


Wintering and Cold Hardiness of the Small Tortoiseshell Aglais urticae (Linnaeus, 1758) (Nymphalidae, Lepidoptera) in the West and East of the Northern Palearctic

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Molecular Genetic Analysis
2.2. Animal Sampling and Care
2.3. Determination of Cold Hardiness
2.3.1. Supercooling Point
2.3.2. Lower Lethal Temperatures
2.3.3. Temperatures in Unheated Buildings and in the Air
2.4. Statistical Analysis
3. Results
3.1. Distribution
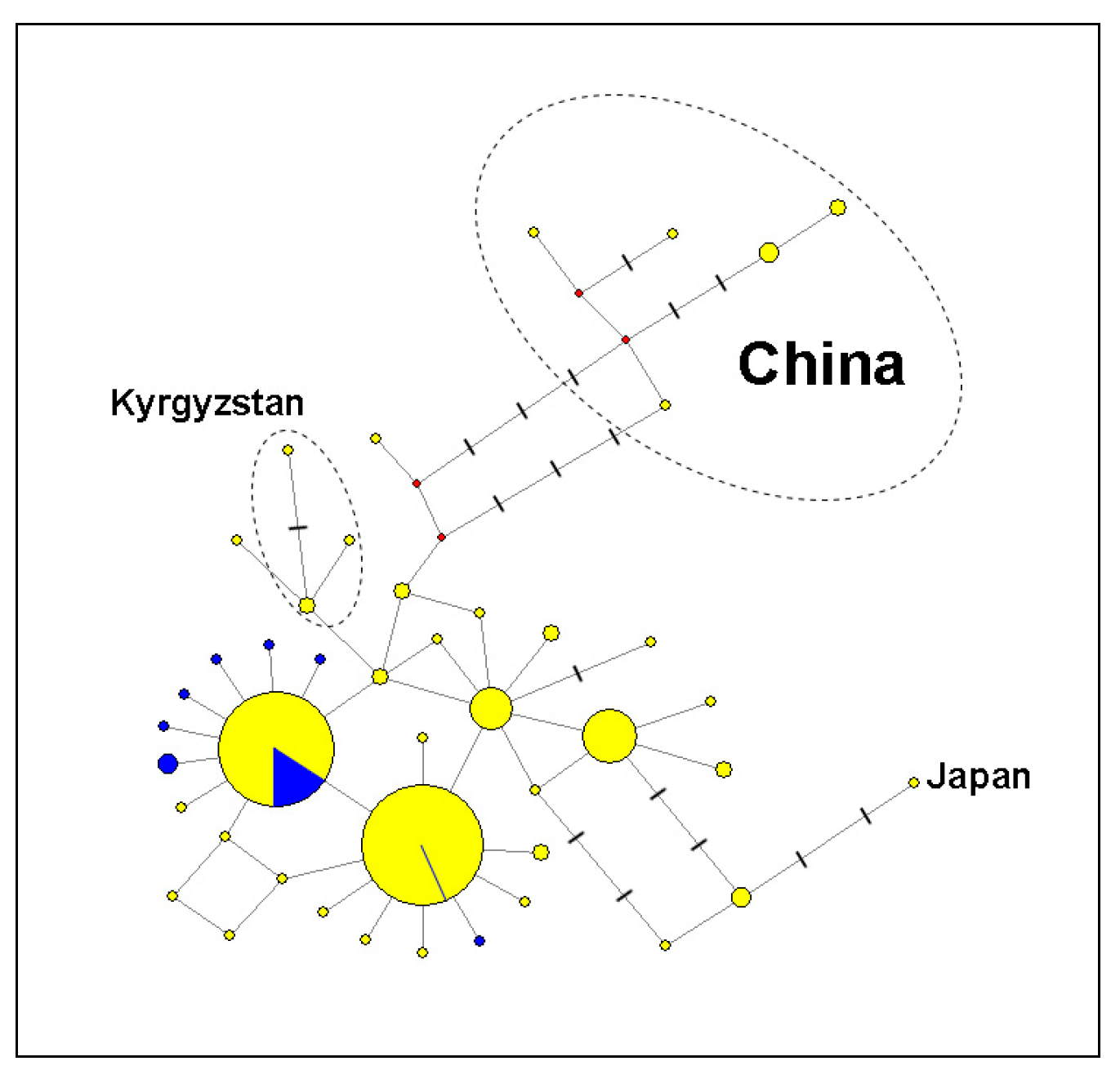
3.2. Molecular Genetic Analysis
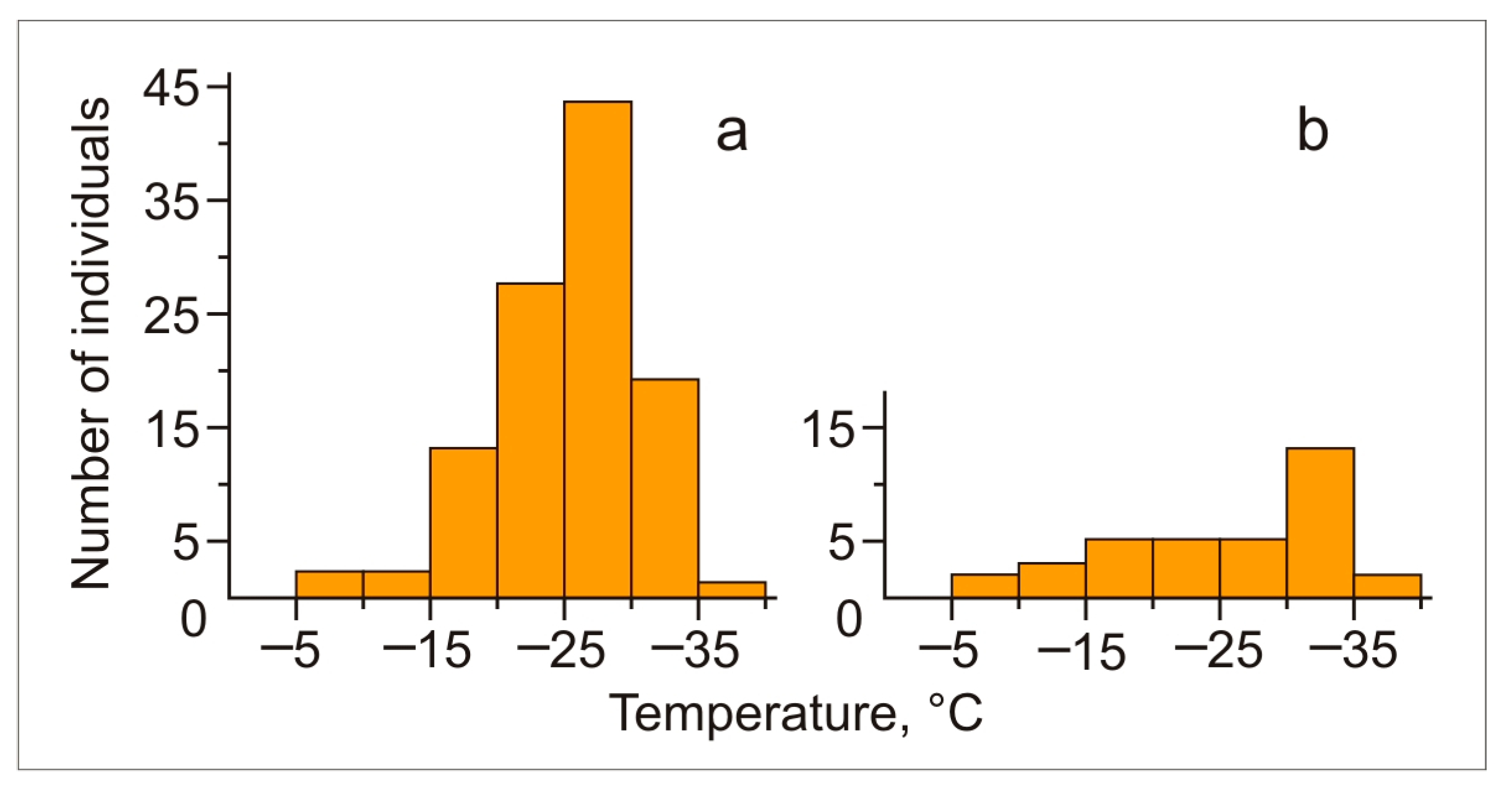
3.3. Cold Hardiness
3.4. The Natural Temperature Conditions of Wintering

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Location | January Average Air Temperature, °C | Averages of the Absolute Minimum |
|---|---|---|---|
| 1 | Magadan [38] | −18.2 | −32 |
| 2 | Talon [38] | −29 | −49 |
| 3 | Palatka [38] | −23.4 | −40 |
| 4 | Madaun [38] | −30.6 | −52 |
| 5 | Ust-Omchug [38] | −35.2 | −53 |
| 6 | Kolymskaya [38] | −35.0 | −51 |
| 7 | Yagodnoye [38] | −35.1 | −50 |
| 8 | Seymchan [38] | −39.1 | −57 |
| 9 | Susuman [38] | −39.8 | −57 |
| 10 | Delyankir [15] | −47.6 | −61 |
| 11 | Oymyakon [15] | −50.0 | −64 |
| 12 | Moma [15] | −46.8 | −60 |
| 13 | Yakutsk [15] | −43.2 | −57 |
3.5. Temperatures in Unheated Buildings
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ring, R.A.; Tesar, D. Adaptations to cold in Canadian arctic insects. Cryobiology 1981, 18, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Bird, J.M.; Hodkinson, I.D. Species at the edge of their range: The significance of the thermal environment for the distribution of congeneric Craspedolepta species (Sternorrhncha: Psylloidea) living on Chamerion angustifolium (Onagraceae). Eur. J. Entomol. 1999, 96, 103–109. [Google Scholar]
