



Museum-Based Research on the Lice (Insecta: Phthiraptera) Infestations of Hummingbirds (Aves: Trochilidae)—Prevalence, Genus Richness and Parasite Associations


Abstract
:1. Introduction
2. Materials and Methods
3. Results
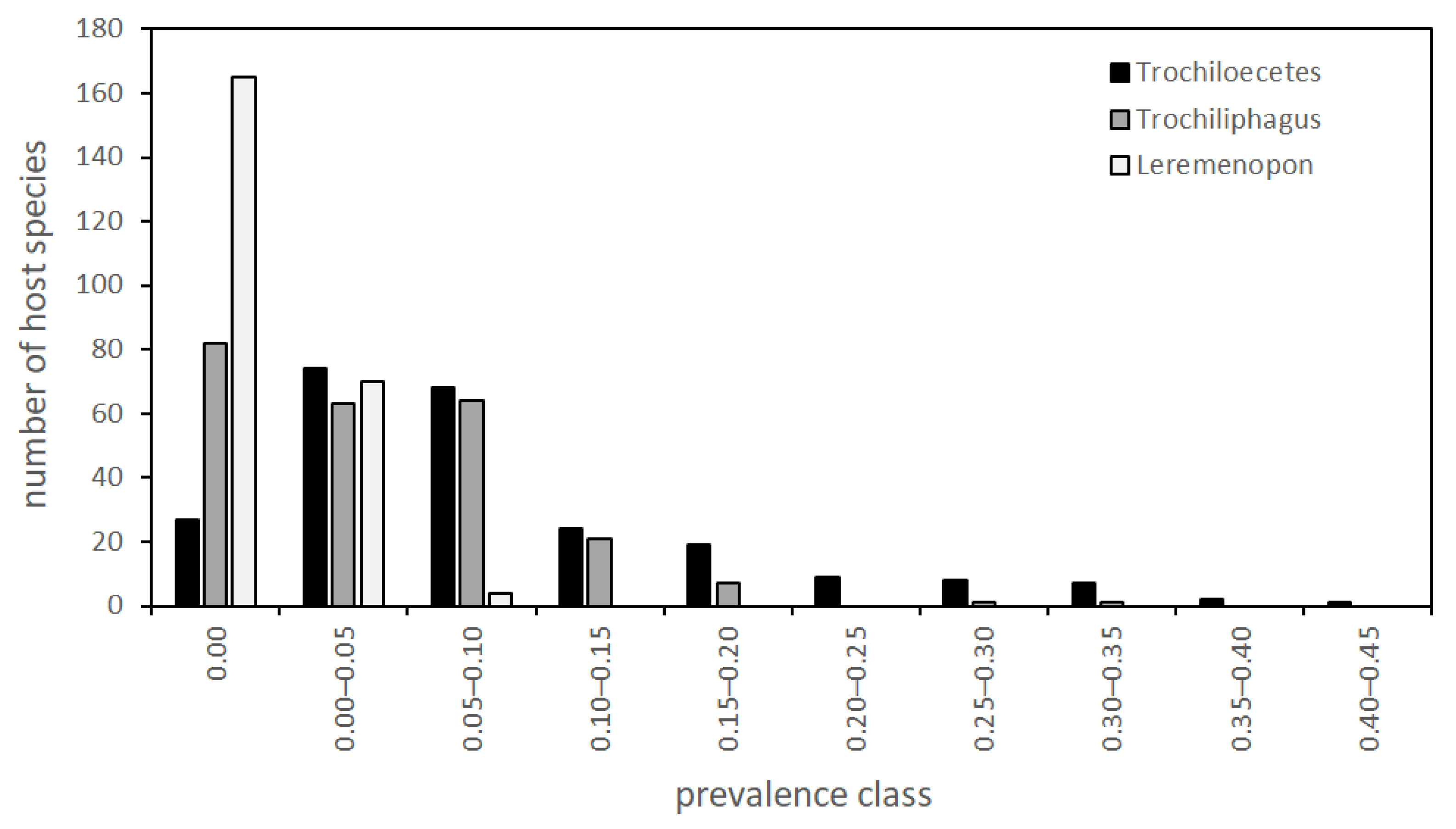
3.1. Prevalence of Infestations
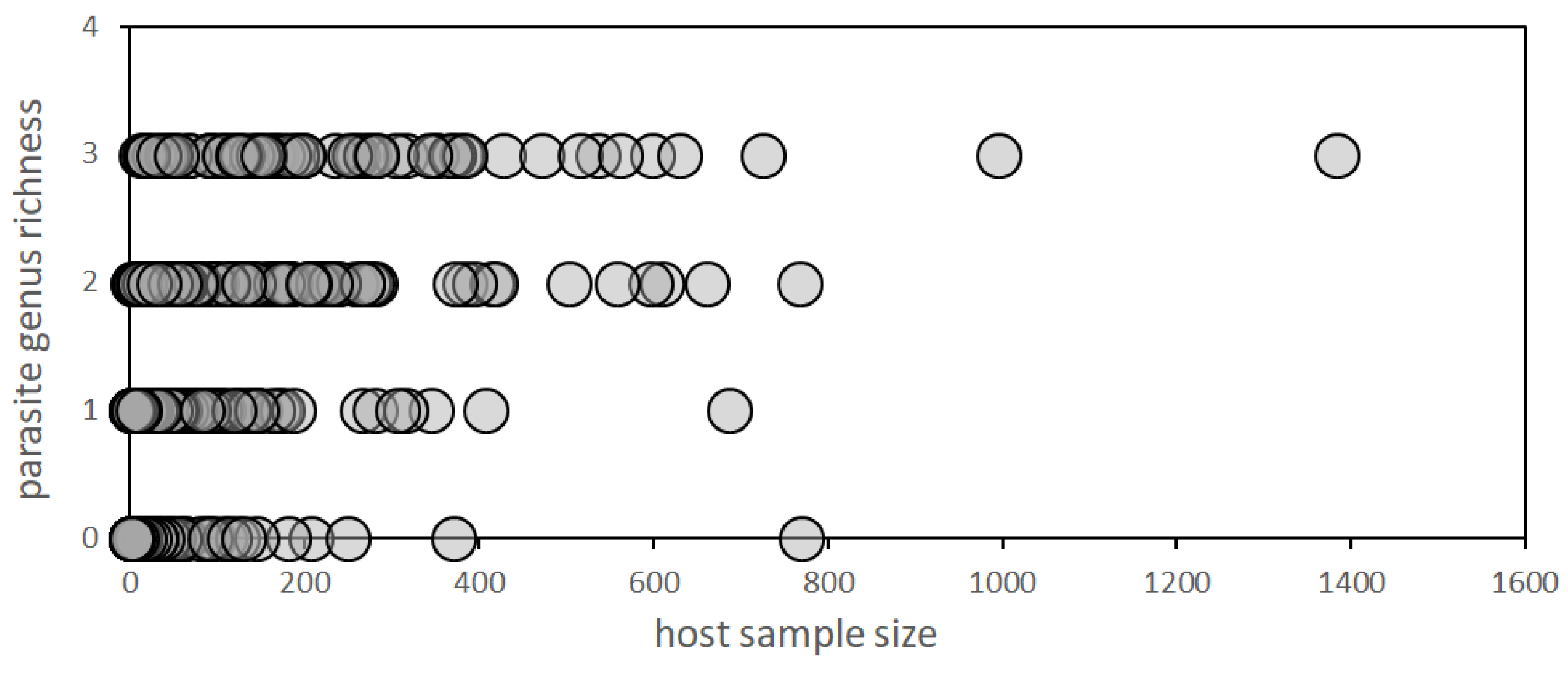
3.2. Host Sample Size and Parasite Genus Richness
3.3. Host Body Mass and Parasite Genus Richness
3.4. Host Body Mass and the Prevalence of Infestations
3.5. Relationships between the Prevalence of Different Parasites across Host Taxa
3.6. Parasite Co-infestations within Particular Host Taxa
3.7. Host Sexual Size Dimorphism and the Prevalence of Parasites
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnson, K.P.; Clayton, D.H. The biology, ecology, and evolution of chewing lice. In The Chewing Lice: World Checklist and Biological Overview; Price, R.D., Hellenthal, R.A., Palma, R.L., Johnson, K.P., Clayton, D.H., Eds.; Illinois Natural History Survey Special Publications 24: Champaign-Urbana, IL, USA, 2003; pp. 451–475. [Google Scholar]
- Clayton, D.H.; Tompkins, D.M. Comparative effects of mites and lice on the reproductive success of rock doves (Columba livia). Parasitology 1995, 110, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Clayton, D.H. Mate choice in experimentally parasitized rock doves: Lousy males lose. Am. Zool. 1990, 30, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Booth, D.T.; Clayton, D.H.; Block, B.A. Experimental demonstration of the energetic cost of parasitism in free-ranging hosts. Proc. R. Soc. Lond. B 1993, 253, 125–129. [Google Scholar] [CrossRef]
- Brown, C.R.; Brown, M.B.; Rannala, B. Ectoparasites reduce long-term survival of their avian host. Proc. R. Soc. Lond. B 1995, 262, 313–319. [Google Scholar] [CrossRef]
- Saxena, A.K.; Arya, G.; Bansal, N. Egg laying site and oviposition pattern of two phthirapteran species parasitizing red whiskered Bulbul (Pycnonotus jocosus). Turk. J. Parasitol. 2012, 36, 166–168. [Google Scholar] [CrossRef] [PubMed]
- Galloway, T.D.; Proctor, H.C.; Mironov, S.V. Chewing lice (Insecta: Phthiraptera: Amblycera, Ischnocera) and feather mites (Acari: Astigmatina: Analgoidea, Pterolichoidea): Ectosymbionts of grassland birds in Canada. In Arthropods of Canadian Grasslands Vol. 3: Biodiversity and Systematics Part 1; Cárcamo, H.A., Giberson, D.J., Eds.; Biological Survey of Canada: Ottawa, ON, Canada, 2014; pp. 139–188. [Google Scholar] [CrossRef]
- Møller, A.P.; Rózsa, L. Parasite biodiversity and host defenses: Chewing lice and immune response of their avian hosts. Oecologia 2005, 142, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Gill, F.; Donsker, D.; Rasmussen, P. (Eds.) IOC World Bird List (v.12.1). 2022. Available online: https://www.worldbirdnames.org/ioc-lists/crossref/ (accessed on 29 November 2022).
