

Climatic and Non-Climatic Drivers of Plant Diversity along an Altitudinal Gradient in the Taihang Mountains of Northern China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Survey
2.3. Environmental Data Source
2.4. α-Diversity and β-Diversity Indices
2.5. Data Analysis
3. Results
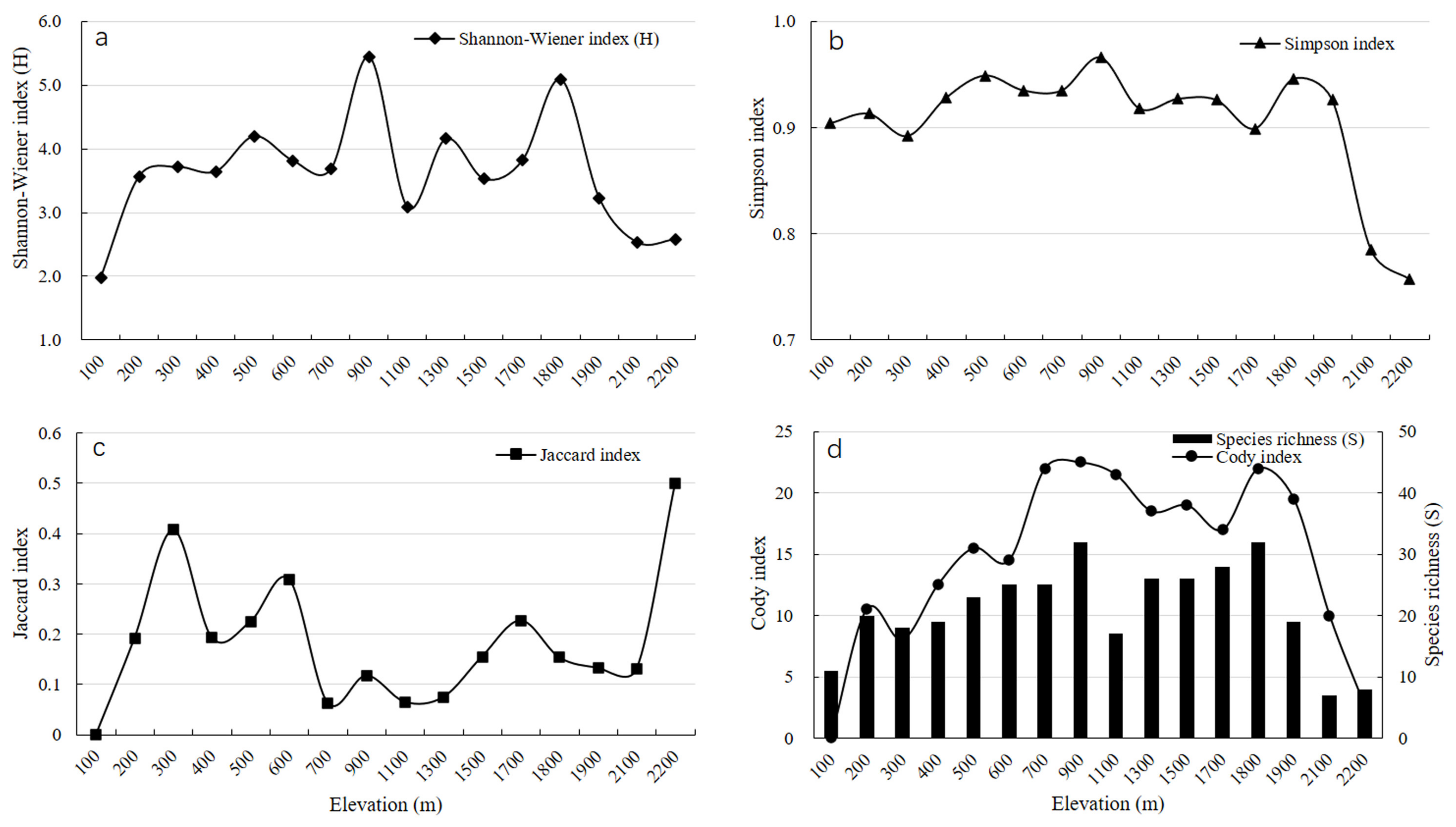
3.1. Altitudinal Distribution of A-Diversity in Plants
3.2. Β-Diversity of Plants along the Altitudinal Gradient
3.3. Relationships between Plant Diversity and Environmental Factors along the Altitudinal Gradient
4. Discussion
4.1. Plant Diversity Pattern along the Altitudinal Gradient
4.2. Characteristics of the Β-Diversity Pattern of the Altitudinal Gradient
4.3. Effects of Climatic and Non-Climatic Factors on the Plant Diversity Altitudinal Pattern
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dullinger, S.; Gattringer, A.; Thuiller, W.; Moser, D.; Zimmermann, N.E.; Guisan, A.; Willner, W.; Plutzar, C.; Leitner, M.; Mang, T.; et al. Extinction debt of high-mountain plants under twenty-first-century climate change. Nat. Clim. Change 2012, 2, 619–622. [Google Scholar] [CrossRef]
- Gottfried, M.; Pauli, H.; Futschik, A.; Akhalkatsi, M.; Barančok, P.; Benito Alonso, J.L.; Coldea, G.; Dick, J.; Erschbamer, B.; Fernández Calzado, M.R.; et al. Continent-wide response of mountain vegetation to climate change. Nat. Clim. Change 2012, 2, 111–115. [Google Scholar] [CrossRef]
- Beniston, M. Climatic change in mountain regions: A review of possible impacts. Clim. Change 2003, 59, 5–31. [Google Scholar] [CrossRef]
- Engler, R.; Randin, C.F.; Thuiller, W.; Dullinger, S.; Zimmermann, N.E.; Araújo, M.B.; Pearman, P.B.; Le Lay, G.; Piedallu, C.; Albert, C.H.; et al. 21st century climate change threatens mountain flora unequally across Europe. Glob. Change Biol. 2011, 17, 2330–2341. [Google Scholar] [CrossRef]
- McFadden, I.R.; Sandel, B.; Tsirogiannis, C.; Morueta-Holme, N.; Svenning, J.; Enquist, B.J.; Kraft, N.J. Temperature shapes opposing latitudinal gradients of plant taxonomic and phylogenetic β diversity. Ecol. Lett. 2019, 22, 1126–1135. [Google Scholar] [CrossRef] [PubMed]
- Gallardo-Cruz, J.A.; Perez-Garcia, E.A.; Meave, J.A. β-diversity and vegetation structure as influenced by slope aspect and altitude in a seasonally dry tropical landscape. Landsc. Ecol. 2009, 24, 473–482. [Google Scholar] [CrossRef]
- Loarie, S.R.; Duffy, P.B.; Hamilton, H.; Asner, G.P.; Field, C.B.; Ackerly, D.D. The velocity of climate change. Nature 2009, 462, 1052–1055. [Google Scholar] [CrossRef]
- Kattan, G.H.; Franco, P.; Rojas, V.; Morales, G. Biological diversification in a complex region: A spatial analysis of faunistic diversity and biogeography of the Andes of Colombia. J. Biogeogr. 2004, 31, 1829–1839. [Google Scholar] [CrossRef]
- Seipel, T.; Kueffer, C.; Rew, L.J.; Daehler, C.C.; Pauchard, A.; Naylor, B.J.; Alexander, J.M.; Edwards, P.J.; Parks, C.G.; Arevalo, J.R.; et al. Processes at multiple scales affect richness and similarity of non-native plant species in mountains around the world. Glob. Ecol. Biogeogr. 2012, 21, 236–246. [Google Scholar] [CrossRef]
