


Altitudinal Patterns of Native and Invasive Alien Herbs along Roadsides in the Dayao Mountain National Nature Reserve, Guangxi, China


Abstract
:1. Introduction
2. Materials and Methods
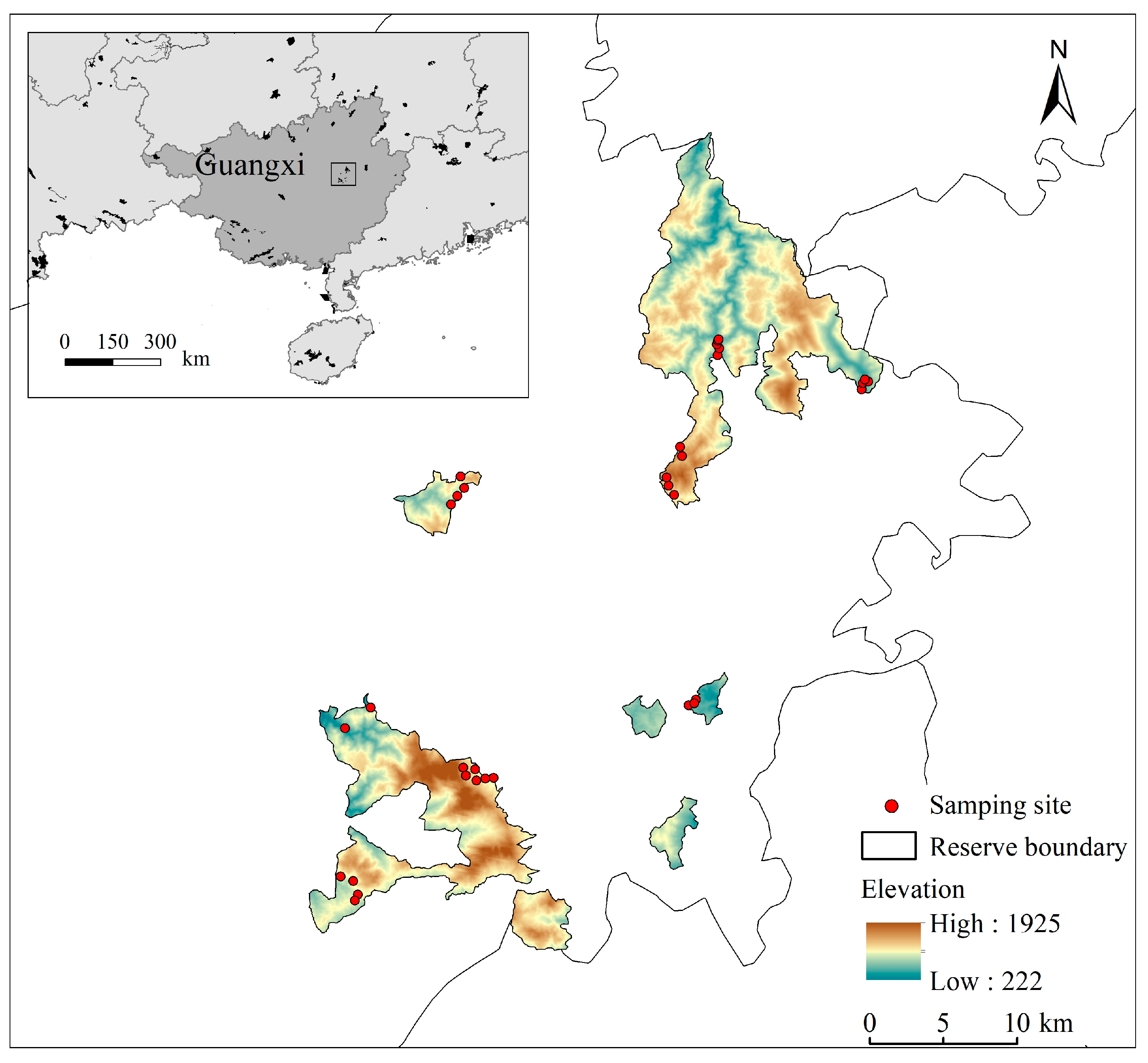
2.1. Study Area
2.2. Data Collection
2.3. Climatic and Human Disturbance Factors
2.4. Data Analysis
3. Results
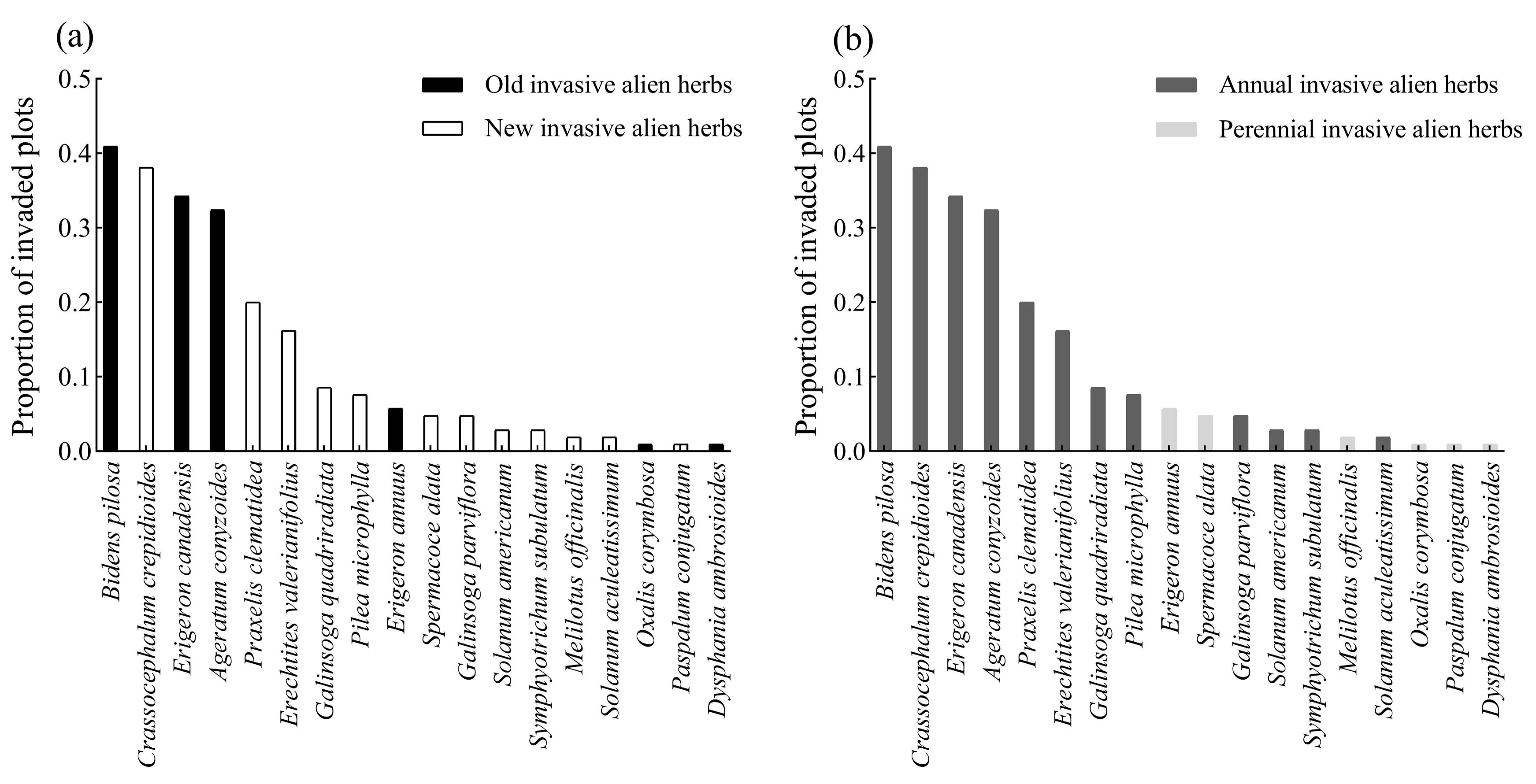
3.1. Composition and Distribution of Native and Invasive Alien Herbs
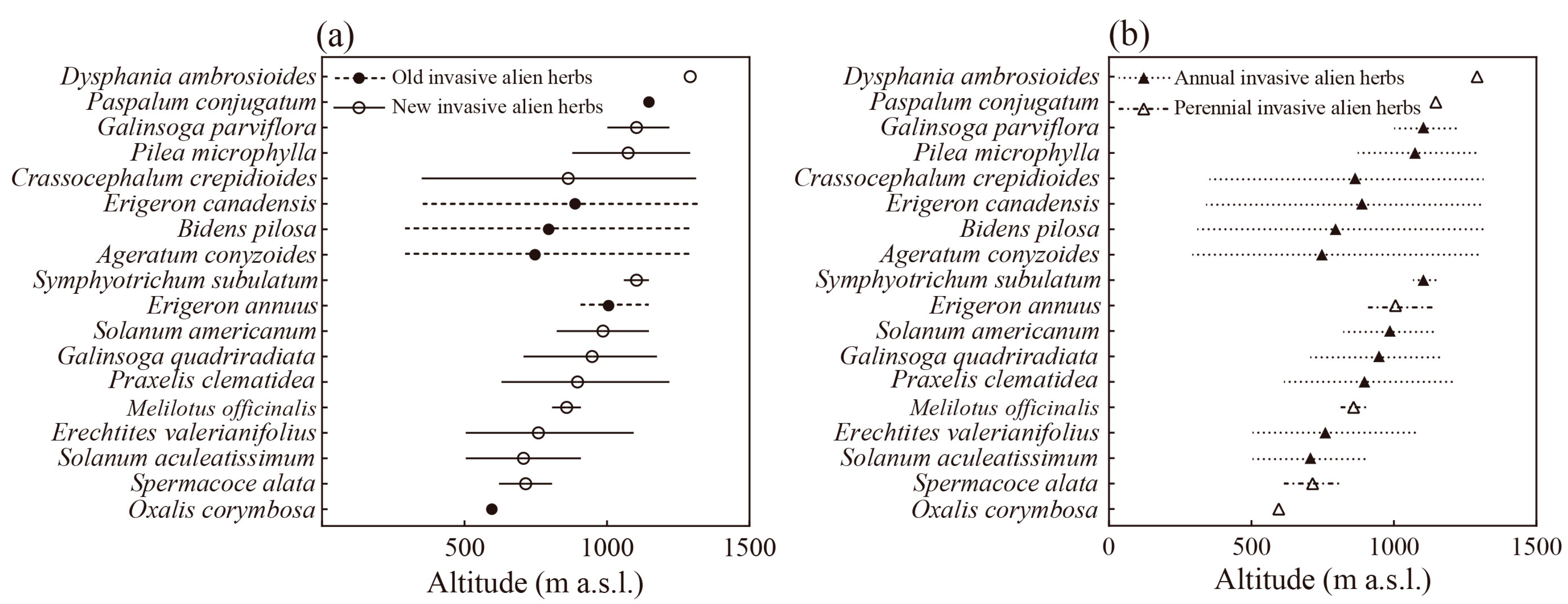
3.2. Altitude Distribution Patterns of Native and Invasive Alien Herbs
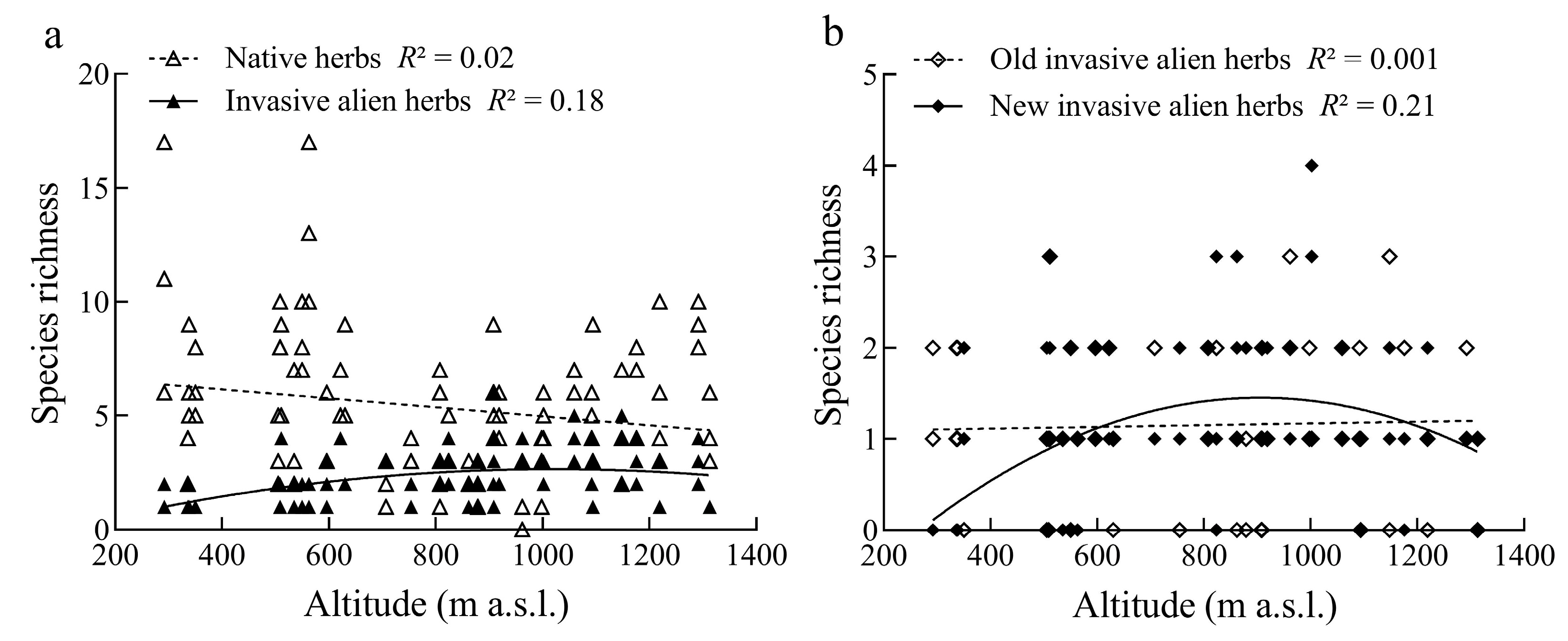
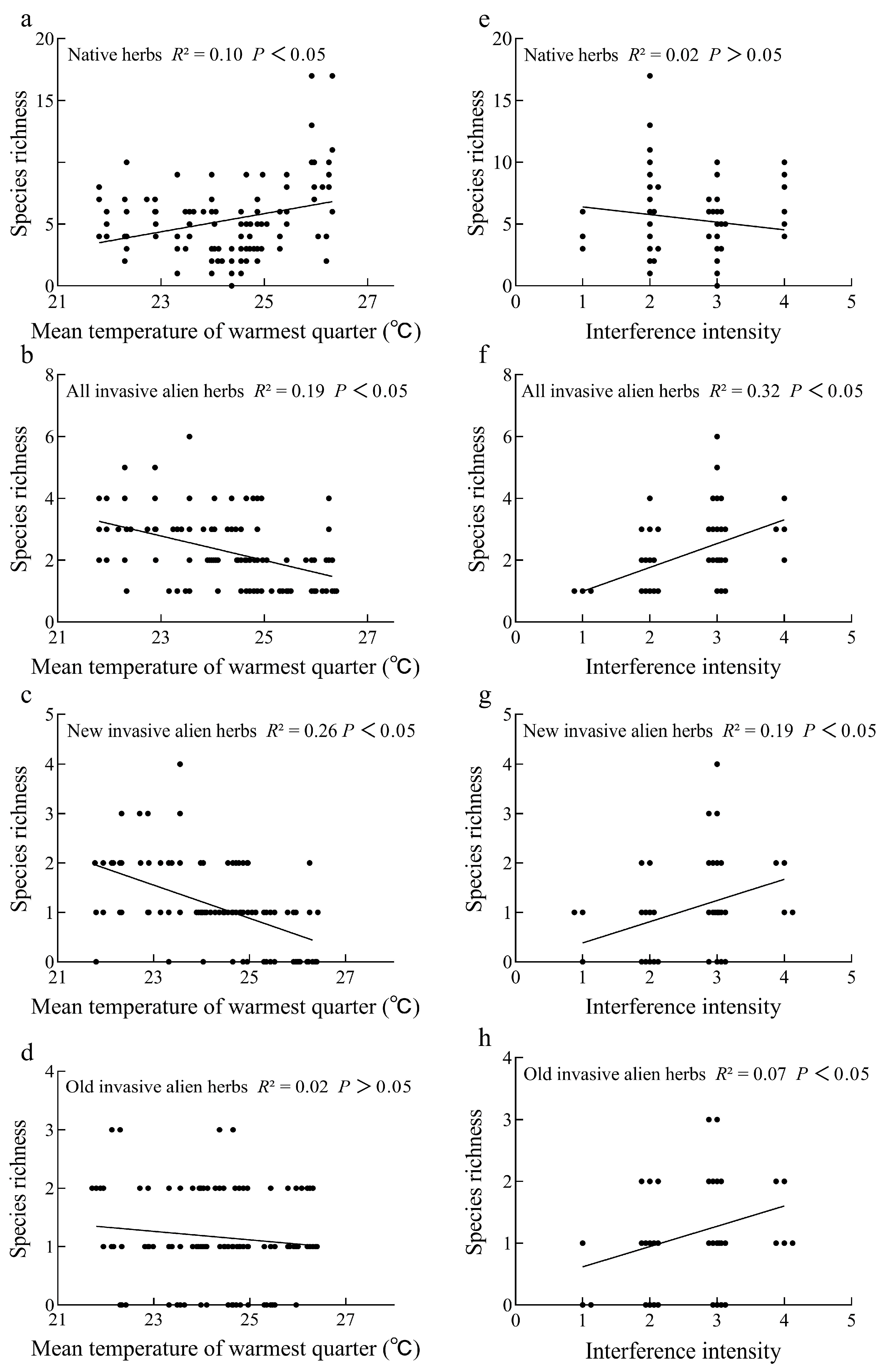
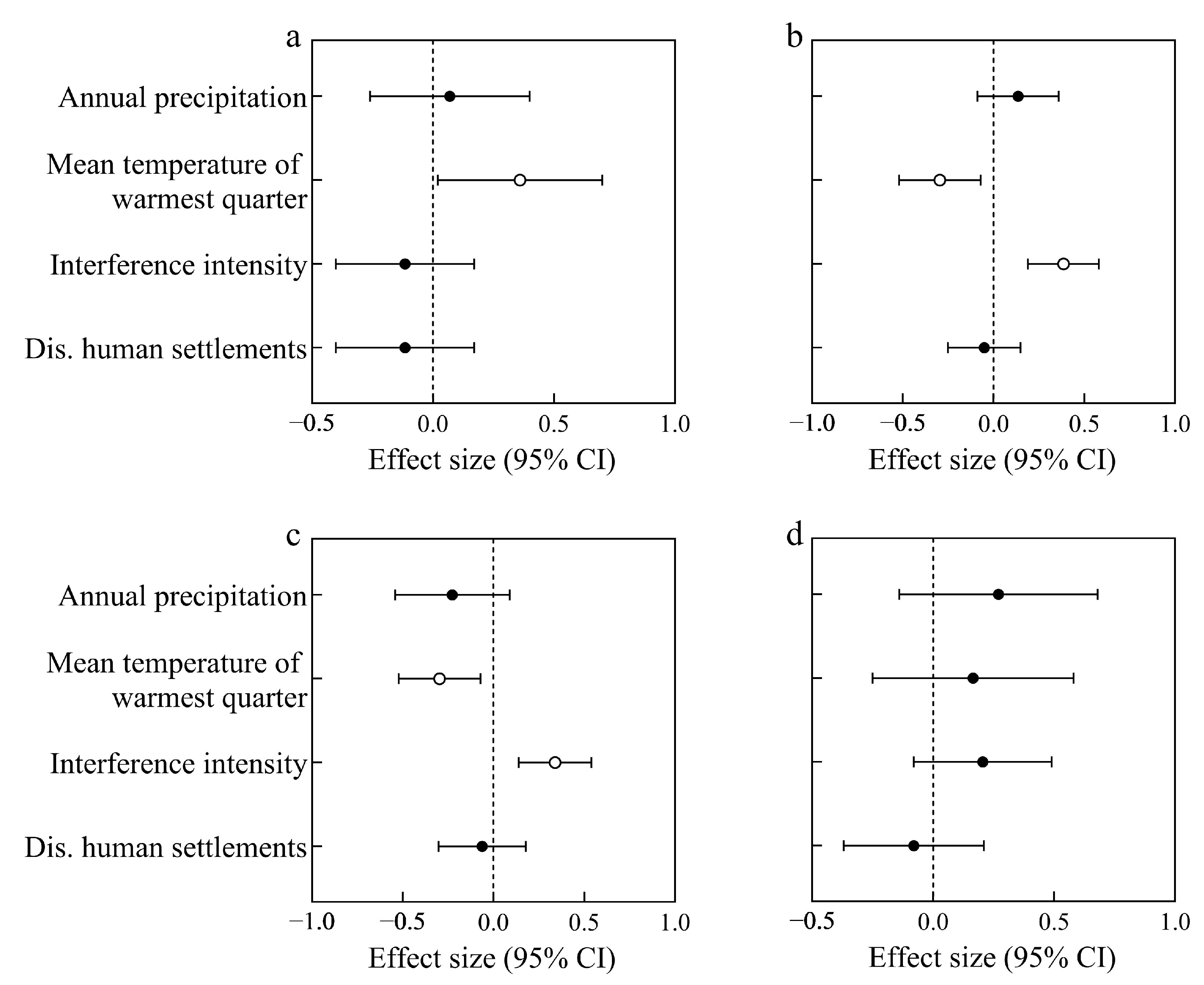
3.3. Factors Determining the Distribution Patterns of Native and Invasive Herbs
4. Discussion
4.1. Altitudinal Patterns of Native and Invasive Alien Herbs
4.2. Effects of Environmental Factors on the Richness of Native and Invasive Herbs
4.3. Effects of Environmental Factors on the Richness of Invasive Alien Herbs at Different Introduction Times
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Perrings, C.; Naeem, S.; Ahrestani, F.; Bunker, D.E.; Burkill, P.; Canziani, G.; Elmqvist, T.; Ferrati, R.; Fuhrman, J.A.; Jaksic, F.; et al. Ecosystem Services for 2020. Science 2010, 330, 323–324. [Google Scholar] [CrossRef] [PubMed]
- Hulme, P.E. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Kalwij, J.M.; Robertson, M.P.; van Rensburg, B.J. Annual monitoring reveals rapid upward movement of exotic plants in a montane ecosystem. Biol. Invasions 2015, 17, 3517–3529. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Shen, Z. Roles of Dispersal Limit and Environmental Filtering in Shaping the Spatiotemporal Patterns of Invasive Alien Plant Diversity in China. Front. Ecol. Evol. 2020, 8, 544670. [Google Scholar] [CrossRef]
- Tittensor, D.P.; Walpole, M.; Hill, S.L.L.; Boyce, D.G.; Britten, G.L.; Burgess, N.D.; Butchart, S.H.M.; Leadley, P.W.; Regan, E.C.; Alkemade, R.; et al. A mid-term analysis of progress toward international biodiversity targets. Science 2014, 346, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Braun, M.; Schindler, S.; Essl, F. Distribution and management of invasive alien plant species in protected areas in Central Europe. J. Nat. Conserv. 2016, 33, 48–57. [Google Scholar] [CrossRef]
- Bhatta, S.; Joshi, L.R.; Shrestha, B.B. Distribution and impact of invasive alien plant species in Bardia National Park, western Nepal. Environ. Conserv. 2020, 47, 197–205. [Google Scholar] [CrossRef]
