Chang. 2013, 3, 268–272. [Google Scholar] [CrossRef]
- Diaz-Pulido, G.; Nash, M.C.; Anthony, K.R.N.; Bender, D.; Opdyke, B.N.; Reyes-Nivia, C.; Troitzsch, U. Greenhouse Conditions Induce Mineralogical Changes and Dolomite Accumulation in Coralline Algae on Tropical Reefs. Nat. Commun. 2014, 5, 3310. [Google Scholar] [CrossRef] [PubMed]
- Golubic, S.; Radtke, G.; Campion-Alsumard, T.L. Endolithic Fungi in Marine Ecosystems. Trends Microbiol. 2005, 13, 229–235. [Google Scholar] [CrossRef]
- Gutiérrez-Isaza, N.; Espinoza-Avalos, J.; León-Tejera, H.P.; González-Solís, D. Endolithic Community Composition of Orbicella Faveolata (Scleractinia) underneath the Interface between Coral Tissue and Turf Algae. Coral Reefs 2015, 34, 625–630. [Google Scholar] [CrossRef]
- Keats, D.W.; Groener, A.; Chamberlain, Y.M. Cell Sloughing in the Littoral Zone Coralline Alga, Spongites Yendoi (Foslie) Chamberlain (Corallinales, Rhodophyta). Phycologia 1993, 32, 143–150. [Google Scholar] [CrossRef]
- Owen, G.; Williams, A. The Caecum of Articulate Brachiopoda. Proc. R. Soc. Lond. B Biol. Sci. 1969, 172, 187–201. [Google Scholar]
- Scardino, A.; De Nys, R.; Ison, O.; O’Connor, W.; Steinberg, P. Microtopography and Antifouling Properties of the Shell Surface of the Bivalve Molluscs Mytilus Galloprovincialis and Pinctada Imbricata. Biofouling 2003, 19, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Scardino, A.; de Nys, R. Fouling Deterrence on the Bivalve Shell Mytilus Galloprovincialis: A Physical Phenomenon? Biofouling 2004, 20, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Bers, A.V.; Díaz, E.R.; da Gama, B.A.P.; Vieira-Silva, F.; Dobretsov, S.; Valdivia, N.; Thiel, M.; Scardino, A.J.; McQuaid, C.D.; Sudgen, H.E.; et al. Relevance of Mytilid Shell Microtopographies for Fouling Defence—A Global Comparison. Biofouling 2010, 26, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Prusina, I.; Peharda, M.; Ezgeta-Balic, D.; Puljas, S.; Glamuzina, B.; Golubic, S. Life-History Trait of the Mediterranean Keystone Species Patella Rustica: Growth and Microbial Bioerosion. Mediterr. Mar. Sci. 2015, 16, 393. [Google Scholar] [CrossRef]
- Odum, H.T.; Odum, E.P. Trophic Structure and Productivity of a Windward Coral Reef Community on Eniwetok Atoll. Ecol. Monogr. 1955, 25, 291–320. [Google Scholar] [CrossRef]
- Fine, M.; Roff, G.; Ainsworth, T.D.; Hoegh-Guldberg, O. Phototrophic Microendoliths Bloom during Coral “White Syndrome”. Coral Reefs 2006, 25, 577–581. [Google Scholar] [CrossRef]
- Grange, J.S.; Rybarczyk, H.; Tribollet, A. The Three Steps of the Carbonate Biogenic Dissolution Process by Microborers in Coral Reefs (New Caledonia). Environ. Sci. Pollut. Res. 2015, 22, 13625–13637. [Google Scholar] [CrossRef]
- Le Campion-Alsumard, T. Les Cyanophycées endolithes marines. Systématique, ultrastructure, écologie et biodestruction. Oceanol. Acta 1979, 2, 143–156. [Google Scholar]
- Tribollet, A.; Golubic, S. Reef Bioerosion: Agents and Processes. In Coral Reefs: An Ecosystem in Transition; Dubinsky, Z., Stambler, N., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 435–450. ISBN 978-94-007-0113-7. [Google Scholar]
- Fordyce, A.J.; Ainsworth, T.D.; Leggat, W. Microalgae, a Boring Bivalve and a Coral—A Newly Described Association Between Two Coral Reef Bioeroders Within Their Coral Host. Integr. Org. Biol. 2020, 2, obaa035. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.; Torunski, H. Biokarst on Limestone Coasts, Morphogenesis and Sediment Production. Mar. Ecol. 1983, 4, 45–63. [Google Scholar] [CrossRef]
- Nicholson, G.M.; Clements, K.D. Resolving Resource Partitioning in Parrotfishes (Scarini) Using Microhistology of Feeding Substrata. Coral Reefs 2020, 39, 1313–1327. [Google Scholar] [CrossRef]