- Oniki-Willis, Y.; Willis, E.O.; Machado, V.L.L. On feeding habits of hummingbirds from five Brazilian states. Atual. Ornitológicas 2020, 218, 9–15. [Google Scholar]
- Colwell, R.K. Rensch’s rule crosses the line: Convergent allometry of sexual size dimorphism in Hummingbirds and flower mites. Am. Nat. 2000, 156, 495–510. [Google Scholar] [CrossRef]
- Nuñez-Rosas, L.; Arizmendi, M.C.; Cueva del Castillo, R.; Serrano-Meneses, M.A. Mating system, male territoriality and agility as predictors of the evolution of sexual size dimorphism in hummingbirds (Aves: Trochilidae). Behaviour 2017, 154, 1297–1341. [Google Scholar] [CrossRef]
- Parra, J.L. Color evolution in the hummingbird genus Coeligena. Evolution 2010, 64, 324–335. [Google Scholar] [CrossRef]
- Rico-Guevara, A.; Araya-Salas, M. Bills as daggers? A test for sexually dimorphic weapons in a lekking hummingbird. Behav. Ecol. 2015, 26, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Carriker, M.A. Studies in neotropical Mallophaga, XVII. A new family (Trochiliphagidae) and a new genus of the lice of hummingbirds. Proc. United States Natl. Mus. 1960, 112, 307–342. [Google Scholar] [CrossRef] [Green Version]
- Price, R.D.; Hellenthal, R.A.; Palma, R.L. 2003. World checklist of chewing lice with host associations and keys to families and genera. In The Chewing Lice: World Checklist and Biological Overview; Price, R.D., Hellenthal, R.A., Palma, R.L., Johnson, K.P., Clayton, D.H., Eds.; Illinois Natural History Survey Special Publications 24: Champaign-Urbana, IL, USA, 2003; pp. 1–448. [Google Scholar]
- Dalgleish, R.C.; Price, R.D. Two new species of Myrsidea (Phthiraptera: Amblycera: Menoponidae) from hummingbirds (Apodiformes: Trochilidae). Occas. Pap. West. Found. Vertebr. Zool. 2003, 6, 1–9. [Google Scholar]
- Dalgleish, R.C.; Price, R.D. A new genus and four new species of chewing lice (Phthiraptera: Amblycera: Menoponidae) from hummingbirds (Apodiformes: Trochilidae). Occas. Pap. West. Found. Vertebr. Zool. 2003, 7, 1–9. [Google Scholar]
- Rheinwald, G. The position of Trochiliphagus Carriker within the Ricinidae (Insecta: Phthiraptera). Bonn. Zool. Beiträge 2007, 55, 37. [Google Scholar]
- Valan, M. Chewing Lice of Hummingbirds and Wild Living Birds. Ph.D. Thesis, University of Veterinary and Pharmaceutical Sciences, Brno, Czech Republic, 2018. [Google Scholar]
- Harnos, A.; Lang, Z.; Petrás, D.; Bush, S.E.; Szabó, K.; Rózsa, L. Size matters for lice on birds: Coevolutionary allometry of host and parasite body size. Evolution 2017, 71, 421–431. [Google Scholar] [CrossRef]
- Clay, T. Piercing mouth-parts in the biting lice (Mallophaga). Nature 1949, 164, 617. [Google Scholar] [CrossRef] [PubMed]
- McGuire, J.A.; Witt, C.C.; Remsen, J.V.; Corl, A.; Rabosky, D.L.; Altshuler, D.L.; Dudley, R. Molecular phylogenetics and the diversification of hummingbirds. Curr. Biol. 2014, 24, 910–916. [Google Scholar] [CrossRef] [Green Version]
- Valan, M.; Sychra, O.; Literák, I. Chewing lice (Phthiraptera: Amblycera) of hummingbirds: Present status and future research. In Proceedings of the Abstract book of 5th International Conference on Phthiraptera, Park City, UT, USA, 2–7 August 2014. [Google Scholar]