- Byers, J.E. Impact of non-indigenous species on natives enhanced by anthropogenic alteration of selection regimes. Oikos 2002, 97, 449–458. [Google Scholar] [CrossRef]
- Foxcroft, L.C.; Jarosik, V.; Pysek, P.; Richardson, D.M.; Rouget, M. Protected-Area Boundaries as Filters of Plant Invasions. Conserv. Biol. 2011, 25, 400–405. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Moreno, P.; Diez, J.M.; Ibanez, I.; Font, X.; Vila, M. Plant invasions are context-dependent: Multiscale effects of climate, human activity and habitat. Divers. Distrib. 2014, 20, 720–731. [Google Scholar] [CrossRef]
- Thalmann, D.J.K.; Kikodze, D.; Khutsishvili, M.; Kharazishvili, D.; Guisan, A.; Broennimann, O.; Mueller-Schaerer, H. Areas of high conservation value in Georgia: Present and future threats by invasive alien plants. Biol. Invasions 2015, 17, 1041–1054. [Google Scholar] [CrossRef] [Green Version]
- Shackleton, R.T.; Foxcroft, L.C.; Pysek, P.; Wood, L.E.; Richardson, D.M. Assessing biological invasions in protected areas after 30 years: Revisiting nature reserves targeted by the 1980s SCOPE programme. Biol. Conserv. 2020, 243, 108424. [Google Scholar] [CrossRef]
- Tyser, R.W.; Worley, C.A. Alien flora in grasslands adjacent to road and trail corridors in Glacier National-park, Montana (USA). Conserv. Biol. 1992, 6, 253–262. [Google Scholar] [CrossRef]
- McDougall, K.L.; Lembrechts, J.; Rew, L.J.; Haider, S.; Cavieres, L.A.; Kueffer, C.; Milbau, A.; Naylor, B.J.; Nunez, M.A.; Pauchard, A.; et al. Running off the road: Roadside non-native plants invading mountain vegetation. Biol. Invasions 2018, 20, 3461–3473. [Google Scholar] [CrossRef] [Green Version]
- Liedtke, R.; Barros, A.; Essl, F.; Lembrechts, J.J.; Wedegartner, R.E.M.; Pauchard, A.; Dullinger, S. Hiking trails as conduits for the spread of non-native species in mountain areas. Biol. Invasions 2020, 22, 1121–1134. [Google Scholar] [CrossRef] [Green Version]
- Gong, L.; Li, J.; Liu, X.; Zhao, X.; Zhao, C. Analysis of invasive alien species in Chinese national nature reserves. Ecol. Sci. 2017, 36, 210–216. [Google Scholar] [CrossRef]
- Pollnac, F.; Seipel, T.; Repath, C.; Rew, L.J. Plant invasion at landscape and local scales along roadways in the mountainous region of the Greater Yellowstone Ecosystem. Biol. Invasions 2012, 14, 1753–1763. [Google Scholar] [CrossRef]
