

Inventory and Ecological Characterization of Ichthyofauna of Nine Lakes in the Adamawa Region (Northern Cameroon, Central Africa)

 , , and
, , and
Abstract
:1. Introduction
2. Material and Methods
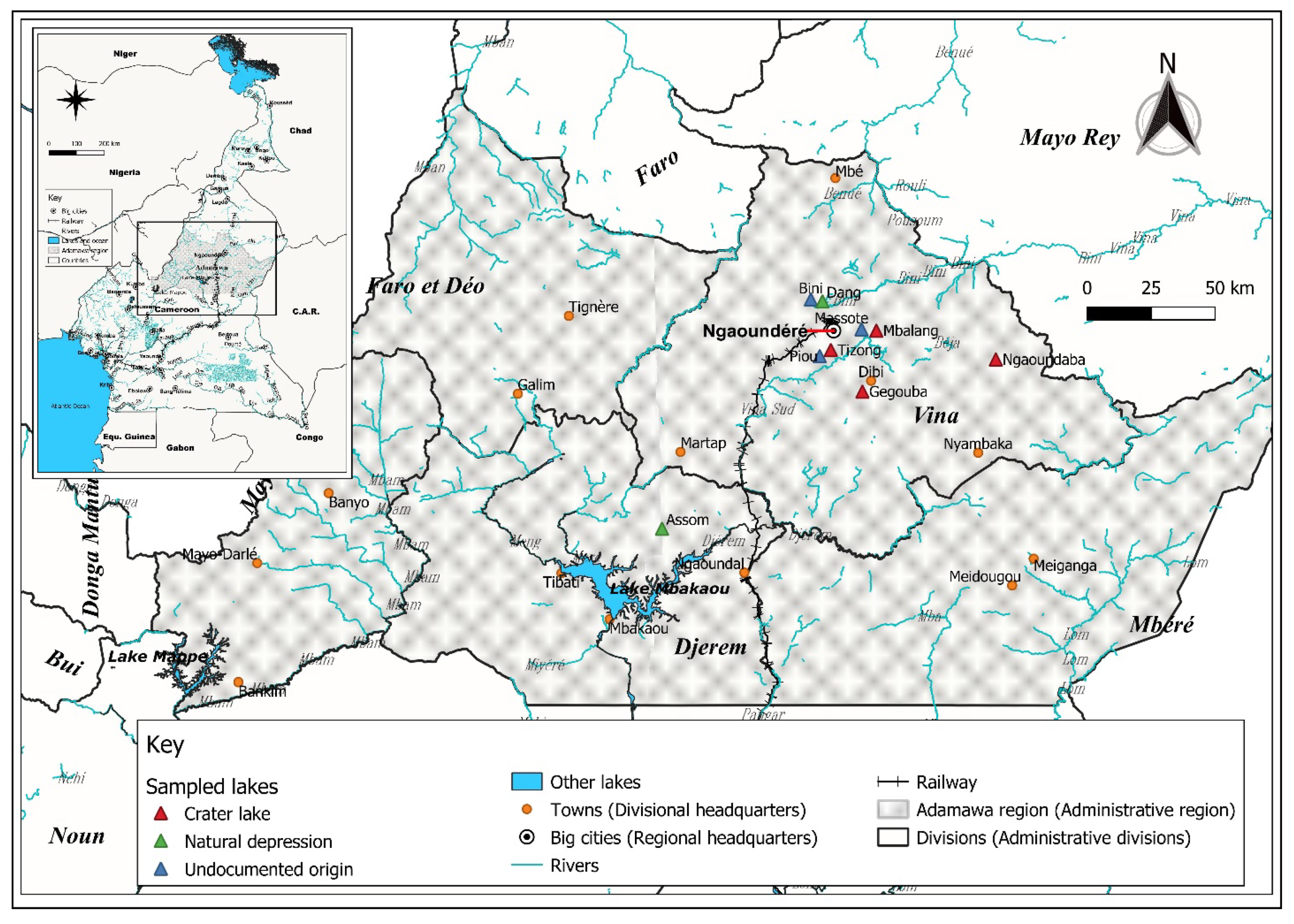
2.1. Study Sites
2.2. Data Collection
2.2.1. Measurement of Environmental Variables
2.2.2. Capture and Storage of Fish Samples
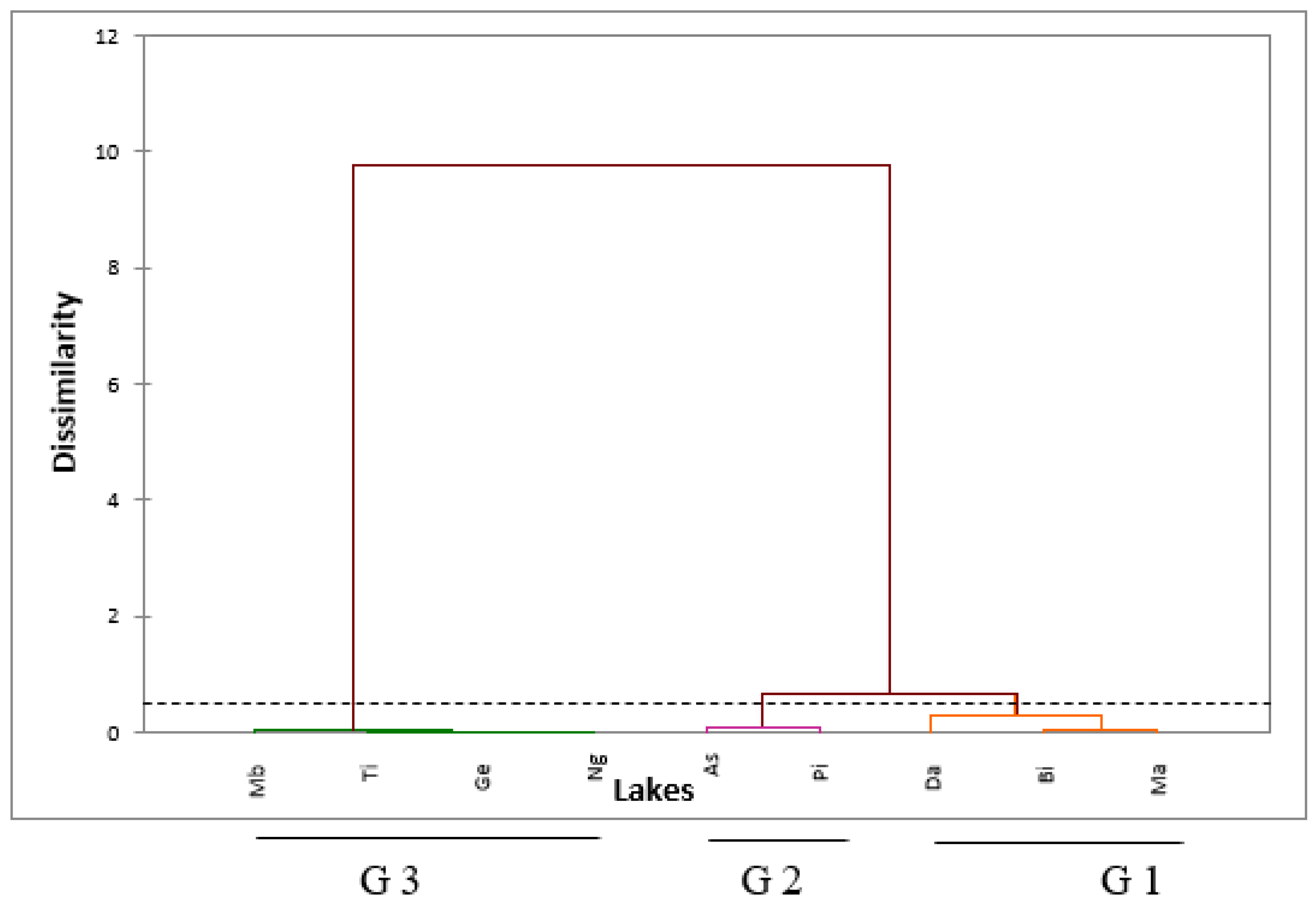
2.3. Data Analysis
3. Results
3.1. Physicochemical Characteristics of the Waters of the Lakes Studied
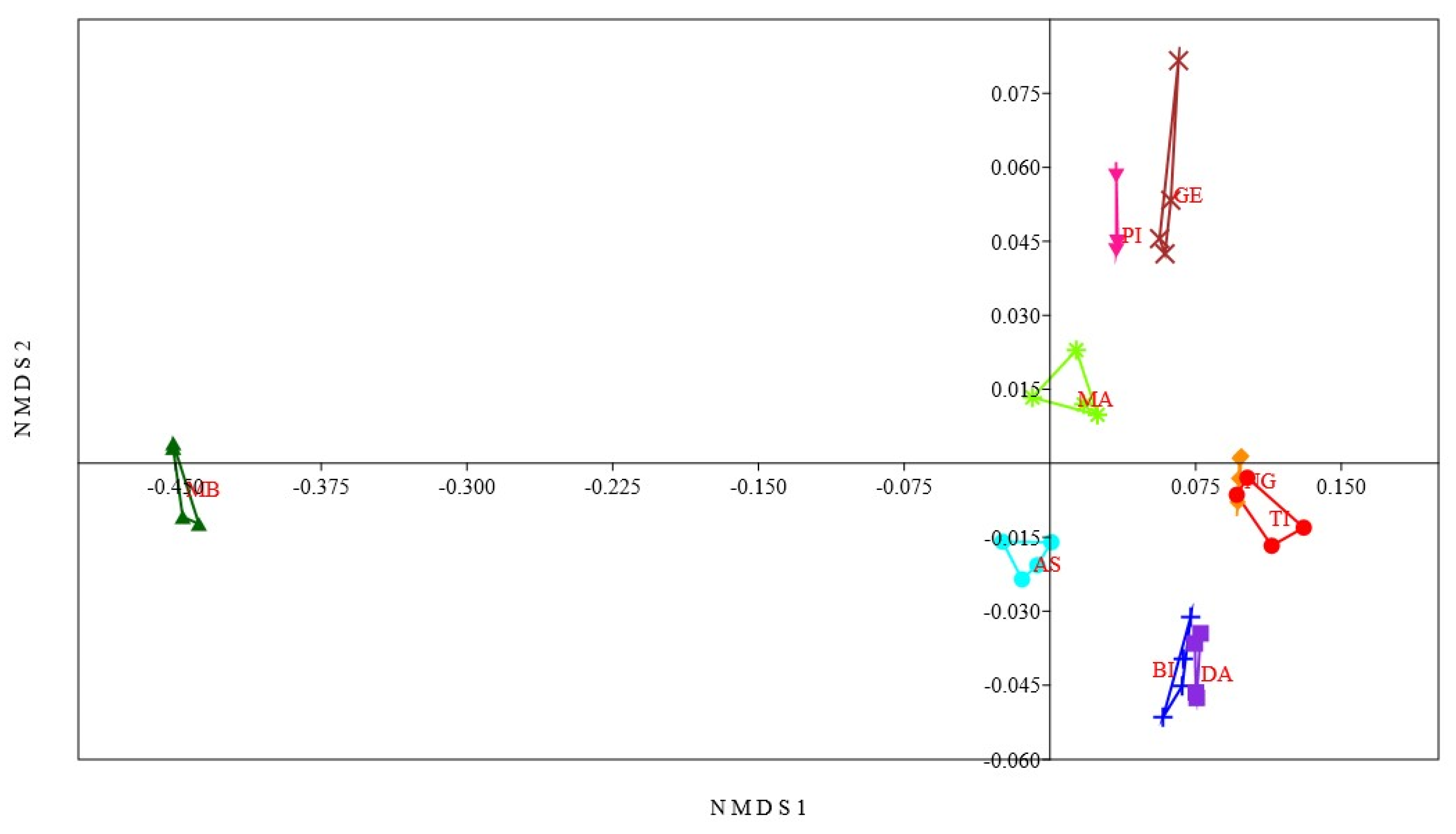
3.2. Relation between Lakes Based on Physicochemical Parameters
3.3. Abundance of Taxa
3.4. Frequency of Trophic Groups
3.5. Frequency of Breeding Groups
3.6. Ecological Diversity of Fish Communities
3.7. Patterns of Fish Species Diversity and Its Distribution
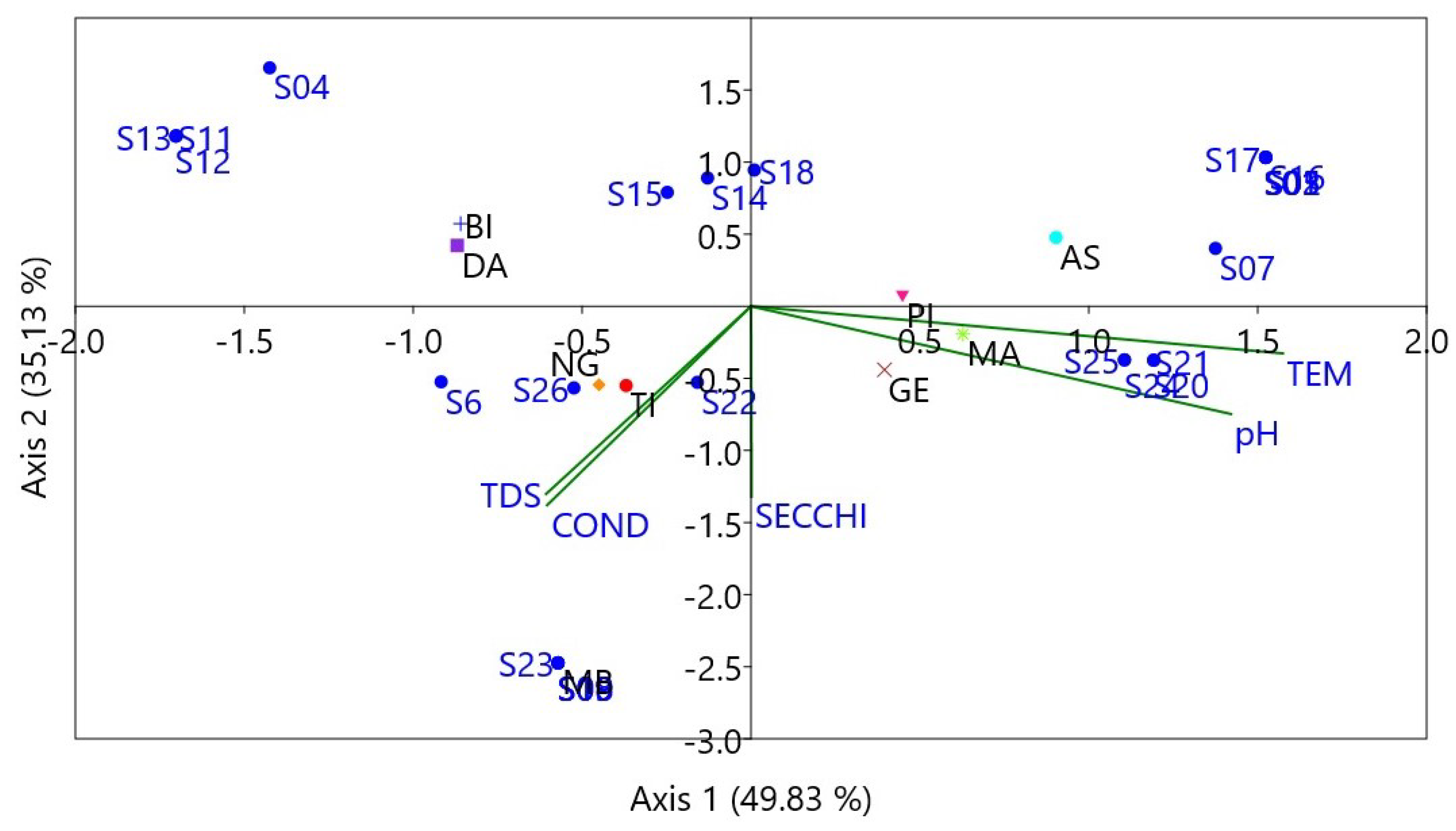
3.8. Relationships between Environmental Variables and Fish Species
4. Discussion
4.1. Comparative Limnology
4.2. Fish Species Composition and Distribution
4.3. Specific Wealth
4.4. Origin of Ichthyofauna and Speciation in the Adamawa Lakes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miqueleiz, I.; Böhm, M.; Ariño, A.H.; Miranda, R. Assessment Gaps and Biases in Knowledge of Conservation Status of Fishes. Aquat. Conserv. 2020, 30, 225–236. [Google Scholar] [CrossRef]
- Barbarossa, V.; Bosmans, J.; Wanders, N.; King, H.; Bierkens, M.F.P.; Huijbregts, M.A.J.; Schipper, A.M. Threats of Global Warming to the World’s Freshwater. Fishes. Nat. Commun. 2021, 12, 1–10. [Google Scholar] [CrossRef]
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.J.; Kidd, K.A.; MacCormack, T.J.; Olden, J.D.; Ormerod, S.J.; et al. Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol. Rev. 2019, 94, 849–873. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.P.; McCormick, M.I.; Srinivasan, M.; Eagle, J.V. Coral decline threatens fish biodiversity in marine reserves. Proc. Natl. Acad. Sci. USA 2004, 101, 8251–8253. [Google Scholar] [CrossRef] [PubMed]