- Vazquez, G.; Givnish, T.J. Altitudinal gradients in tropical forest composition, structure, and diversity in the Sierra de Manantalan. J. Ecol. 1998, 86, 999–1020. [Google Scholar]
- Willig, M.R.; Presley, S.J. The spatial configuration of taxonomic biodiversity along a tropical elevational gradient: α-, β-, and γ-partitions. Biotropica 2019, 51, 104–116. [Google Scholar] [CrossRef]
- Britton, A.J.; Beale, C.M.; Towers, W.; Hewison, R.L. Biodiversity gains and losses: Evidence for homogenization of Scottish alpine vegetation. Biol. Conserv. 2009, 142, 1728–1739. [Google Scholar] [CrossRef]
- Hoorn, C.; Wesselingh, F.P.; Ter Steege, H.; Bermudez, M.A.; Mora, A.; Sevink, J.; Sanmartín, I.; Sanchez-Meseguer, A.; Anderson, C.L.; Antonelli, A. Amazonia through time: Andean uplift, climate change, landscape evolution, and biodiversity. Science 2010, 330, 927–931. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Wang, Q.-R.; Fan, W.; Song, G.-H. The relationship between secondary forest and environmental factors in the southern Taihang Mountains. Sci. Rep. 2017, 7, 16431. [Google Scholar] [CrossRef] [Green Version]
- Walter, H. Vegetation of the Earth in Relation to Climate and the Eco-Physiological Conditions; English University Press: London, UK, 1973. [Google Scholar]
- Brown, J.H.; Whitham, T.G.; Ernest, S.K.M.; Gehring, C.A. Complex species interactions and the dynamics of ecological systems: Long-term experiments. Science 2001, 293, 643–650. [Google Scholar] [CrossRef] [Green Version]
- Pauli, H.; Gottfried, M.; Dullinger, S.; Abdaladze, O.; Akhalkatsi, M.; Alonso, J.L.B.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, R.F.; et al. Recent plant diversity changes on Europe’s mountain summits. Science 2012, 336, 353–356. [Google Scholar] [CrossRef] [Green Version]
- Gaston, K.J. Global patterns in biodiversity. Nature 2000, 405, 220–227. [Google Scholar] [CrossRef]
- Fine, P.V.A. Ecological and evolutionary drivers of geographic variation in species diversity. Annu. Rev. Ecol. Evol. Syst. 2015, 46, 369–392. [Google Scholar] [CrossRef] [Green Version]
- Peters, M.; Hemp, A.; Appelhans, T.; Behler, C.; Classen, A.; Detsch, F.; Ensslin, A.; Ferger, S.W.; Frederiksen, S.B.; Gebert, F.; et al. Predictors of elevational biodiversity gradients change from single taxa to the multi-taxa community level. Nat. Commun. 2016, 7, 13736. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, P.; Tasser, E.; Leitinger, G.; Tappeiner, U. Effects of land-use and land-cover pattern on landscape-scale biodiversity in the European Alps. Agriculture. Ecosyst. Environ. 2010, 139, 13–22. [Google Scholar] [CrossRef]
- Linder, H.P.; De Klerk, H.M.; Born, J.; Burgess, N.D.; Fjeldså, J.; Rahbek, C. The partitioning of Africa: Statistically denied biogeographical regions in sub-Saharan Africa. J. Biogeogr. 2012, 39, 1189–1205. [Google Scholar] [CrossRef]