- Zubia, M.; Peyrot-Clausade, M. Internal Bioerosion of Acropora Formosa in Réunion (Indian Ocean): Microborer and Macroborer Activities. Oceanol. Acta 2001, 24, 251–262. [Google Scholar] [CrossRef]
- Rice, M.M.; Maher, R.L.; Correa, A.M.S.; Moeller, H.V.; Lemoine, N.P.; Shantz, A.A.; Burkepile, D.E.; Silbiger, N.J. Macroborer Presence on Corals Increases with Nutrient Input and Promotes Parrotfish Bioerosion. Coral Reefs 2020, 39, 409–418. [Google Scholar] [CrossRef]
- Pari, N.; Peyrot-Clausade, M.; Le Campion-Alsumard, T.; Hutchings, P.; Chazottes, V.; Golubic, S.; Le Campion, J.; Fontaine, M. Bioerosion of Experimental Substrates on High Islands and on Atoll Lagoons (French Polynesia) after Two Years of Exposure. Mar. Ecol. Prog. Ser. 1998, 166, 119–130. [Google Scholar] [CrossRef]
- Pari, N.; Peyrot-Clausade, M.; Hutchings, P.A. Bioerosion of Experimental Substrates on High Islands and Atoll Lagoons (French Polynesia) during 5 Years of Exposure. J. Exp. Mar. Biol. Ecol. 2002, 276, 109–127. [Google Scholar] [CrossRef]
- Carreiro-Silva, M.; McClanahan, T.R.; Kiene, W.E. The Role of Inorganic Nutrients and Herbivory in Controlling Microbioerosion of Carbonate Substratum. Coral Reefs 2005, 24, 214–221. [Google Scholar] [CrossRef]
- Carreiro-Silva, M.; McClanahan, T.; Kiene, W. Effects of Inorganic Nutrients and Organic Matter on Microbial Euendolithic Community Composition and Microbioerosion Rates. Mar. Ecol. Prog. Ser. 2009, 392, 1–15. [Google Scholar] [CrossRef]
- Carreiro-Silva, M.; Kiene, W.E.; Golubic, S.; McClanahan, T.R. Phosphorus and Nitrogen Effects on Microbial Euendolithic Communities and Their Bioerosion Rates. Mar. Pollut. Bull. 2012, 64, 602–613. [Google Scholar] [CrossRef]
- Godinot, C.; Tribollet, A.; Grover, R.; Ferrier-Pagès, C. Bioerosion by Euendoliths Decreases in Phosphate-Enriched Skeletons of Living Corals. Biogeosciences Discuss. 2012, 9, 2425–2444. [Google Scholar] [CrossRef]
- del Campo, J.; Pombert, J.-F.; Šlapeta, J.; Larkum, A.; Keeling, P.J. The ‘Other’ Coral Symbiont: Ostreobium Diversity and Distribution. ISME J. 2017, 11, 296–299. [Google Scholar] [CrossRef]
- Al-Thukair, A.A. Effect of Oil Pollution on Euendolithic Cyanobacteria of the Arabian Gulf. Environ. Microbiol. 2002, 4, 125–129. [Google Scholar] [CrossRef]
- Cherchi, A.; Buosi, C.; Zuddas, P.; De Giudici, G. Bioerosion by Microbial Euendoliths in Benthic Foraminifera from Heavy Metal-Polluted Coastal Environments of Portovesme (South-Western Sardinia, Italy). Biogeosciences 2012, 9, 4607–4620. [Google Scholar] [CrossRef]
- Cherchi, A.; Da Pelo, S.; Ibba, A.; Mana, D.; Buosi, C.; Floris, N. Benthic Foraminifera Response and Geochemical Characterization of the Coastal Environment Surrounding the Polluted Industrial Area of Portovesme (South-Western Sardinia, Italy). Mar. Pollut. Bull. 2009, 59, 281–296. [Google Scholar] [CrossRef]
- Cabioch, G.; Montaggioni, L.F.; Faure, G.; Ribaud-Laurenti, A. Reef Coralgal Assemblages as Recorders of Paleobathymetry and Sea Level Changes in the Indo-Pacific Province. Quat. Sci. Rev. 1999, 18, 1681–1695. [Google Scholar] [CrossRef]
- Perry, C.T.; Macdonald, I.A. Impacts of Light Penetration on the Bathymetry of Reef Microboring Communities: Implications for the Development of Microendolithic Trace Assemblages. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2002, 186, 101–113. [Google Scholar] [CrossRef]
- Swinchatt, J.P. Algal Boring: A Possible Depth Indicator in Carbonate Rocks and Sediments. Geol. Soc. Am. Bull. 1969, 80, 1391. [Google Scholar] [CrossRef]
- Massé, A.; Domart-Coulon, I.; Golubic, S.; Duché, D.; Tribollet, A. Early Skeletal Colonization of the Coral Holobiont by the Microboring Ulvophyceae Ostreobium Sp. Sci. Rep. 2018, 8, 2293. [Google Scholar] [CrossRef] [PubMed]