- Rothschild, M.; Clay, T. Fleas, Flukes and Cuckoos—A Study of Bird Parasites; Macmillan Press: New York, NY, USA, 1957; pp. 1–305. [Google Scholar]
- Rózsa, L. Patterns in the abundance of avian lice (Phthiraptera: Amblycera, Ischnocera). J. Avian Biol. 1997, 28, 249–254. [Google Scholar] [CrossRef] [Green Version]
- Nelson, B.C. A revision of the New World species of Ricinus (Mallophaga) occurring on Passeriformes (Aves). Univ. Calif. Publ. Entomol. 1972, 68, 1–175. [Google Scholar] [CrossRef]
- Hamilton, W.D.; Zuk, M. Heritable true fitness and bright birds: A role for parasites? Science 1982, 218, 384–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balenger, S.L.; Zuk, M. Testing the Hamilton–Zuk hypothesis: Past, present, and future. Integr. Comp. Biol. 2014, 54, 601–613. [Google Scholar] [CrossRef] [Green Version]
- Gupta, N.; Khan, V.; Kumar, S.; Saxena, S.; Rashmi, A.; Saxena, A.K. Eggshell morphology of selected Indian bird lice species (Phthiraptera: Amblycera, Ischnocera). Entomol. News 2008, 120, 327–336. [Google Scholar] [CrossRef]
- Ahmad, A.; Khan, V.; Badola, S.; Arya, G.; Bansal, N.; Saxena, A.K. Population characteristics and the nature of egg shells of two Phthirapteran species parasitizing Indian cattle egrets. J. Insect Sci. 2010, 10, 163. [Google Scholar] [CrossRef] [PubMed]
- Foster, M. The eggs of three species of Mallophaga and their significance in ecological studies. J. Parasitol. 1969, 55, 435–456. [Google Scholar] [CrossRef]
- Foster, M. Synchronized life cycles in the Orange-crowned Warbler and its Mallophagan parasites. Ecology 1969, 50, 315–323. [Google Scholar] [CrossRef]
- Wetmore, A. The Birds of the REPUBLIC of Panamá, Part 2, Columbidae (Pigeons) to Picidae (Woodpeckers); Smithsonian Miscellaneous Collections 150(2); Smithsonian Institution Press: Washington, DC, USA, 1968; pp. 1–605. [Google Scholar]
- Ffrench, R. A Guide to the Birds of Trinidad & Tobago, 2nd ed.; Comstock Publishing Associates: Ithaca, NY, USA, 1991; pp. 1–426. [Google Scholar]
- Ruschi, A. Aves do Brasil, vol. IV, Beija-Flores; Expressão e Cultura: Rio de Janeiro, Brazil, 1982; pp. 1–208. [Google Scholar]
- Ruschi, A. Aves do Brasil, vol. V, Beija-Flores; Expressão e Cultura: Rio de Janeiro, Brazil, 1982; pp. 209–454. [Google Scholar]
- Grantsau, R. Os Beija-Flores do Brasil; Expressão e Cultura: Rio de Janeiro, Brazil, 1988; pp. 1–234. [Google Scholar]
- Dunning, J.B. CRC Handbook of Avian Body Masses, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2007; pp. 1–574. [Google Scholar]
- Piacentini, V.Q. Taxonomia e Distribuição Geográfica dos Representantes do Gênero Phaethornis Swainson, 1827 (Aves: Trochilidae). Ph.D. Thesis, Universidade de São Paulo, São Paulo, Brazil, 2011. [Google Scholar]
- Wilman, H.; Belmaker, J.; Simpson, J.; de la Rosa, C.; Rivadeneira, M.M.; Jetz, W. 2014. EltonTraits 1.0: Species-level foraging attributes of the world’s birds and mammals. Ecology 2014, 95, 2027. [Google Scholar] [CrossRef] [Green Version]
- de la Peña, M. Aves Argentinas: Descripción, comportamiento, reproducción y distribución. Columbidae, Cuculidae, Tytonidae, Strigidae, Nyctibiidae, Caprimulgidae, Apodidae y Trochilidae. In Comunicaciones del Museo Provincial de Ciencias Naturales “Florentino Ameghino” (Nueva Serie); El Museo: Santa Fe, Argentina, 2019; pp. 1–344. [Google Scholar]