- Lembrechts, J.J.; Alexander, J.M.; Cavieres, L.A.; Haider, S.; Lenoir, J.; Kueffer, C.; McDougall, K.; Naylor, B.J.; Nunez, M.A.; Pauchard, A.; et al. Mountain roads shift native and non-native plant species’ ranges. Ecography 2017, 40, 353–364. [Google Scholar] [CrossRef]
- Guo, Q.; Fei, S.; Shen, Z.; Iannone, B.V., III; Knott, J.; Chown, S.L. A global analysis of elevational distribution of non-native versus native plants. J. Biogeogr. 2018, 45, 793–803. [Google Scholar] [CrossRef]
- Pauchard, A.; Kueffer, C.; Dietz, H.; Daehler, C.C.; Alexander, J.; Edwards, P.J.; Arevalo, J.R.; Cavieres, L.A.; Guisan, A.; Haider, S.; et al. Ain’t no mountain high enough: Plant invasions reaching new elevations. Front. Ecol. Environ. 2009, 7, 479–486. [Google Scholar] [CrossRef]
- Barros, A.; Pickering, C.M. Non-native Plant Invasion in Relation to Tourism Use of Aconcagua Park, Argentina, the Highest Protected Area in the Southern Hemisphere. Mt. Res. Dev. 2014, 34, 13–26. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Lu, Z.; Fan, Z.; Liu, X.; Hens, L.; De Wulf, R.; Ou, X. Distribution of non-native plant species along elevation gradients in a protected area in the eastern Himalayas, China. Alp. Bot. 2018, 128, 169–178. [Google Scholar] [CrossRef]
- Ibanez, T.; Hart, P.; Ainsworth, A.; Gross, J.; Monello, R. Factors associated with alien plant richness, cover and composition differ in tropical island forests. Divers. Distrib. 2019, 25, 1910–1923. [Google Scholar] [CrossRef] [Green Version]
- Landi, S.; Tordoni, E.; Amici, V.; Bacaro, G.; Carboni, M.; Filibeck, G.; Scoppola, A.; Bagella, S. Contrasting patterns of native and non-native plants in a network of protected areas across spatial scales. Biodivers. Conserv. 2020, 29, 2035–2053. [Google Scholar] [CrossRef]
- Rojas-Sandoval, J.; Ackerman, J.D.; Tremblay, R.L. Island biogeography of native and alien plant species: Contrasting drivers of diversity across the Lesser Antilles. Divers. Distrib. 2020, 26, 1539–1550. [Google Scholar] [CrossRef]
- Pretto, F.; Celesti-Grapow, L.; Carli, E.; Brundu, G.; Blasi, C. Determinants of non-native plant species richness and composition across small Mediterranean islands. Biol. Invasions 2012, 14, 2559–2572. [Google Scholar] [CrossRef]
- Paudel, S.; Benavides, J.C.; MacDonald, B.; Longcore, T.; Wilson, G.W.T.; Loss, S.R. Determinants of native and non-native plant community structure on an oceanic island. Ecosphere 2017, 8, e01927. [Google Scholar] [CrossRef] [Green Version]