- Ndhlovu, A.; McQuaid, C.D.; Monaco, C.J. Ectoparasites Reduce Scope for Growth in a Rocky-Shore Mussel (Perna Perna) by Raising Maintenance Costs. Sci. Total Environ. 2021, 753, 142020. [Google Scholar] [CrossRef]
- Curry, G.B. Microborings in Recent Brachiopods and the Functions of Caeca. Lethaia 1983, 16, 119–127. [Google Scholar] [CrossRef]
- Schlichter, D.; Zscharnack, B.; Krisch, H. Transfer of Photoassimilates from Endolithic Algae to Coral Tissue. Naturwissenschaften 1995, 82, 1–564. [Google Scholar] [CrossRef]
- Schlichter, D.; Kampmann, H.; Conrady, S. Trophic Potential and Photoecology of Endolithic Algae Living within Coral Skeletons. Mar. Ecol. 1997, 18, 299–317. [Google Scholar] [CrossRef]
- Marquet, N.; Nicastro, K.R.; Gektidis, M.; McQuaid, C.D.; Pearson, G.A.; Serrão, E.A.; Zardi, G.I. Comparison of Phototrophic Shell-Degrading Endoliths in Invasive and Native Populations of the Intertidal Mussel Mytilus Galloprovincialis. Biol. Invasions 2013, 15, 1253–1272. [Google Scholar] [CrossRef]
- Ndhlovu, A.; McQuaid, C.D.; Nicastro, K.R.; Zardi, G.I. Parasitism by Endolithic Cyanobacteria Reduces Reproductive Output and Attachment Strength of Intertidal Ecosystem Engineers. Mar. Biol. 2022, 169, 37. [Google Scholar] [CrossRef]
- Goldberg, W.M.; Makemson, J.C.; Colley, S.B. Entocladia Endozoica Sp. Nov., a Pathogenic Chlorophyte: Structure, Life History, Physiology, and Effect on Its Coral Host. Biol. Bull. 1984, 166, 368–383. [Google Scholar] [CrossRef]
- Nicastro, K.R.; McQuaid, C.D.; Zardi, G.I. Between a Rock and a Hard Place: Combined Effect of Trampling and Phototrophic Shell-Degrading Endoliths in Marine Intertidal Mussels. Mar. Biodivers. 2019, 49, 1581–1586. [Google Scholar] [CrossRef]
- Nolan, C.P. Size, Shape and Shell Morphology in the Antarctic Limpet Nacella Concinna at Signy Island, South Orkney Islands. J. Molluscan Stud. 1991, 57, 225–238. [Google Scholar] [CrossRef]
- Fine, M.; Steindler, L.; Loya, Y. Endolithic Algae Photoacclimate to Increased Irradiance during Coral Bleaching. Mar. Freshw. Res. 2004, 55, 115. [Google Scholar] [CrossRef]
- Fine, M.; Meroz-Fine, E.; Hoegh-Guldberg, O. Tolerance of Endolithic Algae to Elevated Temperature and Light in the Coral Montipora Monasteriata from the Southern Great Barrier Reef. J. Exp. Biol. 2005, 208, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Nivia, C.; Diaz-Pulido, G.; Kline, D.; Guldberg, O.-H.; Dove, S. Ocean Acidification and Warming Scenarios Increase Microbioerosion of Coral Skeletons. Glob. Change Biol. 2013, 19, 1919–1929. [Google Scholar] [CrossRef] [PubMed]
- Risk, M.J.; Sammarco, P.W.; Edinger, E.N. Bioerosion in Acropora across the Continental Shelf of the Great Barrier Reef. Coral Reefs 1995, 14, 79–86. [Google Scholar] [CrossRef]
- Le Campion-Alsumard, T.; Golubic, S.; Priess, K. Fungi in Corals: Symbiosis or Disease? Interaction between Polyps and Fungi Causes Pearl-like Skeleton Biomineralization. Mar. Ecol. Prog. Ser. 1995, 117, 137–147. [Google Scholar] [CrossRef]
- Morse, D.E.; Morse, A.; Duncan, H.; Trench, R.K. Algal Tumors in the Caribbean Octocorallian, Gorgonia Ventalina: II. Biochemical Characterization of the Algae, and First Epidemiological Observations. Bull. Mar. Sci. 1981, 31, 399–409. [Google Scholar]
- Morse, D.E.; Morse, A.; Duncan, H. Algal Tumors in the Caribeean Sea-Fan, Gorgonia Ventalina. In Proceeding of Third International Coral Reef Symposium; Taylor, D.L., Ed.; Rosenstiel School of Marine and Atmospheric Science: Miami, FL, USA, 1977; Volume 1, pp. 623–629. [Google Scholar]