- Kalushkov, P.; Nedved, O. Cold hardiness of Pyrrhocoris apterus (Heteroptera: Pyrrhocoridae) from central and southern Europe. Eur. J. Entomol. 2000, 97, 149–153. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, L.M.; Holmstrup, M. Geographic variation of freeze-tolerance in the earthworm Dendrobaena octaedra. J. Comp. Physiol. B 2002, 172, 91–698. [Google Scholar]
- Bale, J.S.; Hayward, S.A.L. Insect overwintering in a changing climate. J. Exp. Biol. 2010, 213, 980–994. [Google Scholar] [CrossRef] [Green Version]
- Meshcheryakova, E.N.; Bulakhova, N.A. Cold resistance of three species of earthworms (Oligohaeta, Lumbricidae) in Western Siberia. Intern. J. Environ. Stu. 2014, 71, 749–754. [Google Scholar] [CrossRef]
- Meshcheryakova, E.N.; Berman, D.I. Cold hardiness and geographic distribution of earthworms (Oligochaeta, Lumbricidae, Moniligastridae). Entomol. Rev. 2014, 94, 486–497. [Google Scholar] [CrossRef]
- Battisti, A.; Larsson, S. Climate change and insect pest distribution range. In Climate Change and Insect Pests; Björkman, C., Niemelä, P., Eds.; CABI International: Wallingford, UK, 2015; pp. 1–16. [Google Scholar]
- Ananko, G.G.; Kolosov, A.V. Asian gypsy moth (Lymantria dispar L.) populations: Tolerance of eggs to extreme winter temperatures. J. Therm. Biol. 2021, 102, 103–123. [Google Scholar] [CrossRef]
- Daneck, H.; Barth, M.B.; Geck, M.; Hundsdoerfer, A.K. Super cooling point phenotypes and old resistance in Hyles euphorbiae hawk moths from different climate zones. Diversity 2021, 13, 207. [Google Scholar] [CrossRef]
- Gorbunov, P.; Kosterin, O. The Butterflies of North Asia in Nature; Aidis Producers House, Rodina & Fodio: Moscow, Russia, 2007; Volume 2, pp. 1–412. [Google Scholar]