- Haider, S.; Kueffer, C.; Bruelheide, H.; Seipel, T.; Alexander, J.M.; Rew, L.J.; Ramon Arevalo, J.; Cavieres, L.A.; McDougall, K.L.; Milbau, A.; et al. Mountain roads and non-native species modify elevational patterns of plant diversity. Glob. Ecol. Biogeogr. 2018, 27, 667–678. [Google Scholar] [CrossRef] [Green Version]
- Marini, L.; Battisti, A.; Bona, E.; Federici, G.; Martini, F.; Pautasso, M.; Hulme, P.E. Alien and native plant life-forms respond differently to human and climate pressures. Glob. Ecol. Biogeogr. 2012, 21, 534–544. [Google Scholar] [CrossRef]
- Sullivan, J.J.; Williams, P.A.; Timmins, S.M.; Smale, M.C. Distribution and spread of environmental weeds along New Zealand roadsides. N. Z. J. Ecol. 2009, 33, 190–204. [Google Scholar]
- Pauchard, A.; Milbau, A.; Albihn, A.; Alexander, J.; Burgess, T.; Daehler, C.; Englund, G.; Essl, F.; Evengard, B.; Greenwood, G.B.; et al. Non-native and native organisms moving into high elevation and high latitude ecosystems in an era of climate change: New challenges for ecology and conservation. Biol. Invasions 2016, 18, 345–353. [Google Scholar] [CrossRef]
- Dainese, M.; Aikio, S.; Hulme, P.E.; Bertolli, A.; Prosser, F.; Marini, L. Human disturbance and upward expansion of plants in a warming climate. Nat. Clim. Change 2017, 7, 577–580. [Google Scholar] [CrossRef]
- Carboni, M.; Gueguen, M.; Barros, C.; Georges, D.; Boulangeat, I.; Douzet, R.; Dullinger, S.; Klonner, G.; van Kleunen, M.; Essl, F.; et al. Simulating plant invasion dynamics in mountain ecosystems under global change scenarios. Glob. Change Biol. 2018, 24, E289–E302. [Google Scholar] [CrossRef] [Green Version]
- Steyn, C.; Greve, M.; Robertson, M.P.; Kalwij, J.M.; le Roux, P.C. Alien plant species that invade high elevations are generalists: Support for the directional ecological filtering hypothesis. J. Veg. Sci. 2017, 28, 337–346. [Google Scholar] [CrossRef] [Green Version]
- Felde, V.A.; Kapfer, J.; Grytnes, J.-A. Upward shift in elevational plant species ranges in Sikkilsdalen, central Norway. Ecography 2012, 35, 922–932. [Google Scholar] [CrossRef]
- Pysek, P.; Manceur, A.M.; Alba, C.; McGregor, K.F.; Pergl, J.; Stajerova, K.; Chytry, M.; Danihelka, J.; Kartesz, J.; Klimesova, J.; et al. Naturalization of central European plants in North America: Species traits, habitats, propagule pressure, residence time. Ecology 2015, 96, 762–774. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pysek, P. Plant invasions: Merging the concepts of species invasiveness and community invasibility. Prog. Phys. Geogr. -Earth Environ. 2006, 30, 409–431. [Google Scholar] [CrossRef] [Green Version]