- Nicastro, K.R.; Seuront, L.; McQuaid, C.D.; Zardi, G.I. Symbiont-Induced Intraspecific Phenotypic Variation Enhances Plastic Trapping and Ingestion in Biogenic Habitats. Sci. Total Environ. 2022, 826, 153922. [Google Scholar] [CrossRef] [PubMed]
- Stolarski, J. Gardineria—A Scleractinian Living Fossil. Acta Palaeontol. Pol. 1996, 41, 339–367. [Google Scholar]
- Brown, B.E.; Tudhope, A.W.; Le Tissier, M.D.A.; Scoffin, T.P. A Novel Mechanism for Iron Incorporation into Coral Skeletons. Coral Reefs 1991, 10, 211–215. [Google Scholar] [CrossRef]
- Lazier, A.V.; Smith, J.E.; Risk, M.J.; Schwarcz, H.P. The Skeletal Structure of Desmophyllum Cristagalli: The Use of Deep-Water Corals in Sclerochronology. Lethaia 1999, 32, 119–130. [Google Scholar] [CrossRef]
- Steneck, R.S. The Ecology of Coralline Algal Crusts: Convergent Patterns and Adaptative Strategies. Annu. Rev. Ecol. Syst. 1986, 17, 3–303. [Google Scholar] [CrossRef]
- Laborel, J.; Le Campion-Alsumard, T. Infestation Massive Du Squelette de Coraux Vivants Par Des Rhodophycées de Type Conchocelis. Comptes Rendus Acad. Sci. Paris 1979, 288, 1575–1577. [Google Scholar]
- Ralph, P.J.; Larkum, A.W.D.; Kühl, M. Photobiology of Endolithic Microorganisms in Living Coral Skeletons: 1. Pigmentation, Spectral Reflectance and Variable Chlorophyll Fluorescence Analysis of Endoliths in the Massive Corals Cyphastrea Serailia, Porites Lutea and Goniastrea Australensis. Mar. Biol. 2007, 152, 395–404. [Google Scholar] [CrossRef]
- Ghirardelli, L.A. An Endolithic Cyanophyte in the Cell Wall of Calcareous Algae. Bot. Mar. 1998, 41, 367–373. [Google Scholar] [CrossRef]
- Duerden, J.E. Boring Algae as Agents in the Disintegration of Corals. Bull. Am. Mus. Nat. Hist. 1902, 16, 323–332. [Google Scholar]
- Highsmith, R.C. Lime-Boring Algae in Hermatypic Coral Skeletons. J. Exp. Mar. Biol. Ecol. 1981, 55, 267–281. [Google Scholar] [CrossRef]
- Delvoye, L. Endolithic Algae in Living Stony Corals: Algal Concentrations under Influence of Depth-Dependent Light Conditions and Coral Tissue Fluorescence in Agaricia agaricites (L.) and Meandrina meandrites (L.) (Scleractinia, Anthozoa). Stud. Nat. Hist. Caribb. Reg. 1992, 71, 24–41. [Google Scholar]
- Tribollet, A.; Pica, D.; Puce, S.; Radtke, G.; Campbell, S.E.; Golubic, S. Euendolithic Conchocelis Stage (Bangiales, Rhodophyta) in the Skeletons of Live Stylasterid Reef Corals. Mar. Biodivers. 2018, 48, 1855–1862. [Google Scholar] [CrossRef]
- LaJeunesse, T.C.; Parkinson, J.E.; Gabrielson, P.W.; Jeong, H.J.; Reimer, J.D.; Voolstra, C.R.; Santos, S.R. Systematic Revision of Symbiodiniaceae Highlights the Antiquity and Diversity of Coral Endosymbionts. Curr. Biol. 2018, 28, 2570–2580. [Google Scholar] [CrossRef] [PubMed]
- Marshall, A.T. Calcification in Hermatypic and Ahermatypic Corals. Science 1996, 271, 637–639. [Google Scholar] [CrossRef]
- Aeby, G.S.; Williams, G.J.; Franklin, E.C.; Haapkyla, J.; Harvell, C.D.; Neale, S.; Page, C.A.; Raymundo, L.; Vargas-Ángel, B.; Willis, B.L.; et al. Growth Anomalies on the Coral Genera Acropora and Porites Are Strongly Associated with Host Density and Human Population Size across the Indo-Pacific. PLoS ONE 2011, 6, e16887. [Google Scholar] [CrossRef] [PubMed]
- McClanahan, T.R.; Weil, E.; Maina, J. Strong Relationship between Coral Bleaching and Growth Anomalies in Massive Porites. Glob. Chang. Biol. 2009, 15, 1804–1816. [Google Scholar] [CrossRef]
- Miller, A.; Blackwelder, P.; Al-Sayegh, H.; Richardson, L. Fine-Structural Analysis of Black Band Disease-Infected Coral Reveals Boring Cyanobacteria and Novel Bacteria. Dis. Aquat. Organ. 2011, 93, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Kanwisher, J.W.; Wainwright, S.A. Oxygen Balance in Some Reef Corals. Biol. Bull. 1967, 133, 378–390. [Google Scholar] [CrossRef]