- Lalèyè, P.A.; Philipart, J.C.; Heymans, J.C. Cycle aannuel de l’indice et de la condition chez deux espèces de Chrysichthys (Siluriformes, Bagridae) au lac Nokoué et la lagune de Porto -Novo. Cybium 1995, 19, 131–142. [Google Scholar]
- Lévêque, C.; Paugy, D. Impacts des activités humaines. In Les Poissons d’Eaux Continentales Africaines: Diversité, Ecologie, Utilisation par l’Homme; IRD: Paris, France, 2006; pp. 395–413. [Google Scholar]
- Kling, G.W. Comparative transparency, depth of mixing, and stability of stratification in lakes of Cameroon, West Africa. Limnol. Oceanogr. 1988, 33, 27–40. [Google Scholar] [CrossRef]
- Bitja Nyom, A.R. Taxonomy, Phylogeny and Biogeography of Cichlids (Teleostei, Perciformes) from Cameroon South of the North Parallel 8th. Ph.D. Thesis, Unpublished Thesis. University of Yaoundé I, Yaoundé, Cameroon, 2012; 261p. [Google Scholar]
- Paugy, D.; Levêque, C. Fish communities in small aquatic ecosystems: Caves, gueltas, crater and salt lakes. In The Inland Water Fishes of Africa Diversity, Ecology and Human Use; IRD: Paris, France, 2017; pp. 397–416. [Google Scholar]
- Cornen, G.; Bande, Y.; Giresse, P.; Maley, J. The nature and chronostratigraphy of Quaternary pyroclastic accumulations from Lake Barombi Mbo (West-Cameroon). J. Volcanol. Geotherm. Res. 1992, 51, 357–374. [Google Scholar] [CrossRef]
- Richards, E.J.; Poelstra, J.W.; Martin, C.H. Don’t throw out the sympatric speciation with the crater lake water: Fine-scale investigation of introgression provides equivocal support for causal role of secondary gene flow in one of the clearest examples of sympatric speciation. Evol. Let. 2018, 2, 524–540. [Google Scholar] [CrossRef]
- Schliewen, U.K.; Klee, B. Reticulate sympatric speciation in Cameroonian crater lake cichlids. Front. Zool. 2004, 1, 5. [Google Scholar] [CrossRef]
- Trewavas, E. Fishes of the crater lakes of the northwestern Cameroons. Bonn. Zool. Beitr. 1962, 13, 146–190. [Google Scholar]