- Richardson, D.M.; Pysek, P. Naturalization of introduced plants: Ecological drivers of biogeographical patterns. New Phytol. 2012, 196, 383–396. [Google Scholar] [CrossRef]
- Polce, C.; Kunin, W.E.; Biesmeijer, J.C.; Dauber, J.; Phillips, O.L.; Network, A.F.S. Alien and native plants show contrasting responses to climate and land use in Europe. Glob. Ecol. Biogeogr. 2011, 20, 367–379. [Google Scholar] [CrossRef]
- Eichenberg, D.; Bowler, D.E.; Bonn, A.; Bruelheide, H.; Grescho, V.; Harter, D.; Jandt, U.; May, R.; Winter, M.; Jansen, F. Widespread decline in Central European plant diversity across six decades. Glob. Change Biol. 2021, 27, 1097–1110. [Google Scholar] [CrossRef]
- Chen, B.; Wang, L. Investigation and application of wild ornamental ferns in Dayao Mountain National Nature Reserve, Guangxi. Contemp. Hortic. 2021, 44, 83–86. [Google Scholar] [CrossRef]
- Chen, B.; Wang, L.; Sun, C. Investigation of wild woody ornamental plant resources in Dayao Mountain National Nature Reserve. Chin. Wild Plant Resour. 2021, 40, 61–67. [Google Scholar] [CrossRef]
- Wang, L.; Luo, B.; Tan, M.; Chen, B. Study on Flora of Dayao Mountain Nature Reserve in Guangxi. In Proceedings of the Second China Forestry Academic Conference—S8 Wildlife, Wetland and Nature Reserve, Nanning, China, 7 November 2009; pp. 132–136. [Google Scholar]
- Ma, J. Flora of Invasive Aliens in China, 1st ed.; Shanghai Jiao Tong University Press: Shanghai, China, 2021; pp. 39–378. [Google Scholar]
- Xu, Z.; Feng, Z.; Yang, J.; Zheng, J.; Zhang, F. Nowhere to Invade: Rumex crispus and Typha latifolia Projected to Disappear under Future Climate Scenarios. PLoS ONE 2013, 8, e70728. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wei, H.; Zhao, Z.; Liu, J.; Zhang, Q.; Zhang, X.; Gu, W. The Global Potential Distribution of Invasive Plants: Anredera cordifolia under Climate Change and Human Activity Based on Random Forest Models. Sustainability 2020, 12, 1491. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Yin, D.; Huang, D.; Du, N.; Liu, J.; Guo, W.; Wang, R. Altitudinal patterns illustrate the invasion mechanisms of alien plants in temperate mountain forests of northern China. For. Ecol. Manag. 2015, 351, 1–8. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 12 January 2022).
- Koerner, C. The use of ‘altitude’ in ecological research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef]