- Bryant, S.R.; Thomas, C.D.; Bale, J.S. Nettle-feeding nymphalid butterflies: Temperature, development and distribution. Ecol. Entomol. 1997, 22, 390–398. [Google Scholar] [CrossRef]
- Lebedev, A.N. (Ed.) Western European Climate Guide; Gidrometeoizdat: Leningrad, Russia, 1979; pp. 1–678. (In Russian) [Google Scholar]
- Antonova, E.M.; Berman, D.I. Diurnal butterflies of the relict steppes of northeastern Yakutia. In Insects of the Meadow-Taiga Biocenoses of Yakutia; Publishing house of Academy of Sciences USSR: Yakutsk, Russia, 1988; pp. 57–59. (In Russian) [Google Scholar]
- Izyumenko, S.A. (Ed.) Climate of the USSR: An Applied Reference Book; Gidrometeoizdat: Leningrad, Russia, 1966; Volume 24, pp. 1–398. (In Russian) [Google Scholar]
- Pullin, A.S.; Bale, J.S. Effects of low temperature on diapausing Aglais urticae and Inachis io (Lepidoptera: Nymphalidae): Cold hardiness and overwintering survival. J. Insect Physiol. 1989, 35, 277–281. [Google Scholar] [CrossRef]
- Kaimuk, E.L.; Vinokurov, N.N.; Burnasheva, A.P. Insects of Yakutia. Butterflies; IBPC SB RAS: Yakutsk, Russia, 2005; pp. 1–88. (In Russian) [Google Scholar]
- Vlasova, A.A.; Potapov, G.S. Fauna of the butterflies (Lepidoptera: Rhopalocera) the lower reaches of the Ob river Yamal-Nenets autonomous District. In Arctic Research from Extensive Development to Integrated Development; Northern (Arctic) Federal University: Arkhangelsk, Russia, 2018; pp. 242–243. (In Russian) [Google Scholar]
- Strouhal, H.; Vornatscher, J. Katalog der rezenten Höhlentiere Österreichs. Ann. Des Nat. Mus. Wien 1975, 79, 401–542. [Google Scholar]