- Baldo, L.; Pretus, J.L.; Riera, J.L.; Musilova, Z.; Bitja Nyom, A.R.; Salzburger, W. Convergence of gut microbiotas in the adaptive radiations of African cichlid fishes. ISME J. 2017, 11, 1975–1987. [Google Scholar] [CrossRef] [PubMed]
- Galvez, J.R.; McLean, K.; Touokong, D.C.; Gonwouo, N.L.; Martin, C.H. Trophic Specialization on unique resources despite limited niche divergence in a celebrated example of sympatric speciation. Ecol. Freshw. Fish. 2021, 00, 1–18. [Google Scholar] [CrossRef]
- Trewavas, E.; Green, J.; Corbet, S.A. Ecological studies on crater lakes in West Cameroon, Fishes of Barombi Mbo. J. Zool. Soc. Lond. 1972, 167, 41–95. [Google Scholar] [CrossRef]
- Stiassny, M.L.J.; Schliewen, U.K.; Dominey, W.J. A new species flock of cichlid fishes from Lake Bermin, Cameroon with a description of eight new species of Tilapia (Labroidei: Cichlidae). Ichthyol. Explor. Freshw. 1992, 34, 311–346. [Google Scholar]
- Dunz, A.R.; Schliewen, U.K. Description of a Tilapia (Coptodon) species flock of lake Ejagham (Cameroon), including a redescription of Tilapia deskerti Thys van den Audenaerde. Spixiana 2010, 33, 251–280. [Google Scholar]
- Neumann, D.; Stiassny, M.L.J.; Schliewen, U.K. Two new sympatric Sarotherodon species (Pisces: Cichlidae) endemic to lake Ejagham, Cameroon, west-central Africa, with comments on the Sarotherodon galilaeus species complex. Zootaxa 2011, 2765, 1–20. [Google Scholar] [CrossRef]
- Kusakabe, M. Lakes Nyos and Monoun gas disasters (Cameroon)- Limnic eruptions caused by excessive accumulation of magmatic CO2 in crater lakes. Geochem. Monogr. Ser. 2017, 1, 1–50. [Google Scholar] [CrossRef]
- Martin, C.H.; Cutler, J.S.; Friel, J.P.; Touokong, D.C.; Coop, G.; Wainwright, P.C. Complex histories of repeated gene flow in Cameroon crater lake cichlids cast doubt on one of the clearest examples of sympatric speciation. Evolution 2015, 69, 1406–1422. [Google Scholar] [CrossRef]
- Ramsar, Ramsar Sites Information Service. 2006. Available online: https://rsis.ramsar.org/ris/1643?language=en (accessed on 11 August 2022).