- Haider, S.; Alexander, J.; Dietz, H.; Trepl, L.; Edwards, P.J.; Kueffer, C. The role of bioclimatic origin, residence time and habitat context in shaping non-native plant distributions along an altitudinal gradient. Biol. Invasions 2010, 12, 4003–4018. [Google Scholar] [CrossRef]
- Applestein, C.; Germino, M.J. Patterns of post-fire invasion of semiarid shrub-steppe reveals a diversity of invasion niches within an exotic annual grass community. Biol. Invasions 2022, 24, 741–759. [Google Scholar] [CrossRef]
- Roy, K.; Goldberg, E.E. Origination, extinction, and dispersal: Integrative models for understanding present-day diversity gradients. Am. Nat. 2007, 170, S71–S85. [Google Scholar] [CrossRef]
- Clark-Tapia, R.; Gonzalez-Adame, G.; Campos, J.E.; Aguirre-Hidalgo, V.; Pacheco-Cruz, N.; Ugalde, J.J.V.T.; Campista-Leon, S.; Peinado-Guevara, L.I.; Alfonso-Corrado, C. Effects of Habitat Loss on the Ecology of Pachyphytum caesium (Crassulaceae), a Specialized Cliff-Dwelling Endemic Species in Central Mexico. Diversity 2021, 13, 421. [Google Scholar] [CrossRef]
- Sheppard, C.S.; Brendel, M.R. Competitive ability of native and alien plants: Effects of residence time and invasion status. Neobiota 2021, 65, 47–69. [Google Scholar] [CrossRef]
- Becker, T.; Dietz, H.; Billeter, R.; Buschmann, H.; Edwards, P.J. Altitudinal distribution of alien plant species in the Swiss Alps. Perspect. Plant Ecol. Evol. Syst. 2005, 7, 173–183. [Google Scholar] [CrossRef]
- Ross, L.C.; Lambdon, P.W.; Hulme, P.E. Disentangling the roles of climate, propagule pressure and land use on the current and potential elevational distribution of the invasive weed Oxalis pes-caprae L. on Crete. Perspect. Plant Ecol. Evol. Syst. 2008, 10, 251–258. [Google Scholar] [CrossRef]
- Roy, S.; Simon, J.P.; Lapointe, F.J. Determination of the origin of the cold-adapted populations of barnyard grass (Echinochloa crus-galli) in eastern North America: A total-evidence approach using RAPD DNA and DNA sequences. Can. J. Bot. -Rev. Can. De Bot. 2000, 78, 1505–1513. [Google Scholar] [CrossRef]
- Essl, F.; Glaser, M.; Schertler, A. New and old invaders in forests in eastern Austria: The role of species attributes and invasion history. Flora 2021, 283, 151922. [Google Scholar] [CrossRef]
- Padalia, H.; Bushra, A.; Singh, G.; Nandy, S.; Rai, I.D.; Rawat, G.S. Spatial patterns of plant functional types and environmental proxies of plant richness in alpine region of Western Himalaya, India. Biodivers. Conserv. 2019, 28, 2221–2244. [Google Scholar] [CrossRef]