- Dvorak, L.; Belicek, J.; Fric, Z. Observations of overwintering nymphalid butterflies in underground shelters in SW and W Bohemia (Czech Republic) (Lepidoptera: Nymphalidae: Nymphalini). J. Res. Lepidop. 2009, 41, 45–52. [Google Scholar] [CrossRef]
- Pan’kov, O.S.; Starova, O.S. Invertebrate animals of the Perm region cave. In Peshchery Caves: Collection of Scientific Transactions; Natural Science Institute of Perm State Research University: Perm, Russia, 2013; Volume 36, pp. 103–111. (In Russian) [Google Scholar]
- Lewington, R. Pocket Guide to the Butterflies of Great Britain and Ireland; Bloomsbury Publishing: Oxford, UK, 2016; pp. 1–144. [Google Scholar]
- Poulton, E. Aglais urticae L. with symmetrical injuries possibly inflicted before hibernation, flying in house, 30 march, 1936, St. Helens, isle of wight. In Proceedings of the Royal Entomological Society of London. Series A, General Entomology; Blackwell Publishing Ltd.: Oxford, UK, 1936; Volume 11, p. 98. [Google Scholar]
- Wiklund, C.; Vallin, A.; Friberg, M.; Jakobsson, S. Rodent predation on hibernating peacock and small tortoiseshell butterflies. Behav. Ecol. and Sociobiol. 2008, 62, 379–389. [Google Scholar] [CrossRef]
- Lozina-Lozinskii, L.K. Studies in Cryobiology; Nauka: Leningrad, Russia, 1972; pp. 1–288. (In Russian) [Google Scholar]
- Shekhovtsov, S.V.; Golovanova, E.V.; Peltek, S.E. Cryptic diversity within the Nordenskiold’s earthworm, Eisenia nordenskioldi subsp. nordenskioldi (Lumbricidae, Annelida). Eur. J. Soil Biol. 2013, 58, 13–18. [Google Scholar] [CrossRef]
- Folmer, O.; Hoeh, W.R.; Black, M.B.; Vrijenhoek, R.C. Conserved primers for PCR amplification of mitochondrial DNA from different invertebrate phyla. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Chromas 2.6.6. Available online: http://technelysium.com.au (accessed on 20 September 2022).