- Marques, J.F.; Cabral, H.N. Effects of sample size on fish parasite prevalence, mean abundance and mean intensity estimates. J. Appl. Ichthyol. 2007, 23, 158–162. [Google Scholar] [CrossRef]
- Walther, B.A.; Cotgreave, P.; Gregory, R.D.; Price, R.D.; Clayton, D.H. Sampling effort and parasite species richness. Parasitol. Today 1995, 11, 306–310. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| N Taxa | Sample Size | rho | p |
|---|---|---|---|
| 398 | 1–1384 | 0.0099 | 0.8436 |
| 79 | 1–11 | 0.1847 | 0.1032 |
| 81 | 12–38 | 0.0495 | 0.6609 |
| 79 | 39–79 | −0.1419 | 0.2124 |
| 80 | 80–173 | −0.1308 | 0.2476 |
| 79 | 174–1384 | −0.0618 | 0.5885 |
| Variables | N | rho | p |
|---|---|---|---|
| Host body mass and Trochiloecetes prevalence | 213 | −0.0335 | 0.6264 |
| Host body mass and Trochiliphagus prevalence | 213 | −0.1037 | 0.1316 |
| Host body mass and Leremenopon prevalence | 213 | 0.1317 | 0.0550 |
| Lice Genera | N | rho | p |
|---|---|---|---|
| Trochiloecetes and Trochiliphagus | 239 | 0.0224 | 0.7305 |
| Trochiloecetes and Leremenopon | 239 | −0.0522 | 0.4215 |
| Trochiliphagus and Leremenopon | 239 | 0.2660 | <0.0001 |
| Host Species | N | Tcet + Tpha + | Tcet -Tpha - | Tcet + Tpha - | Tcet - Tpha + | Fisher’s Exact p |
|---|---|---|---|---|---|---|
| Archilochus colubris | 725 | 7 | 615 | 85 | 18 | 0.0294 |
| Chlorostilbon mellisugus | 599 | 6 | 528 | 37 | 28 | 0.0281 |
| Selasphorus scintilla | 557 | 4 | 514 | 31 | 8 | 0.0045 |
| Chrysolampis mosquitus | 428 | 8 | 361 | 33 | 26 | 0.0098 |
| Selasphorus platycercus | 394 | 3 | 370 | 20 | 1 | 0.0007 |
| Heliangelus exortis | 351 | 2 | 317 | 32 | 0 | 0.0091 |
| Variables | N | rho | p |
|---|---|---|---|
| Host SSD and Trochiloecetes prevalence | 74 | −0.0983 | 0.4045 |
| Host SSD and Trochiliphagus prevalence | 74 | −0.0895 | 0.4480 |
| Host SSD and Leremenopon prevalence | 74 | −0.1384 | 0.2395 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oniki-Willis, Y.; Willis, E.O.; Lopes, L.E.; Rózsa, L. Museum-Based Research on the Lice (Insecta: Phthiraptera) Infestations of Hummingbirds (Aves: Trochilidae)—Prevalence, Genus Richness and Parasite Associations. Diversity 2023, 15, 54. https://doi.org/10.3390/d15010054
Oniki-Willis Y, Willis EO, Lopes LE, Rózsa L. Museum-Based Research on the Lice (Insecta: Phthiraptera) Infestations of Hummingbirds (Aves: Trochilidae)—Prevalence, Genus Richness and Parasite Associations. Diversity. 2023; 15(1):54. https://doi.org/10.3390/d15010054
Chicago/Turabian StyleOniki-Willis, Yoshika, Edwin O. Willis, Leonardo E. Lopes, and Lajos Rózsa. 2023. "Museum-Based Research on the Lice (Insecta: Phthiraptera) Infestations of Hummingbirds (Aves: Trochilidae)—Prevalence, Genus Richness and Parasite Associations" Diversity 15, no. 1: 54. https://doi.org/10.3390/d15010054
APA StyleOniki-Willis, Y., Willis, E. O., Lopes, L. E., & Rózsa, L. (2023). Museum-Based Research on the Lice (Insecta: Phthiraptera) Infestations of Hummingbirds (Aves: Trochilidae)—Prevalence, Genus Richness and Parasite Associations. Diversity, 15(1), 54. https://doi.org/10.3390/d15010054