- Gao, D.; Fu, L.; Sun, J.; Li, Y.; Cao, Z.; Liu, Y.; Xu, P.; Zhao, J. The mid-domain effect and habitat complexity applied to elevational gradients: Moss species richness in a temperate semihumid monsoon climate mountain of China. Ecol. Evol. 2021, 11, 7448–7460. [Google Scholar] [CrossRef]
- Alexander, J.M.; Kueffer, C.; Daehler, C.C.; Edwards, P.J.; Pauchard, A.; Seipel, T.; Consortium, M. Assembly of nonnative floras along elevational gradients explained by directional ecological filtering. Proc. Natl. Acad. Sci. USA 2011, 108, 656–661. [Google Scholar] [CrossRef] [Green Version]
- Khuroo, A.A.; Weber, E.; Malik, A.H.; Reshi, Z.A.; Dar, G.H. Altitudinal distribution patterns of the native and alien woody flora in Kashmir Himalaya, India. Environ. Res. 2011, 111, 967–977. [Google Scholar] [CrossRef]
- Davis, M.A.; Grime, J.P.; Thompson, K. Fluctuating resources in plant communities: A general theory of invasibility. J. Ecol. 2000, 88, 528–534. [Google Scholar] [CrossRef] [Green Version]
- Rose, M.; Hermanutz, L. Are boreal ecosystems susceptible to alien plant invasion? Evidence from protected areas. Oecologia 2004, 139, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Paiaro, V.; Cabido, M.; Pucheta, E. Altitudinal distribution of native and alien plant species in roadside communities from central Argentina. Austral Ecol. 2011, 36, 176–184. [Google Scholar] [CrossRef]
- Woitke, M.; Dietz, H. Shifts in dominance of native and invasive plants in experimental patches of vegetation. Perspect. Plant Ecol. Evol. Syst. 2002, 5, 165–184. [Google Scholar] [CrossRef]
- Gasso, N.; Pysek, P.; Vila, M.; Williamson, M. Spreading to a limit: The time required for a neophyte to reach its maximum range. Divers. Distrib. 2010, 16, 310–311. [Google Scholar] [CrossRef]
- Huebner, C.D.; Tobin, P.C. Invasibility of mature and 15-year-old deciduous forests by exotic plants. Plant Ecol. 2006, 186, 57–68. [Google Scholar] [CrossRef]
- Tanaka, T.; Sato, T. Contemporary patterns and temporal changes in alien plant species richness along an elevational gradient in central Japan. Plant Ecol. Evol. 2016, 149, 177–188. [Google Scholar] [CrossRef]