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Berman, D.I.; Alfimov, A.V.; Zhigulskaya, Z.A.; Leirikh, A.N. Overwintering and Cold Hardiness of Ants in the Northeast of Asia; Pensoft Publishers: Sofia-Moscow, Russia, 2010; pp. 1–294. [Google Scholar]
- Berman, D.I.; Bulakhova, N.A.; Alfimov, A.V.; Meshcheryakova, E.N. How the most northern lizard, Zootoca vivipara, overwinters in Siberia. Polar Biol. 2016, 39, 2411–2425. [Google Scholar] [CrossRef]
- Sinclair, B.J.; Alvarado, L.E.C.; Ferguson, L.V. An invitation to measure insect cold tolerance: Methods, approaches, and workflow. J. Therm. Biol. 2015, 53, 180–197. [Google Scholar] [CrossRef] [Green Version]
- Berman, D.I.; Meshcheryakova, E.N.; Alfimov, A.V.; Leirikh, A.N. The distribution of the earthworm Dendrobaena octaedra (Lumbricidae: Oligochaeta) in the north of the Holarctic is limited by insufficient frost resistance. Zool. Zhurnal 2002, 81, 1210–1221. (In Russian) [Google Scholar]
- Berman, D.I.; Meshcheryakova, E.N.; Mikhaljova, E.V. Cold hardiness and range of the myriapod Angarozonium amurense (Polyzoniidae, Diplopoda, Arthropoda) in permafrost environments. CryoLetters 2015, 36, 237–242. [Google Scholar]
- Khokhryakov, A.P. Flora of the Magadan Region; Nauka: Moscow, Russia, 1985; p. 397. (In Russian) [Google Scholar]
- Terziev, F.S. (Ed.) Climate of the USSR: An Applied Reference Book; Gidrometeoizdat: Leningrad, Russia, 1965; Volume 3, pp. 1–144. (In Russian) [Google Scholar]
- Pakhnev, S.Y. (Ed.) Climate of the USSR: An Applied Reference Book; Gidrometeoizdat: Leningrad, Russia, 1965; Volume 20, pp. 1–396. (In Russian) [Google Scholar]
- Melnikova, T.V. (Ed.) Climate of the USSR: An Applied Reference Book; Gidrometeoizdat: Leningrad, Russia, 1966; Volume 33, pp. 1–288. (In Russian) [Google Scholar]
- Shtandfuss, M. Life of Butterflies, Their Catching, Education and Preservation; Education: St. Petersburg, Russia, 1901; pp. 1–315. (In Russian) [Google Scholar]
- Vandewoestijne, S.; Baguette, M.; Btakefield, P.M.; Saccheri, L.J. Phylogeography of Aglais urticae (Lepidoptera) based on DNA sequences of the mitochondrial COI gene and control region. Mol. Phylogenetics Evol. 2004, 31, 630–646. [Google Scholar] [CrossRef] [PubMed]
- Alfimov, A.B. Thermal regime of the upper layers of soil in the main ecosystems of the open forest belt of the Upper Kolyma basin. In Belt of Open Forests of the Upper Kolyma (the Area of Construction of the Kolyma Hydroelectric Power Station); DVNTs AN USSR: Vladivostok, Russia, 1985; pp. 9–29. (In Russian) [Google Scholar]
- Dennis, R.L.H. Retreats for peacock butterflies Inachis io L. (Lep.: Nymphalidae) in changing weather conditions. Ent. Rec. 2005, 117, 175. [Google Scholar]
- Goltzberg, I.A. (Ed.) Agro-Climatic Atlas of the World; Gidrometeoizdat: Moscow-Leningrad, Russia, 1972; pp. 1–146. [Google Scholar]
- Alfimov, A.V.; Berman, D.I. Two poles of cold under the Eurasian snow cover. Priroda 2006, 6, 27–33. (In Russian) [Google Scholar]