- Starnes, T.; Spiliopoulou, K. Zones clés pour la biodiversité. In Identification et Validation des Zones clés pour la Biodiversité des eaux douces d’Afrique Occidentale; UICN: Gland, Suisse, 2021; pp. 116–118. [Google Scholar]
- Stagera, J.C.; Alton, K.; Martin, C.H.; King, D.T., Jr.; Petruny, L.W.; Wiltse, B.; Livingstone, D.A. On the Age and Origin of Lake Ejagham, Cameroon, and Its Endemic Fishes. Quat. Res. 2017, 89, 1–12. [Google Scholar] [CrossRef]
- Schliewen, U.K.; Rassmann, K.; Markmann, M.; Markert, J.; Kocher, T.; Tautz, D. Genetic and ecological divergence of a monophyletic cichlid species pair under fully sympatric conditions in Lake Ejagham, Cameroon. Mol. Ecol. 2001, 10, 1471–1488. [Google Scholar] [CrossRef]
- Freeth, S.J. Life (and death) in Lake Nyos. Nature 1987, 329, 491. [Google Scholar] [CrossRef]
- Ohba, T.; Oginuma, Y.; Saiki, K.; Kusakabe, M.; Issa Fouepe, T.A.; Ntchantcho, R.; Tanyileke, G.; Hell, J.V.A. Depression Containing CO2-Enriched Water at the Bottom of Lake Monoun, Cameroon, and Implications for the 1984 Limnic Eruption. Front. Earth Sci. 2022, 10, 766–791. [Google Scholar] [CrossRef]
- Stiassny, M.L.J.; Teugels, G.; Hopkins, C. Poissons d’eaux Douces et Saumâtres de Basse Guinée, ouest de l’Afrique Centrale, 2nd ed.; MRAC/IRD: Tervuren, Paris; Belgique, France, 2007; Volumes 1 and 2.
- Halbwachs, M.; Sabroux, J.C.; Grangeon, J.; Kayser, G.; Tochon-Danguy, J.C.; Felix, A.; Béard, J.C.; Villevieille, A.; Vitter, G.; Richon, P.; et al. Degassing the Killer Lakes Nyos and Monoun. Eos 2011, 85, 281–285. [Google Scholar] [CrossRef]
- Bitja Nyom, A.R.; Kitio, H.I.; Njom, S.D.; Snoeks, J.; Pariselle, A.; Bilong Bilong, C.F. Morphological variations, length-weight relationships and condition factors of Hemichromis camerounensis (Cichliformes, Cichlidae) in three lakes in northern Cameroon. Int. J. Fish. Aquat. Stud. 2021, 9, 139–145. [Google Scholar] [CrossRef]
- Ngos, S., III; Giresse, P.; Maley, J. Palaeoenvironments of Lake Assom near Tibati (south Adamawa, Cameroon), What hap pened in Tibati around 1700 years BP. J. Afr. Earth Sci. 2003, 37, 35–45. [Google Scholar] [CrossRef]
- Giresse, P. Tropical and Sub-Tropical West Africa—Marine and Continental Changes during the Late Quaternary, 1st ed.; Elsevier Science: Amsterdam Pays-Bas, The Netherlands, 2007. [Google Scholar]
- Olivry, J.C. Fleuves et rivières du Cameroun; MESRES-ORTSOM: Paris, France, 1986; 754p. [Google Scholar]
- Letouzey, R. Etude phytogéographique du Cameroun; P. Lechevalier: Paris, France, 1968; 511p. [Google Scholar]
- Oumar, B.; Ekengéle, N.L.; Balla, O.A.D. Evaluation du niveau de pollution par les métaux lourds des lacs Bini et Dang région de l’Adamaoua—Cameroun. Afr. Sci. 2014, 102, 184–198. [Google Scholar]
- Daïwe, N.; Ngounou Ngatcha, B. The study of silting in the lake of Dang (Ngaoundéré, Cameroon) and the estimation of the suspended sediment transport. Pangea 2010, 47/48, 63–67. [Google Scholar]
- Wirrmann, D. Les lacs Baledjam, Gagouba, Mbalang, Tabéré et Tizon, région de Ngoundéré, province de l’Adamaoua; ORSTOM: Yaoundé, Cameroon, 1994; 16p. [Google Scholar]
- Nguetsop, V.F.; Bentaleb, I.; Favier, C.; Martin, C.; Bietrix, C.; Giresse, P.; Servant-Vildary, S.; Servant, M. Past environmental and climatic changes during the last 7200 cal yr BP in Adamawa plateau (Northern-Cameroun) based on fossil diatoms and sedimentary carbon isotopic records from Lake Mbalang. Clim. Past. 2011, 7, 1371–1393. [Google Scholar] [CrossRef]
- Bessa, E.A.Z.; Armstrong-Altrin, J.S.; Calistus, F.G.; Betsi, B.T.; Kelepile, T.; Ndjigui, P.D. Mineralogy and geochemistry of the Ngaoundaba Crater Lake sediments, northern Cameroon: Implications for provenance and trace metals status. Acta Geochim. 2021, 40, 718–738. [Google Scholar] [CrossRef]
- Bitja Nyom, A.R. Etat Initial des Peuplements Piscicoles. Rapport Final. 2015. Available online: http://documents.worldbank.org/curated/pt/930081498550117079/pdf/116 (accessed on 12 September 2021).
- Bitja Nyom, A.R.; Gilles, A.; Pariselle, A.; Snoeks, J.; Bilong Bilong, C.F. Divergences morphologiques allopatriques et sympatriques de Tilapia kottae (Perciformes, Cichlidae) endémique des lacs Barombi Kotto et Mboandong et affinités avec des tilapias de la Ligne Volcanique du Cameroun. Cybium 2012, 362, 335–348. [Google Scholar]
- Paugy, D.; Lévêque, C.; Teugels, G.G. Poissons d’eaux douces et saumâtres de l’Afrique de l’Ouest; IRD: Paris, France, 2003; Volume 1 et 2.