- Irl, S.D.H.; Schweiger, A.H.; Steinbauer, M.J.; Ah-Peng, C.; Arevalo, J.R.; Beierkuhnlein, C.; Chiarucci, A.; Daehler, C.C.; Fernandez-Palacios, J.M.; Flores, O.; et al. Human impact, climate and dispersal strategies determine plant invasion on islands. J. Biogeogr. 2021, 48, 1889–1903. [Google Scholar] [CrossRef]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pysek, P.; Winter, M.; Arianoutsou, M.; et al. No saturation in the accumulation of alien species worldwide. Nat. Commun. 2017, 8, 14435. [Google Scholar] [CrossRef]
- Lososova, Z.; Chytry, M.; Tichy, L.; Danihelka, J.; Fajmon, K.; Hajek, O.; Kintrova, K.; Kuehn, I.; Lanikova, D.; Otypkova, Z.; et al. Native and alien floras in urban habitats: A comparison across 32 cities of central Europe. Glob. Ecol. Biogeogr. 2012, 21, 545–555. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Category | Environment Variables | t | p |
|---|---|---|---|
| Native herbs | Mean temperature of warmest quarter | 3.008 | 0.003 |
| Annual precipitation | 0.415 | 0.679 | |
| Interference intensity | −1.038 | 0.302 | |
| Dis. human settlement | −1.060 | 0.292 | |
| All invasive alien herbs | Mean temperature of warmest quarter | −2.469 | 0.015 |
| Annual precipitation | 0.191 | 0.848 | |
| Interference intensity | 3.913 | <0.001 | |
| Dis. human settlement | −0.371 | 0.712 | |
| New invasive alien herbs | Mean temperature of warmest quarter | −4.723 | <0.001 |
| Annual precipitation | −1.733 | 0.086 | |
| Interference intensity | 3.233 | 0.002 | |
| Dis. human settlement | 0.429 | 0.668 | |
| Old invasive alien herbs | Mean temperature of warmest quarter | 1.194 | 0.235 |
| Annual precipitation | 1.767 | 0.08 | |
| Interference intensity | 1.875 | 0.05 | |
| Dis. human settlement | −0.830 | 0.408 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, B.; Ni, X.; Zhao, C. Altitudinal Patterns of Native and Invasive Alien Herbs along Roadsides in the Dayao Mountain National Nature Reserve, Guangxi, China. Diversity 2023, 15, 105. https://doi.org/10.3390/d15010105
Li B, Ni X, Zhao C. Altitudinal Patterns of Native and Invasive Alien Herbs along Roadsides in the Dayao Mountain National Nature Reserve, Guangxi, China. Diversity. 2023; 15(1):105. https://doi.org/10.3390/d15010105
Chicago/Turabian StyleLi, Bai, Xinying Ni, and Caiyun Zhao. 2023. "Altitudinal Patterns of Native and Invasive Alien Herbs along Roadsides in the Dayao Mountain National Nature Reserve, Guangxi, China" Diversity 15, no. 1: 105. https://doi.org/10.3390/d15010105
APA StyleLi, B., Ni, X., & Zhao, C. (2023). Altitudinal Patterns of Native and Invasive Alien Herbs along Roadsides in the Dayao Mountain National Nature Reserve, Guangxi, China. Diversity, 15(1), 105. https://doi.org/10.3390/d15010105