- Halldal, P. Photosynthetic Capacities and Photosynthetic Action Spectra of Endozoic Algae of the Massive Coral Favia. Biol. Bull. 1968, 134, 411–424. [Google Scholar] [CrossRef]
- Magnusson, S.; Fine, M.; Kühl, M. Light Microclimate of Endolithic Phototrophs in the Scleractinian Corals Montipora Monasteriata and Porites Cylindrica. Mar. Ecol. Prog. Ser. 2007, 332, 119–128. [Google Scholar] [CrossRef]
- Shibata, K.; Haxo, F.T. Light Transmission and Spectral Distribution through Epi- and Endozoic Algal Layers in the Brain Coral, Favia. Biol. Bull. 1969, 136, 461–468. [Google Scholar] [CrossRef]
- Shashar, N.; Stambler, N. Endolithic Algae within Corals—Life in an Extreme Environment. J. Exp. Mar. Biol. Ecol. 1992, 163, 277–286. [Google Scholar] [CrossRef]
- Galindo-Martínez, C.T.; Weber, M.; Avila-Magaña, V.; Enríquez, S.; Kitano, H.; Medina, M.; Iglesias-Prieto, R. The Role of the Endolithic Alga Ostreobium Spp. during Coral Bleaching Recovery. Sci. Rep. 2022, 12, 2977. [Google Scholar] [CrossRef] [PubMed]
- Schlichter, D. Cnidaria: Permability, Epidermal Transport and Related Phenoma. In Biology of the Integument; Bereiter-Hahn, J., Matoltsy, A.G., Richards, K.S., Eds.; Springer: Berlin/Heidelberg, Germany, 1984; pp. 79–95. ISBN 978-3-642-51595-8. [Google Scholar]
- Kühl, M.; Holst, G.; Larkum, A.W.D.; Ralph, P.J. Imaging of Oxygen Dynamics within the Endolithic Algal Community of the Massive Coral Porites Lobata. J. Phycol. 2008, 44, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Titlyanov, E.A.; Kiyashko, S.I.; Titlyanova, T.V.; Kalita, T.L.; Raven, J.A. δ13C and δ15N Values in Reef Corals Porites Lutea and P. Cylindrica and in Their Epilithic and Endolithic Algae. Mar. Biol. 2008, 155, 353–361. [Google Scholar] [CrossRef]
- Fine, M.; Zibrowius, H.; Loya, Y. Oculina Patagonica: A Non-Lessepsian Scleractinian Coral Invading the Mediterranean Sea. Mar. Biol. 2001, 138, 1195–1203. [Google Scholar] [CrossRef]
- Nothdurft, L.D.; Webb, G.E. Earliest Diagenesis in Scleractinian Coral Skeletons: Implications for Palaeoclimate-Sensitive Geochemical Archives. Facies 2009, 55, 161–201. [Google Scholar] [CrossRef]
- Harmelin-Vivien, M.L. The Effects of Storms and Cyclones on Coral Reefs: A Review. J. Coast. Res. Spec. Issue 1994, 12, 211–231. [Google Scholar]
- Littler, M.M.; Littler, D.S. Impact of CLOD Pathogen on Pacific Coral Reefs. Science 1995, 267, 1356–1360. [Google Scholar] [CrossRef]
- Harrington, L.; Fabricius, K.; De’ath, G.; Negri, A. Recognition and Selection of Settlement Substrata Determine Post-Settlement Survival in Corals. Ecology 2004, 85, 3428–3437. [Google Scholar] [CrossRef]
- Hayakawa, J.; Kawamura, T.; Ohashi, S.; Horii, T.; Watanabe, Y. Habitat Selection of Japanese Top Shell (Turbo Cornutus) on Articulated Coralline Algae: Combination of Preferences in Settlement and Post-Settlement Stage. J. Exp. Mar. Biol. Ecol. 2008, 363, 118–123. [Google Scholar] [CrossRef]
- Scoffin, T.P.; Alexandersson, E.T.; Bowes, G.E.; Clokie, J.J.; Farrow, G.E.; Milliman, J.D. Recent, Temperate, Sub-Photic, Carbonate Sedimentation: Rockall Bank, Northeast Atlantic. J. Sediment. Petrol. 1980, 50, 331–356. [Google Scholar]
- Eakin, C.M. Where Have All the Carbonates Gone? A Model Comparison of Calcium Carbonate Budgets before and after the 1982-1983 El Niño at Uva Island in the Eastern Pacific. Coral Reefs 1996, 15, 109–119. [Google Scholar]
- Glynn, P.W. State of Coral Reefs in the Galápagos Islands: Natural vs Anthropogenic Impacts. Mar. Pollut. Bull. 1994, 29, 131–140. [Google Scholar] [CrossRef]
- Raghukumar, C.; Sharma, S.; Lande, V. Distribution and Biomass Estimation of Shell-Boring Algae in the Intertidal at Goa, India. Phycologia 1991, 30, 303–309. [Google Scholar] [CrossRef]