- Zhang, Z.; Grabchak, M. Entropic representation and estimation of diversity indices. J. Nonparametric. Stat. 2016, 28, 563–575. [Google Scholar] [CrossRef]
- Dajoz, R. Précis d’écologie, 7th ed.; Dunod: Paris, France, 2000. [Google Scholar]
- Abdel-Fattah, M.K.; Abd-Elmabod, S.K.; Aldosari, A.A.; Elrys, A.S.; Mohamed, E.S. Multivariate Analysis for Assessing Irrigation Water Quality: A Case Study of the Bahr Mouise Canal, Eastern Nile Delta. Water 2020, 12, 2537. [Google Scholar] [CrossRef]
- Hammam, A.A.; Mohamed, W.S.; Sayed, S.E.-E.; Kucher, D.E.; Mohamed, E.S. Assessment of Soil Contamination Using GIS and Multi-Variate Analysis: A Case Study in El-Minia Governorate, Egypt. Agronomy 2022, 12, 1197. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Austral. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Similarity-based testing for community pattern: The two-way layout with no replication. Mar. Boil. 1994, 118, 167–176. [Google Scholar] [CrossRef]
- Braak, C.J.E.; Verdonschot, P.E.M. Canonical correspondence analysis and related multivariate methods in aquatic ecology. Aquat. Sci. 1995, 57, 255–289. [Google Scholar] [CrossRef]
- Mamilov, N.; Sharakhmetov, S.; Amirbekova, F.; Bekkozhayeva, D.; Sapargaliyeva, N.; Kegenova, G.; Tanybayeva, A.; Abilkasimov, K. Past, Current and Future of Fish Diversity in the Alakol Lakes (Central Asia: Kazakhstan). Diversity 2022, 14, 11. [Google Scholar] [CrossRef]
- Krumhansl, K.; Jamieson, R.; Krkosek, W. Using species traits to assess human impacts on near shore benthic ecosystems in the Canadian. Arctic. Ecol. Indic. 2016, 60, 495–502. [Google Scholar] [CrossRef]
- Bliss, S.M.; Lennox, R.J.; Midwood, J.D.; Cooke, S.J. Temporally stable and distinct fish assemblages between stream and earthen stormwater drain reaches in an urban watershed. Urban Ecosyst. 2017, 20, 1045–1055. [Google Scholar] [CrossRef]
- Paugy, D.; Lévêque, C. Régimes alimentaires et réseaux trophiques. In Les Poissons d’Eaux Continentales Africaines: Diversité, Ecologie, Utilisation par l’Homme; IRD: Paris, France, 2006; pp. 191–215. [Google Scholar]
- Bitja Nyom, A.R.; Njom, S.D.; Kitio, H.I.; Bilong Bilong, C.F. Diversité de l’ichtyofaune de la rivière Djerem: Impact du barrage de Mbakaou et enjeux pour la conservation des poissons dans le Parc National du Mbam et Djerem (Cameroun). Int. J. Biol. Chem. Sci. 2020, 14, 1520–1535. [Google Scholar] [CrossRef]
- Ibanez, C.; Berdorff, T.; Teugels, G.; Mamononekene, V.; Lavoué, S.; Fermon, Y.; Paugy, D.; Toham, A.K. Fish assemblage structure and function along environmental gradients in rivers of Gabon (Africa). Ecol. Freshw. Fish 2007, 16, 315–334. [Google Scholar] [CrossRef]
- Øyvind, H. Paleontological Statistics Version 3.0. Reference Manual; Natural History Museum University of Oslo: Oslo, Norway, 2013; 221p. [Google Scholar]
- Chikou, A.; Lalèyè, P.A.; Raemakers, V.; Vandewalle, P.; Philippart, J.-C. Etude de l’âge et de la croissance chez Clarias gariepinus (Pisces, Clariidae) dans le delta de l’Ouémé au Benin (Afrique de l’Ouest). Int. J. Biol. Chem. Sci. 2008, 2, 157–167. [Google Scholar] [CrossRef]
- Lemoalle, J. La diversité des milieux aquatiques. In Les poissons d’eaux Continentales Africaines: Diversité, écologie, utilisation par l’homme; IRD: Paris, France, 2006; pp. 11–30. [Google Scholar]
- Lin, C.; Sovan, L.; Géraldine, L.; Sithan, L.; Zhongjie, L. Variations of fish composition and diversity related to environmental variables in shallow lakes in the Yangtze River basin. Aquat. Living Resour. 2011, 23, 417–426. [Google Scholar] [CrossRef]
- Kouamé, K.A.; Yao, S.S.; Gooré Bi, G.; Kouamélan, E.P.; N’Douba, V.; Kouassi, N.J. Influential environmental gradients and patterns of fish assemblages in a West African basin. Hydrobiologia 2008, 603, 159–169. [Google Scholar] [CrossRef]
- Aboua, B.R.D.; N’Zi, K.G.; Kouamélan, E.P.; Berté, S.; Bamba, M. Spatial organization of the fish population in Bandama. Int. J. Biol. Chem. Sci. 2010, 4, 1480–1493. [Google Scholar]
- Kamelan, T.M.; Yao, S.S.; Kouamé, K.A.; N’Zi, K.G.; Kouamélan, E.P. Ichtyofaune de la rivière Dodo (Côte d’Ivoire, Afrique de l’Ouest): Mise à jour et influence des variables environnementales sur la distribution des espèces. J. Appl. Biosci. 2013, 71, 5773–5785. [Google Scholar] [CrossRef]
- Kamdem, T.A.; Teugels, G.G. Patterns of microhabitat use among fourteen fishes of the Lower Ntem River Basin (Cameroon). Aquat. Living Resour. 1997, 10, 289–298. [Google Scholar] [CrossRef]
- Patrick, A.E.S. Influence of rainfall and water level on inland fisheries production: A review. Arch. Appl. Sci. Res. 2016, 8, 44–51. [Google Scholar]
- Hossain, M.S.; Das, N.G.; Sarker, S.; Rahaman, M.Z. Fish diversity and habitat relationship with environmental variables at Meghna river estuary, Bangladesh. Egypt. J. Aquat. Res. 2012, 38, 213–226. [Google Scholar] [CrossRef] [Green Version]
- Jean-Jacques, A. Fish communities in in estuaries and lagoons. In The Inland Water Fishes of Africa Diversity, Ecology and Human Use; IRD: Paris, France, 2017; pp. 417–440. [Google Scholar]