- Sanders, N.J.; Rahbek, C. The patterns and causes of elevational diversity gradients. Ecography 2012, 35, 1–3. [Google Scholar] [CrossRef]
- Guo, Q.; Kelt, D.A.; Sun, Z.; Liu, H.; Hu, L.-J.; Ren, H.; Wen, J. Global variation in elevational diversity patterns. Sci. Rep. 2013, 3, srep03007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whittaker, R.H. Vegetation of the Siskiyou Mountains, Oregon and California. Ecol. Monogr. 1960, 30, 279–338. [Google Scholar] [CrossRef]
- De Frenne, P.; Rodríguez-Sánchez, F.; Coomes, D.A.; Baeten, L.; Verstraeten, G.; Vellend, M.; Bernhardt-Römermann, M.; Brown, C.D.; Brunet, J.; Cornelis, J.; et al. Microclimate moderates plant responses to macroclimate warming. Proc. Natl. Acad. Sci. USA 2013, 110, 18561–18565. [Google Scholar] [CrossRef] [Green Version]
- Chun, J.H.; Lee, C.B. Partitioning the regional and local drivers of phylogenetic and functional diversity along temperate elevational gradients on an East Asian peninsula. Sci. Rep. 2018, 8, 2853. [Google Scholar] [CrossRef] [Green Version]
- Horák, J.; Materna, J.; Halda, J.P.; Mladenović, S.; Bogusch, P.; Pech, P. Biodiversity in remnants of natural mountain forests under conservation-oriented management. Sci. Rep. 2019, 9, 89. [Google Scholar] [CrossRef] [Green Version]
- Naud, L.; Måsviken, J.; Freire, S.; Angerbjörn, A.; Dalén, L.; Dalerum, F. Altitude effects on spatial components of vascular plant diversity in a subarctic mountain tundra. Ecol. Evol. 2019, 9, 4783–4795. [Google Scholar] [CrossRef] [Green Version]
- Lomolino, M.V. Elevation gradients of species-density: Historical and prospective views. Glob. Ecol. Biogeogr. 2001, 10, 3–13. [Google Scholar] [CrossRef]
- Hansen, A.J.; Spies, T.A.; Swanson, F.J.; Ohmann, J.L. Conserving biodiversity in Managed forests. BioScience 1991, 21, 382–392. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Wang, J.; Yu, F.; Zhang, X.; Yao, Y.; Zhang, B. Altitudinal biodiversity gradient and ecological drives for different lifeforms in the Baotianman Nature Reserve of the eastern Qinling Mountains. Forests 2019, 10, 332. [Google Scholar] [CrossRef]
- Cabrera, O.; Benítez, A.; Cumbicus, N.; Naranjo, C.; Ramón, P.; Tinitana, F.; Escudero, A. Geomorphology and altitude effects on the diversity and structure of the vanishing montane forest of southern Ecuador. Diversity 2019, 11, 32. [Google Scholar] [CrossRef] [Green Version]
- Dhakal, B.; Kattel, R.R. Effects of global changes on ecosystems services of multiple natural resources in mountain agricultural landscapes. Sci. Total Environ. 2019, 676, 665–682. [Google Scholar] [CrossRef]
- Xu, M.; Zhang, S.; Wen, J.; Yang, X. Multiscale spatial patterns of species and biomass together with their correlations along geographical gradients in subalpine meadows. PLoS ONE 2019, 14, e0211560. [Google Scholar] [CrossRef] [Green Version]