- Alfaro, A.C.; Webb, S.C.; Barnaby, C. Variability of Growth, Health, and Population Turnover within Mussel Beds of Perna Canaliculus in Northern New Zealand. Mar. Biol. Res. 2008, 4, 376–383. [Google Scholar] [CrossRef]
- Ndhlovu, A.; McQuaid, C.D.; Nicastro, K.; Marquet, N.; Gektidis, M.; Monaco, C.J.; Zardi, G. Biogeographical Patterns of Endolithic Infestation in an Invasive and an Indigenous Intertidal Marine Ecosystem Engineer. Diversity 2019, 11, 75. [Google Scholar] [CrossRef] [Green Version]
- Gardner, J.; Thomas, M. Growth, Mortality and Production of Organic Matter by a Rocky Intertidal Population of Mytilus Edulis in the Quoddy Region of the Bay of Fundy. Mar. Ecol. Prog. Ser. 1987, 39, 31–36. [Google Scholar] [CrossRef]
- Griffiths, C.L.; King, J.A. Energy Expended on Growth and Gonad Output in the Ribbed Mussel Aulacomya Ater. Mar. Biol. 1979, 53, 217–222. [Google Scholar] [CrossRef]
- Wahl, M. Ecological Lever and Interface Ecology: Epibiosis Modulates the Interactions between Host and Environment. Biofouling 2008, 24, 427–438. [Google Scholar] [CrossRef]
- Thieltges, D.W.; Buschbaum, C. Vicious Circle in the Intertidal: Facilitation between Barnacle Epibionts, a Shell Boring Polychaete and Trematode Parasites in the Periwinkle Littorina Littorea. J. Exp. Mar. Biol. Ecol. 2007, 340, 90–95. [Google Scholar] [CrossRef]
- Gutiérrez, J.L.; Jones, C.G.; Strayer, D.L.; Iribarne, O.O. Mollusks as Ecosystem Engineers: The Role of Shell Production in Aquatic Habitats. Oikos 2003, 101, 79–90. [Google Scholar] [CrossRef]
- Witman, J.D. Refuges, Biological Disturbance, and Rocky Subtidal Community Structure in New England. Ecol. Monogr. 1985, 55, 421–445. [Google Scholar] [CrossRef]
- Suchanek, T.H. Mussels and Their Role in Structuring Rocky Shore Communities. In The Ecology of Rocky Coasts; Moore, P.J., Seed, R., Eds.; Colombia Univ. Press: New York, NY, USA, 1986; pp. 70–96. [Google Scholar]
- Stephens, E.G.; Bertness, M.D. Mussel Facilitation of Barnacle Survival in a Sheltered Bay Habitat. J. Exp. Mar. Biol. Ecol. 1991, 145, 33–48. [Google Scholar] [CrossRef]
- Jurgens, L.J.; Gaylord, B. Physical Effects of Habitat-Forming Species Override Latitudinal Trends in Temperature. Ecol. Lett. 2018, 21, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Thiel, M.; Dernedde, T. Recruitment of Shore Crabs Carcinus Maenas on Tidal Flats: Mussel Clumps as an Important Refuge for Juveniles. Helgoländer Meeresunters. 1994, 48, 321–332. [Google Scholar] [CrossRef]
- Tsuchiya, M.; Nishihira, M. Islands of Mytilus as a Habitat for Small Intertidal Animals: Effect of Island Size on Community Structure. Mar. Ecol. Prog. Ser. 1985, 25, 71–81. [Google Scholar] [CrossRef]
- Piazza, B.P.; Banks, P.D.; La Peyre, M.K. The Potential for Created Oyster Shell Reefs as a Sustainable Shoreline Protection Strategy in Louisiana. Restor. Ecol. 2005, 13, 499–506. [Google Scholar] [CrossRef]
- Borsje, B.W.; van Wesenbeeck, B.K.; Dekker, F.; Paalvast, P.; Bouma, T.J.; van Katwijk, M.M.; de Vries, M.B. How Ecological Engineering Can Serve in Coastal Protection. Ecol. Eng. 2011, 37, 113–122. [Google Scholar] [CrossRef]
- Peck, L.S.; Clarke, A.; Holmes, L.J. Size, Shape and the Distribution of Organic Matter in the Recent Antarctic Brachiopod Liothyrella Uva. Lethaia 1987, 20, 33–40. [Google Scholar] [CrossRef]
- Walker, S.E. Endobionts on Modern and Fossil Turritella from the Northern Gulf of California Region. Ichnos 1998, 6, 99–115. [Google Scholar] [CrossRef]
- Day, E.G.; Branch, G.M.; Viljoen, C. How Costly Is Molluscan Shell Erosion? A Comparison of Two Patellid Limpets with Contrasting Shell Structures. J. Exp. Mar. Biol. Ecol. 2000, 243, 185–208. [Google Scholar] [CrossRef]