- Fermon, Y. Les poissons des grands lacs d’Afrique de l’Est. In Le Rift Est-Africain: Une Singularité Plurielle; IRD: Paris, France, 2009; pp. 213–222. [Google Scholar]
- Lévêque, C.; Quensière, J. Les peuplements ichtyologiques des lacs peu profonds. In Biology and Ecology of African Freshwater Fishes; Orstom: Paris, France, 1988; pp. 303–324. [Google Scholar]
- Snoeks, J. The Cichlid Diversity of Lake Malawi/Nyasa/Niassa: Identification, Distribution and Taxonomy; Cichlid Press: El Paso, TX, USA, 2004. [Google Scholar]
- Anabi, A.T.; Issiaka, Y. diversité ichtyologique d’un petit lac tropical: Cas du lac Madarounfa. Niger. Rev. Ivoir. Sci. Technol. 2021, 37, 143–154. [Google Scholar]
- Musilová, Z.; Indermaur, A.; Bitja-Nyom, A.R.; Omelchenko, D.; Kłodawska, M.; Albergati, L.; Remišová, K.; Salzburger, W. Evolution of visual sensory system in cichlid fishes from crater lake Barombi Mbo in Cameroon. Mol. Ecol. 2019, 28, 5010–5031. [Google Scholar] [CrossRef] [PubMed]
- Richards, K. Preliminary results of pollen analysis of a 6000 years core from Mboandong, a crater Lake in Cameroon. Hull Univ. Geogr. Dept. Misc. Ser. 1986, 32, 14–28. [Google Scholar]
- Maley, J.; Brenac, P. Analyses polliniques préliminaires du Quaternaire récent de l’Ouest du Cameroun: Mise en évidence des refuges forestiers et discussion des problèmes paléoclimatiques. Mém. Trav. E.P.H.E. Inst. Montpellier 1987, 17, 129–142. [Google Scholar]
- Giresse, P.; Maley, J.; Brenac, P. Late Quaternary paleoenvironments in the Lake Barombi Mbo (West Cameroon) deduced from pollen and carbon isotopes of organic matter. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1994, 107, 65–78. [Google Scholar] [CrossRef]
- Maley, J. The African rain forest vegetation and paleoenvironments during Late Quaternary. Clim. Chang. 1991, 19, 79–98. [Google Scholar] [CrossRef]
- Missoup, A.D. Systématique et biogéographie des rongeurs des forêts de la ligne volcanique du Cameroun (Afrique ouest centrale). Ph.D. These, Muséum National d’Histoire Naturelle, Paris, France, Université de Yaoundé, Yaoundé, Cameroon, 2010. [Google Scholar]
- Loheac, B.; Caudron, A.; Guillard, J. Communautés piscicoles introduites des lacs d’altitude: Approches scientifiques et influences idéologiques. Rev. Des. Sci. De L’Eau 2019, 32, 39–50. [Google Scholar] [CrossRef]
- Daget, J. Les poissons du Fouta Djalon et de la Basse Guinée. Mém. Inst. Franc Afrique Noire 1962, 65, 1–210. [Google Scholar]
- Lévêque, C.; Paugy, D. Les Poissons des eaux Continentales Africaines: Diversité, écologie, Utilisation par l’homme; IRD: Paris, France, 2006.
- Temdjim, R. The Volcanism of the Ngaoundere Region (Adamaoua-Cameroon). Volcanological and Petrological Study. Ph.D. Thesi, University of Clermont-Ferrand II, Clermont-Ferrand, France, 1986; 185p. [Google Scholar]
- Vreven, E.J.; Musschoot, T.; Boden, G.; Stiassny, M.L.J. Espèces étrangères ou introduites en basse Guinée. In Poissons d’Eaux Douces et Saumâtres de Basse Guinée, Ouest de l’Afrique Centrale; Stiassny, M., Teugels, G., Hopkins, C., Eds.; IRD: Paris, France; MRAC: Tervuren, Belgium; AMNH: New York, NY, USA, 2007. [Google Scholar]
- Lévêque, C. Les introductions d’espèces dans les milieux naturels et leurs conséquences. In Les Poissons d’Eaux Continentales Africaines: Diversité, Ecologie, Utilisation par l’Homme; Lévêque, C., Paugy, D., Eds.; IRD: Paris, France, 2006; pp. 382–394. [Google Scholar]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Res. 2005, 81, 163–182. [Google Scholar] [CrossRef]
- Leprieur, F.; Brosse, S.; Garcia-Berthou, E.; Oberdorff, T.; Olden, J.D.; Townsend, C.R. Scientific uncertainty and the assessment of risks posed by non-native freshwater fishes. Fish Fish. 2009, 10, 88–97. [Google Scholar] [CrossRef]
- De Vos, L.; Jos, S.; Dirk, F.E. Thys van den Audenaerde, The effects of Tilapia introductions in Lake Luhondo, Rwanda. Environ. Biol. Fishes 1990, 27, 303–308. [Google Scholar] [CrossRef]
- Canonico, G.C.; Arthington, A.; Mccrary, J.K.; Thieme, M. The effect of introduced tilapias on native biodiversity. Aquat. Conser. Mar. Fresh. Ecos. 2005, 15, 463–483. [Google Scholar] [CrossRef]
- Darwall, W.R.T.; Tweddle, D.; Skelton, P.H.; Smith, K.S. Background. In The Status and Distribution of Freshwater Biodiversity in Southern Africa; IUCN: Gland, Switzerland; SAIAB: Grahamstown, South Africa, 2009; pp. 1–15. [Google Scholar]
- Martens, K. Speciation in ancient lakes. Trends Ecol. Evol. 1997, 12, 177–182. [Google Scholar] [CrossRef]
- De Vos, L.; Snoeks, J. The non-cichlid fishes of the Lake Tanganyika basin. Arch. Für Hydrobiol. –Beih. Ergeb. Der Limnol. 1994, 44, 391–405. [Google Scholar]
- Snoeks, J. How well known is the Ichthyodiversity of the Large East African Lakes? Adv. Ecol. Res. 2000, 31, 17–38. [Google Scholar]
- Salzburger, W. Understanding explosive diversification through cichlid fish genomics. Nat. Rev. Genet. 2018, 19, 705–717. [Google Scholar] [CrossRef]
- Lévêque, C.; Fermon, Y.; Duponchelle, F. Communautés des lacs profonds In Les Poissons des eaux Continentales Africaines. Diversité, écologie, Utilisation par l’homme; IRD: Paris, France, 2006; pp. 321–340.