- Fan, X. The Road to the Taihang Mountains; China Forestry Publishing House: Beijing, China, 2015. [Google Scholar]
- Liu, X.; Ge, J.; Feng, X. Study on ecological security of land resources in Taihang Mountain Hebei. J. Arid Land Resour. Environ. 2007, 21, 68–74. [Google Scholar]
- Gao, H.; Fu, T.G.; Liu, J.T.; Liang, H.; Han, L. Ecosystem services management based on differentiation and regionalization along altitudinal gradient in Taihang Mountain, China. Sustainability 2018, 10, 986. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, E.M.; Arroyo-Rodríguez, V.; Santos, B.A.; Tabarelli, M.; Leal, I.R. Chronic anthropogenic disturbance drives the biological impoverishment of the Brazilian Caatinga vegetation. J. Appl. Ecol. 2015, 52, 611–620. [Google Scholar] [CrossRef]
- Wolf, H.D.J. Diversity patterns and biomass of epiphytic bryophytes and lichens along an altitudinal gradient in the northern Andes. Ann. Mo. Bot. Gard. 1993, 4, 928–960. [Google Scholar]
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and qualitative β diversity measures lead to different insights into factors that structure microbial communities. Appl. Environ. Microbiol. 2007, 73, 1576–1585. [Google Scholar] [CrossRef] [Green Version]
- Jankowski, J.E.; Ciecka, A.L.; Meyer, N.Y.; Rabenold, K.N. Beta diversity along environmental gradients: Implications of habitat specialization in tropical montane landscapes. J. Anim. Ecol. 2009, 78, 315–327. [Google Scholar] [CrossRef]
- Koellner, T.; Hersperger, A.M.; Wohlgemuth, T. Rarefaction method for assessing plant species diversity on a regional scale. Ecography 2004, 27, 532–544. [Google Scholar] [CrossRef]
- Guisan, A.; Weiss, S.B.; Weiss, A. GLM versus CCA spatial modeling of plant species distribution. Plant Ecol. 1999, 143, 107–122. [Google Scholar] [CrossRef]
- Kessler, M. Patterns of diversity and range size of selected plant groups along an elevational transect in the Bolivian Andes. Biodivers. Conserv. 2001, 10, 1897–1921. [Google Scholar] [CrossRef]
- Wang, G.; Zhou, G.; Yang, L.; Li, Z. Distribution, species diversity and life-form spectra of plant communities along an altitudinal gradient in the northern slopes of Qilianshan Mountains, Gansu, China. Plant Ecol. 2002, 165, 169–181. [Google Scholar] [CrossRef]
- Sanchez-Gonzalez, A.; Lopez-Mata, L. Plant species richness and diversity along an altitudinal gradient in the Sierra Nevada, Mexico. Divers. Distrib. 2005, 11, 567–575. [Google Scholar] [CrossRef]
- Cardelus, C.L.; Colwell, R.K.; Watkins, J.E. Vascular epiphyte distribution patterns: Explaining the mid-elevation richness peak. J. Ecol. 2006, 94, 144–156. [Google Scholar] [CrossRef]
- Buckley, L.B.; Jetz, W. Linking global turnover of species and environments. Proc. Natl. Acad. Sci. USA 2008, 105, 17836–17841. [Google Scholar] [CrossRef] [Green Version]