- Parke, M.W.; Moore, H.B. The Biology of Balanus Balanoides. II. Algal Infection of the Shell. J. Mar. Biol. Assoc. UK 1935, 20, 49–56. [Google Scholar] [CrossRef]
- Peebles, M.W.; Lewis, R.D. Differential Infestation of Shallow-Water Benthic Foraminifera by Microboring Organisms: Possible Biases in Preservation Potential. PALAIOS 1988, 3, 345. [Google Scholar] [CrossRef]
- Kobluk, D.R.; Risk, M.J. Calcification of Exposed Filaments of Endolithic Algae, Micrite Envelope Formation and Sediment Production. SEPM J. Sediment. Res. 1977, 47, 517–528. [Google Scholar] [CrossRef]
- Bathurst, R.G.C. Boring Algae, Micrite Envelopes and Lithification of Molluscan Biosparites. Geol. J. 1966, 5, 15–32. [Google Scholar] [CrossRef]
- Raven, J.A. Ocean Acidification Due to Increasing Atmospheric Carbon Dioxide; The Royal Society: London, UK, 2005. [Google Scholar]
- Andersson, A.J. Coastal Ocean and Carbonate Systems in the High CO2 World of the Anthropocene. Am. J. Sci. 2005, 305, 875–918. [Google Scholar] [CrossRef]
- Schönberg, C.H.L.; Fang, J.K.H.; Carreiro-Silva, M.; Tribollet, A.; Wisshak, M. Bioerosion: The Other Ocean Acidification Problem. ICES J. Mar. Sci. 2017, 74, 895–925. [Google Scholar] [CrossRef]
- Mendoza, V.; Pazos, M.; Garduño, R.; Mendoza, B. Thermodynamics of Climate Change between Cloud Cover, Atmospheric Temperature and Humidity. Sci. Rep. 2021, 11, 21244. [Google Scholar] [CrossRef]
- Gruber, N.; Sarmiento, J.L.; Stocker, T.F. An Improved Method for Detecting Anthropogenic CO2 in the Oceans. Glob. Biogeochem. Cycles 1996, 10, 809–837. [Google Scholar] [CrossRef]
- Caldeira, K.; Wickett, M.E. Anthropogenic Carbon and Ocean PH. Nature 2003, 425, 365. [Google Scholar] [CrossRef] [PubMed]
- Sabine, C.L.; Feely, R.A.; Gruber, N.; Key, R.M.; Lee, K.; Bullister, J.L.; Wanninkhof, R.; Wong, C.S.; Wallace, D.W.R.; Tilbrook, B.; et al. The Oceanic Sink for Anthropogenic CO2. Science 2004, 305, 367–371. [Google Scholar] [CrossRef]
- Key, R.M.; Kozyr, A.; Sabine, C.L.; Lee, K.; Wanninkhof, R.; Bullister, J.L.; Feely, R.A.; Millero, F.J.; Mordy, C.; Peng, T.-H. A Global Ocean Carbon Climatology: Results from Global Data Analysis Project (GLODAP). Glob. Biogeochem. Cycles 2004, 18, GB4031. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O. Climate Change, Coral Bleaching and the Future of the World’s Coral Reefs. Mar. Freshw. Res. 1999, 50, 839–866. [Google Scholar] [CrossRef]
- Gattuso, J.-P.; Allemand, D.; Frankignoulle, M. Photosynthesis and Calcification at Cellular, Organismal and Community Levels in Coral Reefs: A Review on Interactions and Control by Carbonate Chemistry. Am. Zool. 1999, 39, 160–183. [Google Scholar] [CrossRef]
- Petes, L.E.; Menge, B.A.; Murphy, G.D. Environmental Stress Decreases Survival, Growth, and Reproduction in New Zealand Mussels. J. Exp. Mar. Biol. Ecol. 2007, 351, 83–91. [Google Scholar] [CrossRef]
- O’Donnell, M.J.; George, M.N.; Carrington, E. Mussel Byssus Attachment Weakened by Ocean Acidification. Nat. Clim. Chang. 2013, 3, 587–590. [Google Scholar] [CrossRef]
- Enochs, I.C.; Manzello, D.P.; Tribollet, A.; Valentino, L.; Kolodziej, G.; Donham, E.M.; Fitchett, M.D.; Carlton, R.; Price, N.N. Elevated Colonization of Microborers at a Volcanically Acidified Coral Reef. PLoS ONE 2016, 11, e0159818. [Google Scholar] [CrossRef]
- Marcelino, V.R.; Morrow, K.M.; Oppen, M.J.H.; Bourne, D.G.; Verbruggen, H. Diversity and Stability of Coral Endolithic Microbial Communities at a Naturally High pCO2 Reef. Mol. Ecol. 2017, 26, 5344–5357. [Google Scholar] [CrossRef]