- Lavoué, S. A new species of Petrocephalus Marcusen 1854 (Osteoglossomorpha: Mormyridae) from the Sanaga River basin, Cameroon. Zootaxa 2011, 2934, 20–28. [Google Scholar] [CrossRef]
- Bitja Nyom, A.R.; Agnèse, J.-F.; Pariselle, A.; Bilong Bilong, C.F.; Gilles, A.; Snoeks, J. A systematic revision of the five-spotted Hemichromis complex (Cichiformes: Cichlidae) from West Africa and Lower Guinea, with the description of a new species from Cameroon. Hydrobiologia 2021, 848, 3779–3803. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lakes | Latitude (°N) | Longitude (°E) | Altitude (m) | Surface (ha) | Zm (m) | Z (m) | Hydrographic System |
|---|---|---|---|---|---|---|---|
| Bi *** | 07°25′54.01′′ | 13°30′26.63′′ | 1076 | 20 [7] | / | / | Lake Chad Basin [33] |
| Da ** | 07°25′29.01′′ | 13°32′54.01′′ | 1072 | 80 [7] | / | 2.5 [34] | Lake Chad Basin [33] |
| As ** | 06°37′22.36′′ | 12°58′56.51′′ | 898 | / | 3–4 [32] | / | Sanaga Basin [32] |
| Ge * | 07°06′23.14′′ | 13°41′23.9′′ | 1181 | 20 [7] | 104 [7] | / | Sanaga Basin [33] |
| Ma *** | 07°19′35.5′′ | 13°41′10.8′′ | 1073 | 5 [7] | / | / | Sanaga Basin [33] |
| Mb * | 07°19′17.53′′ | 13°44′18.73′′ | 1110 | 50 [7] | 52 [7] | / | Sanaga Basin [33] |
| Ng * | 07°13′11.57′′ | 13°69′37.01′′ | 1178 | 10 [7] | 62 [7] | / | Sanaga Basin [33] |
| Pi *** | 07°13′53.7′′ | 13°32′19.7′′ | 1185 | / | / | / | Sanaga Basin [33] |
| Ti * | 07°15′11.00′′ | 13°34′40.6′′ | 1065 | 8 [7] | 48 [7] | 26.2 [7] | Sanaga Basin [32] |
| Lakes | Parameters | ||||
|---|---|---|---|---|---|
| T (°C) | pH | TDS (ppm) | CON (µs/cm) | Secchi (m) | |
| As D | 25.95 ± 0.34 a | 8.26 ± 0.18 a | 8.6 ± 0.1 d | 14.5 ± 0.12 jk | 1.15 ± 0.09 g |
| As W | 26.1 ± 0.11 a | 5.98 ± 0.08 f | 8.11 ± 0.02 d | 12.17 ± 0.21 k | 0.7± 0.02 ij |
| Da D | 22.2 ± 0.5 f | 5.18 ±1.10 g | 14.89 ±1.41 d | 24.96± 0.78 fg | 0.45 ±0.02 kl |
| Da W | 23.74 ± 0.08 de | 5.06 ± 0.05 gh | 19.84 ± 1.14 d | 27.34 ± 1.75 f | 0.35 ± 0.01 l |
| Bi D | 23.55 ± 0.4 e | 5.13 ± 0.05 gh | 10.27 ± 0.28 d | 17.32 ± 0.3 ij | 0.55 ± 0.01 jk |
| Bi W | 24.59 ± 0.07 cd | 4.76 ± 0.06 h | 12.47 ± 0.76 d | 17.9 ± 1.24 hij | 0.4 ± 0.0 kl |
| Ge D | 25.75 ± 0.09 ab | 7.25 ± 0.02 cd | 121.15 ± 0.23 b | 171.43 ± 0.09 c | 6.25 ± 0.06 a |
| Ge W | 25.82 ± 0.1 a | 7.20 ± 0.03 cd | 120.12 ± 0.5 b | 169.62 ± 0.32 cd | 5.32 ± 0.04 b |
| Ma D | 25.35 ± 0.19 abc | 7.52 ± 0.02 bc | 14.31 ± 0.17 d | 22.512 ± 0.14 gh | 1.01 ± 0.03 gh |
| Ma W | 25.4 ± 0.1 abc | 8.56 ± 0.01 a | 14.8± 0.2 d | 21.8 ± 0.2 ghi | 0.78 ± 0.02 i |
| Mb D | 24.7 ± 0.01 c | 6.73 ± 0.05 e | 67.83 ± 12.88 c | 165.36 ± 0.1 d | 3.73 ± 0.06 e |
| Mb W | 25.38 ± 0.06 abc | 7.20 ± 0.02 cd | 114.45 ± 1.35 b | 157.65 ± 0.89 e | 3.33 ± 0.03 f |
| Ng D | 24.78 ± 0.12 bc | 7.27 ± 0.02 bcd | 121.71 ± 0.54 b | 170.45 ± 0.47 c | 4.5 ± 0.0 c |
| Ng W | 26 ± 0.07 a | 7.247 ± 0.02 cd | 121 ± 0.57 b | 169.05 ± 0.15 cd | 3.35 ± 0.03 f |
| Pi D | 25.49 ± 0.19 abc | 7.63 ± 0.01 bc | 8.41 ± 0.05 d | 17.75 ± 0.11 hij | 0.87 ± 0.02 hi |
| Pi W | 25.8 ± 0.1 a | 7.60 ± 0.24 bc | 11.31± d | 18.4 ± 0.88 hij | 0.73 ± 0.01 ij |
| Ti D | 22.08 ± 0.3 f | 7.747 ± 0.17 b | 165.05 ± 5.34 a | 273.5 ± 2.82 a | 4.18 ± 0.03 d |
| Ti W | 25.73 ± 0.11 ab | 6.97 ± 0.04 de | 181 ± 0.18 a | 256.475 ± 0.16 b | 3.38 ± 0.04 f |
| F | 41.92 | 158.83 | 337.8 | 9068.25 | 1992.41 |
| p | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Taxa | Ecological Features | Presence–Absence in the Station | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Orders Families | Species | BC | TG | RG | IUCN | As | Bi | Da | Ge | Ma | Mb | Ng | Pi | Ti | Code |
| Osteoglossiformes Mormyridae | Petrocephalus similis (Lavoué, 2011) | E | INV | SUB | NE | † | S01 | ||||||||
| Marcusenius mento (Boulenger, 1890) | N | INV | NES | LC | † | S02 | |||||||||
| Paramormyrops kingsleyae (Günther, 1896) | N | INV | NES | DD | † | S03 | |||||||||
| Brevimyrus niger (Günther, 1866) | N | INV | SUB | LC | † | S04 | |||||||||
| Brienomyrus brachyitius (Gill, 1862) | N | INV | SUB | LC | † | S05 | |||||||||
| Arapaimidae | Heterotis niloticus (Cuvier, 1829) | I | OMV | NES | LC | † | † | † | S06 | ||||||
| Cypriniformes Cyprinidae | Enteromius martorelli (Roman, 1971) | N | OMV | SUB | LC | † | † | S07 | |||||||
| Enteromius sp. «mbalang» | N | OMV | SUB | † | S08 | ||||||||||
| Garra cf ornata | N | HER | SUB | † | S09 | ||||||||||
| Garra cf dembeensis | N | HER | SUB | † | S10 | ||||||||||
| Anabantiformes Anabantidae | Ctenopoma sp. | N | INV | SUB | † | † | S11 | ||||||||
| Channidae | Parachanna obscura (Günther, 1861) | I | PIS | NES | LC | † | † | S12 | |||||||
| Perciformes Latidae | Lates niloticus (Linnaeus, 1758) | I | PIS | SUB | LC | † | † | S13 | |||||||
| Siluriformes Clariidae | Clarias camerunensis (Lönnberg, 1895) | N | OMV | SUB | LC | † | † | † | S14 | ||||||