- Stohlgren, T.J.; Binkley, D.; Chong, G.W.; Kalkhan, M.A.; Schell, L.D.; Bull, K.A.; Otsuki, Y.; Newman, G.; Bashkin, M.; Son, Y. Exotic plant species invade hot spots of native plant diversity. Ecol. Monogr. 1999, 69, 25–46. [Google Scholar] [CrossRef]
- Graham, C.H.; Fine, P.V.A. Phylogenetic beta diversity: Linking ecological and evolutionary processes across space in time. Ecol. Lett. 2008, 11, 1265–1277. [Google Scholar] [CrossRef]
- Callaway, R.M.; Brooker, R.W.; Choler, P.; Kikvidze, Z.; Lortie, C.J.; Michalet, R.; Paolini, L.; Pugnaire, F.I.; Newingham, B.; Aschehoug, E.T.; et al. Positive interactions among alpine plants increase with stress. Nature 2002, 417, 844–848. [Google Scholar] [CrossRef]
- Ohmann, J.L.; Spies, T.A. Regional gradient analysis and spatial pattern of woody plant communities of Oregon forest. Ecol. Monogr. 1998, 68, 151–182. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Min | Max | Mean | Standard Deviation | Skewness | Kurtosis | |
|---|---|---|---|---|---|---|---|
| Climatic factors | Temperature (°C) | 7.40 | 11.56 | 8.91 | 1.26 | 0.43 | −0.47 |
| Precipitation (mm) | 491 | 547 | 529 | 15.68 | −1.01 | 0.86 | |
| Non-climatic factors | Slope (°) | 3.13 | 43.84 | 17.65 | 12.51 | 0.88 | 0.46 |
| Hfp | 21 | 43 | 28.56 | 7.31 | 0.50 | −0.80 | |
| Hii | 14 | 28 | 19.13 | 4.26 | 0.57 | −0.22 | |
| NPP (gC·m−2·a−1) | 139.93 | 412.47 | 339.01 | 62.21 | −2.27 | 7.15 | |
| Population density (p/km2) | 0 | 258 | 43.88 | 93.22 | 1.88 | 1.97 | |
| pH | 5.31 | 6.91 | 6.07 | 0.41 | −0.20 | 0.52 | |
| VWC (100%) | 0.02 | 0.61 | 0.20 | 0.03 | 1.40 | 1.61 | |
| Factors | CCA1 | CCA2 | r2 | Pr (>r) |
|---|---|---|---|---|
| Hfp | −0.66 | 0.75 | 0.69 | 0.002 ** |
| Temperature | −0.88 | 0.48 | 0.66 | 0.001 *** |
| Hii | −0.71 | 0.70 | 0.62 | 0.002 ** |
| Population density | −0.26 | 0.96 | 0.55 | 0.005 ** |
| Precipitation | 0.82 | -0.58 | 0.52 | 0.004 ** |
| VWC (%) | 0.99 | -0.07 | 0.40 | 0.033 * |
| NPP | −0.64 | -0.77 | 0.28 | 0.09 |
| pH | 0.99 | -0.17 | 0.02 | 0.85 |
| Slope | -0.74 | -0.67 | 0.01 | 0.90 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, H.; Fu, T.; Gao, H.; Li, M.; Liu, J. Climatic and Non-Climatic Drivers of Plant Diversity along an Altitudinal Gradient in the Taihang Mountains of Northern China. Diversity 2023, 15, 66. https://doi.org/10.3390/d15010066
Liang H, Fu T, Gao H, Li M, Liu J. Climatic and Non-Climatic Drivers of Plant Diversity along an Altitudinal Gradient in the Taihang Mountains of Northern China. Diversity. 2023; 15(1):66. https://doi.org/10.3390/d15010066
Chicago/Turabian StyleLiang, Hongzhu, Tonggang Fu, Hui Gao, Min Li, and Jintong Liu. 2023. "Climatic and Non-Climatic Drivers of Plant Diversity along an Altitudinal Gradient in the Taihang Mountains of Northern China" Diversity 15, no. 1: 66. https://doi.org/10.3390/d15010066
APA StyleLiang, H., Fu, T., Gao, H., Li, M., & Liu, J. (2023). Climatic and Non-Climatic Drivers of Plant Diversity along an Altitudinal Gradient in the Taihang Mountains of Northern China. Diversity, 15(1), 66. https://doi.org/10.3390/d15010066