- Tribollet, A.; Atkinson, M.J.; Christopher, L. Effects of Elevated pCO2 on Epilithic and Endolithic Metabolism of Reef Carbonates. Glob. Chang. Biol. 2006, 12, 2200–2208. [Google Scholar] [CrossRef]
- Tribollet, A.; Godinot, C.; Atkinson, M.; Langdon, C. Effects of Elevated pCO2 on Dissolution of Coral Carbonates by Microbial Euendoliths. Glob. Biogeochem. Cycles 2009, 23, 3008. [Google Scholar] [CrossRef]
- Tribollet, A.; Chauvin, A.; Cuet, P. Carbonate Dissolution by Reef Microbial Borers: A Biogeological Process Producing Alkalinity under Different pCO2 Conditions. Facies 2019, 65, 9. [Google Scholar] [CrossRef]
- Diaz-Pulido, G.; Anthony, K.R.N.; Kline, D.I.; Dove, S.; Hoegh-Guldberg, O. Interactions between Ocean Acidification and Warming on the Mortality and Dissolution of Coralline Algae. J. Phycol. 2012, 48, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Leggat, W.P.; Camp, E.F.; Suggett, D.J.; Heron, S.F.; Fordyce, A.J.; Gardner, S.; Deakin, L.; Turner, M.; Beeching, L.J.; Kuzhiumparambil, U.; et al. Rapid Coral Decay Is Associated with Marine Heatwave Mortality Events on Reefs. Curr. Biol. 2019, 29, 2723–2730.e4. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Responses to Endolithic Infestation | Live Calcifying Hosts | References | |||
|---|---|---|---|---|---|
| Corals | Coralline Algae | Bivalves | Others | ||
| Physiological Parameters | |||||
| Growth | ↓ = | ↓ | ↓ | [22,24,143,167,168,169] | |
| General condition | = | ↓ | [22,96,107,167,170,171,172] | ||
| Reproduction | ↓ | = | [22,168,173,174] | ||
| Attachment strength | ↓ | [107,172,173] | |||
| General survival | ↑ = | ↓ ✞ | ↓ | [14,22,73,143,144,170,171,173,175,176] | |
| Individual survival to heat stress | ↑ (lim) | ↑ (lim) | [23,25,33,108,109,115,177,178] | ||
| Calcified structures | |||||
| Microbioerosion | ↑ | ↑↓ | ↑ | ↑ | [13,15,22,108,129,143,169,179] |
| Thickness | ↑ | ↓ ✞ | ↓ | [22,24,98,143] | |
| Strength | ↓ | ↓ ✞ | ↓ | [22,107,143,169,172,174,175,176,180] | |
| Porosity | ↑ | ↑ | ↑ | [13,15,24,108] | |
| Deformations | ↑ | ↑ ✞ | ↑ | [14,24,96,174,181,182,183] | |
| Maintenance costs | ↑ | ↑ | ↑ | [22,24,96,98,143,168,181] | |
| Mineralogy | ~ | ~ | [96,135] | ||
| Biological interactions | |||||
| Epibionts | ↑ | [172] | |||
| Predators | ↑ | ↑ | [107] | ||
| Grazers | ↑ | ↑ | ↑ | [151,176] | |
| Photoautotrophic euendoliths | ↔ | ↔ | ↔ | [15,24,108,144,170,171] | |
| Bioengineered ecosystems | |||||
| Architectural complexity | ↑↓ | ↓ | [129,135] | ||
| Coastal protection from waves and other stressors | ↓ | ↑↓ | ↓ | ↓ | [107,129,135,174,175,179] |
| Mitigation of environmental stressors for associated species | ↑ | [33,109,115] | |||
| Resistance to anthropogenic stressors | ↓ | [175,184] | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dievart, A.M.; McQuaid, C.D.; Zardi, G.I.; Nicastro, K.R.; Froneman, P.W. Photoautotrophic Euendoliths and Their Complex Ecological Effects in Marine Bioengineered Ecosystems. Diversity 2022, 14, 737. https://doi.org/10.3390/d14090737
Dievart AM, McQuaid CD, Zardi GI, Nicastro KR, Froneman PW. Photoautotrophic Euendoliths and Their Complex Ecological Effects in Marine Bioengineered Ecosystems. Diversity. 2022; 14(9):737. https://doi.org/10.3390/d14090737
Chicago/Turabian StyleDievart, Alexia M., Christopher D. McQuaid, Gerardo I. Zardi, Katy R. Nicastro, and Pierre W. Froneman. 2022. "Photoautotrophic Euendoliths and Their Complex Ecological Effects in Marine Bioengineered Ecosystems" Diversity 14, no. 9: 737. https://doi.org/10.3390/d14090737
APA StyleDievart, A. M., McQuaid, C. D., Zardi, G. I., Nicastro, K. R., & Froneman, P. W. (2022). Photoautotrophic Euendoliths and Their Complex Ecological Effects in Marine Bioengineered Ecosystems. Diversity, 14(9), 737. https://doi.org/10.3390/d14090737