| Clarias gariepinus (Burchell, 1822) | I | OMV | SUB | LC | † | † | † | † | S15 | ||||||
| Clarias jaensis (Boulenger, 1909) | N | OMV | SUB | LC | † | † | S16 | ||||||||
| Clarias longior (Boulenger, 1907) | N | OMV | SUB | LC | † | S17 | |||||||||
| Clarias pachynema (Boulenger, 1903) | N | OMV | SUB | LC | † | † | S18 | ||||||||
| Clarias sp. «mbalang» | N | OMV | SUB | † | S19 | ||||||||||
| Clarias sp. «massote» | N | OMV | SUB | † | S20 | ||||||||||
| Mochokidae | Synodontis rebeli (Holly, 1926) | E | OMV | SUB | DD | † | S21 | ||||||||
| Cichliformes Cichlidae | Oreochromis niloticus (Linnaeus, 1758) | I | HER | MOU | LC | † | † | † | † | † | † | † | † | S22 | |
| Oreochromis macrochir (Boulenger, 1912) | I | HER | MOU | LC | † | S23 | |||||||||
| Coptodon cameronensis (Holly, 1927) | E | HER | NES | LC | † | † | † | S24 | |||||||
| Coptodon rendalli (Boulenger, 1897) | I | HER | SUB | LC | † | † | † | † | S25 | ||||||
| Hemichromis camerounensis (Bitja Nyom et al., 2021) | N | PIS | NES | LC | † | † | † | † | † | † | † | S26 | |||
| Total | 26 | 13 | 10 | 7 | 3 | 8 | 6 | 4 | 4 | 2 | |||||
| Lakes | Seasons | Species Richness |
|---|---|---|
| Bi | Dry | 10 |
| Wet | 7 | |
| Da | Dry | 7 |
| Wet | 6 | |
| As | Dry | 13 |
| Wet | 11 | |
| Ge | Dry | 3 |
| Wet | 3 | |
| Ma | Dry | 8 |
| Wet | 8 | |
| Mb | Dry | 3 |
| Wet | 6 | |
| Pi | Dry | 4 |
| Wet | 4 | |
| Ng | Dry | 4 |
| Wet | 3 | |
| Ti | Dry | 2 |
| Wet | 1 |
| Parameters | Lakes | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| As | Bi | Da | Ge | Mas | Mb | Ng | Pi | Ti | ||
| Trophic groups | HER | 23.1 | 20 | 14.3 | 66.7 | 37.5 | 66.7 | 25 | 50 | 50 |
| INV | 30.8 | 20 | 14.3 | |||||||
| OMV | 38.5 | 40 | 42.8 | 50 | 33.3 | 25 | 50 | |||
| PIS | 7.7 | 20 | 28.6 | 33.3 | 12.5 | 50 | 50 | |||
| Breeding groups | MOU | 7.7 | 10 | 14.2 | 33.33 | 12.5 | 16.7 | 25 | 25 | 50 |
| NES | 30.8 | 30 | 42.9 | 33.33 | 25 | 50 | 25 | 50 | ||
| SUB | 61.5 | 60 | 42.9 | 33.33 | 62.5 | 83.3 | 25 | 50 | ||
| Ecological Indices | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| H’ | J | D | |||||||
| Sites | General Values | Seasonal Values | General Values | Seasonal Values | General Values | Seasonal Values | |||
| Wet | Dry | Wet | Dry | Wet | Dry | ||||
| As | 2.50 | 2.48 | 2.41 | 0.97 | 0.96 | 0.97 | 1.62 | 1.74 | 1.71 |
| Ma | 2.04 | 2.04 | 2.03 | 0.98 | 0.98 | 0.97 | 1.09 | 1.17 | 1.29 |
| Bi | 1.94 | 1.99 | 1.84 | 0.84 | 0.86 | 0.94 | 1.21 | 1.99 | 0.89 |
| Da | 1.57 | 1.63 | 1.51 | 0.81 | 0.84 | 0.84 | 0.81 | 0.89 | 0.75 |
| Pi | 1.24 | 1.26 | 1.25 | 0.90 | 0.91 | 0.90 | 0.44 | 0.48 | 0.48 |
| Mb | 1.22 | 0.99 | 1.34 | 0.68 | 0.90 | 0.75 | 0.72 | 0.32 | 0.78 |
| Ge | 0.99 | 1.01 | 1.00 | 0.91 | 0.92 | 0.91 | 0.35 | 0.39 | 0.41 |
| Ng | 0.81 | 0.93 | 0.66 | 0.58 | 0.67 | 0.60 | 0.49 | 0.55 | 0.37 |
| Ti | 0.57 | 0.67 | 0.29 | 0.83 | 0.97 | 0.42 | 0.19 | 0.21 | 0.22 |
| Bi | 0.0277 (0.70) | |||||||
| Da | 0.0284 (0.62) | 0.0304 (0.73) | ||||||
| Ge | 0.0283 (0.53) | 0.0261 (0.55) | 0.0318 (0.50) | |||||
| Ma | 0.0257 (0.64) | 0.0289 (0.67) | 0.0291 (0.61) | 0.0285 (0.69) | ||||
| Mb | 0.0312 (0.75 | 0.0296 (0.77) | 0.0295 (0.75) | 0.0271 (0.81) | 0.0288 (0.80) | |||
| Ng | 0.0268 (0.71) | 0.0289 (0.66) | 0.0292 (0.69) | 0.0272 (0.72) | 0.0271 (0.61) | 0.0318 (0.80) | ||
| Pi | 0.0277 (0.61) | 0.0291 (0.69) | 0.0275 (0.67) | 0.0274 (0.42) | 0.0302 (0.53) | 0.028 (0.82) | 0.0269 (0.73) | |
| Ti | 0.0273 (0.70) | 0.0272 (0.73) | 0.0293 (0.72) | 0.0274 (0.79) | 0.0259 (0.74) | 0.0289 (0.83) | 0.0277 (0.30) | 0.0221 (0.78) |
| As | Bi | Da | Ge | Ma | Mb | Ng | Pi |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kitio, H.I.; Bitja Nyom, A.R.; Pariselle, A.; Bilong Bilong, C.F. Inventory and Ecological Characterization of Ichthyofauna of Nine Lakes in the Adamawa Region (Northern Cameroon, Central Africa). Diversity 2022, 14, 770. https://doi.org/10.3390/d14090770
Kitio HI, Bitja Nyom AR, Pariselle A, Bilong Bilong CF. Inventory and Ecological Characterization of Ichthyofauna of Nine Lakes in the Adamawa Region (Northern Cameroon, Central Africa). Diversity. 2022; 14(9):770. https://doi.org/10.3390/d14090770
Chicago/Turabian StyleKitio, Hermann I., Arnold R. Bitja Nyom, Antoine Pariselle, and Charles F. Bilong Bilong. 2022. "Inventory and Ecological Characterization of Ichthyofauna of Nine Lakes in the Adamawa Region (Northern Cameroon, Central Africa)" Diversity 14, no. 9: 770. https://doi.org/10.3390/d14090770
APA StyleKitio, H. I., Bitja Nyom, A. R., Pariselle, A., & Bilong Bilong, C. F. (2022). Inventory and Ecological Characterization of Ichthyofauna of Nine Lakes in the Adamawa Region (Northern Cameroon, Central Africa). Diversity, 14(9), 770. https://doi.org/10.3390/d14090770