- Berman, D.I.; Leirikh, A.N. Overwintering and Cold Hardiness of the Invertebrates in the Northeast Asia; Saulich, F.H., Ed.; KMK Scientific Press: Moscow, Russia, 2019; pp. 1–314. (In Russian) [Google Scholar]







| Location | Coordinates | n |
|---|---|---|
| Chester, United Kingdom | 53°12′ N 2°53′ W | 1 |
| Oriketo District, Turku, Finland | 60°28′ N 22°19′ E | 5 |
| Saint Petersburg, Russia | 59°59′ N 30°23′ E | 4 |
| Novgorod, Russia | 58°31′ N 31°16′ E | 1 |
| Istra District, Moscow Oblast, Russia | 55°55′ N 36°52′ E | 1 |
| Pinega Village, Arkhangelsk Oblast, Russia | 64°42′ N 43°22′ E | 3 |
| Podgornoye Village, Tomsk Oblast, Russia | 57°47′ N 82°39′ E | 1 |
| Novosibirsk, Russia | 55°01′ N 82°55′ E | 3 |
| Yakutsk, Sakha Republic, Russia | 62°3′ N 129°29′ E | 3 |
| Yagodnoye Village, Magadan Oblast, Russia, | 62°31′ N 149°37′ E | 1 |
| Magadan, Russia | 59°34′ N 150°48′ E | 3 |
| Seymchan Village, Magadan Oblast, Russia | 62°52′ N 152°23′ E | 3 |
| Total | 29 |
| Temperature, °C | Duration, Days |
|---|---|
| 16 | 1 |
| 14 | 1 |
| 11 | 3 |
| 10 | 2 |
| 8 | 3 |
| 5 | 21 |
| 0 | 21 |
| Population | Magadan | Seymchan | Podgornoye | |||
|---|---|---|---|---|---|---|
| Year | 2018 * | 2018 ** | 2020 * | 2022 * | 2022 * | 2022 * |
| n | 32 | 19 | 29 | 16 | 35 | 5 |
| SCP mean ±SE | −24.0 ± 0.8 | −25.0 ± 1.0 | −28.1 ± 1.0 | −23.1 ± 1.8 | −25.3 ± 1.4 | −19.4 ± 3.1 |
| SCP min | −34.8 | −29.3 | −35.2 | −32.8 | −35.5 | −26.1 |
| Location | Altitude, m | Air Temperature, °C | |
|---|---|---|---|
| Average, January | Averages of the Absolute Minimum | ||
| London [13] | 22 | 3.8 | 3.3 |
| Saint Petersburg [36] | 3 | −7.8 | −26 |
| Podgornoye [37] | 97 | −20.6 | −44 |
| Magadan [38] | 115 | −18.2 | −32 |
| Seymchan [38] | 207 | −39.1 | −57 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meshcheryakova, E.N.; Bulakhova, N.A.; Zhigulskaya, Z.A.; Shekhovtsov, S.V.; Berman, D.I. Wintering and Cold Hardiness of the Small Tortoiseshell Aglais urticae (Linnaeus, 1758) (Nymphalidae, Lepidoptera) in the West and East of the Northern Palearctic. Diversity 2023, 15, 72. https://doi.org/10.3390/d15010072
Meshcheryakova EN, Bulakhova NA, Zhigulskaya ZA, Shekhovtsov SV, Berman DI. Wintering and Cold Hardiness of the Small Tortoiseshell Aglais urticae (Linnaeus, 1758) (Nymphalidae, Lepidoptera) in the West and East of the Northern Palearctic. Diversity. 2023; 15(1):72. https://doi.org/10.3390/d15010072
Chicago/Turabian StyleMeshcheryakova, Ekaterina N., Nina A. Bulakhova, Zoya A. Zhigulskaya, Sergei V. Shekhovtsov, and Daniil I. Berman. 2023. "Wintering and Cold Hardiness of the Small Tortoiseshell Aglais urticae (Linnaeus, 1758) (Nymphalidae, Lepidoptera) in the West and East of the Northern Palearctic" Diversity 15, no. 1: 72. https://doi.org/10.3390/d15010072
APA StyleMeshcheryakova, E. N., Bulakhova, N. A., Zhigulskaya, Z. A., Shekhovtsov, S. V., & Berman, D. I. (2023). Wintering and Cold Hardiness of the Small Tortoiseshell Aglais urticae (Linnaeus, 1758) (Nymphalidae, Lepidoptera) in the West and East of the Northern Palearctic. Diversity, 15(1), 72. https://doi.org/10.3390/d15010072